
Contributions on Lindane Degradation by Microcystis aeruginosa PCC 7806
by
 ,
,
Cristina Sarasa-Buisán
, 
Jorge Guío
 ,
,
Carolina Castro
,
María Teresa Bes
,
María F. Fillat
,
María Luisa Peleato
and
Emma Sevilla
* Department of Biochemistry, Molecular Biology, Institute of Biocomputation, Physics of Complex Systems, Universidad de Zaragoza, 50009 Zaragoza, Spain
*
Author to whom correspondence should be addressed.
Water 2022, 14(8), 1219; https://doi.org/10.3390/w14081219
Submission received: 23 March 2022
/
Revised: 6 April 2022
/
Accepted: 8 April 2022
/
Published: 11 April 2022
(This article belongs to the Special Issue Emerging Pollutants in Aquatic Environments and Chemical-Biological Remediation)
Abstract
:Cyanobacteria are able to tolerate, and even metabolize, moderate doses of organochlorine pesticides, such as lindane (γ-hexachlorocyclohexane), one of the most persistent and widely used in recent decades. Previous work showed that Microcystis aeruginosa PCC 7806 degrades lindane and that, in the presence of the pesticide, microcystin synthesis is enhanced. In this work, using in silico approaches, we have identified in M. aeruginosa putative homologues of the lin genes, involved in lindane degradation in Sphingobium japonicum UT26S. Real-time RT-PCR assays showed that the putative linC gene was induced in the presence of 7 mg/L of lindane. Additionally, prxA, encoding a peroxiredoxin, and involved in oxidative stress response, was also induced when lindane was present. Taking into account these results, M. aeruginosa PCC 7806 may degrade lindane through a metabolic pathway involving a putative 2,5-dichloro-2,5-cyclohexadiene-1,4-diol dehydrogenase encoded by a linC homologue. However, the low similarity of the other potential lin homologues suggest the existence of an alternative pathway different to that of heterotrophic microorganisms such as S. japonicum.
1. Introduction
Cyanobacteria are organisms with an outstanding capacity to adapt and survive in extreme or highly degraded environments. Their metabolic plasticity includes the synthesis of a high level of potentially toxic secondary metabolites, such as cyanotoxins, that cause serious health and environmental problems. Cyanobacteria are able to tolerate and even metabolize moderate doses of organochlorine pesticides, such as lindane (γ-hexachlorocyclohexane) [1,2,3,4,5], one of the most persistent and widely used in recent decades. Previous work showed that Microcystis aeruginosa PCC 7806 degrades lindane, and that in the presence of lindane, microcystin synthesis is enhanced [6] and oxidative stress responses are induced [6]. However, there is not yet information concerning the degradative pathway in cyanobacteria.
The aerobic degradation pathway of lindane was described in Sphingobium japonicum UT26S, which is able to use lindane as a unique carbon and energy source [7]. The main reactions that occur during the aerobic degradation of lindane include dehydrogenation, dechlorination, hydroxylation, dehydrochlorination, and mineralization [8], and the enzymes involved in the pathway are encoded by the lin genes [7].
Lindane is a persistent pesticide that has been widely used in the past. Lindane is toxic and bioaccumulative and, even though its production has been eliminated in most of countries, spillages from former factories and illegal dumping of a high amount of subproducts (other hexachlorocyclohexane isomers) have given risen to serious environmental concerns. These highly polluted sites spill HCH into the surface of soils, and to the surface and groundwater. Cyanobacteria are a good candidate to bioremediate HCH from surface waters, due to its photoautotrophic growth, plasticity, and ability to survive in a wide range of environmental conditions. Previous work suggested the presence of potential metabolic machinery to degrade HCH [1,2,3,4,5,6], which makes cyanobacteria a good candidate for the bioremediation of lindane in polluted aquatic ecosystems.
In this work we have identified genes potentially involved in the aerobic degradation of lindane by M. aeruginosa, and we have studied the transcriptional response in the presence of lindane of a putative linC homologue, the gene showing the highest similarity with the lin genes of S. japonicum UT26S. Before now, nothing was known about the degradative pathway of this organochlorine in cyanobacteria, and this is the first work that tries to decipher the metabolic transformations that occur in those photosynthetic cells.
2. Materials and Methods
2.1. Growth Conditions and Cell Visualization
The axenic strain Microcystis aeruginosa PCC 7806 was provided by the Pasteur Culture Collection (Paris, France), and grown in BG11 media [9] with 2 mM of NaNO3, as indicated by the Pasteur Institute. Cells were grown in batch conditions with continuous agitation at 25 °C (150 rpm). The cyanobacteria were grown using a light intensity of 40 μmol of photons m−2 s−1, unless indicated. Light was measured using a quantum sensor photometer (Skye Instruments, SKP 200). Every culture, control, and lindane stressed, was started with equal aliquots of 0.3 OD (700 nm). Three different experiments were performed in duplicate, using 250 mL of medium with cells in 500 mL Erlenmeyer flasks to avoid substantial volume changes during the sampling. Three different absorbance measurements at 700 nm (OD) were taken for each sample. Cells after 4 h of treatment were measured in a Nikon Eclipse 50i epi-fluorescent, provided with a micrometer scale. Fifty cells of each culture (with and without lindane) were measured. Microcystis tends to aggregate, so 0.5 M of KOH was used to individualize each cell. It was tested that the treatment had no effects on size and appearance with a light microscope.
2.2. In Silico Studies of lin Genes
All the in silico analyses were performed in M. aeruginosa NIES-843, since the genome of the strain PCC 7806 is not available. In order to identify putative lin genes in M. aeruginosa, the sequences of the proteins codified by genes linA, linB, linC, linD, linE, and linR from S. japonicum UT26S were retrieved from UniProt [10]. Using these sequences as queries and an expectancy value threshold of 0.005, a protein BLAST (Basic Local Alignment Search Tool) using the CyanoBase similarity search [11] with default parameters was performed on the M. aeruginosa NIES-843 genome. In all cases, the sequence with the lowest expectation value was selected and a pairwise global alignment using EMBOSS Needle [12] with default settings was performed to obtain the percentages of identity and similarity. Following the criteria proposed by Rost [13], which establish that 30% of identity is an appropriate cut-off to determine whether two protein sequences are homologous, only the protein sequences of M. aeruginosa showing more than 30% of identity, with respect to the corresponding sequences of S. japonicum UT26S, were considered as lin genes homologues.
The LinC phylogenetic tree was generated via maximum likelihood analysis using Phylogeny.fr. [14,15] Sequences of proteins codified by putative linC genes in other cyanobacteria were obtained performing a protein BLAST restricted to cyanobacteria (taxid: 117), using the LinC protein sequence from S. japonicum UT26S as query and the NIH BLASTp suite [16], and then selecting in each case the sequence with the lowest expectation value. Multiple sequence alignment of these sequences was performed using MUSCLE [17], while sequences were curated using G-blocks, with options for a less stringent selection [18]. A maximum likelihood phylogenetic tree was built using PhyML with default settings [19]. To assure statistical significance, 100 bootstraps were used in the computation of the tree.
2.3. RNA Isolation and RealTime RT-PCR
Sampling was performed very carefully to avoid RNA degradation during manipulation. Aliquots of 25 mL of the cultures were harvested by centrifugation at 4000× g for 4 min at 4 °C. After removing the supernatant, each cell pellet was resuspended in 600 µL of 50 mM Tris-HCl (pH 8), 100 mM EDTA, and 130 µL of chloroform, and incubated in ice for 3 min to eliminate external RNases. The buffer was removed by centrifugation at 13000× g for 5 min at 4 °C. Finally, the cell pellets were frozen in liquid nitrogen and kept at –80 °C until RNA isolation was achieved. Cells were lysed using TRIZOL (Invitrogen) according to the manufacturer’s instructions. After the chloroform extraction, RNA was collected from the aqueous layer, and precipitated in isopropanol and liquid nitrogen. The RNA pellet was washed twice with 75% ethanol.
Prior to retrotranscription, the total RNA was treated with 40 units of DNase I (Pharmacia) in a volume of 100 µL, using a buffer containing 4 mL of 1 M Tris-HCl (pH 7.5) and 0.6 mL 1 M of MgCl2 in DEPC-H2O. The sample was incubated at 37 °C for 1 h. After digestion, the enzyme was inactivated by phenol acid and chloroform extraction, and RNA was precipitated with absolute ethanol. The successful digestion of DNA was assessed via PCR, with primers targeting the 16S rRNA gene (Table 1). RNA integrity was checked using a 1% agarose gel, and the concentration was determined by measuring the absorbance at 260 nm. Its purity was assessed by the ratio A260nm/A280nm. For reverse transcription, 1 µg of total RNA was mixed with 150 ng of random hexamer primers (Invitrogen) and diluted with the annealing buffer (10 mM Tris-HCl (pH 8), 1 mM EDTA, 150 mM KCl) to a final volume of 10 µL. The mixture was heated at 85 °C for 10 min and then incubated at 50 °C for 1 h. After this, the RNA was reverse-transcribed with 200 U of SuperScriptTM (GibcoBRL) in the presence of 2 µL of deoxyribonucleoside triphosphate mixture (2.5 mM each), 2 µL of dithiothreitol (100 mM), and 4 µL of the 5× buffer provided by the manufacturer with the reverse transcriptase enzyme kit. The volume was adjusted to 20 µL in DEPC-H2O. The mixture was incubated at 47 °C for 1 h and finally heated at 75 °C for 15 min.
Real-time PCR was performed using the ViiA™ 7 Real-Time PCR System (Applied Biosystems). Each reaction was set up by mixing 12.5 μL of SYBR Green PCR Master Mix with 0.4 μL of 25 μM primer mixture and 10 ng of cDNA template in a final volume of 30 μL. Amplification was performed at 60 °C. Negative controls with no cDNA were included. Transcript levels of target genes were normalized to those of the housekeeping 16S rRNA. The sequences of the specific primers are shown in Table S1. Relative quantification of gene expression was performed according to the comparative Ct method (the ΔΔCt method) [20]. The minimum fold change threshold was set to ±1.5 fold.
3. Results
3.1. In Silico Identification of lin Gene Homologues
In silico studies allowed the identification of genes homologous to the lin genes of S. japonicum UT26S in M. aeruginosa NIES-843 genome. The results of these in silico studies are summarized in Table 1.
As can be seen in Table 1, only the protein encoded by the gene MAE_33900 of M. aeruginosa NIES-843, which corresponds to the possible linC homologue, presented more than 30% of identity with respect to the corresponding sequence of S. japonicum UT26S, whereas the rest of the sequences presented identities lower than 30%. For this reason, following the criteria proposed by Rost [13], which established that 30% of identity is an appropriate cut-off to determine whether two protein sequences are homologous, the gene MAE_33900 was proposed as a putative linC gene of M. aeruginosa. Additionally, bidirectional BLAST showed that the protein encoded by the linC gene of S. japonicum UT26S, despite not being the best hit, also appears as a putative homologue of the protein encoded by the gene MAE_33900 of Microcystis aeruginosa NIES-843 with an expectancy value of 1·10−37.
3.2. M. aeruginosa PCC 7806 Growth in the Presence of Lindane
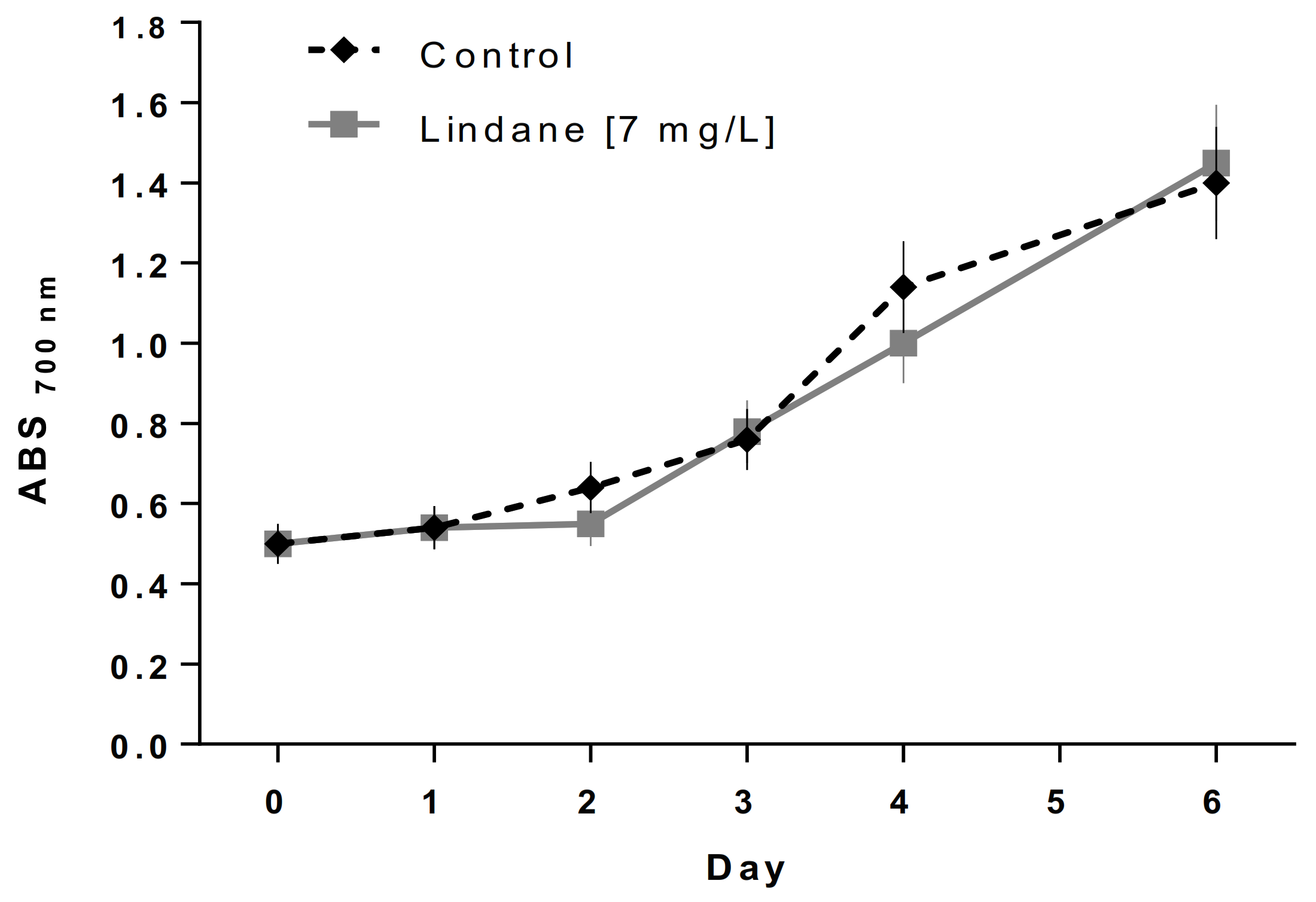
Lindane was used at its water solubility limit of 7 mg/L to avoid the undesired effects of the solvents. Visual observation after 4 h of treatment using an optical microscope with an oil-immersion objective lens (total magnification 1000×) indicates that lindane-treated M. aeruginosa shows a larger size (6 ± 0.5 mm diameter versus 4 ± 0.3 mm of the control culture of cells grown without the pesticide) (data not shown). Additionally, thylakoids seem to be less packed, and gas vesicles were absent when lindane was present in the culture media. Figure 1 shows the small differences in growth in both M. aeruginosa PCC 7806 cultures, control versus 7 mg/L lindane.
3.3. linC and prxA Are Induced in the Presence of Lindane in M. aeruginosa PCC 7806
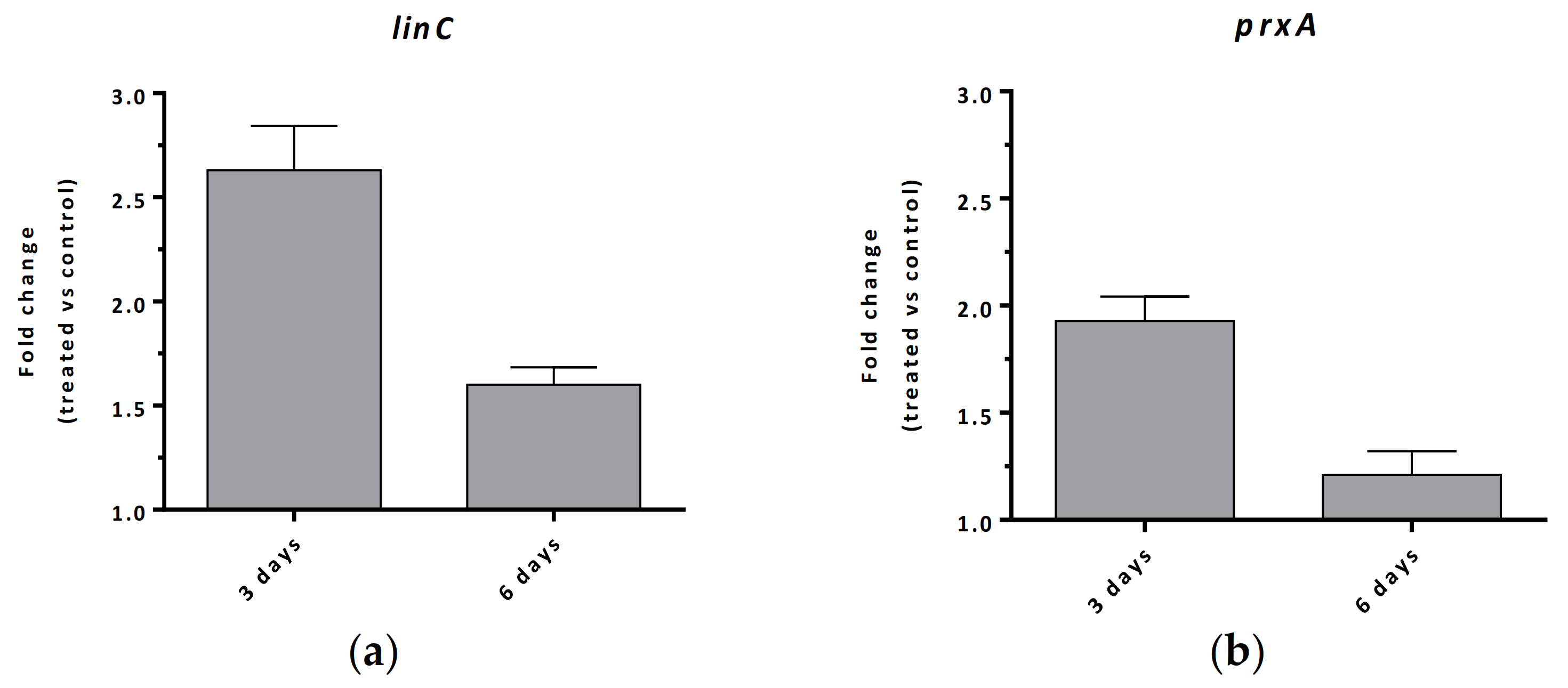
Cells from the cultures shown in Figure 1, at day 3 and day 6, were used to perform transcriptional analysis. RNA from M. aeruginosa PCC 7806 cell cultures performed in the presence and in absence of lindane was isolated to carry out the transcriptional response of the proposed linC gene (MAE_33900) for M. aeruginosa as well as the gene MAE_35830, hereafter prxA, that encode the 2-cys type peroxiredoxin PrxA orthologue (87% identity to PrxA from Anabaena sp. PCC 7120). The fold change of those genes expression after 3 and 6 days of lindane exposure is reported in Figure 2.
As seen in Figure 2a, the proposed linC gene for M. aeruginosa PCC 7806 increased its transcription after 3 and 6 days of treatment with lindane (7 mg/L). The value obtained at time 3 days was 2.6-fold, while at time 6 days it was 1.6-fold (Figure 2a). The decrease in linC induction at 6 days with respect to 3 days indicates that the transcriptional response to that lindane concentration begins to decline after 3 days, even though from our data it is not possible to set the time of the initiation of the decline. In the case of the peroxiredoxin prxA, it also displayed an increase in gene expression after 3 days of lindane exposure, up to 1.9-fold, whereas after 6 days no significant change versus control cells was observed (Figure 2b).
4. Discussion
As previously described [6], M. aeruginosa PCC 7806 not only tolerates high lindane concentrations but it is also able to degrade it, similarly to other cyanobacteria [1,2,3,4,5]. The lindane degradative pathway is unknown in cyanobacteria, and for this reason we considered it interesting to carry out an in silico study to try to identify the presence of homologues for lin genes described in Sphingomonadaceae [7] in M. aeruginosa PCC 7806. The initial reason for performing the experimental work with M. aeruginosa PCC 7806 was because this strain is a model strain, and the literature concerning Microcystis and microcystins is the most abundant. The entire genome of M. aeruginosa PCC 7806 is not yet available and, for this reason, the genome of M. aeruginosa NIES-843 has been used for bioinformatic studies. The high identity of the known sequences may permit the extrapolation of results [21], but the possibility of differences between the two genomes should not be ruled out. The identification of lin gene homologues in the M. aeruginosa NIES-843 genome has only been partial and, in fact, only a potential linC homologue has been found (Table 1). In the case of linA, linD, and linE, no results were obtained when a protein BLAST was performed, suggesting that either these genes are not present in M. aeruginosa NIES-843, or their sequences are highly divergent with respect to those of S. japonicum. This is not strange, since photosynthetic organisms may have different degradative pathways than the ones present in heterotrophs. With respect to the linB and linR genes, two putative genes were found in M. aeruginosa NIES-843 (Table 1), but their percentages of identity (26.2% and 15.2%, respectively) were lower than 30% and, consequently, were not high enough to be considered homologues of their respective lin genes of S. japonicum according to the criteria established by Rost [13].
In the case of the putative linC gene, the sequence found in M. aeruginosa NIES-843 holds a 34.4% of identity with respect to the linC gene of S. japonicum (Table 1). Consequently, as the percentage of identity was higher than 30% [13], it can be proposed as the putative linC of M. aeruginosa NIES-843. A phylogenetic tree of proteins codified by putative linC genes of other cyanobacteria revealed that this gene is also present in other species of cyanobacteria (Supplementary Material, Figure S4). Interestingly, in some of these bacteria, such as Synechococcus sp. WH 8020, Crocosphaera chwakensis CCY0110, and Microcoleus asticus IPMA8, it is already annotated as 2,5-dichloro-2,5-cyclohexadiene-1,4-diol dehydrogenase. These findings suggest that the 2,5-dichloro-2,5-cyclohexadiene-1,4-diol dehydrogenase enzyme could be present in several cyanobacteria and, perhaps, could allow them to tolerate and metabolize lindane.
The transcriptional analysis of the putative linC for M. aeruginosa revealed its induction after treatment with lindane (7 mg/L) (Figure 2a). In the heterotrophic bacteria S. japonicum UT26, linC is constitutively expressed [22]. However in M. aeruginosa PCC 7806, linC increased its expression after 3 and 6 days of lindane exposure (Figure 2a), suggesting that linC is an inducible gene that may take part in the lindane degradative pathway in this cyanobacterium. Moreover, the decrease in linC induction at 6 days with respect to 3 days suggest that the lower transcriptional response could be related to the time course of the intracellular concentration of lindane [6]. This behavior has been observed with linD and linE genes in S. japonicum UT26, whose induction disappears after 1 to 2 h, paralleling the degradation of lindane [22].
It was previously reported that in M. aeruginosa PCC 7806, the degradation of lindane by this cyanobacterium is coupled to an induction of a gene related to its own degradation, nirA, and genes related to the nitrogen and iron metabolism, ntcA and furA, respectively [6]. Additionally, mcy operon gene expression, as well as microcystin production, are likely enhanced due to oxidative stress caused by the pesticide [6]. The fact that in this work we found that the prxA gene increases the expression (Figure 2b) in the presence of lindane supports such a hypothesis. Peroxiredoxins are thought to play an important role in the reduction of endogenously generated H2O2 [23]. Therefore, the fact that expression of prxA increases after treatment with lindane (Figure 2b) would indicate that the internalization and/or degradation of lindane by this cyanobacterium is an example of it deploying its defenses against oxidative stress.
The identification of potential gene candidates to be involved in lindane degradation is just the first step in understanding the fate of lindane when is metabolized by cyanobacteria. Putative linC seems to be involved in the degradative pathway, based on its induction in the presence of pesticide. Future work needs to complete the identification of more candidates, for instance obtaining information by the determination of intermediaries of the degradation pathway. The interest of this work is that cyanobacteria present ideal ecophysiology characteristics to arise as a promising strategy to eliminate lindane residues in aquatic environments.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w14081219/s1, Table S1: Primers used in Real-Time PCR analyses, Figure S1: Pairwise alignment between LinB from S. japonicum UT26S and the protein codified by the gene MAE_40040 of Microcystis aeruginosa NIES-843, Figure S2: Pairwise alignment between LinC from S. japonicum UT26S and the protein codified by the gene MAE_33900 of Microcystis aeruginosa NIES-843, Figure S3: Pairwise alignment between LinR from S. japonicum UT26S and the protein codified by the gene MAE_62070 of Microcystis aeruginosa NIES-843, Figure S4: Phylogenetic tree of proteins codified by cyanobacterial putative linC genes. The Maximum Likelihood phylogenetic tree was built in Phylogeny [14] using PhyML with default parameters. To assure statistical significance, 100 bootstraps were used in the computation of the tree (bootstrap values are indicated in the tree branches). The annotation of each sequence is represented, and the name of the organism appears in brackets. The sequence corresponding to M. aeruginosa NIES-843 is highlighted in bold.
Author Contributions
Conceptualization, E.S., M.T.B., M.F.F. and M.L.P.; methodology, C.S.-B.; software, J.G.; formal analysis, C.S.-B.; investigation, C.C.; writing—original draft preparation, E.S., M.L.P.; writing—review and editing, C.S.-B., M.F.F., J.G. and M.L.P.; supervision, E.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work has been supported by grants E35_17R Biología Estructural from Gobierno de Aragón, BFU2016-77671-P/ FEDER and PID2019-104889 GB-I00 from MINECO and Ministerio de Ciencia e Innovación.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Kuritz, T.; Wolk, C.P. Use of filamentous cyanobacteria for biodegradation of organic pollutants. Appl. Environ. Microbiol. 1995, 61, 234–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuritz, T. An easy colorimetric assay for screening and qualitative assessment of deiodination and dehalogenation by bacterial cultures. Lett. Appl. Microbiol. 1999, 28, 445–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, G.S.; Hans, R.K.; Singh, J.; Viswanathan, P.N.; Joshi, P.C. Effect of Lindane on the Growth and Metabolic Activities of Cyanobacteria. Ecotoxicol. Environ. Saf. 2001, 48, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Bueno, M.; Fillat, M.F.; Strasser, R.J.; Maldonado-Rodriguez, R.; Marina, N.; Smienk, H.; Gomez-Moreno, C.; Barja, F. Effects of lindane on the photosynthetic apparatus of the cyanobacterium Anabaena: Fluorescence induction studies and immunolocalization of ferredoxin-NADP+ reductase. Environ. Sci. Pollut. Res. Int. 2004, 11, 98–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, R.; García-Balboa, C.; Rouco, M.; Lopez-Rodas, V.; Costas, E. Adaptation of microalgae to lindane: A new approach for bioremediation. Aquat. Toxicol. 2012, 109, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Ceballos-Laita, L.; Calvo-Begueria, L.; Lahoz, J.; Bes, M.-T.; Fillat, M.F.; Peleato, M.-L. γ-Lindane Increases Microcystin Synthesis in Microcystis aeruginosa PCC7806. Mar. Drugs 2015, 13, 5666–5680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, Y.; Endo, R.; Ito, M.; Ohtsubo, Y.; Tsuda, M. Aerobic degradation of lindane (γ-hexachlorocyclohexane) in bacteria and its biochemical and molecular basis. Appl. Microbiol. Biotechnol. 2007, 76, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Cuozzo, S.A.; Sineli, P.E.; Davila Costa, J.; Tortella, G. Streptomyces sp. is a powerful biotechnological tool for the biodegradation of HCH isomers: Biochemical and molecular basis. Crit. Rev. Biotechnol. 2018, 38, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- UniPro. Available online: http://www.uniprot.org (accessed on 2 September 2021).
- CyanoBase: Similarity Search. Available online: http://genome.microbedb.jp/blast/blast_search/cyanobase/genes (accessed on 2 September 2021).
- EMBOSS Needle. Available online: https://www.ebi.ac.uk/Tools/psa/emboss_needle/ (accessed on 2 September 2021).
- Rost, B. Twilight zone of protein sequence alignments. Protein Eng. 1999, 12, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Phylogeny.fr. Available online: https://www.phylogeny.fr/ (accessed on 2 September 2021).
- NIH BLASTp suite. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE=Proteins (accessed on 2 September 2021).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Nakajima, N.; Okamoto, S.; Suzuki, I.; Tanabe, Y.; Tamaoki, M.; Nakamura, Y.; Kasai, F.; Watanabe, A.; Kawashima, K.; et al. Complete Genomic Structure of the Bloom-forming Toxic Cyanobacterium Microcystis aeruginosa NIES-843. DNA Res. 2007, 14, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Suar, M.; van der Meer, J.R.; Lawlor, K.; Holliger, C.; Lal, R. Dynamics of Multiple lin Gene Expression in Sphingomonas paucimobilis B90A in Response to Different Hexachlorocyclohexane Isomers. Appl. Environ. Microbiol. 2004, 70, 6650–6656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latifi, A.; Ruiz, M.; Jeanjean, R.; Zhang, C.-C. PrxQ-A, a member of the peroxiredoxin Q family, plays a major role in defense against oxidative stress in the cyanobacterium Anabaena sp. strain PCC7120. Free Radic. Biol. Med. 2007, 42, 424–431. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
M. aeruginosa PCC 7806 growth curve. ♦ Control: cells grown in BG 11 with 2 mM NaNO3, ■ Cells grown in the same conditions in the presence of 7 mg/L of lindane.
Figure 1.
M. aeruginosa PCC 7806 growth curve. ♦ Control: cells grown in BG 11 with 2 mM NaNO3, ■ Cells grown in the same conditions in the presence of 7 mg/L of lindane.

Figure 2.
Relative transcription in M. aeruginosa PCC 7806 as a consequence of 7 mg/L lindane treatment of the (a) putative linC and (b) prxA genes. Values are expressed as fold change and correspond to the average of three biological and two technical replicates; the standard deviation is indicated.
Figure 2.
Relative transcription in M. aeruginosa PCC 7806 as a consequence of 7 mg/L lindane treatment of the (a) putative linC and (b) prxA genes. Values are expressed as fold change and correspond to the average of three biological and two technical replicates; the standard deviation is indicated.


{kind=link}
{kind=link}
Table 1.
Results of the identification of putative lin genes in Microcystis aeruginosa NIES-843. For each of the lin genes of Sphingobium japonicum UT26S, the gene of M. aeruginosa NIES-843 that presented the lowest expectancy value according to the protein BLAST is annotated, and the parameters of the pairwise alignment between these two sequences are indicated (sequence alignments are included in Supplementary Material, Figures S1–S3). In the case of linA, linD, and linE no sequences presenting an expectancy value lower than the threshold (0.005) were found.
Table 1.
Results of the identification of putative lin genes in Microcystis aeruginosa NIES-843. For each of the lin genes of Sphingobium japonicum UT26S, the gene of M. aeruginosa NIES-843 that presented the lowest expectancy value according to the protein BLAST is annotated, and the parameters of the pairwise alignment between these two sequences are indicated (sequence alignments are included in Supplementary Material, Figures S1–S3). In the case of linA, linD, and linE no sequences presenting an expectancy value lower than the threshold (0.005) were found.
| Sphingobium japonicum UT26S | Microcystis aeruginosa NIES-843 | Pairwise Alignment | |||||
|---|---|---|---|---|---|---|---|
| Gene | Anotation | Gene | Anotation | Identity | Similarity | Gaps | Score |
| linA | Gamma-hexachlorocyclohexane dehydrochlorinase | - | - | - | - | - | - |
| linB | Haloalkane dehalogenase | MAE_40040 | putative hydrolase | 26.2% | 41.2% | 17.2% | 171.0 |
| linC | 2,5-dichloro-2,5-cyclohexadiene-1,4-diol dehydrogenase | MAE_33900 | 3-oxoacyl-[acyl-carrier protein] reductase | 34.4% | 50.0% | 7.6% | 346.0 |
| linD | 2,5-dichlorohydroquinone reductive dechlorinase | - | - | - | - | - | - |
| linE | Chlorohydroquinone/hydroquinone 1,2-dioxygenase | - | - | - | - | - | - |
| linR | HTH-type transcriptional regulator LinR | MAE_62070 | transcriptional regulator | 15.2% | 27.8% | 46.2% | 71.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sarasa-Buisán, C.; Guío, J.; Castro, C.; Bes, M.T.; Fillat, M.F.; Peleato, M.L.; Sevilla, E. Contributions on Lindane Degradation by Microcystis aeruginosa PCC 7806. Water 2022, 14, 1219. https://doi.org/10.3390/w14081219
AMA Style
Sarasa-Buisán C, Guío J, Castro C, Bes MT, Fillat MF, Peleato ML, Sevilla E. Contributions on Lindane Degradation by Microcystis aeruginosa PCC 7806. Water. 2022; 14(8):1219. https://doi.org/10.3390/w14081219
Chicago/Turabian StyleSarasa-Buisán, Cristina, Jorge Guío, Carolina Castro, María Teresa Bes, María F. Fillat, María Luisa Peleato, and Emma Sevilla. 2022. "Contributions on Lindane Degradation by Microcystis aeruginosa PCC 7806" Water 14, no. 8: 1219. https://doi.org/10.3390/w14081219
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.