
The Coupling Response between Different Bacterial Metabolic Functions in Water and Sediment Improve the Ability to Mitigate Climate Change

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mesocosm Experiment Set up and Management
2.2. Sampling and Environmental Physicochemical Properties
2.3. DNA Extraction, PCR Amplification
2.4. Illumina MiSeq Sequencing
2.5. Sequence Denoising, OTU Clustering and FAPROTAX Function Prediction
2.6. Co-Occurrence Network Analysis
2.7. Statistical Analysis
3. Results
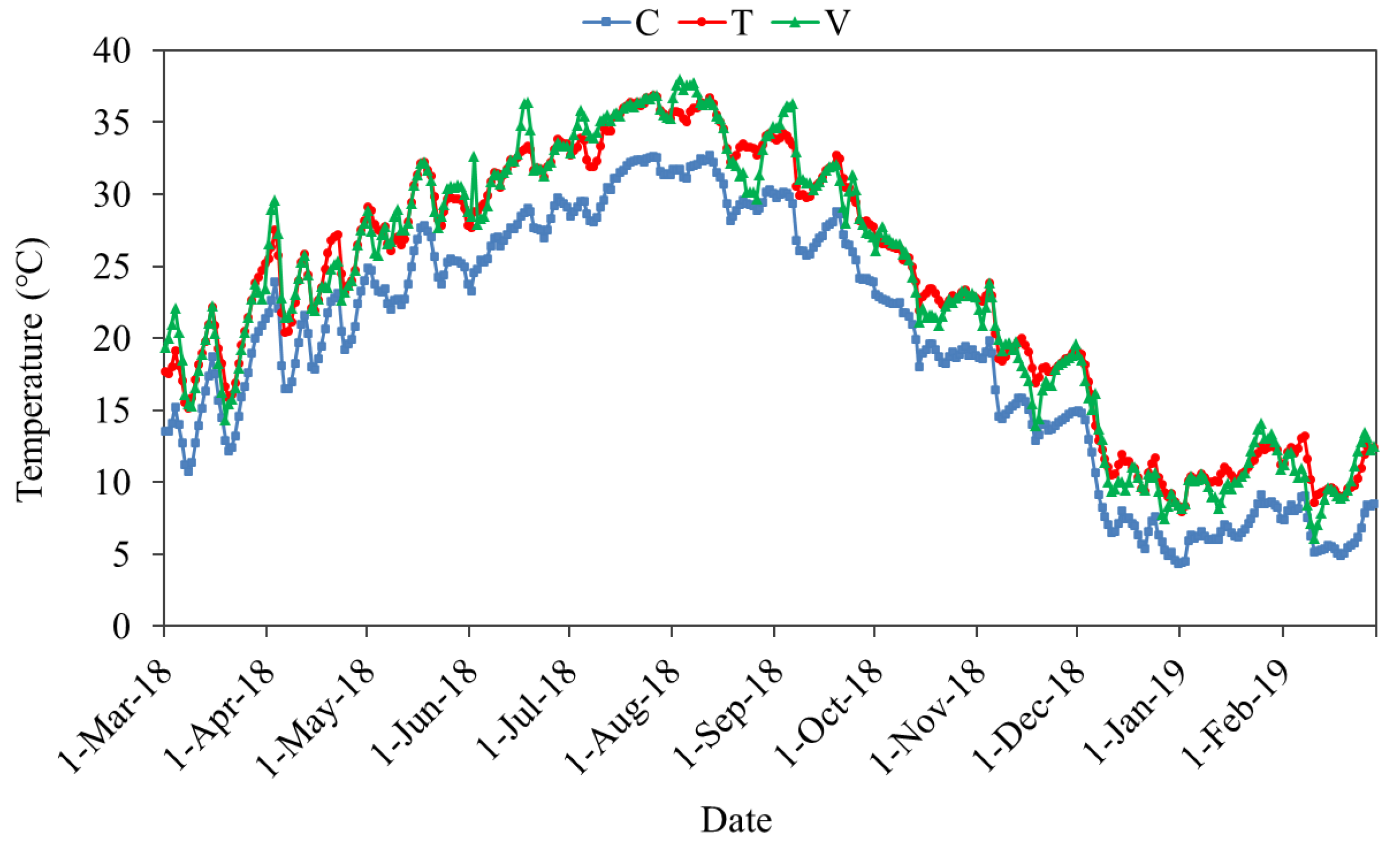
3.1. Warming Effects on Physical and Chemical Characteristics
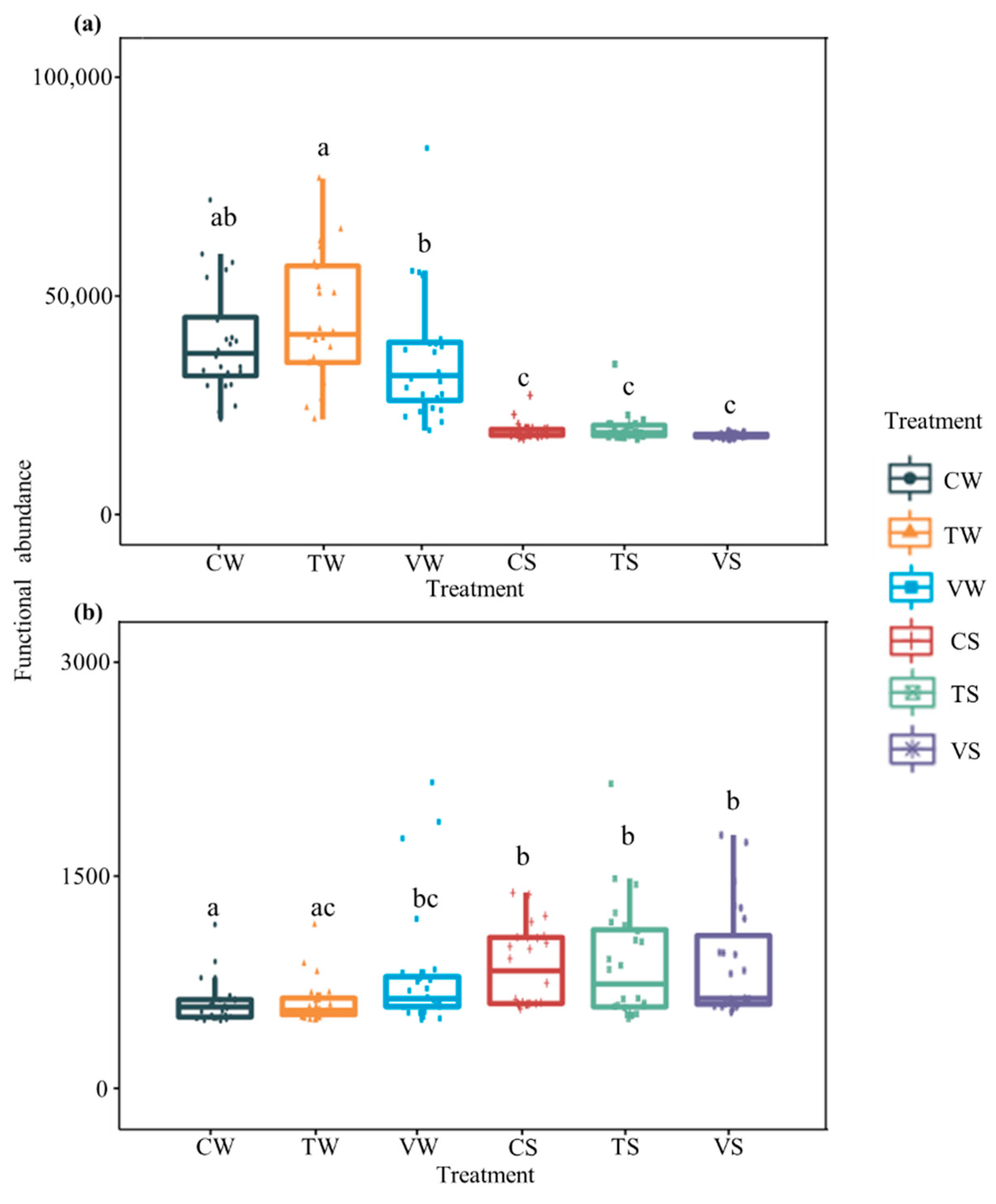
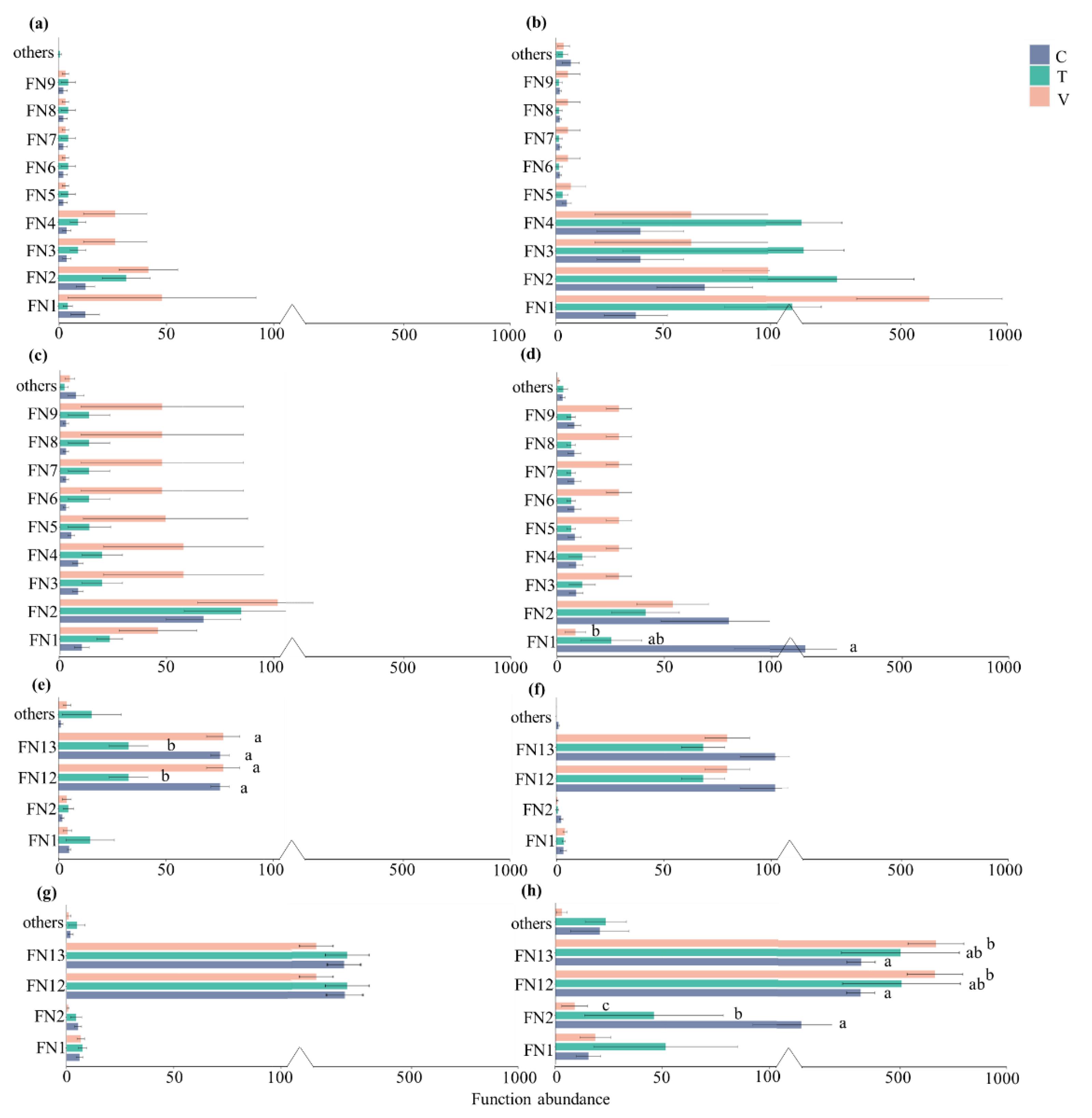
3.2. Bacterial Functional Abundance Analysis
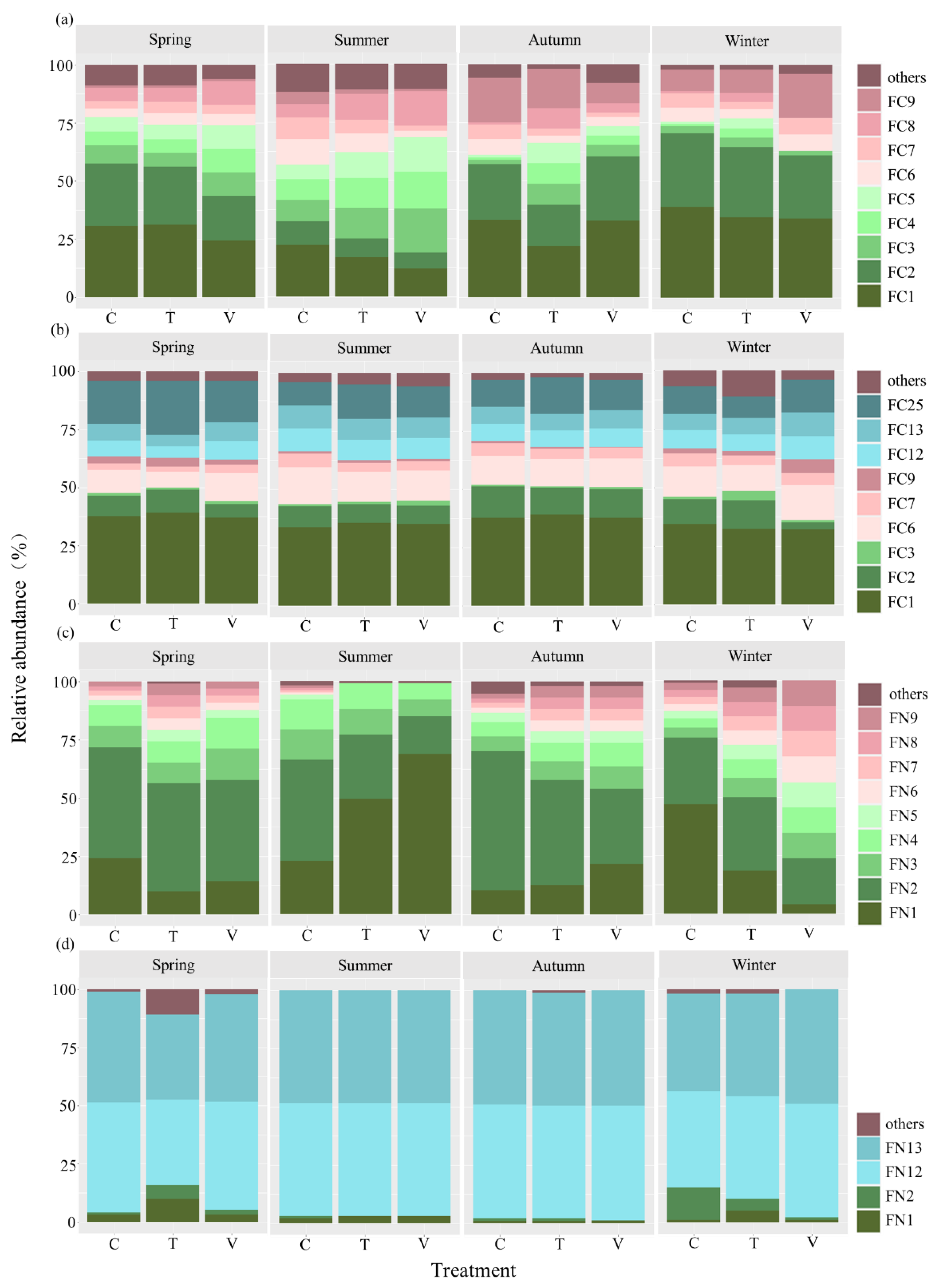
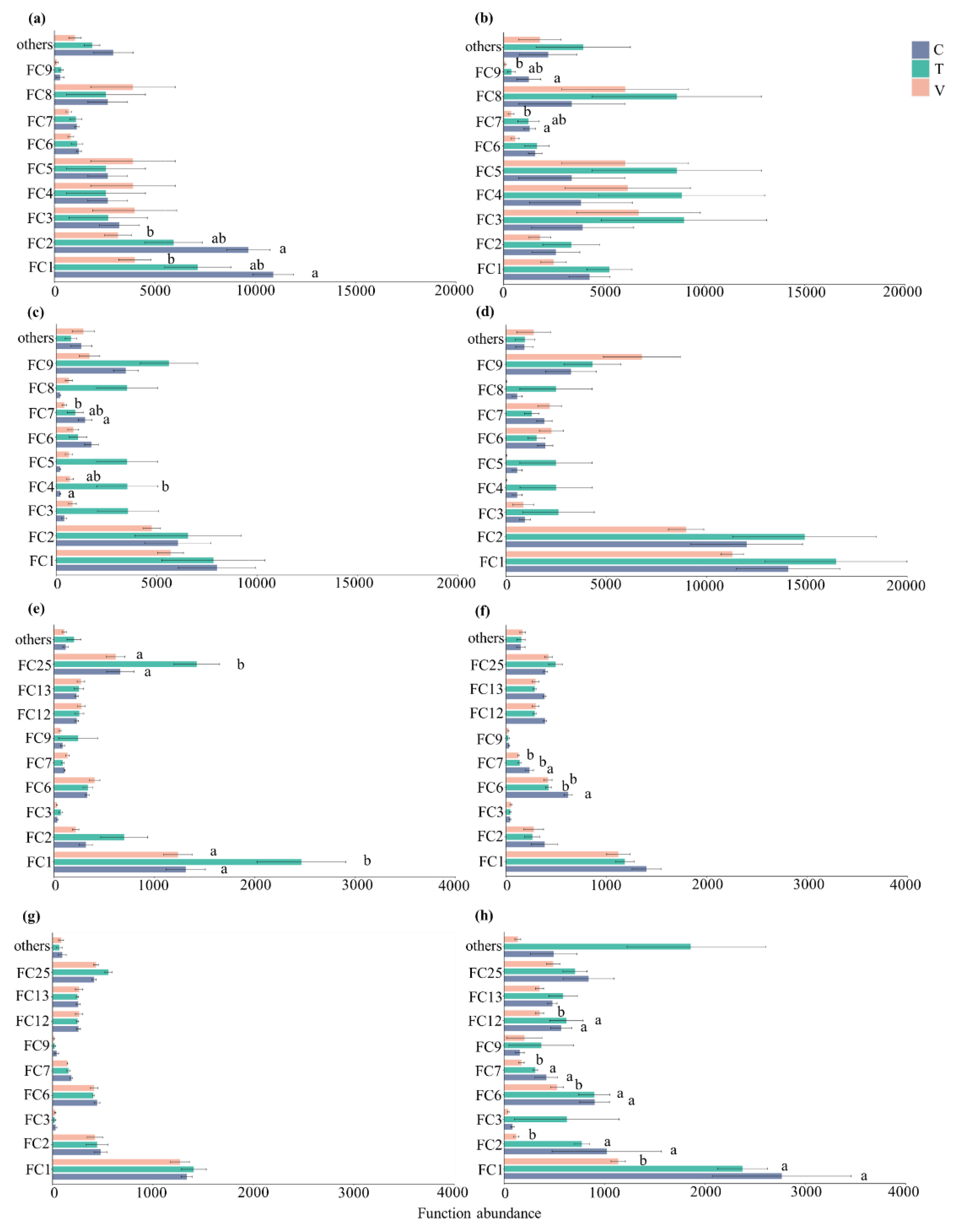
3.3. Composition of Bacterial Functional Community
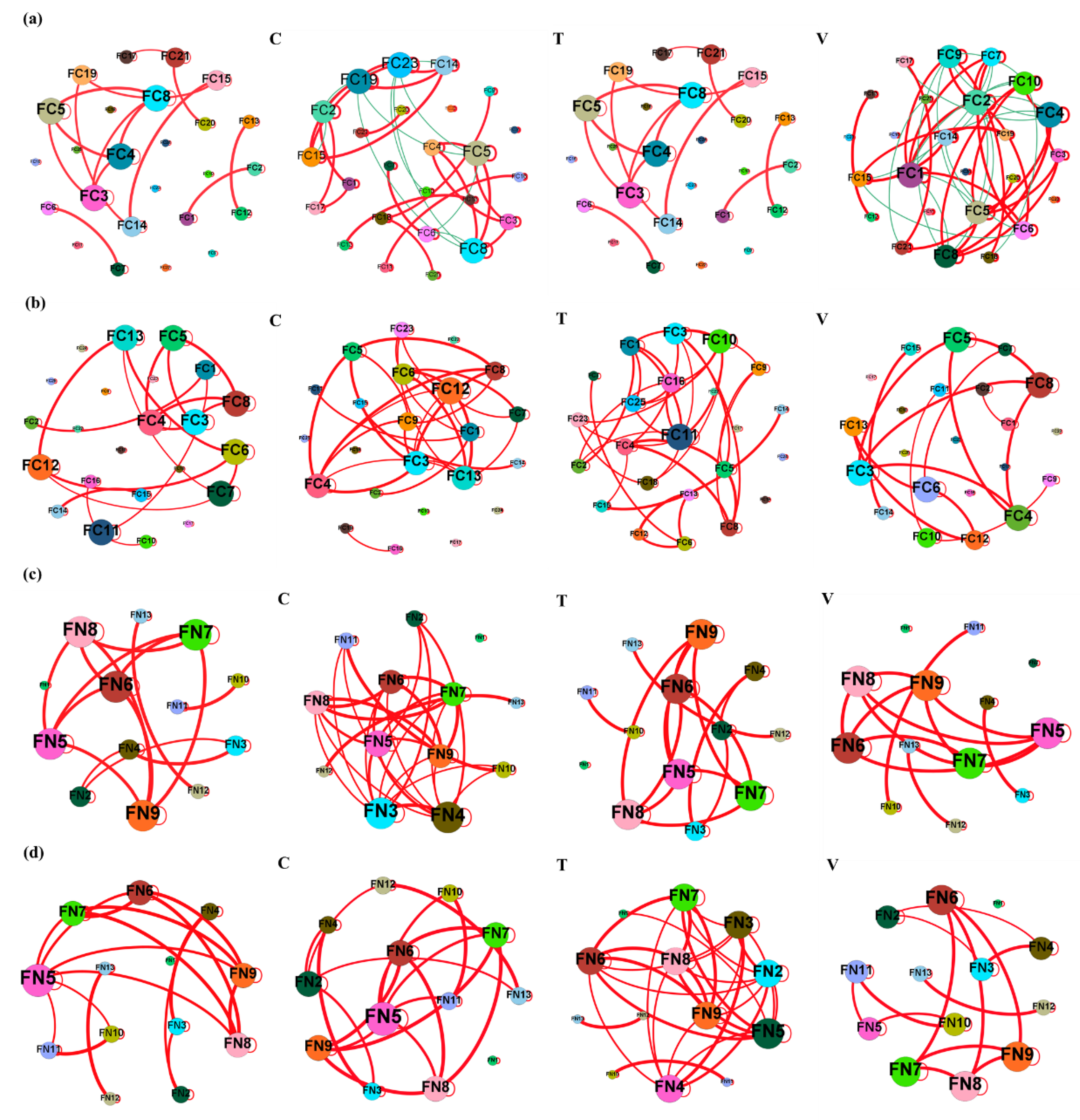
3.4. Co-Occurrence Networks and Topological Features
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Tignor, M., Allen, S., Boschung, J., Nauels, A., Xia, Y., Bex, V., Pm, M., Eds.; Cambridge University Press: New York, NY, USA, 2013; p. 1535. [Google Scholar]
- O’Reilly, C.M.; Sharma, S.; Gray, D.K.; Hampton, S.E.; Read, J.S.; Rowley, R.J.; Schneider, P.; Lenters, J.D.; McIntyre, P.B.; Kraemer, B.M.; et al. Rapid and highly variable warming of lake surface waters around the globe. Geophys. Res. Lett. 2015, 42, 773–781. [Google Scholar] [CrossRef] [Green Version]
- Seneviratne, S.I.; Donat, M.G.; Mueller, B.; Alexander, L.V. No pause in the increase of hot temperature extremes. Nat. Clim. Chang. 2014, 4, 161–163. [Google Scholar] [CrossRef]
- Davidson, T.A.; Audet, J.; Jeppesen, E.; Landkildehus, F.; Lauridsen, T.L.; Søndergaard, M.; Syväranta, J. Synergy between nutrients and warming enhances methane ebullition from experimental lakes. Nat. Clim. Chang. 2018, 8, 156–160. [Google Scholar] [CrossRef]
- Downing, J.A.; Prairie, Y.T.; Cole, J.J.; Duarte, C.M.; Tranvik, L.J.; Striegl, R.G.; McDowell, W.H.; Kortelainen, P.; Caraco, N.F.; Melack, J.M.; et al. The global abundance and size distribution of lakes, ponds, and impoundments. Limnol. Oceanogr. 2006, 51, 2388–2397. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; He, L.; Zhang, H.; Urrutia-Cordero, P.; Ekvall, M.K.; Hollander, J.; Hansson, L.A. Climate warming and heat waves affect reproductive strategies and interactions between submerged macrophytes. Glob. Chang. Biol. 2016, 23, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Praetzel, L.S.E.; Schmiedeskamp, M.; Knorr, K. Temperature and sediment properties drive spatiotemporal variability of methane ebullition in a small and shallow temperate lake. Limnol. Oceanogr. 2021, 66, 2598–2610. [Google Scholar] [CrossRef]
- Parmesan, C. Ecological and Evolutionary Responses to Recent Climate Change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- Reyer, C.P.O.; Leuzinger, S.; Rammig, A.; Wolf, A.; Bartholomeus, R.P.; Bonfante, A.; De Lorenzi, F.; Dury, M.; Gloning, P.; Jaoudé, R.A.; et al. A plant’s perspective of extremes: Terrestrial plant responses to changing climatic variability. Glob. Chang. Biol. 2012, 19, 75–89. [Google Scholar] [CrossRef]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: Impacts across multiple levels of organization. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2093–2106. [Google Scholar] [CrossRef] [Green Version]
- Newton, R.J.; Jones, S.E.; Eiler, A.; Mcmahon, K.D.; Bertilsson, S. A guide to the natural history of freshwater lake bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macarena, M.; Jeannette, V. Microorganisms that participate in biochemical cycles in wetlands. Can. J. Microbiol. 2021, 67, 771–788. [Google Scholar] [CrossRef]
- Zhang, L.; Lv, J. Land-use change from cropland to plantations affects the abundance of nitrogen cycle-related microorganisms and genes in the Loess Plateau of China. Appl. Soil Ecol. 2021, 161, 103873. [Google Scholar] [CrossRef]
- Zingel, P.; Cremona, F.; Nõges, T.; Cao, Y.; Neif, É.; Coppens, J.; Işkın, U.; Lauridsen, T.L.; Davidson, T.A.; Søndergaard, M. Effects of warming and nutrients on the microbial food web in shallow lake mesocosms. Eur. J. Protistol. 2018, 64, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Liu, Y.; Lauridsen, T.L.; Sndergaard, M.; Han, B.; Wang, J.; Jeppesen, E.; Zhou, J.; Wu, Q.L. Warming exacerbates the impact of nutrient enrichment on microbial functional potentials important to the nutrient cycling in shallow lake mesocosms. Limnol. Oceanogr. 2021, 66, 2481–2495. [Google Scholar] [CrossRef]
- Fierer, N.; Ladau, J.; Clemente, J.C.; Leff, J.W.; Owens, S.M.; Pollard, K.S.; Knight, R.; Gilbert, J.A.; Mcculley, R.L. Reconstructing the Microbial Diversity and Function of Pre-Agricultural Tallgrass Prairie Soils in the United States. Science 2013, 342, 621–624. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, S.M. Microbial community ecology: Function over phylogeny. Nat. Ecol. Evol. 2017, 1, 32. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.B.; Martiny, A.C.; Martiny, J.B.H. Global biogeography of microbial nitrogen-cycling traits in soil. Proc. Natl. Acad. Sci. USA 2016, 113, 8033–8040. [Google Scholar] [CrossRef] [Green Version]
- Coles, V.J.; Stukel, M.R.; Brooks, M.T.; Burd, A.; Crump, B.C.; Moran, M.A.; Paul, J.H.; Satinsky, B.M.; Yager, P.L.; Zielinski, B.L. Ocean biogeochemistry modeled with emergent trait-based genomics. Science 2017, 358, 1149–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanwonterghem, I.; Jensen, P.D.; Rabaey, K.; Tyson, G.W. Genome-centric resolution of microbial diversity, metabolism and interactions in anaerobic digestion. Environ. Microbiol. 2016, 18, 3144–3158. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Doebeli, M. Taxonomic variability and functional stability in microbial communities infected by phages. Environ. Microbiol. 2017, 19, 3863–3878. [Google Scholar] [CrossRef] [PubMed]
- Barberán, A.; Fernández-Guerra, A.; Bohannan, B.; Casamayor, E.O. Exploration of community traits as ecological markers in microbial metagenomes. Mol. Ecol. 2012, 21, 1909–1917. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, M.I.; Piehler, M.F.; Leech, D.M.; Anton, A.; Bruno, J.F. Warming and Resource Availability Shift Food Web Structure and Metabolism. PLoS Biol. 2009, 7, 1000178. [Google Scholar] [CrossRef]
- Özen, A.; Šorf, M.; Trochine, C.; Liboriussen, L.; Jeppesen, E. Long-term effects of warming and nutrients on microbes and other plankton in mesocosms. Freshw. Biol. 2013, 58, 483–493. [Google Scholar] [CrossRef]
- Posch, T.; Köster, O.; Salcher, M.M.; Pernthaler, J. Harmful filamentous cyanobacteria favoured by reduced water turnover with lake warming. Nat. Clim. Chang. 2012, 2, 809–813. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Shen, T.; Cheng, Y.; Zhao, T.; Qi, P. Temporal and spatial variations in the bacterial community composition in Lake Bosten, a large, brackish lake in China. Sci. Rep. 2020, 10, 304. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, M.; Liu, F.; Chen, L.; Li, Y.; Li, Y.; Xiao, R.; Wu, J. Seasonality distribution of the abundance and activity of nitrification and denitrification microorganisms in sediments of surface flow constructed wetlands planted with Myriophyllum elatinoides during swine wastewater treatment. Bioresour. Technol. 2018, 248, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, Y.; He, W.; Wu, M.; Xing, M.; Yang, J.; Gao, N.; Pan, M. Impacts of temperature and nitrifying community on nitrification kinetics in a moving-bed biofilm reactor treating polluted raw water. Chem. Eng. J. 2014, 236, 242–250. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, D.; Saha, S. Response of Soil Properties and Soil Microbial Communities to the Projected Climate Change. In Advances in Crop Environment Interaction; Springer: Singapore, 2018; pp. 87–136. [Google Scholar]
- Zhou, X.; Li, B.; Wei, J.; Ye, Y.; Lu, C. Temperature Influenced the Comammox Community Composition in Drinking Water and Wastewater Treatment Plants. Microb. Ecol. 2021, 82, 870–884. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, Y.; He, Y.; Brookes, P.C.; Xu, J. Elevated temperature increased nitrification activity by stimulating AOB growth and activity in an acidic paddy soil. Plant Soil 2019, 445, 71–83. [Google Scholar] [CrossRef]
- Razavi, B.S.; Liu, S.; Kuzyakov, Y. Hot experience for cold-adapted microorganisms: Temperature sensitivity of soil enzymes. Soil Biol. Biochem. 2017, 105, 236–243. [Google Scholar] [CrossRef]
- Minett, D.A.; Cook, P.L.M.; Kessler, A.J.; Cavagnaro, T.R. Root effects on the spatial and temporal dynamics of oxygen in sand-based laboratory-scale constructed biofilters. Ecol. Eng. 2013, 58, 414–422. [Google Scholar] [CrossRef]
- Thompson, R.M.; Beardall, J.; Beringer, J.; Grace, M.; Sardina, P.; Lloret, F. Means and extremes: Building variability into community-level climate change experiments. Ecol. Lett. 2013, 16, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.; Reichstein, M.; Bahn, M.; Thonicke, K.; Frank, D.; Mahecha, M.D.; Smith, P.; van der Velde, M.; Vicca, S.; Babst, F.; et al. Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts. Glob. Chang. Biol. 2015, 21, 2861–2880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audet, J.; Neif, É.M.; Cao, Y.; Hoffmann, C.C.; Lauridsen, T.L.; Larsen, S.E.; Søndergaard, M.; Jeppesen, E.; Davidson, T.A. Heat-wave effects on greenhouse gas emissions from shallow lake mesocosms. Freshw. Biol. 2017, 62, 1130–1142. [Google Scholar] [CrossRef]
- Font, R.A.; Khamis, K.; Milner, A.M.; Smith, G.; Ledger, M.E. Low flow and heatwaves alter ecosystem functioning in a stream mesocosm experiment. Sci. Total Environ. 2021, 777, 146067. [Google Scholar] [CrossRef] [PubMed]
- Misson, L.; Degueldre, D.; Collin, C.; Rodriguez, R.; Rocheteau, A.; Ourcival, J.M.; Rambal, S. Phenological responses to extreme droughts in a Mediterranean forest. Glob. Chang. Biol. 2011, 17, 1036–1048. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, H.; Wang, H.; Hilt, S.; Li, C.; Yu, C.; Zhang, M.; Xu, J. Warming alters juvenile carp effects on macrophytes resulting in a shift to turbid conditions in freshwater mesocosms. J. Appl. Ecol. 2022, 59, 165–175. [Google Scholar] [CrossRef]
- Wu, L.; Yang, F.; Feng, J.; Tao, X.; Qi, Q.; Wang, C.; Schuur, E.A.G.; Bracho, R.; Huang, Y.; Cole, J.R.; et al. Permafrost thaw with warming reduces microbial metabolic capacities in subsurface soils. Mol. Ecol. 2022, 31, 1403–1415. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, T.; Molinos, J.G.; Li, C.; Hu, B.; Pan, M.; Zhang, M. Effects of warming, climate extremes and phosphorus enrichment on the growth, sexual reproduction and propagule carbon and nitrogen stoichiometry of Potamogeton crispus L. Environ. Int. 2020, 137, 105502. [Google Scholar] [CrossRef]
- Diehl, S.; Berger, S.A.; Uszko, W.; Stibor, H. Stoichiometric mismatch causes a warming-induced regime shift in experimental plankton communities. Ecology 2022, e3674. [Google Scholar] [CrossRef] [PubMed]
- Moran, R.; Harvey, I.; Moss, B.; Feuchtmayr, H.; Hatton, K.; Heyes, T.; Atkinson, D. Influence of simulated climate change and eutrophication on three-spined stickleback populations: A large scale mesocosm experiment. Freshw. Biol. 2010, 55, 315–325. [Google Scholar] [CrossRef]
- Demko, A.M.; Patin, N.V.; Jensen, P.R. Microbial diversity in tropical marine sediments assessed using culture-dependent and culture-independent techniques. Environ. Microbiol. 2021, 23, 6859–6875. [Google Scholar] [CrossRef] [PubMed]
- Liao, B.; Yan, X.; Zhang, J.; Chen, M.; Li, Y.; Huang, J.; Lei, M.; He, H.; Wang, J. Microbial community composition in alpine lake sediments from the Hengduan Mountains. MicrobiologyOpen 2019, 8, e00832. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; Mcdonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knightet, R. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Naturebiotechnology 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Ward, T.; Larson, J.; Meulemans, J.; Hillmann, B.; Lynch, J.; Sidiropoulos, D.; Spear, J.R.; Caporaso, G.; Blekhman, R.; Knight, R.; et al. BugBase predicts organism-level microbiome phenotypes. bioRxiv 2017, 133462. [Google Scholar]
- Aßhauer, K.; Wemheuer, B.; Daniel, R.; Peter, M. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef]
- Pan, M.; Wang, T.; Hu, B.W.; Shi, P.L.; Xu, J.; Zhang, M. Mesocosm Experiments Reveal Global Warming Accelerates Macrophytes Litter Decomposition and Alters Decomposition-Related Bacteria Community Structure. Water 2021, 13, 1940. [Google Scholar] [CrossRef]
- Ren, L.; He, D.; Chen, Z.; Jeppesen, E.; Lauridsen, T.L.; Sondergaard, M.; Liu, Z.; Wu, Q.L. Warming and nutrient enrichment in combination increase stochasticity and beta diversity of bacterioplankton assemblages across freshwater mesocosms. ISME J. 2017, 11, 613–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Luo, Y.; Xia, J.; Jiang, L.; Zhou, X.; Lu, M.; Liang, J.; Shi, Z.; Shelton, S.; Cao, J. Stronger warming effects on microbial abundances in colder regions. Sci. Rep. 2015, 5, 18032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, P.; Tao, Y.; Jeppesen, E.; Wu, Q.L. Comparing microbial composition and diversity in freshwater lakes between Greenland and the Tibetan Plateau. Limnol. Oceanogr. 2021, 66, S142–S156. [Google Scholar] [CrossRef]
- Dong, W.; Min, S.; Tang, L.; Zeng, Q.; Yang, J.; Yao, M. Research of Eutrophication of Honghu Lake and Liangzi Lake. Environ. Prot. Sci. 2016, 42, 66–70. [Google Scholar]
- Li, C.; Wang, T.; Zhang, M.; Xu, J. Maternal Environment Effect of Warming and Eutrophication on the Emergence of Curled Pondweed, Potamogeton crispus L. Water 2018, 10, 1285. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Huang, J.; Luo, Y.; Zhao, Z.; Xu, Y. Projection of heat waves over China for eight different global warming targets using 12 CMIP5 models. Theor. Appl. Climatol. 2017, 128, 507–522. [Google Scholar] [CrossRef]
- Jin, X.C.; Tu, Q.Y. Investigation Specifications for Lake Eutrophication, 2nd ed.; China Environmental Science Press: Beijing, China, 1990. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Jespersen, A.; Christoffersen, K.J. Measurements of chlorophyll-a from phytoplankton using ethanol as extraction solvent. Arch. Hydrobiol. 1987, 109, 445–454. [Google Scholar]
- Mori, H.; Maruyama, F.; Kato, H.; Toyoda, A.; Dozono, A.; Ohtsubo, Y.; Nagata, Y.; Fujiyama, A.; Tsuda, M.; Kurokawa, K. Design and Experimental Application of a Novel Non-Degenerate Universal Primer Set that Amplifies Prokaryotic 16S rRNA Genes with a Low Possibility to Amplify Eukaryotic rRNA Genes. DNA Res. 2014, 21, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Jia, G. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Handelsman, J. Introducing DOTUR, a Computer Program for Defining Operational Taxonomic Units and Estimating Species Richness. Appl. Environ. Microb. 2005, 71, 1501–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picazo, A.; Villaescusa, J.A.; Rochera, C.; Miralles-Lorenzo, J.; Quesada, A.; Camacho, A. Functional Metabolic Diversity of Bacterioplankton in Maritime Antarctic Lakes. Microorganisms 2021, 9, 2077. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Deng, J.; Jiang, Y.; Wu, S.; Zhou, Y.; Zhu, W. Functional Distribution of Bacterial Community under Different Land Use Patterns Based on FaProTax Function Prediction. Pol. J. Environ. Stud. 2020, 29, 1245–1261. [Google Scholar] [CrossRef]
- Wei, H.; Peng, C.; Yang, B.; Song, H.; Li, Q.; Jiang, L.; Wei, G.; Wang, K.; Wang, H.; Liu, S.; et al. Contrasting Soil Bacterial Community, Diversity, and Function in Two Forests in China. Front. Microbiol. 2018, 9, 1693. [Google Scholar] [CrossRef] [PubMed]
- Barberan, A.; Bates, S.T.; Casamayor, E.O.; Fiere, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Shi, Y.; Jing, X.; He, J.-S.; Sun, R.; Yang, Y.; Shade, A.; Chu, H. Effects of Short-Term Warming and Altered Precipitation on Soil Microbial Communities in Alpine Grassland of the Tibetan Plateau. Front. Microbiol. 2016, 7, 1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, Q.; Yan, Q.; Deng, Y.; Michaletz, S.T.; Zhou, J. Biogeographic patterns of microbial co-occurrence ecological networks in six American forests. Soil Biol. Biochem. 2020, 148, 107897. [Google Scholar] [CrossRef]
- Jiao, S.; Yang, Y.; Xu, Y.; Zhang, J.; Lu, Y. Balance between community assembly processes mediates species coexistence in agricultural soil microbiomes across eastern China. ISME J. 2020, 14, 202–216. [Google Scholar] [CrossRef]
- Newman, M. Modularity and community structure in networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. ICWSM 2009, 3, 361–362. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. Statistics and Computing. arXiv 2014, arXiv:14065823. [Google Scholar]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Package “Emmeans”. R Package Version 4.0-3. 2018. Available online: http://cran.r-project.org/package=emmeans (accessed on 15 February 2021).
- Budy, P.; Pennock, C.A.; Giblin, A.E.; Luecke, C.; Kling, G.W. Understanding the effects of climate change via disturbance on pristine arctic lakes-multitrophic level response and recovery to a 12-yr, low-level fertilization experiment. Limnol. Oceanogr. 2021, 67, S224–S241. [Google Scholar] [CrossRef]
- Belle, S.; Tõnno, I.; Vrede, T.; Freiberg, R.; Nilsson, J.L.; Goedkoop, W. Both climate trends and short-term fluctuations affected algae–zooplankton interactions in a boreal lake during the late Holocene. Freshwater Biol. 2021, 66, 2076–2085. [Google Scholar] [CrossRef]
- Wei, Z.; Liu, Y.; Kai, F.; Li, S.; Shang, W.; Jin, D.; Yu, Z.; Chen, H.; Yin, H.; Xu, M. The divergence between fungal and bacterial communities in seasonal and spatial variations of wastewater treatment plants. Sci. Total Environ. 2018, 628, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Xiang, X.; Yang, Y.; Huang, G.; Fu, K.; Che, R.; Chen, L. Seasonal effects of river flow on microbial community coalescence and diversity in a riverine network. FEMS Microbiol. Ecol. 2020, 96, fiaa132. [Google Scholar] [CrossRef]
- Rivett, D.W.; Bell, T. Abundance determines the functional role of bacterial phylotypes in complex communities. Nat. Microbiol. 2018, 3, 767–772. [Google Scholar] [CrossRef]
- Pan, M. Effects of Climate Warming on Aquatic Bacterial Community Structure and Decomposition of Submerged Macrophytes Litter. Master Thesis, HuaZhong Agricultural University, Wuhan, China, 2021. [Google Scholar]
- Dutta, H.; Dutta, A. The microbial aspect of climate change. Energy Ecol. Environ. 2016, 1, 209–232. [Google Scholar] [CrossRef] [Green Version]
- Hasler, C.T.; Butman, D.; Jeffrey, J.D.; Suski, C.D. Freshwater biota and rising pCO2? Ecol. Lett. 2015, 19, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Hood, J.M.; Benstead, J.P.; Cross, W.F.; Huryn, A.D.; Johnson, P.W.; Gíslason, G.M.; Junker, J.R.; Nelson, D.; Ólafsson, J.S.; Tran, C. Increased resource use efficiency amplifies positive response of aquatic primary production to experimental warming. Glob. Chang. Biol. 2018, 24, 1069–1084. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Chen, Y.; Qin, M.; Mao, Z.; Yuan, L.; Niu, Q.; Tan, X. Effects of temperature on anammox performance and community structure. Bioresour. Technol. 2018, 260, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Eduok, S.; Ferguson, R.; Jefferson, B.; Villa, R.; Coulon, F. Aged-engineered nanoparticles effect on sludge anaerobic digestion performance and associated microbial communities. Sci. Total Environ. 2017, 609, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeks-Johnson, B.M.; Cotner, J.B. Upper Midwest lakes are supersaturated with N2. Proc. Natl. Acad. Sci. USA 2020, 117, 17063–17067. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hu, M.; Ren, H.; Li, J.; Tong, C.; Musenze, R.S. Seasonal variations of nitrous oxide fluxes and soil denitrification rates in subtropical freshwater and brackish tidal marshes of the Min River estuary. Sci. Total Environ. 2018, 616–617, 1404–1413. [Google Scholar] [CrossRef] [PubMed]
- De Klein, J.J.M.; Overbeek, C.C.; Juncher Jrgensen, C.; Veraart, A.J. Effect of Temperature on Oxygen Profiles and Denitrification Rates in Freshwater Sediments. Wetlands 2017, 37, 975–983. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Chen, N.; Hong, H.; Lu, T.; Wang, L.; Chen, Z. Direct measurement of dissolved N2 and denitrification along a subtropical river-estuary gradient, China. Mar. Pollut. Bull. 2013, 66, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Sand-Jensen, K.; Prahl, C.; Stokholm, H. Oxygen release from roots of submerged aquatic macrophytes. Oikos 1982, 38, 349–354. [Google Scholar] [CrossRef]
- Yao, Y.; Liu, H.; Han, R.; Li, D.; Zhang, L. Identifying the Mechanisms behind the Positive Feedback Loop between Nitrogen Cycling and Algal Blooms in a Shallow Eutrophic Lake. Water 2021, 13, 524. [Google Scholar] [CrossRef]
- Deng, H.; Zhang, J.; Wu, J.; Yao, X.; Yang, L. Biological denitrification in a macrophytic lake: Implications for macrophytes-dominated lake management in the north of China. Environ. Sci. Pollut. Res. 2020, 27, 42460–42471. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Shi, W.; Zhou, J.; Yu, J.; Qin, B. Strong turbulence accelerates sediment nitrification-denitrification for nitrogen loss in shallow lakes. Sci. Total Environ. 2020, 761, 143210. [Google Scholar] [CrossRef]
- Wu, H.; Hao, B.; Zhou, Q.; Xu, K.; Liu, G. Contribution of various categories of environmental factors to sediment nitrogen-removal in a low C/N ratio river. Ecol. Eng. 2021, 159, 106121. [Google Scholar] [CrossRef]
- Liu, W.; Jiang, X.; Zhang, Q.; Feng, L.; Liu, G. Has Submerged Vegetation Loss Altered Sediment Denitrification, N2O Production, and Denitrifying Microbial Communities in Subtropical Lakes? Glob. Biogeochem. Cycles. 2018, 32, 1195–1207. [Google Scholar] [CrossRef]
- Gossiaux, A.; Jabiol, J.; Poupin, P.; Chauvet, E.; Guerold, F. Seasonal variations overwhelm temperature effects on microbial processes in headwater streams: Insights from a temperate thermal spring. Aquat. Sci. 2019, 81, 30. [Google Scholar] [CrossRef] [Green Version]
- Duarte, S.; Cássio, S.; Ferreira, V.; Canhoto, C.; Pascoal, C. Seasonal Variability May Affect Microbial Decomposers and Leaf Decomposition More Than Warming in Streams. Microb. Ecol. 2016, 72, 263–276. [Google Scholar] [CrossRef]
- Brzostek, E.R.; Finzi, A.C. Seasonal variation in the temperature sensitivity of proteolytic enzyme activity in temperate forest soils. J. Geophys. Res. 2012, 117, 1018. [Google Scholar] [CrossRef] [Green Version]
- Berdjeb, L.; Parada, A.; Needham, D.M.; Fuhrman, J.A. Short-term dynamics and interactions of marine protist communities during the spring–summer transition. ISME J. 2018, 12, 1907–1917. [Google Scholar] [CrossRef] [PubMed]
- Lindström, E.S.; Feng, X.M.; Graneli, W.; Kritzbergb, E.S. The interplay between bacterial community composition and the environment determining function of inland water bacteria. Limnol. Oceanogr. 2010, 55, 2052–2060. [Google Scholar] [CrossRef]
- Barbosa da Costa, N.; Fugère, V.; Hébert, M.P.; Xu, C.C.Y.; Barrett, R.D.H.; Beisner, B.E.; Bell, G.; Yargeau, V.; Fussmann, G.F.; Gonzalez, A.; et al. Resistance, resilience, and functional redundancy of freshwater bacterioplankton communities facing a gradient of agricultural stressors in a mesocosm experiment. Mol. Ecol. 2021, 30, 4771–4788. [Google Scholar] [CrossRef]
- Peura, S.; Eiler, A.; Bertilsson, S.; Nykänen, H.; Tiirola, M.; Jones, R. Distinct and diverse anaerobic bacterial communities in boreal lakes dominated by candidate division OD1. ISME J. 2012, 6, 1640–1652. [Google Scholar] [CrossRef]
- Smith, T.P.; Clegg, T.; Bell, T.; Pawar, S. Systematic variation in the temperature dependence of bacterial carbon use efficiency. Ecol. Lett. 2021, 24, 2123–2133. [Google Scholar] [CrossRef] [PubMed]
- Sinsabaugh, R.L.; Follstad Shah, J.J. Integrating resource utilization and temperature in metabolic scaling of riverine bacterial production. Ecology 2010, 91, 1455–1465. [Google Scholar] [CrossRef] [PubMed]
- Degerman, R.; Dinasquet, J.; Riemann, L.; de Luna, S.S.; Andersson, A. Effect of resource availability on bacterial community responses to increased temperature. Aquat. Microb. Ecol. 2013, 68, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogee, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef]
- Jentsch, A.; Kreyling, J.; Elmer, M.; Gellesch, E.; Glaser, B.; Grant, K.; Hein, R.; Lara, M.; Mirzae, H.; Nadler, S.E. Climate extremes initiate ecosystem-regulating functions while maintaining productivity. J. Ecol. 2011, 99, 689–702. [Google Scholar] [CrossRef]
- Correa-Araneda, F.; Tonin, A.M.; Pérez, J.; Álvarez, K.; López-Rojo, N.; Díaz, A.; Esse, C.; Encina-Montoya, F.; Figueroa, R.; Cornejo, A.; et al. Extreme climate events can slow down litter breakdown in streams. Aquat. Sci. 2020, 82, 25. [Google Scholar] [CrossRef]
- Martínez, A.; Larrañaga, A.; Pérez, J.; Descals, E.; Pozo, J. Temperature affects leaf litter decomposition in low-order forest streams: Field and microcosm approaches. FEMS Microbiol. Ecol. 2014, 87, 257–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Z.; Yu, M.; Chen, H.; Zhao, H.; Xu, J. Elevated temperature shifts soil N cycling from microbial immobilization to enhanced mineralization, nitrification and denitrification across global terrestrial ecosystems. Glob. Chang. Biol. 2020, 26, 5267–5276. [Google Scholar] [CrossRef]
- Rysgaard, S.; Risgaard-Petersen, N.; Peter, S.N.; Kim, J.; Peter, N.L. Oxygen regulation of nitrification and denitrification in sediments. Limnol. Oceanogr. 2003, 39, 1643–1652. [Google Scholar] [CrossRef]
- Zhan, Q.; Stratmann, C.N.; van der Geest, H.G.; Veraart, A.J.; Brenzinger, K.; Lürling, M.; de Senerpont Domis, L.N. Effectiveness of phosphorus control under extreme heatwaves: Implications for sediment nutrient releases and greenhouse gas emissions. Biogeochemistry 2021, 156, 421–436. [Google Scholar] [CrossRef]
- Hartmann, A.A.; Barnard, R.L.; Marhan, S.; Niklaus, P.A. Effects of drought and N-fertilization on N cycling in two grassland soils. Oecologia 2013, 171, 705–717. [Google Scholar] [CrossRef]
- Bergen, T.J.H.M.; Barros, N.; Mendonça, R.; Aben, R.C.H.; Althuizen, I.H.J.; Huszar, V.; Lamers, L.P.M.; Lürling, M.; Roland, F.; Kosten, S. Seasonal and diel variation in greenhouse gas emissions from an urban pond and its major drivers. Limnol. Oceanogr. 2019, 64, 2129–2139. [Google Scholar] [CrossRef]
- Peacock, M.; Audet, J.; Jordan, S.; Smeds, J.; Wallin, M.B. Greenhouse gas emissions from urban ponds are driven by nutrient status and hydrology. Ecosphere 2019, 10, e02643. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Treatment | ||
|---|---|---|---|
| C | T | V | |
| DO (mg·L−1) | 6.12 ± 0.27 a | 6.43 ± 0.25 a | 6.06 ± 0.26 a |
| Cond (μs·cm−1) | 218.82 ± 2.26 a | 266.01 ± 3.73 b | 266.83 ± 3.42 b |
| pH | 8.43 ± 0.08 a | 8.59 ± 0.07 a | 8.53 ± 0.07 a |
| Turb (mg·L−1) | 2.41 ± 0.11 a | 3.91 ± 0.23 b | 2.54 ± 0.10 a |
| PO43−-P (μg·L−1) | 11.71 ± 0.90 a | 13.63 ± 0.91 a | 10.20 ± 0.77 a |
| NH4 +-N (mg·L−1) | 0.11 ± 0.01 a | 0.17 ± 0.01 b | 0.15 ± 0.01 b |
| NO3−-N (mg·L−1) | 0.09 ± 0.01 a | 0.11 ± 0.01 b | 0.09 ± 0.00 a |
| Chl.a (μg·L−1) | 8.30 ± 0.67 a | 23.64 ± 1.92 b | 20.49 ± 1.81 b |
| TP (mg·L−1) | 0.06 ± 0.00 a | 0.06 ± 0.00 a | 0.06 ± 0.00 a |
| TN (mg·L−1) | 0.62 ± 0.03 a | 0.87 ± 0.03 b | 0.78 ± 0.03 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, P.; Wang, H.; Feng, M.; Cheng, H.; Yang, Q.; Yan, Y.; Xu, J.; Zhang, M. The Coupling Response between Different Bacterial Metabolic Functions in Water and Sediment Improve the Ability to Mitigate Climate Change. Water 2022, 14, 1203. https://doi.org/10.3390/w14081203
Shi P, Wang H, Feng M, Cheng H, Yang Q, Yan Y, Xu J, Zhang M. The Coupling Response between Different Bacterial Metabolic Functions in Water and Sediment Improve the Ability to Mitigate Climate Change. Water. 2022; 14(8):1203. https://doi.org/10.3390/w14081203
Chicago/Turabian StyleShi, Penglan, Huan Wang, Mingjun Feng, Haowu Cheng, Qian Yang, Yifeng Yan, Jun Xu, and Min Zhang. 2022. "The Coupling Response between Different Bacterial Metabolic Functions in Water and Sediment Improve the Ability to Mitigate Climate Change" Water 14, no. 8: 1203. https://doi.org/10.3390/w14081203