
Hydroclimatic Conditions, Wildfire, and Species Assemblages Influence Co-Occurrence of Bull Trout and Tailed Frogs in Northern Rocky Mountain Streams



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Electrofishing Detection of Tailed Frogs
2.3. Importance of GIS-Derived versus Field-Measured Predictor Variables
2.4. How Often Do Bull Trout and Tailed Frogs Co-Occur?
2.5. What Are the Characteristics of Habitats Occupied by Each Species?
2.6. What Habitat Characteristics Are Associated with Co-Occupancy?
2.7. What Are the Wildfire Trends across Occupancy and Co-Occupancy Categories?
3. Results
3.1. Was Electrofishing Effective for Detecting Tailed Frogs?
3.2. What Was the Relative Importance of Field-Measured versus GIS-Derived Predictor?
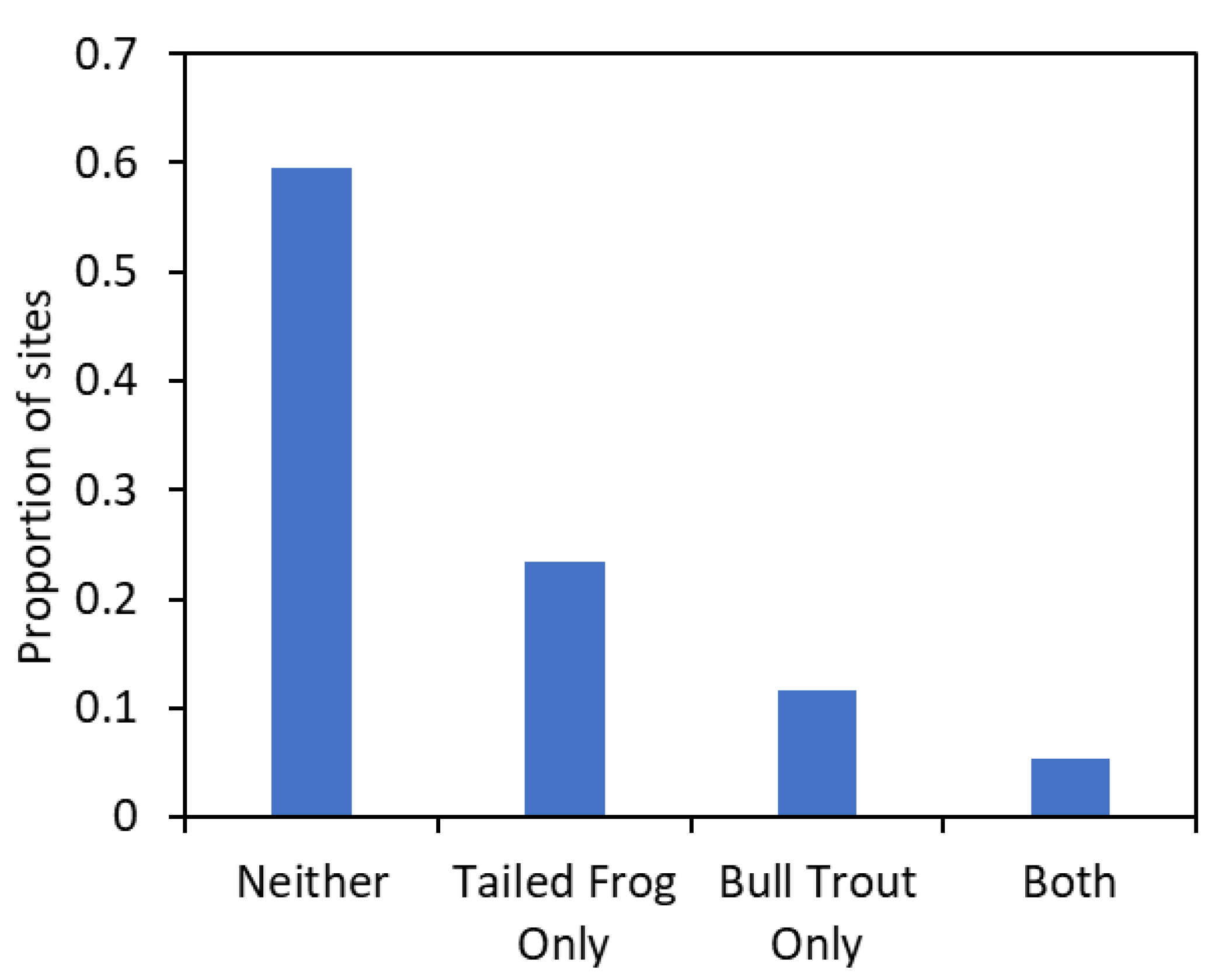
3.3. How Often Do Bull Trout and Tailed Frogs Co-Occur?
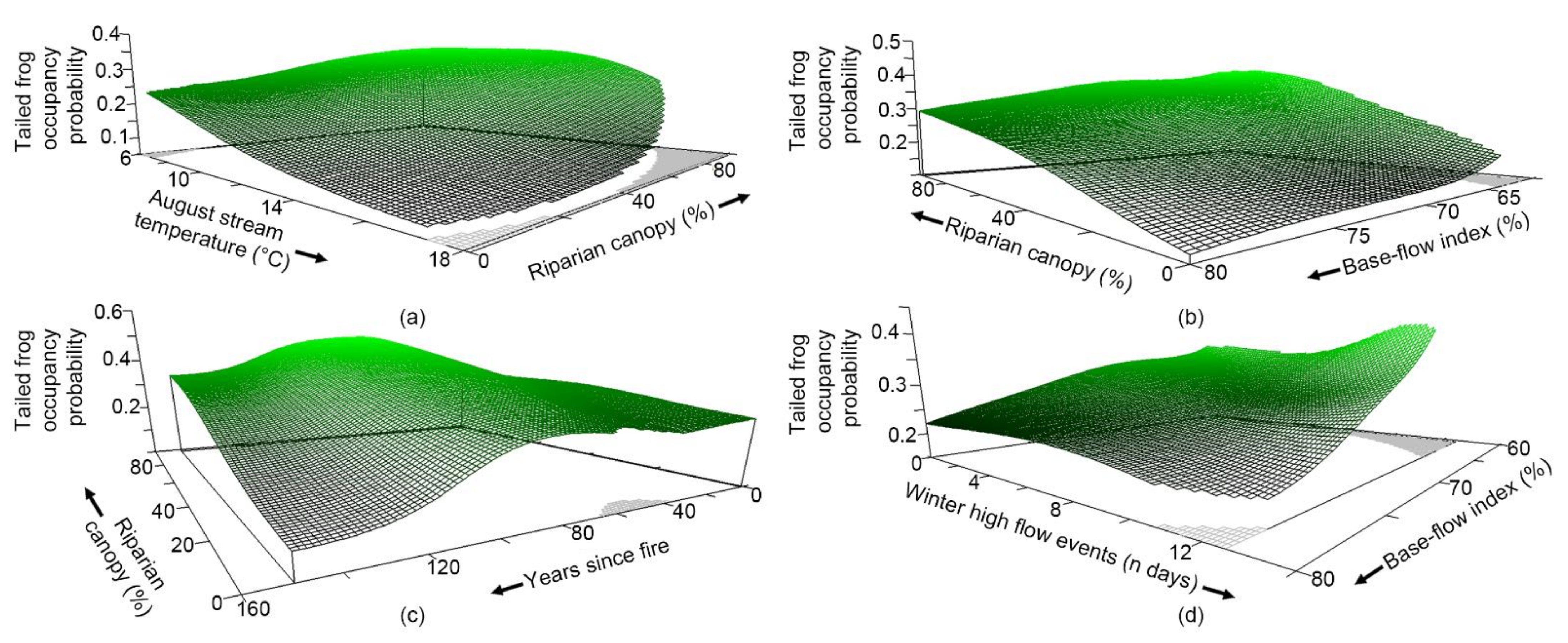
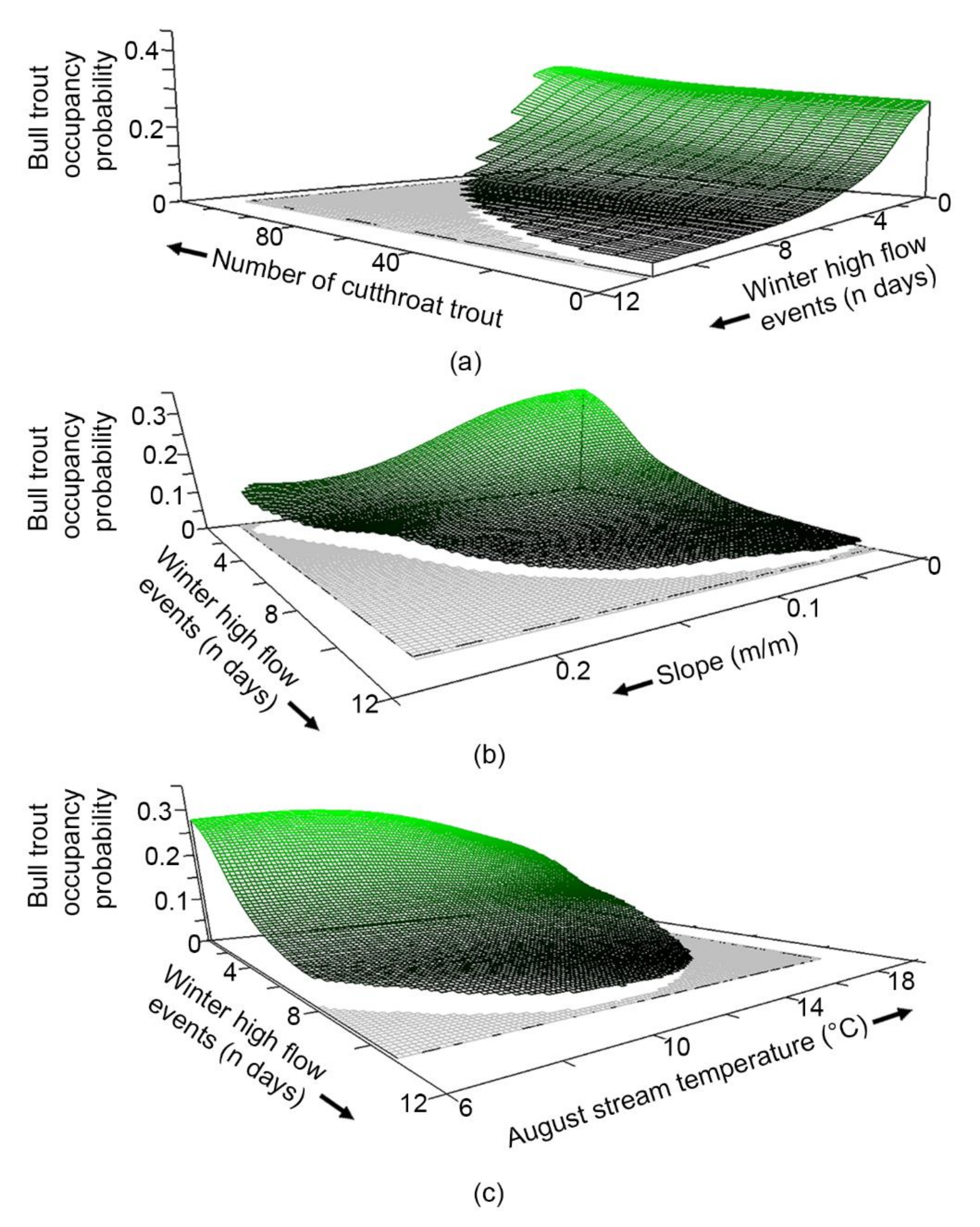
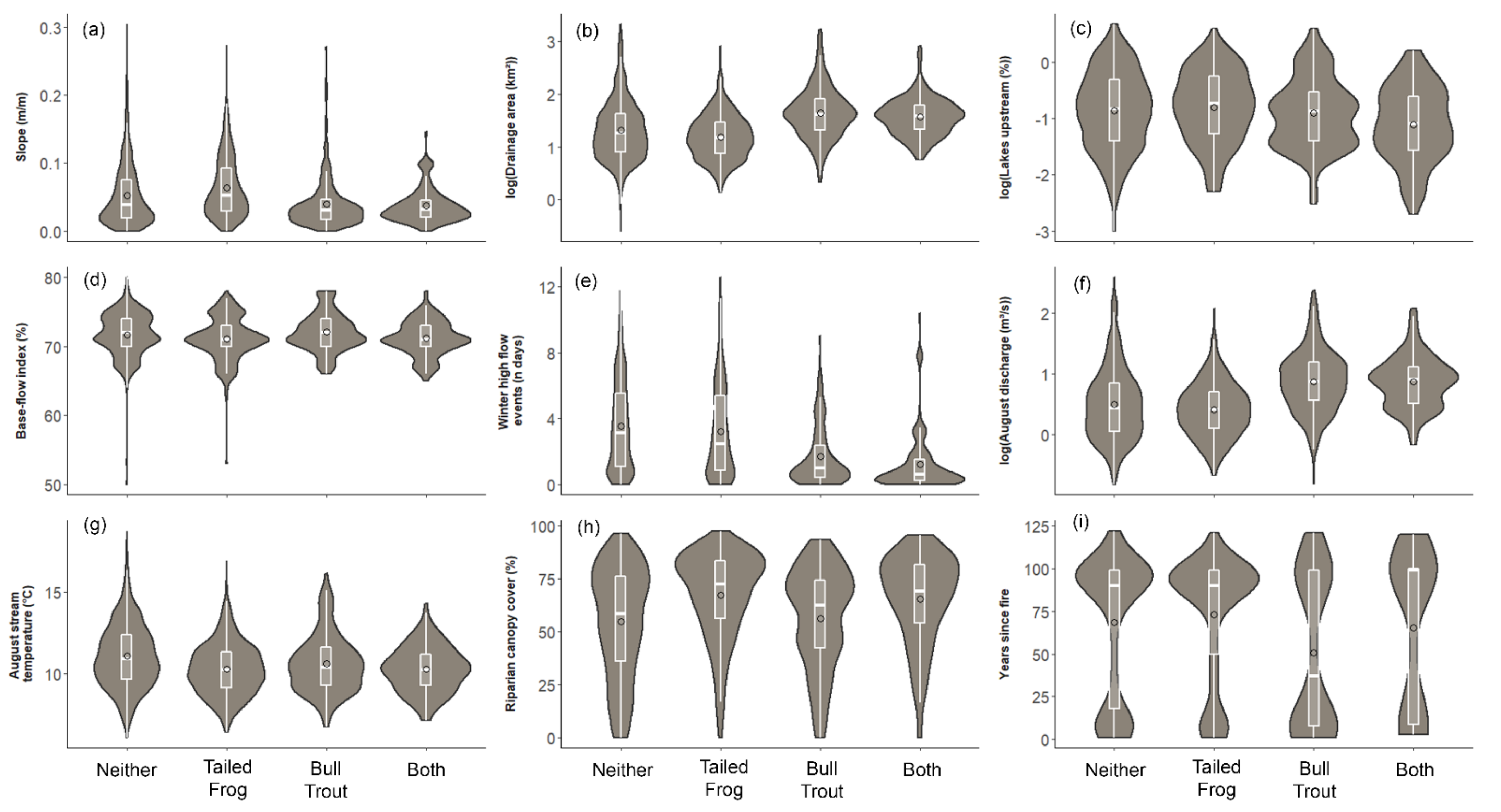
3.4. What Are the Characteristics of Habitats Occupied by Each Species?
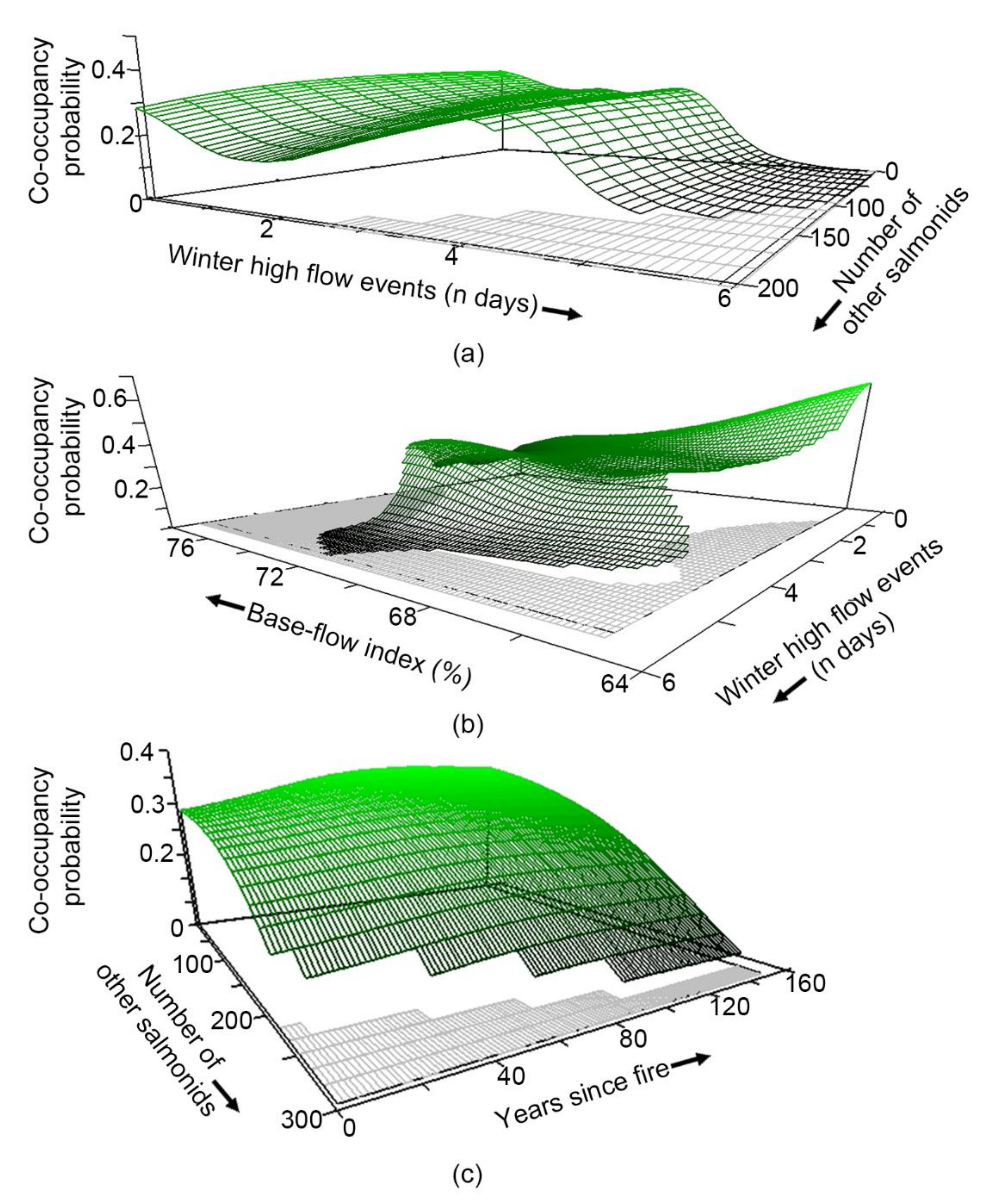
3.5. What Habitat Characteristics Are Associated with Co-Occupancy?
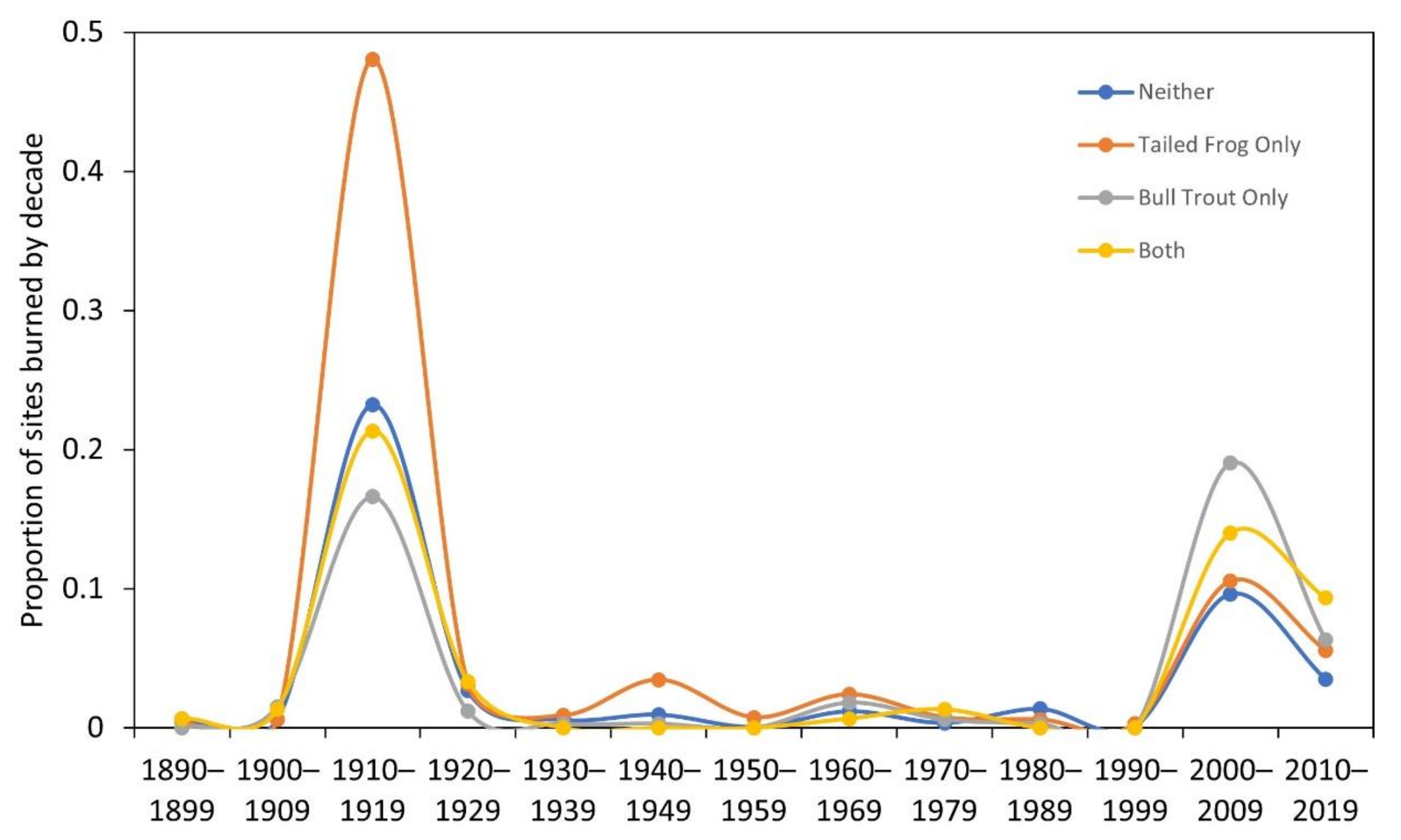
3.6. What Are the Wildfire Trends across Occupancy and Co-Occupancy Categories?
4. Discussion
4.1. Species Occupancy and Co-Occupancy Habitat Models
4.2. Methodological Comparisons
4.3. Implications for Climate Change
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2021: The Physical Science Basis; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Stewart, I.T.; Cayan, D.R.; Dettinger, M.D. Changes in snowmelt runoff timing in western North America under a business as usual climate change scenario. Clim. Chang. 2004, 62, 217–232. [Google Scholar] [CrossRef]
- Mote, P.W.; Li, S.; Lettenmaier, D.P.; Xiao, M.; Engel, R. Dramatic declines in snowpack in the western US. Clim. Atmos. Sci. 2018, 1, 2. [Google Scholar] [CrossRef]
- Hamlet, A.F.; Lettenmaier, D.P. Effects of 20th century warming and climate variability on flood risk in the western U.S. Water Resour. Res. 2007, 43, W06427. [Google Scholar] [CrossRef]
- Luce, C.H.; Holden, Z.A. Declining annual streamflow distributions in the Pacific Northwest United States, 1948–2006. Geophys. Res. Lett. 2009, 36, L16401. [Google Scholar] [CrossRef] [Green Version]
- Leppi, J.C.; DeLuca, T.H.; Harrar, S.W.; Running, S.W. Impacts of climate change on August stream discharge in the Central-Rocky Mountains. Clim. Chang. 2012, 112, 997–1014. [Google Scholar] [CrossRef]
- Isaak, D.; Wollrab, S.; Horan, D.; Chandler, G. Climate change effects on stream and river temperatures across the Northwest U.S. from 1980—2009 and implications for salmonid fishes. Clim. Chang. 2012, 113, 499–524. [Google Scholar] [CrossRef] [Green Version]
- Isaak, D.J.; Luce, C.H.; Horan, D.L.; Chandler, G.; Wollrab, S.; Nagel, D. Global warming of salmon and trout rivers in the northwestern U.S.: Road to ruin or path through purgatory? Trans. Am. Fish. Soc. 2018, 147, 566–587. [Google Scholar] [CrossRef]
- Comte, L.; Buisson, L.; Daufresne, M.; Grenouillet, G. Climate-induced changes in the distribution of freshwater fish: Observed and predicted trends. Freshw. Biol. 2013, 58, 625–639. [Google Scholar] [CrossRef] [Green Version]
- Kovach, R.P.; Muhlfeld, C.C.; Al-Chokhachy, R.; Dunham, J.B.; Letcher, B.H.; Kershner, J.L. Impacts of climatic variation on trout: A global synthesis and path forward. Rev. Fish Biol. Fish. 2016, 26, 135–151. [Google Scholar] [CrossRef]
- Lee, S.Y.; Fullerton, A.H.; Sun, N.; Torgersen, C.E. Projecting spatiotemporally explicit effects of climate change on stream temperature: A model comparison and implications for coldwater fishes. J. Hydrol. 2020, 588, 125066. [Google Scholar] [CrossRef]
- Rahel, F.J.; Olden, J.D. Assessing the effects of climate change on aquatic invasive species. Conserv. Biol. 2008, 22, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Young, M.K.; Isaak, D.J.; McKelvey, K.S.; Wilcox, T.M.; Pilgrim, K.L.; Carim, K.J.; Campbell, M.R.; Corsi, M.P.; Horan, D.L.; Nagel, D.E.; et al. Climate, demography, and zoogeography predict introgression thresholds in salmonid hybrid zones in Rocky Mountain streams. PLoS ONE 2016, 11, e0163563. [Google Scholar] [CrossRef] [PubMed]
- Rubenson, E.S.; Olden, J.D. An invader in salmonid rearing habitat: Current and future distributions of smallmouth bass (Micropterus dolomieu) in the Columbia River Basin. Can. J. Fish. Aquat. Sci. 2020, 77, 314–325. [Google Scholar] [CrossRef]
- Isaak, D.J.; Young, M.K.; Nagel, D.E.; Horan, D.L.; Groce, M.C. The cold-water climate shield: Delineating refugia for preserving salmonid fishes through the 21st century. Glob. Chang. Biol. 2015, 21, 2540–2553. [Google Scholar] [CrossRef] [PubMed]
- Isaak, D.J.; Young, M.K.; Luce, C.H.; Hostetler, S.; Wenger, S.; Peterson, E.E.; Ver hoef, J.M.; Groce, M.; Horan, D.L.; Nagel, D. Slow climate velocities of mountain streams portend their role as refugia for cold-water biodiversity. Proc. Natl. Acad. Sci. USA 2016, 113, 4374–4379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, S.T.; Betancourt, J.L.; Booth, R.K.; Gray, S.T. Ecology and the ratchet of events: Climate variability, niche dimensions, and species distributions. Proc. Natl. Acad. Sci. USA 2009, 106, 19685–19692. [Google Scholar] [CrossRef] [Green Version]
- Goode, J.R.; Buffington, J.M.; Tonina, D.; Isaak, D.J.; Thurow, R.F.; Wenger, S.; Nagel, D.; Luce, C.; Tetzlaff, D.; Soulsby, C. Potential effects of climate change on streambed scour and risks to salmonid survival in snow-dominated mountain basins. Hydrol. Process. 2013, 27, 750–765. [Google Scholar] [CrossRef]
- Koontz, E.D.; Steel, E.A.; Olden, J.D. Stream thermal responses to wildfire in the Pacific Northwest. Freshw. Sci. 2018, 37, 731–746. [Google Scholar] [CrossRef]
- Alizadeh, M.R.; Abatzoglou, J.T.; Luce, C.H.; Adamowski, J.F.; Farid, A.; Sadegh, M. Warming enabled upslope advance in western US forest fires. Proc. Natl. Acad. Sci. USA 2021, 118, e2009717118. [Google Scholar] [CrossRef]
- Ball, G.; Regier, P.; González-Pinzón, R.; Reale, J.; Van Horn, D. Wildfires increasingly impact western US fluvial networks. Nat. Commun. 2021, 12, 2484. [Google Scholar] [CrossRef]
- Hagmann, R.K.; Hessburg, P.F.; Prichard, S.J.; Povak, N.A.; Brown, P.M.; Fulé, P.Z.; Merschel, A.G. Evidence for widespread changes in the structure, composition, and fire regimes of western North American forests. Ecol. Appl. 2021, 31, e02431. [Google Scholar] [CrossRef] [PubMed]
- Gresswell, R.E. Fire and aquatic ecosystems in forested biomes of North America. Trans. Am. Fish. Soc. 1999, 128, 193–221. [Google Scholar] [CrossRef]
- Dunham, J.B.; Young, M.K.; Gresswell, R.E.; Rieman, B.E. Effects of fire on fish populations: Landscape perspectives on persistence of native fishes and nonnative fish invasions. For. Ecol. Manag. 2003, 178, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Arkle, R.S.; Pilliod, D.S. Prescribed fires as ecological surrogates for wildfires: A stream and riparian perspective. For. Ecol. Manag. 2010, 259, 893–903. [Google Scholar] [CrossRef]
- Mahlum, S.K.; Eby, L.A.; Young, M.K.; Clancy, C.G.; Jakober, M. Effects of wildfire on stream temperatures in the Bitterroot River Basin, Montana. Int. J. Wildland Fire 2011, 20, 240–247. [Google Scholar] [CrossRef]
- Dunham, J.B.; Rosenberger, A.E.; Luce, C.H.; Rieman, B.E. Influences of wildfire and channel reorganization on spatial and temporal variation in stream temperature and the distribution of fish and amphibians. Ecosystems 2007, 10, 335–346. [Google Scholar] [CrossRef]
- Johnson, Z.C.; Johnson, B.G.; Briggs, M.A.; Devine, W.D.; Snyder, C.D.; Hitt, N.P.; Hare, D.K.; Minkova, T.V. Paired air-water annual temperature patterns reveal hydrogeological controls on stream thermal regimes at watershed to continental scales. J. Hydrol. 2020, 587, 124929. [Google Scholar] [CrossRef]
- Arkle, R.S.; Pilliod, D.S.; Strickler, K. Fire, flow and dynamic equilibrium in stream macroinvertebrate communities. Freshw. Biol. 2010, 55, 299–314. [Google Scholar] [CrossRef]
- Luce, C.; Morgan, P.; Dwire, K.; Isaak, D.; Holden, Z.; Rieman, B. Climate Change, Forests, Fire, Water, and Fish: Building Resilient Landscapes, Streams, and Managers; Joint Fire Sciences Program, GTR-RMRS-290; U.S. Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2012. [CrossRef] [Green Version]
- Rieman, B.E.; McIntyre, J.D. Demographic and Habitat Requirements for Conservation of Bull Trout; USDA Forest Service General Technical Report INT-302; Intermountain Research Station: Ogden, UT, USA, 1993. [CrossRef]
- Shellberg, J.G.; Bolton, S.B.; Montgomery, D.R. Hydrogeomorphic effects on bedload scour in bull char (Salvalinus confluentus) spawning habitat, western Washington, USA. Can. J. Fish. Aquat. Sci. 2010, 67, 626–640. [Google Scholar] [CrossRef] [Green Version]
- USFWS (U.S. Fish and Wildlife Service). Endangered and threatened wildlife and plants: Determination of threatened status of bull trout in the coterminous. US Fed. Regist. 1999, 64, 58910–58933. [Google Scholar]
- Rieman, B.; Isaak, D.; Adams, S.; Horan, D.; Nagel, D.; Luce, C.; Myers, D. Anticipated climate warming effects on bull trout habitats and populations across the Interior Columbia River Basin. Trans. Am. Fish. Soc. 2007, 136, 1552–1565. [Google Scholar] [CrossRef]
- Eby, L.A.; Helmy, O.; Holsinger, L.M.; Young, M.K. Evidence of climate-induced range contractions for bull trout in a Rocky Mountain watershed, U.S.A. PLoS ONE 2014, 9, e98812. [Google Scholar] [CrossRef] [PubMed]
- Al-Chokhachy, R.; Schmetterling, D.; Clancy, C.; Saffel, P.; Kovach, R.; Nyce, L.; Liermann, B.; Fredenberg, W.; Pierce, R. Are brown trout replacing or displacing bull trout populations in a changing climate? Can. J. Fish. Aquat. Sci. 2016, 73, 1395–1404. [Google Scholar] [CrossRef]
- LeMoine, M.; Clancy, C.; Nyce, L.; Jakober, M.; Eby, L.A.; Isaak, D. Landscape resistance mediates native fish species distribution shifts and vulnerability to climate change in riverscapes. Glob. Chang. Biol. 2020, 26, 5492–5508. [Google Scholar] [CrossRef]
- Bell, D.A.; Kovach, R.P.; Muhlfeld, C.C.; Al-Chokhachy, R.; Cline, T.J.; Whited, D.C.; Schmetterling, D.A.; Lukacs, P.M.; Whiteley, A.R. Climate change and expanding invasive species drive widespread declines of native trout in the northern Rocky Mountains, USA. Sci. Adv. 2021, 7, eabj5471. [Google Scholar] [CrossRef]
- Isaak, D.J.; Luce, C.H.; Rieman, B.E.; Nagel, D.E.; Peterson, E.E.; Horan, D.L.; Parkes, S.; Chandler, G.L. Effects of climate change and wildfire on stream temperatures and salmonid thermal habitat in a mountain river network. Ecol. Appl. 2010, 20, 1350–1371. [Google Scholar] [CrossRef] [Green Version]
- Green, D.M.; Cannatella, D.C. Phylogenetic significance of the amphicoelous frogs, Ascaphidae and Leiopelmatidae. Ethol. Ecol. Evol. 1993, 5, 233–245. [Google Scholar] [CrossRef]
- Metzger, G.; Espindola, A.; Waits, L.P.; Sullivan, J. Genetic structure across broad spatial and temporal scales: Rocky Mountain tailed frogs (Ascaphus montanus; Anura: Ascaphidae) in the inland temperate rainforest. J. Hered. 2015, 106, 700–710. [Google Scholar] [CrossRef] [Green Version]
- Isaak, D.J.; Wenger, S.J.; Young, M.K. Big biology meets microclimatology: Defining thermal niches of ectotherms at landscape scales for conservation planning. Ecol. Appl. 2017, 27, 977–990. [Google Scholar] [CrossRef]
- Adams, S.B.; Frissell, C.A. Thermal Habitat Use and Evidence of Seasonal Migration by Rocky Mountain Tailed Frogs, Ascaphus montanus, in Montana. Can. Field-Nat. 2001, 115, 251–256. [Google Scholar]
- Honeycutt, R.K.; Garwood, J.M.; Lowe, W.H.; Hossack, B.R. Spatial capture–recapture reveals age-and sex-specific survival and movement in stream amphibians. Oecologia 2019, 190, 821–833. [Google Scholar] [CrossRef] [PubMed]
- Hossack, B.R.; Lowe, W.H.; Webb, M.A.; Talbott, M.J.; Kappenman, K.M.; Corn, P.S. Population-level thermal performance of a cold-water ectotherm is linked to ontogeny and local environmental heterogeneity. Freshw. Biol. 2013, 58, 2215–2225. [Google Scholar] [CrossRef]
- Hossack, B.R.; Pilliod, D.S. Amphibian responses to wildfire in the western United States: Emerging patterns from short-term studies. Fire Ecol. 2011, 7, 129–144. [Google Scholar] [CrossRef]
- Peterson, J.T.; Thurow, R.F.; Guzevich, J.W. An evaluation of multipass electrofishing for estimating the abundance of stream-dwelling salmonids. Trans. Am. Fish. Soc. 2004, 133, 462–475. [Google Scholar] [CrossRef]
- Thurow, R.F.; Peterson, J.T.; Guzevich, J.W. Utility and validation of day and night snorkel counts for estimating bull trout abundance in first-to third-order streams. N. Am. J. Fish. Manag. 2006, 26, 217–232. [Google Scholar] [CrossRef]
- Peterson, J.T.; Banish, N.P. The evaluation of sampling conditions across the bull trout range in Washington State; Cooperative Monitoring, Evaluation and Research Report, CMER 01-105. Final Report to the U.S. Fish and Wildlife Service; Aquatic Resources Division: Lacey, WA, USA.
- Cossel, J.O., Jr.; Gaige, M.G.; Sauder, J.D. Electroshocking as a survey technique for stream-dwelling amphibians. Wildl. Soc. Bull. 2012, 36, 358–364. [Google Scholar] [CrossRef]
- Isaak, D.J.; Wenger, S.J.; Peterson, E.E.; Ver Hoef, J.M.; Nagel, D.E.; Luce, C.H.; Hostetler, S.W.; Dunham, J.B.; Roper, B.B.; Wollrab, S.P.; et al. The NorWeST summer stream temperature model and scenarios for the western US: A crowd-sourced database and new geospatial tools foster a user community and predict broad climate warming of rivers and streams. Water Resour. Res. 2017, 53, 9181–9205. [Google Scholar] [CrossRef] [Green Version]
- McCune, B.; Mefford, M.J. HyperNiche: Multiplicative Habitat Modeling; Version 2.30; MjM Software: Glenden Beach, OR, USA, 2009. [Google Scholar]
- McKay, L.; Bondelid, T.; Dewald, T.; Johnston, J.; Moore, R.; Reah, A. NHDPlus Version 2, 2012: User Guide. Available online: https://nhdplus.com/NHDPlus/NHDPlusV2_documentation.php (accessed on 15 May 2021).
- Welty, J.L.; Jeffries, M.I. Combined Wildfire Datasets for the United States and Certain Territories, 1878–2019; U.S. Geological Survey Data: Washington, DC, USA, 2020. [CrossRef]
- Feminella, J.W.; Hawkins, C.P. Tailed frog tadpoles differentially alter their feeding behavior in response to non-visual cues from four predators. J. N. Am. Benthol. Soc. 1994, 13, 310–320. [Google Scholar] [CrossRef]
- Friele, P.A.; Paige, K.; Moore, R.D. Stream temperature regimes and the distribution of the Rocky Mountain tailed frog at its northern range limit, southeastern British Columbia. Northw. Sci. 2016, 90, 159–175. [Google Scholar] [CrossRef]
- Jakober, M.J.; McMahon, T.E.; Thurow, R.F. Diel habitat partitioning by bull charr and cutthroat trout during fall and winter in Rocky Mountain streams. Environ. Biol. Fish. 2000, 59, 79–89. [Google Scholar] [CrossRef]
- Selong, J.H.; McMahon, T.E.; Zale, A.V.; Barrows, F.T. Effect of temperature on growth and survival of bull trout, with application of an improved method for determining thermal tolerance in fishes. Trans. Am. Fish. Soc. 2001, 130, 1026–1037. [Google Scholar] [CrossRef]
- Thurow, R.F. Habitat utilization and diel behavior of juvenile bull trout (Salvelinus confluentus) at the onset of winter. Ecol. Freshw. Fish 1997, 6, 1–7. [Google Scholar] [CrossRef]
- Karraker, N.E.; Pilliod, D.S.; Adams, M.J.; Bull, E.L.; Corn, P.S.; Diller, L.V.; Dupuis, L.A.; Hayes, M.P.; Hossack, B.R.; Hodgson, G.R.; et al. Taxonomic variation in oviposition by tailed frogs (Ascaphus spp.). Northw. Nat. 2006, 87, 87–97. [Google Scholar] [CrossRef]
- Hossack, B.R.; Corn, P.S.; Fagre, D.B. Divergent patterns of abundance and age-class structure of headwater stream tadpoles in burned and unburned watersheds. Can. J. Zool. 2006, 84, 1482–1488. [Google Scholar] [CrossRef] [Green Version]
- McEwan, A.L.; Johnson, C.J.; Todd, M.; Govindarajulu, P. Resource selection and movement of the coastal tailed frog in response to forest harvesting. For. Ecol. Manag. 2021, 497, 119448. [Google Scholar] [CrossRef]
- Spear, S.F.; Storfer, A. Anthropogenic and natural disturbance lead to differing patterns of gene flow in the Rocky Mountain tailed frog, Ascaphus montanus. Biol. Conserv. 2010, 143, 778–786. [Google Scholar] [CrossRef]
- Hossack, B.R.; Honeycutt, R.K. Declines revisited: Long-term recovery and spatial population dynamics of tailed frog larvae after wildfire. Biol. Conserv. 2017, 212, 274–278. [Google Scholar] [CrossRef]
- Rich, C.F., Jr.; McMahon, T.E.; Rieman, B.E.; Thompson, W.L. Local-habitat, watershed, and biotic features associated with bull trout occurrence in Montana streams. Trans. Am. Fish. Soc. 2003, 132, 1053–1064. [Google Scholar] [CrossRef] [Green Version]
- Wenger, S.J.; Isaak, D.J.; Luce, C.H.; Neville, H.M.; Fausch, K.D.; Dunham, J.B.; Dauwalter, D.C.; Young, M.K.; Elsner, M.M.; Rieman, B.E.; et al. Flow regime, temperature, and biotic interactions drive differential declines of trout species under climate change. Proc. Natl. Acad. Sci. USA 2011, 108, 14175–14180. [Google Scholar] [CrossRef] [Green Version]
- Kovach, R.P.; Al-Chokhachy, R.; Whited, D.C.; Schmetterling, D.A.; Dux, A.M.; Muhlfeld, C.C. Climate, invasive species and land use drive population dynamics of a cold-water specialist. J. Appl. Ecol. 2017, 54, 638–647. [Google Scholar] [CrossRef] [Green Version]
- Nakano, S.; Fausch, K.D.; Tanaka, T.; Maekawa, K.; Kawanabe, H. Resource utilization by bull char and cutthroat trout in a mountain stream in Montana, USA. Jpn. J. Ichthyol. 1992, 39, 211–217. [Google Scholar] [CrossRef]
- Arkle, R.S.; Pilliod, D.S. Persistence at distributional edges: Columbia spotted frog habitat in the arid Great Basin, USA. Ecol. Evol. 2015, 5, 3704–3724. [Google Scholar] [CrossRef] [PubMed]
- Gunckel, S.L.; Hemmingsen, A.R.; Li, J.L. Effect of bull trout and brook trout interactions on foraging habitat, feeding behavior, and growth. Trans. Am. Fish. Soc. 2002, 131, 1119–1130. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Peterson, C.R. Local and landscape effects of introduced trout on amphibians in historically fishless watersheds. Ecosystems 2001, 4, 322–333. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Hossack, B.R.; Bahls, P.F.; Bull, E.L.; Corn, P.S.; Hokit, G.; Maxell, B.A.; Munger, J.C.; Wyrick, A. Non-native salmonids affect amphibian occupancy at multiple spatial scales. Divers. Distrib. 2010, 16, 959–974. [Google Scholar] [CrossRef] [Green Version]
- Metter, D.E. A morphological and ecological comparison of two populations of the tailed frog, Ascaphus truei Stejneger. Copeia 1964, 1964, 181–195. [Google Scholar] [CrossRef]
- Kirk, M.A.; Rahel, F.J. Air temperatures over-predict changes to stream fish assemblages with climate warming compared with water temperatures. Ecol. Appl. 2021, 32, e02465. [Google Scholar] [CrossRef]
- Falke, J.A.; Flitcroft, R.L.; Dunham, J.B.; McNyset, K.M.; Hessburg, P.F.; Reeves, G.H. Climate change and vulnerability of bull trout (Salvelinus confluentus) in a fire-prone landscape. Can. J. Fish. Aquat. Sci. 2015, 72, 304–318. [Google Scholar] [CrossRef]
- Jager, H.I.; Long, J.W.; Malison, R.L.; Murphy, B.P.; Rust, A.; Silva, L.G.; Sollmann, R.; Steel, Z.L.; Bowen, M.D.; Dunham, J.B.; et al. Resilience of terrestrial and aquatic fauna to historical and future wildfire regimes in western North America. Ecol. Evol. 2021, 11, 12259–12284. [Google Scholar] [CrossRef]
- Austin, C.S.; Essington, T.E.; Quinn, T.P. Spawning and emergence phenology of bull trout Salvelinus confluentus under differing thermal regimes. J. Fish Biol. 2019, 94, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, J.B.; Fullerton, A.H.; Jordan, C.E.; Ebersole, J.L.; Bellmore, J.R.; Arismendi, I.; Penaluna, B.E.; Reeves, G.H. The importance of warm habitat to the growth regime of cold-water fishes. Nat. Clim. Chang. 2021, 11, 354–361. [Google Scholar] [CrossRef]
- Holsinger, L.; Keane, R.E.; Isaak, D.J.; Eby, L.; Young, M.K. Relative effects of climate change and wildfires on stream temperatures: A simulation modeling approach in a Rocky Mountain watershed. Clim. Chang. 2014, 124, 191–206. [Google Scholar] [CrossRef]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.B.; Pe’er, G.; Singer, A.; Bridle, J.R.; Crozier, L.G.; De Meester, L.; Godsoe, W.; et al. Improving the forecast for biodiversity under climate change. Science 2016, 353, 6304. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Type | Variable Name | Variable Description |
|---|---|---|
| Response variables | Tailed frog occupancy | Binary variable for whether tailed frog tadpoles were detected at a site |
| Bull trout occupancy | Binary variable for whether bull trout were detected at a site | |
| Co-occupancy | Binary variable for whether tailed frog tadpoles and bull trout were detected at a site, where a value of 1 indicates both species were detected and a value of 0 indicates only bull trout were detected | |
| Environmental predictor variables | HUC | 8-digit Hydrologic Unit Code (4th code HUCs) containing the survey site |
| Slope | Slope of the stream reach, calculated as the drop in elevation divided by segment length (m/m) [53] (accessed on 15 May 2021) | |
| Drainage area | Cumulative drainage area in the watershed upstream from the survey site [53] | |
| Lakes upstream | Percentage of watershed upstream composed of lake or reservoir Surfaces [53] | |
| Base-flow index | Base-flow index calculated as the ratio of base flow to total flow (%, https://water.usgs.gov/GIS/metadata/usgswrd/XML/bfi48grd.xml, accessed on 15 May 2021) | |
| Winter high flow events | Number of days during winter when flows are in the upper 95% of the flow record | |
| August discharge | Mean August stream discharge (m3/s; http://waterdata.usgs.gov/nwis/rt, accessed on 15 May 2021) | |
| August stream temperature | Mean August stream temperature (°C) for a baseline climate period (1993–2011) derived from the NorWeST model [51] | |
| Riparian canopy cover | Percent canopy derived from the NLCD 2011 USFS Tree Canopy Cartographic layer (https://www.mrlc.gov/data/nlcd-2011-usfs-tree-canopy-cover-conus, accessed on 15 May 2021) | |
| Wildfire predictor variables | Presurvey fires | The number of wildfires [54], (accessed on 7 October 2021) intersecting a portion of the survey reach’s upstream area, prior to the survey year |
| Years since fire | Years between the most recent wildfire prior to the survey and the survey year [54] | |
| Years since oldest fire | Years between the oldest recorded wildfire prior to the survey and the survey year [54] | |
| Predictor variables for biotic interactions | pSACO | In models predicting tailed frog occupancy, this binary variable indicated whether bull trout were detected at a site |
| nSACO | In models predicting tailed frog occupancy, this variable indicated the number of bull trout detected | |
| pASMO | In models predicting bull trout occupancy, this binary variable indicated whether tailed frog tadpoles were detected at a site | |
| nTrout | The cumulative number of non-bull trout salmonids detected at a site | |
| nONCL | The number of cutthroat trout individuals detected at a site | |
| nSAFO | The number of brook trout individuals detected at a site | |
| nONMY | The number of rainbow trout individuals detected at a site | |
| nSATR | The number of brown trout individuals detected at a site |
| Response | Predictor | Source | Sensitivity | Tolerance | % Tolerance |
|---|---|---|---|---|---|
| Tailed frog occupancy | August stream temperature | GIS | 0.36 | 0.67 | 15 |
| Winter high flow events | GIS | 0.25 | 1.89 | 15 | |
| Percent undercut bank | Field | 0.25 | 14.50 | 20 | |
| Bull trout occupancy (pSACO) | Field | na | na | na | |
| Bull trout occupancy | August stream temperature | GIS | 0.36 | 0.90 | 20 |
| Winter high flow events | GIS | 1.26 | 0.63 | 5 | |
| Riparian canopy cover | GIS | 0.07 | 34.68 | 45 |
| Response | Predictor | Sensitivity | Tolerance | % Tolerance |
|---|---|---|---|---|
| Tailed frog occupancy | Riparian canopy cover | 0.1967 | 19.5 | 20 |
| (n = 2829) | Base-flow index | 0.1841 | 3 | 10 |
| August stream temperature | 0.1781 | 1.91 | 15 | |
| Years since fire | 0.1443 | 22.35 | 15 | |
| Winter high flow events | 0.1126 | 3.138 | 25 | |
| Bull trout occupancy | Winter high flow events | 0.4519 | 1.26 | 10 |
| (n = 2829) | Number of cutthroat trout (nONCL) | 0.4348 | 35.45 | 5 |
| Slope | 0.2767 | 0.03 | 10 | |
| August stream temperature | 0.1062 | 2.534 | 20 | |
| Co-occupancy | Winter high flow events | 0.8312 | 0.52 | 5 |
| (n = 481) | Number of salmonids (nTrout) | 0.6196 | 77.55 | 5 |
| Base-flow index | 0.3674 | 1.95 | 15 | |
| Years since fire | 0.0851 | 52.15 | 35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pilliod, D.S.; Arkle, R.S.; Thurow, R.F.; Isaak, D.J. Hydroclimatic Conditions, Wildfire, and Species Assemblages Influence Co-Occurrence of Bull Trout and Tailed Frogs in Northern Rocky Mountain Streams. Water 2022, 14, 1162. https://doi.org/10.3390/w14071162
Pilliod DS, Arkle RS, Thurow RF, Isaak DJ. Hydroclimatic Conditions, Wildfire, and Species Assemblages Influence Co-Occurrence of Bull Trout and Tailed Frogs in Northern Rocky Mountain Streams. Water. 2022; 14(7):1162. https://doi.org/10.3390/w14071162
Chicago/Turabian StylePilliod, David S., Robert S. Arkle, Russell F. Thurow, and Daniel J. Isaak. 2022. "Hydroclimatic Conditions, Wildfire, and Species Assemblages Influence Co-Occurrence of Bull Trout and Tailed Frogs in Northern Rocky Mountain Streams" Water 14, no. 7: 1162. https://doi.org/10.3390/w14071162