
Reconstruction of Ecological Transitions in a Temperate Shallow Lake of the Middle Yangtze River Basin in the Last Century

 , , , and
, , , and
Abstract
:1. Introduction
2. Material and Methods
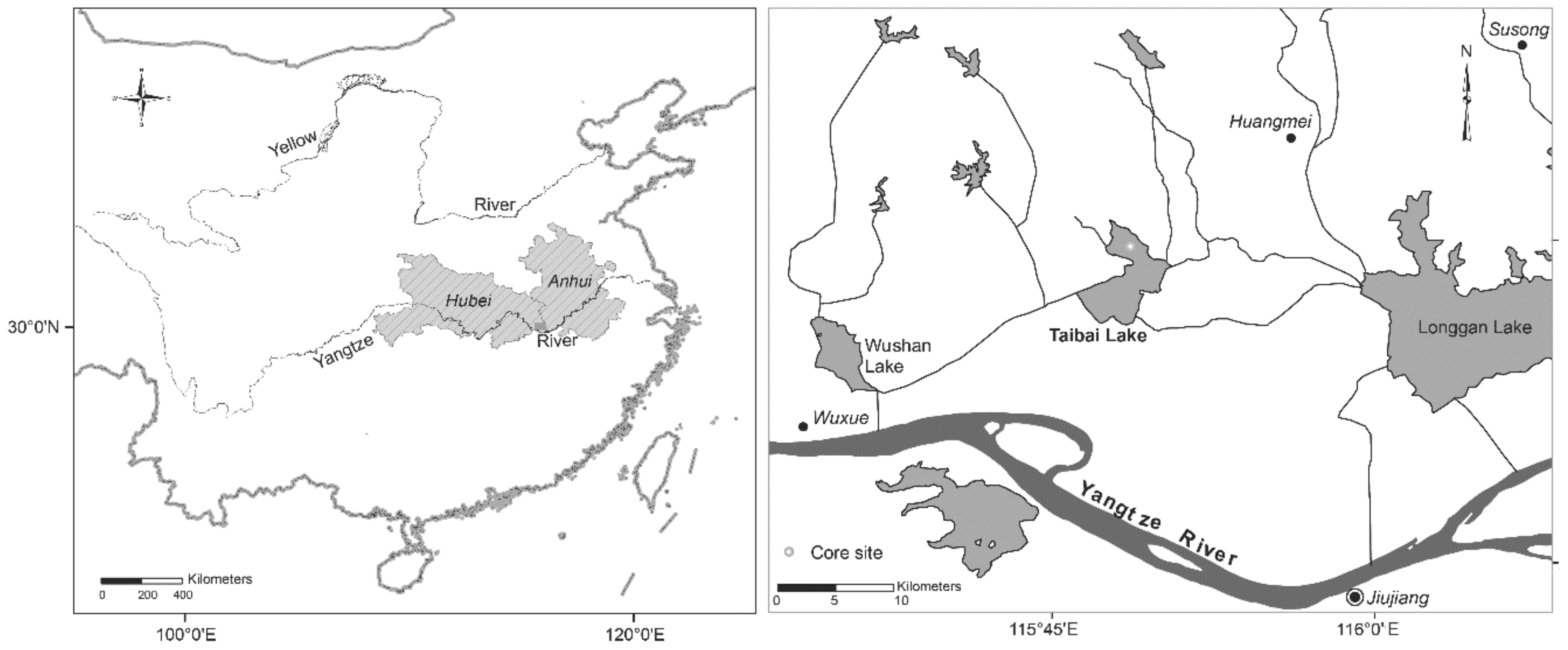
2.1. Lake and Catchment History
2.2. Paleolimnology Reconstruction
2.3. Statistical Analysis
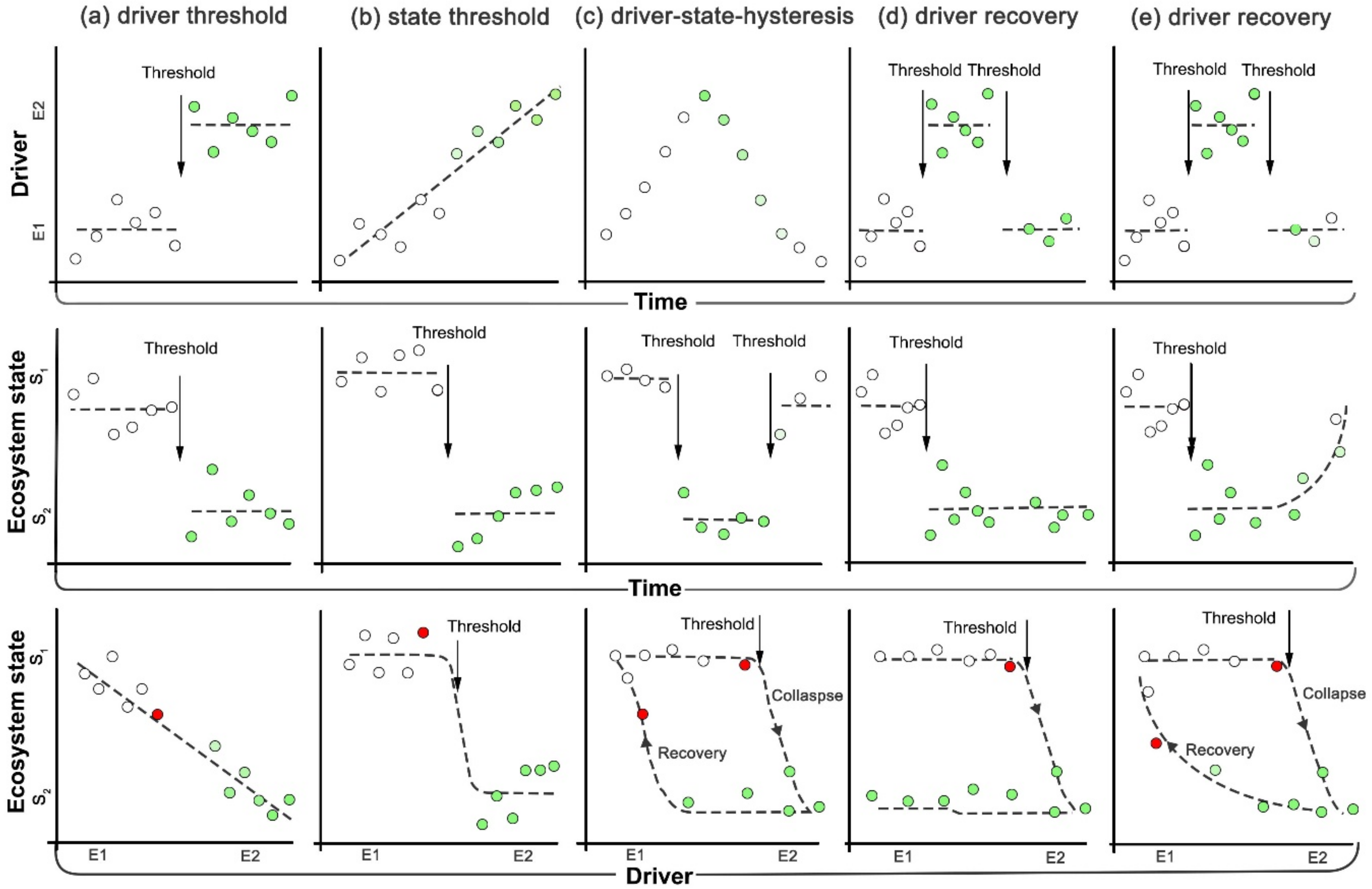
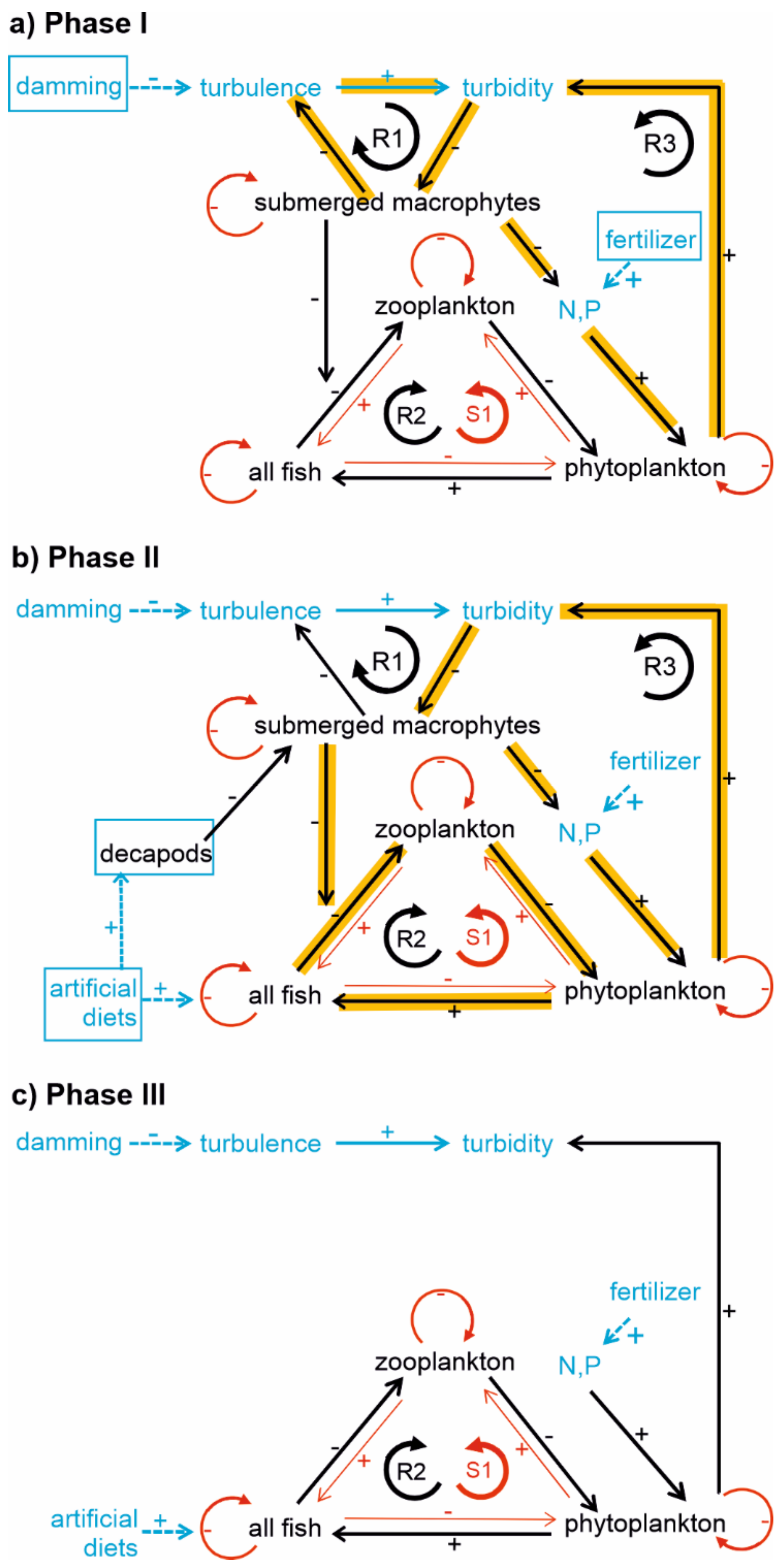
2.4. Diagrams of Interactions and Feedback Loops
3. Results
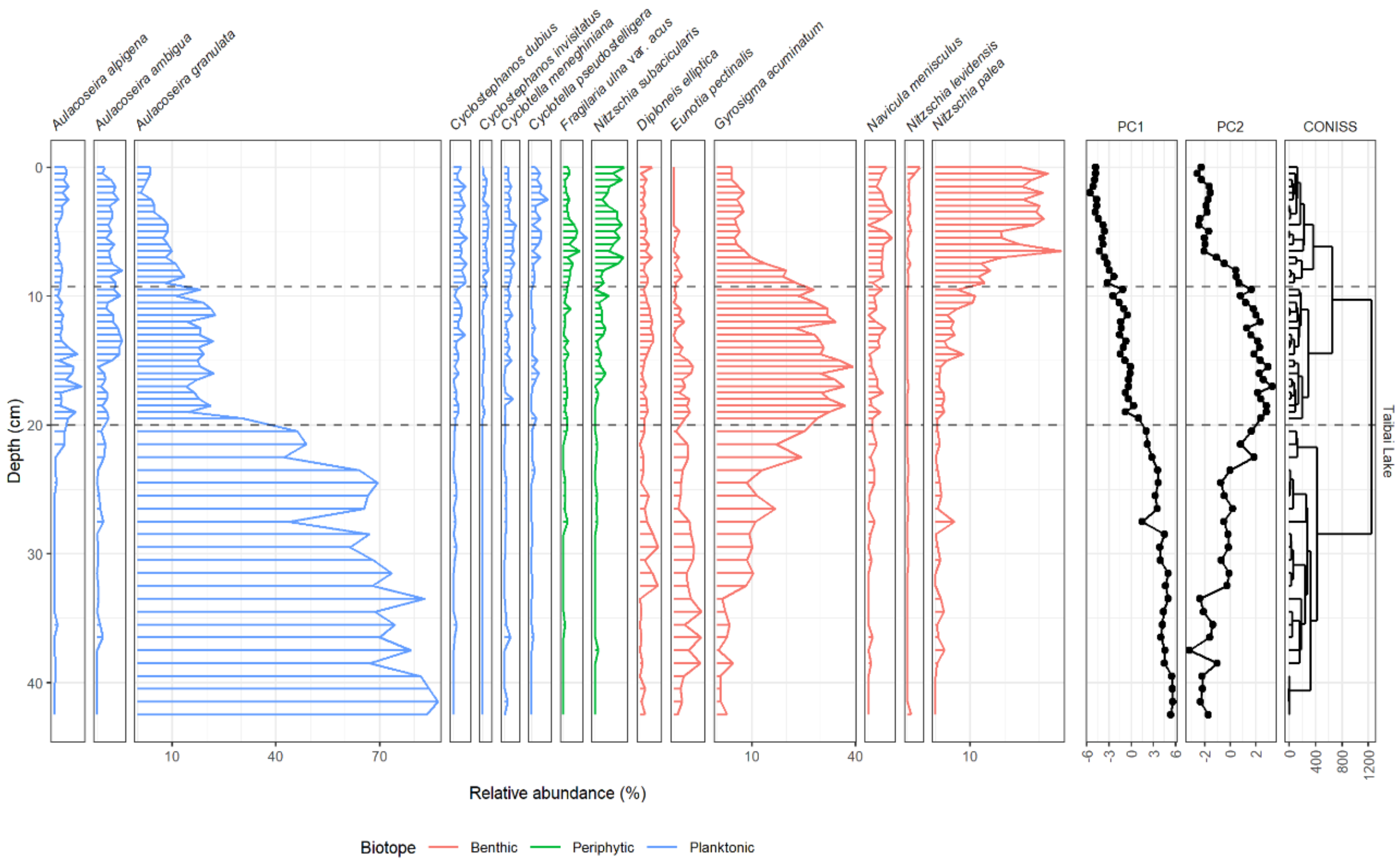
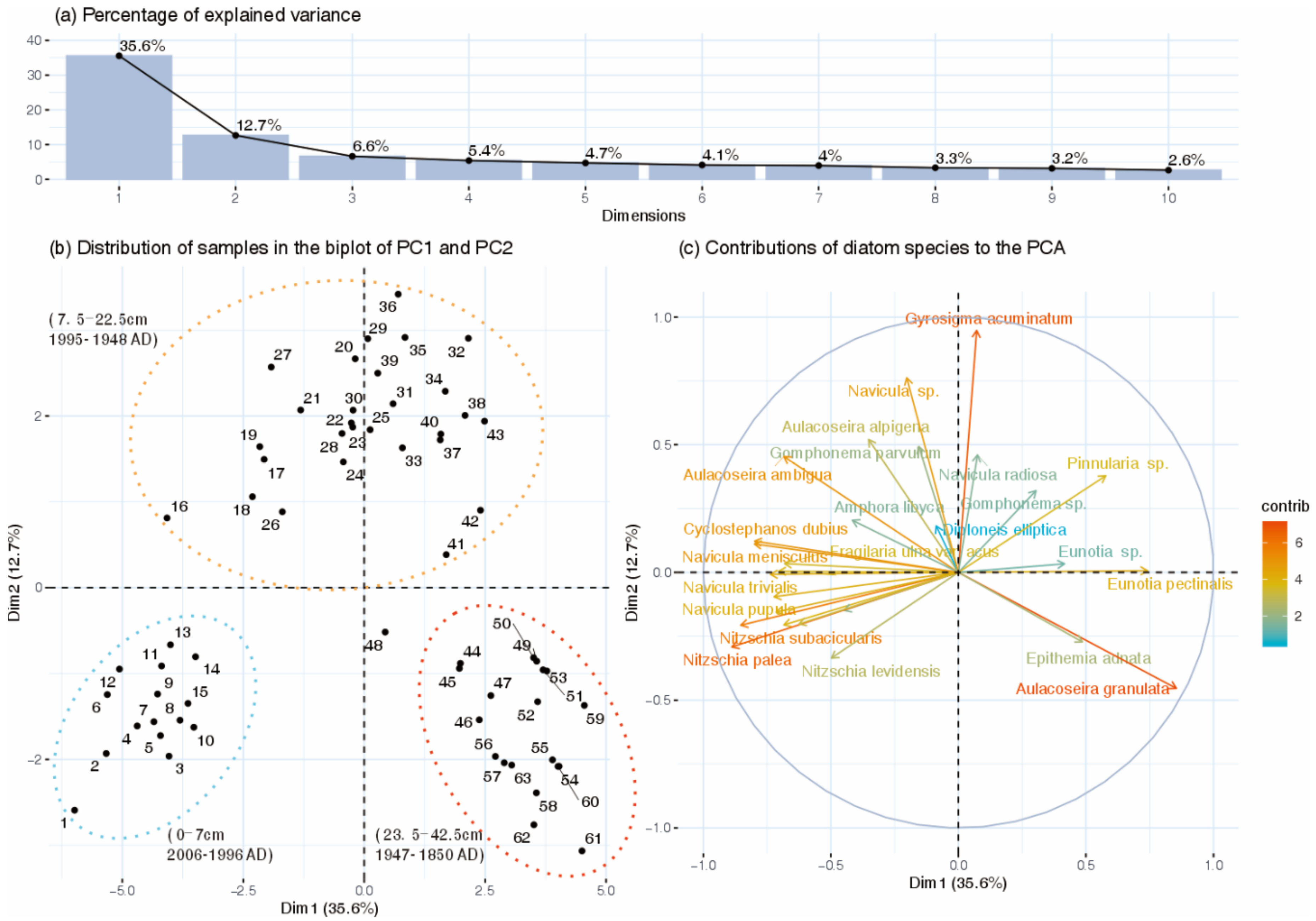
3.1. Changes in Diatom Assemblage Composition
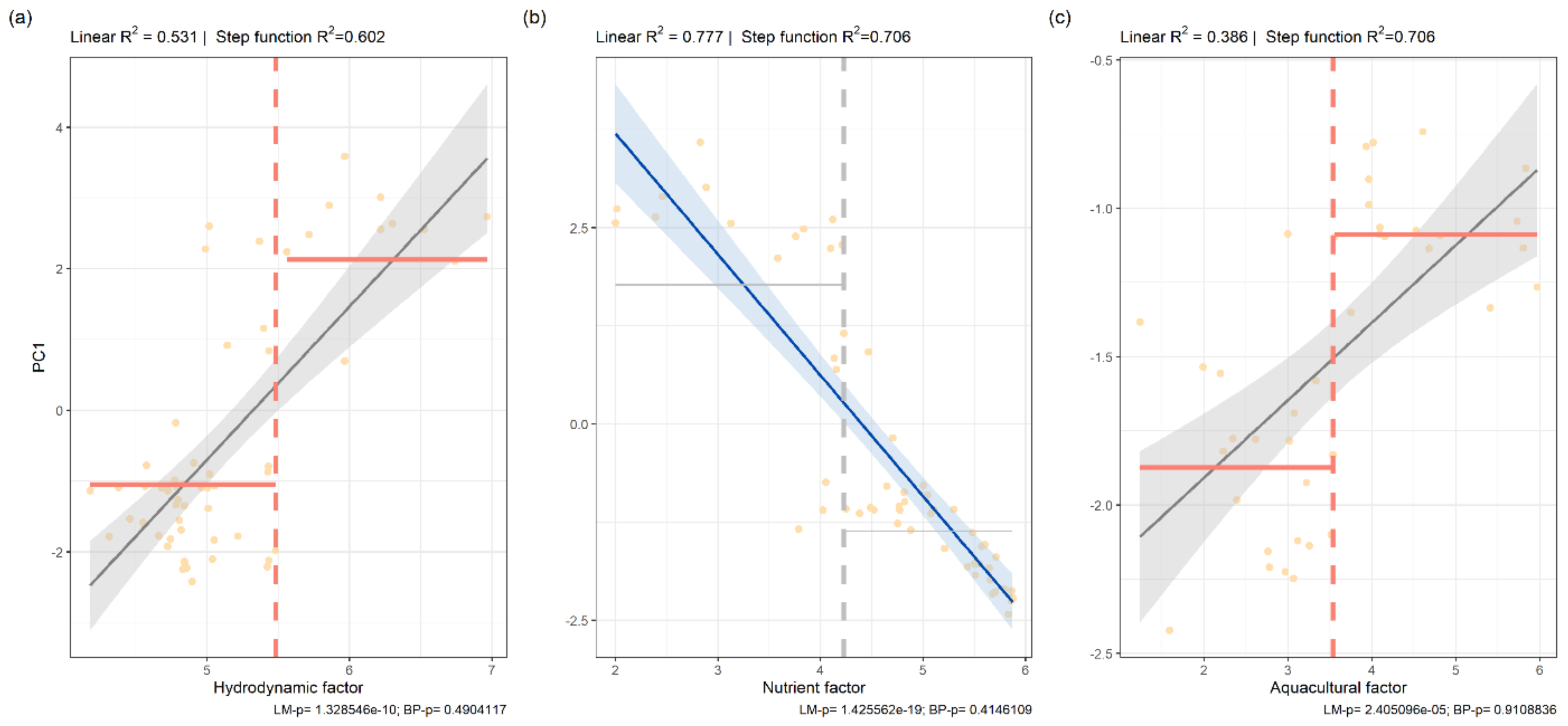
3.2. Tests for Ecological Thresholds
4. Discussion
4.1. Classification of Regime Shifts and Driver-Response Interactions
4.2. Feedback Loops Illustrating Regime Shifts
4.3. Policy Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reynaud, A.; Lanzanova, D. A Global Meta-Analysis of the Value of Ecosystem Services Provided by Lakes. Ecol. Econ. 2017, 137, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Schallenberg, M.; de Winton, M.D.; Verburg, P.; Kelly, D.J.; Hamill, K.D.; Hamilton, D.P. Ecosystem Services of Lakes. In Ecosystem Services in New Zealand—Conditions and Trends; Dymond, J.R., Ed.; Manaaki Whenua Press: Lincoln, New Zealand, 2013; pp. 203–225. [Google Scholar]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B.R. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.W. The dilemma of controlling cultural eutrophication of lakes. Proc. R. Soc. B 2012, 279, 4322–4333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheffer, M.; Hosper, S.H.; Meijer, M.-L.; Moss, B.; Jeppesen, E. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef]
- Qin, B.Q.; Gao, G.; Zhu, G.W.; Zhang, Y.L.; Song, Y.Z.; Tang, X.M.; Xu, H.; Deng, J.M. Lake eutrophication and its ecosystem response. Chin. Sci. Bull. 2013, 58, 961–970. [Google Scholar] [CrossRef] [Green Version]
- Rockström, J.; Steffen, W.; Noone, K.; Persson, Å.; Chapin, F.S., III; Lambin, E.F.; Lenton, T.M.; Scheffer, M.; Folke, C.; Schellnhuber, H.J.; et al. A safe operating space for humanity. Nature 2009, 461, 472–475. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.R. Catastrophic regime shifts in ecosystems: Linking theory to observation. Trends Ecol. Evol. 2003, 18, 648–656. [Google Scholar] [CrossRef]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime shifts, resilience, and biodiversity in ecosystem management. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; He, Q.; Yang, B.; He, W.; Xu, F.; Janssen, A.B.G.; Kuiper, J.J.; van Gerven, L.P.A.; Qin, N.; Jiang, Y.; et al. Hydrological regulation drives regime shifts: Evidence from palaeolimnology and ecosystem modeling of a large shallow Chinese lake. Glob. Chang. Biol. 2017, 23, 737–754. [Google Scholar] [CrossRef]
- Kuehn, C. A mathematical framework for critical transitions: Bifurcations, fast–slow systems and stochastic dynamics. Phys. D Nonlinear Phenom. 2011, 240, 1020–1035. [Google Scholar] [CrossRef] [Green Version]
- Veraart, A.J.; Faassen, E.J.; Dakos, V.; Van Nes, E.H.; Lürling, M.; Scheffer, M. Erratum: Corrigendum: Recovery rates reflect distance to a tipping point in a living system. Nature 2012, 484, 404. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, S.R.; Cole, J.J.; Pace, M.L.; Batt, R.; Brock, W.A.; Cline, T.; Coloso, J.; Hodgson, J.R.; Kitchell, J.F.; Seekell, D.A.; et al. Early Warnings of Regime Shifts: A Whole-Ecosystem Experiment. Science 2011, 332, 1079–1082. [Google Scholar] [CrossRef] [Green Version]
- Seekell, D.A.; Cline, T.J.; Carpenter, S.R.; Pace, M.L. Evidence of alternate attractors from a whole-ecosystem regime shift experiment. Theor. Ecol. 2013, 6, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Søndergaard, M.; Lauridsen, T.L.; Johansson, L.S.; Jeppesen, E. Repeated Fish Removal to Restore Lakes: Case Study of Lake Væng, Denmark—Two Biomanipulations during 30 Years of Monitoring. Water 2017, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- Doncaster, C.P.; Chávez, V.A.; Viguier, C.; Wang, R.; Zhang, E.; Dong, X.; Dearing, J.; Langdon, P.; Dyke, J.G. Early warning of critical transitions in biodiversity from compositional disorder. Ecology 2016, 97, 3079–3090. [Google Scholar] [CrossRef]
- Wang, R.; Dearing, J.A.; Langdon, P.G.; Zhang, E.; Yang, X.; Dakos, V.; Scheffer, M. Flickering gives early warning signals of a critical transition to a eutrophic lake state. Nature 2012, 492, 419–422. [Google Scholar] [CrossRef]
- Andersen, T.; Carstensen, J.; Hernandez-Garcia, E.; Duarte, C.M. Ecological thresholds and regime shifts: Approaches to identification. Trends Ecol. Evol. 2009, 24, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Appelt, M.; Hastrup, K.; Grønnow, B.; Mosbech, A.; Smol, J.P.; Davidson, T.A. Living in an oasis: Rapid transformations, resilience, and resistance in the North Water Area societies and ecosystems. AMBIO 2018, 47, 296–309. [Google Scholar] [CrossRef] [Green Version]
- Patten, B.C. Ecosystem Linearization: An Evolutionary Design Problem. Am. Nat. 1975, 109, 529–539. [Google Scholar] [CrossRef]
- Neutel, A.; Thorne, M. Interaction strengths in balanced carbon cycles and the absence of a relation between ecosystem complexity and stability. Ecol. Lett. 2014, 17, 651–661. [Google Scholar] [CrossRef] [Green Version]
- Kéfi, S.; Holmgren, M.; Scheffer, M. When can positive interactions cause alternative stable states in ecosystems? Funct. Ecol. 2015, 30, 88–97. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.R.; Lenton, T.M.; Bascompte, J.; Brock, W.; Dakos, V.; van de Koppel, J.; van de Leemput, I.A.; Levin, S.A.; van Nes, E.H.; et al. Anticipating Critical Transitions. Science 2012, 338, 344–348. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yang, X.; Anderson, J.; Liu, E. Diatom ecological response to altered hydrological forcing of a shallow lake on the Yangtze floodplain, SE China. Ecohydrology 2012, 268, 256–268. [Google Scholar] [CrossRef]
- Xu, M.; Dong, X.; Yang, X.; Chen, X.; Zhang, Q.; Liu, Q.; Wang, R.; Yao, M.; Davidson, T.A.; Jeppesen, E. Recent Sedimentation Rates of Shallow Lakes in the Middle and Lower Reaches of the Yangtze River: Patterns, Controlling Factors and Implications for Lake Management. Water 2017, 9, 617. [Google Scholar] [CrossRef]
- Wang, R.; Dearing, J.A.; Doncaster, C.P.; Yang, X.; Zhang, E.; Langdon, P.; Yang, H.; Dong, X.; Hu, Z.; Xu, M.; et al. Network parameters quantify loss of assemblage structure in human-impacted lake ecosystems. Glob. Chang. Biol. 2019, 25, 3871–3882. [Google Scholar] [CrossRef]
- Yang, X.; Shen, J.; Dong, X.; Liu, E.; Wang, S. Historical trophic evolutions and their ecological responses from shallow lakes in the middle and lower reaches of the Yangtze River: Case studies on Longgan Lake and Taibai Lake. Sci. China Ser. D Earth Sci. 2006, 49, 51–61. [Google Scholar] [CrossRef]
- Jian, Y. A comparative study of aquatic plant diversity of Haikou, Taibai and Wushan Lake in Hubei Province of China. Acta Ecol. Sin. 2001, 21, 1815–1824. [Google Scholar]
- Liu, E.; Yang, X.; Shen, J.; Dong, X.; Zhang, E.; Wang, S. Environmental response to climate and human impact during the last 400 years in Taibai Lake catchment, middle reach of Yangtze River, China. Sci. Total Environ. 2007, 385, 196–207. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, R.; Yang, X.; Dong, X.; Xu, M. Regime shifts revealed by paleoecological records in Lake Taibai’s ecosystem in the middle and lower Yangtze River Basin during the last century. J. Lake Sci. 2016, 28, 1381–1390. [Google Scholar] [CrossRef] [Green Version]
- Gui, J.; Tang, Q.; Li, Z.; Liu, J.; De Silva, S.S. Aquaculture in China: Success Stories and Modern Trends; Wiley-Blackwell: London, UK, 2018. [Google Scholar] [CrossRef]
- National Bureau of Statistics of China. Huanggang Statistical Yearbook-2010; China Statistics Press: Beijing, China, 2010.
- Battarbee, R.W.; Jones, V.J.; Flower, R.J.; Cameron, N.G.; Bennion, H.; Carvalho, L.; Juggins, S. Diatoms; Smol, J., Birks, H.J., Last, W., Bradley, R., Alverson, K., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 155–202. [Google Scholar]
- Smol, J.P.; Stoermer, E.F. (Eds.) The Diatoms: Applications for the Environmental and Earth Sciences, 2nd ed.; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Appleby, P. Three decades of dating recent sediments by fallout radionuclides: A review. Holocene 2008, 18, 83–93. [Google Scholar] [CrossRef]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae(1-4Teil), Süßwasserflora von Mitteleuropa; Spektrum Akademischer Verlag: Heidelberg, Germany, 1999. [Google Scholar]
- Bennett, K. Determination of the number of zones in a biostratigraphical sequence. New Phytol. 1996, 132, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Seckbach, J.; Kociolek, J.P. The Diatom World, Cellular Origin, Life in Extreme Habitats and Astrobiology; Springer: Dordrecht, The Netherlands, 2011. [Google Scholar] [CrossRef]
- Smol, J.P.; Birks, H.J.B.; Last, W.M. Tracking Environmental Change Using Lake Sediments. Volume 3, Terrestrial, Algal, and Siliceous Indicators, Developments in Palaeoenvironmental Research; Kluwer Academic: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Reid, M.A.; Ogden, R.W. Factors affecting diatom distribution in floodplain lakes of the southeast Murray Basin, Australia and implications for palaeolimnological studies. J. Paleolimnol. 2008, 41, 453–470. [Google Scholar] [CrossRef]
- Dearing, J.A. Sedimentary indicators of lake-level changes in the humid temperate zone: A critical review. J. Paleolimnol. 1997, 18, 1–14. [Google Scholar] [CrossRef]
- Chiverrell, R.; Sear, D.; Warburton, J.; Macdonald, N.; Schillereff, D.; Dearing, J.; Croudace, I.; Brown, J.; Bradley, J. Using lake sediment archives to improve understanding of flood magnitude and frequency: Recent extreme flooding in northwest UK. Earth Surf. Process. Landforms 2019, 44, 2366–2376. [Google Scholar] [CrossRef]
- Dearing, J.A.; Jones, R.T.; Shen, J.; Yang, X.; Boyle, J.F.; Foster, G.C.; Crook, D.S.; Elvin, M.J.D. Using multiple archives to understand past and present climate—human-environment interactions: The lake Erhai catchment, Yunnan Province, China. J. Paleolimnol. 2008, 40, 3–31. [Google Scholar] [CrossRef]
- Xue, J.; Li, J.; Dang, X.; Meyers, A.P.; Huang, X. Paleohydrological changes over the last 4000 years in the middle and lower reaches of the Yangtze River: Evidence from particle size and n-alkanes from Longgan Lake. Holocene 2017, 27, 1318–1324. [Google Scholar] [CrossRef]
- Ju, X.T.; Xing, G.X.; Chen, X.P.; Zhang, S.L.; Zha, L.J.; Liu, X.J.; Cui, Z.L.; Yin, B.; Christie, P.; Zhu, Z.L.; et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 8077. [Google Scholar] [CrossRef] [Green Version]
- Rodionov, S.N. A sequential algorithm for testing climate regime shifts. Geophys. Res. Lett. 2004, 31, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Zeileis, A.; Kleiber, C.; Krämer, W.; Hornik, K. Testing and dating of structural changes in practice. Comput. Stat. Data Anal. 2003, 44, 109–123. [Google Scholar] [CrossRef] [Green Version]
- Van de Leemput, I.A.; Hughes, T.P.; van Nes, E.H.; Scheffer, M. Multiple feedbacks and the prevalence of alternate stable states on coral reefs. Coral Reefs 2016, 35, 857–865. [Google Scholar] [CrossRef]
- Rooney, N.; McCann, K.S.; Gellner, G.; Moore, J.C. Structural asymmetry and the stability of diverse food webs. Nature 2006, 442, 265–269. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Lauridsen, T.; Møller, P.H.; Sandby, K. Changes in nitrogen retention in shallow eutrophic lakes following a decline in density of cyprinids. Fundam. Appl. Limnol. 1998, 142, 129–151. [Google Scholar] [CrossRef]
- Qin, B.; Zhu, G. The nutrient forms, cycling and exchange flux in the sediment and overlying water system in lakes from the middle and lower reaches of Yangtze River. Sci. China Ser. D Earth Sci. 2006, 49, 1–13. [Google Scholar] [CrossRef]
- Biggs, R.; Peterson, G.D.; Rocha, J.C. The Regime Shifts Database: A framework for analyzing regime shifts in social-ecological systems. Ecol. Soc. 2018, 23, 9. [Google Scholar] [CrossRef]
- Chen, T.; Wang, Y.; Gardner, C.; Wu, F. Threats and protection policies of the aquatic biodiversity in the Yangtze River. J. Nat. Conserv. 2020, 58, 125931. [Google Scholar] [CrossRef]
- Xu, H.; McCarthy, M.J.; Paerl, H.W.; Brookes, J.D.; Zhu, G.; Hall, N.S.; Qin, B.; Zhang, Y.; Zhu, M.; Hampel, J.J.; et al. Contributions of external nutrient loading and internal cycling to cyanobacterial bloom dynamics in Lake Taihu, China: Implications for nutrient management. Limnol. Oceanogr. 2021, 66, 1492–1509. [Google Scholar] [CrossRef]
- Anderson, N.J.; Jeppesen, E.; Søndergaard, M. Ecological effects of reduced nutrient loading (oligotrophication) on lakes: An introduction. Freshw. Biol. 2005, 50, 1589–1593. [Google Scholar] [CrossRef]
- Brucet, S.; Tavşanoğlu, Ü.N.; Özen, A.; Levi, E.E.; Bezirci, G.; Çakıroğlu, A.I.; Jeppesen, E.; Svenning, J.-C.; Ersoy, Z.; Beklioğlu, M. Size-based interactions across trophic levels in food webs of shallow Mediterranean lakes. Freshw. Biol. 2017, 62, 1819–1830. [Google Scholar] [CrossRef]
- Zou, Y. Population Time Series of Longgan-Taibai Lake Catchment during 1391–2006 and its Sediment Response. J. Chin. Hist Geogr 2011, 03, 41–59. [Google Scholar]
- National Bureau of Statistics of China. Hubei Statistical Yearbook-2013; China Statistics Press: Beijing, China, 2013.
- Cao, Y.; Zhang, E.; Langdon, P.G.; Liu, E.; Shen, J. Chironomid-inferred environmental change over the past 1400 years in the shallow, eutrophic Taibai Lake (south-east China): Separating impacts of climate and human activity. Holocene 2014, 24, 581–590. [Google Scholar] [CrossRef]
- Scheffer, M. Ecology of Shallow Lakes. In Population and Community Biology Series; Kluwer Academic Publishers: London, UK, 1998. [Google Scholar] [CrossRef]
- Jeppesen, E.; Søndergaard, M.; Søndergaard, M.; Christoffersen, K. (Eds.) The Structuring Role of Submerged Macrophytes in Lakes; Springer: New York, NY, USA, 1998. [Google Scholar] [CrossRef]
- Meiburg, E.; Kneller, B. Turbidity currents and their deposits. Annu. Rev. Fluid Mech 2010, 42, 135–156. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Periods | Human Activity and Paleolimnological Evidence |
|---|---|
| 1970s–present | 2000–present: Reducing vegetation coverage [28] |
| 2000–present: Widespread adoption of flush toilets that are unconnected with local piped sewage systems and a lack of new treatment plants have added to the increase in nutrient loading [29] | |
| 1987–present: Fish farm owners introduced economic domestic fishes into the lake, and the aquaculture intensity upgraded to a high level, resulting in exponential growth in aquaculture products [30] | |
| 1983–1993: High sedimentation flux correlated to soil loss due to intensive cultivation development [29,31] | |
| 1978–2013: In the lake catchment, the population increased and human settlements expanded, while farmland and lake area shrank (Supplementary Material 1.1) [32] | |
| 1976–present: Tongsipai Floodgate cut the dispersal route for juvenile fishes [30] | |
| The 1950s–1980s | The 1950s–1978: Rapidly developing local industrialization, mainly chemical fertilizer factories [29]. Thereafter, the lake received more nutrient loading from increasing domestic sewage, poorly treated industrial waste, and flushed chemical fertilizer [28] |
| 1958–1970: High sedimentation flux correlated with the land reclamation around Taibai [29] | |
| 1955–1962: Damming construction: three reservoirs named Jingzhu, Kaotian, and Xianrenba were built in the upper reaches of Taibai. The lake outflow passes through lake Longgan to the east of Taibai and drains to the Yangtze River [29] | |
| Previous to 1950 | The lake had periodic direct inflow connections with the Yangtze during floods [29,30] |
| Intercept ± SE | Slope ± SE | R2 | Pr(>|t|) | F-Test of Model Fit | |
|---|---|---|---|---|---|
| PC1~MD | 11.5268 ± 1.4251 | 2.1657 ± 0.2746 | 0.5308 | 1.33 × 10−10 | F (1,55) = 62.22, p < 0.001 |
| PC1~FA | 6.7809 ± 0.5283 | −1.5404 ± 0.1112 | 0.7771 | <2 × 10−16 | F (1,55) = 191.8, p < 0.001 |
| PC1~FP | −2.43085 ± 0.20452 | 0.26146 ± 0.05417 | 0.3864 | 2.41 × 10−5 | F (1,37) = 23.3, p < 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Wang, R.; Yang, X.; Dearing, J.A.; Doncaster, C.P.; Langdon, P.; Dong, X. Reconstruction of Ecological Transitions in a Temperate Shallow Lake of the Middle Yangtze River Basin in the Last Century. Water 2022, 14, 1136. https://doi.org/10.3390/w14071136
Zhao Y, Wang R, Yang X, Dearing JA, Doncaster CP, Langdon P, Dong X. Reconstruction of Ecological Transitions in a Temperate Shallow Lake of the Middle Yangtze River Basin in the Last Century. Water. 2022; 14(7):1136. https://doi.org/10.3390/w14071136
Chicago/Turabian StyleZhao, Yanjie, Rong Wang, Xiangdong Yang, John A. Dearing, Charles Patrick Doncaster, Peter Langdon, and Xuhui Dong. 2022. "Reconstruction of Ecological Transitions in a Temperate Shallow Lake of the Middle Yangtze River Basin in the Last Century" Water 14, no. 7: 1136. https://doi.org/10.3390/w14071136