
Simple Is the Best: An Alternative Method for the Analysis of Free-Living Nematode Assemblage Structure


1
Department of Biomolecular Sciences (DiSB), University of Urbino, 61029 Urbino, Italy
2
Fano Marine Center, The Inter-Institute Center for Research on Marine Biodiversity, Resources and Biotechnologies, 61032 Fano, Italy
3
Consorzio Nazionale Interuniversitario per le Scienze del Mare (CoNISMa) Rome, URL-Centro Direzionale—Isola C4, 80143 Naples, Italy
*
Author to whom correspondence should be addressed.
Water 2022, 14(7), 1114; https://doi.org/10.3390/w14071114
Submission received: 14 February 2022
/
Revised: 27 March 2022
/
Accepted: 28 March 2022
/
Published: 31 March 2022
(This article belongs to the Special Issue Ecological Quality Status Assessment of Aquatic Ecosystems: New Methods and Perspectives for the Future)
Abstract
:An alternative approach to notice changes of the nematode community structure was evaluated in five study cases in the Mediterranean Sea. In detail, we suggested a combination of morpho-functional traits (i.e., amphid, cuticle, buccal cavity, and tail shape) as an alternative to the taxonomic identification of nematodes. The results clearly demonstrated that the matrixes made using the trait code numbers perfectly mirror the changes of the nematode community structure at the genus level. The combination of the morpho-functional traits more frequently observed in areas under human pressure were ascribable to genera belonging to Xyalidae, Linhomoeidae and Chromadoridae families. This approach might greatly speed the analyses of nematode fauna in biomonitoring programs and might also be adopted for other meiobenthic organisms that may be categorized in functional groups opening new perspectives in the ecological assessment of meiofauna.
1. Introduction
In aquatic habitats, sediments characterized by large amounts of silt, clay, and organic load adsorb persistent pollutants (e.g., metals, PCBs) and polycyclic aromatic hydrocarbons (i.e., PAHs), becoming the main sink of contaminants brought into the marine system [1,2,3]. These sediment-bound pollutants can alter the state of the seabed, benthic processes, and functioning of ecosystems [4,5,6]. One of the most consistent effects of contamination in sea bottoms is a change in the structure of benthic biological assemblages, as pollution-sensitive species are lost and replaced by more tolerant ones, e.g., [7,8,9]. This makes the composition/structure changes of the benthic assemblages in themselves robust signals of alterations in the ecological quality status of the sediments. Therefore, noticing early alterations and warning conditions represent worldwide aims, as highlighted by all governmental regulations and sediment quality guidelines. However, routine monitoring still focuses mainly on surface waters, while sediment quality assessment remains a major concern; see [3,10] and references therein.
Numerous indices exist that consider diversity, abundance, and autoecology of benthic taxa for the quality assessment of sediments, but they mainly focus on macrofauna due to a long tradition and an extensive taxonomic and ecological knowledge of macrobenthos, see [11,12,13] for review. Notwithstanding macroinvertebrates showing consistent responses to anthropogenic impact [14,15], they are only one face of the benthic coin [16]. Indeed, benthic nematodes are the dominant and most diverse group in marine ecosystems [17,18,19], that suggest a high degree of specificity in their choice of natural environment. Nematodes living between sand particles are strongly influenced in their structure by environmental matrix variations in physicochemical properties and contaminant concentrations [20]. This issue, along with a generally high population stability, short generation times, low mobility, and direct benthic development, results in a wide range of specificity and rapid responses of nematodes to in situ disturbances [2]. The entire life cycle of numerous nematode species is completed in a few weeks, resulting in a much higher production-to-biomass ratio of meiofauna compared with macrofauna [21]. Moreover, nematodes play a crucial role as a trophic link between bacteria and larger fauna, enhancing the rate of: (a) carbon mineralization by stimulating microbial activity through predation; and/or (b) consumption of detritus by larger deposit-feeders [22]. Given their high abundance and wide distribution in all types of habitats, they provide a highly reliable dataset for statistical analysis, even when only limited sediment volumes are collected [23]. Therefore, despite nematodes being invisible to the human eye, their changes in assemblage structures might provide remarkable advantages for preventing actions in the management of coastal systems [2,8,24,25].
A general limitation of ecological investigations based on nematodes is related to the difficult and time-consuming taxonomic identification of species that requires the aid of expert taxonomists [26]. The implementation of molecular techniques and particularly environmental DNA (eDNA) metabarcoding has certainly created new perspectives, making nematode identification easier and faster and increasing the taxonomic resolution of the investigations [3,27]. Studies of pollution response have found metabarcoding to be more sensitive in revealing assemblage changes than the traditional morphological approach. However, drawbacks (possible biodiversity assessment bias by capturing signals from dead organisms, extracellular DNA; false readings due to taxonomic selectivity and restricted sensitivity of primers; unavoidability of primers and amplification bias; lack of comprehensive genetic databases) are still present and so they require caution in the interpretation of the quantitative results [28]. Thus, although molecular methods provide numerous benefits, the promise of barcoding as a routine and cheaper alternative to current biomonitoring practices is not yet realized and more user-friendly methods of data exploration are needed.
A way to overcome some of these limits might be the use of functional traits. They capture characteristics of organisms (i.e., morphological, physiological, phenological, and/or behavioral) that are linked to life-history and ecological functioning, and, at the same time, detect responses to anthropogenic disturbances and effects on ecological processes and services [29]. Species in functional groups share morphological traits that are thought to be linked to significant ecological functions [30]. Functional groups of nematodes can be regarded as groups of species that have similar influences on ecosystem processes [31]. Morphological structure of the buccal cavity, for example, was recognized by Wieser as related to specific trophic roles of the nematodes [32,33]; tail shape has an important part in locomotion, feeding, and reproduction [34,35]; body size may be related to species life history, physiology, and energy requirements [36,37,38]; amphids, as the main chemosensory organs of the nematode cephalic region, [39] take a role in the search for food and partners [40]; and body cuticles (i.e., morphology and thickness) as anti-predator, pollution, and hydrodynamic energy defense [41,42,43].
Thus, a combining approach of a few functional traits (i.e., buccal cavities, amphids, cuticles, and the shapes of tails) that can be easily identified in a short time by inexperienced people might also reflect a possible loss of ecosystem functionality after pollution.
The aim of the present study is to demonstrate that an approach of combining functional traits may efficiently mirror the nematode taxonomic structure and document any variation of the nematode community. To deal with this aim, we have considered a number of study cases focused on various typologies of human disturbances (i.e., ports, marinas, fish farms, urban riverine discharges) carried out in the Mediterranean Sea. We compared the results of the changes in the structure of the nematode assemblages identified at the genus level and the combination of the four morpho-functional traits in five study cases. Thus, we first assigned the four morpho-functional traits to each taxon found in the single dataset, then we obtained a new data matrix based on the traits’ number codes and compared the trends observed using the nematode taxonomic composition. This approach, if recognized as able to capture nematode variations, might be a very low-cost solution (both in terms of time and money) to overcome the problem of nematode taxonomic identification and an alternative method easily applicable in biomonitoring by inexperienced people (e.g., students, environmental agency staff, voluntary citizens). In this paper, three main questions are addressed: Does the combination of morpho-functional traits reflect the taxonomic composition of nematode fauna? Does the combination of morpho-functional traits detect the effects of a different human pressure? Are there recurrent combinations of morpho-functional traits in nematode communities in polluted sediments?
2. Materials and Methods
2.1. Study Areas
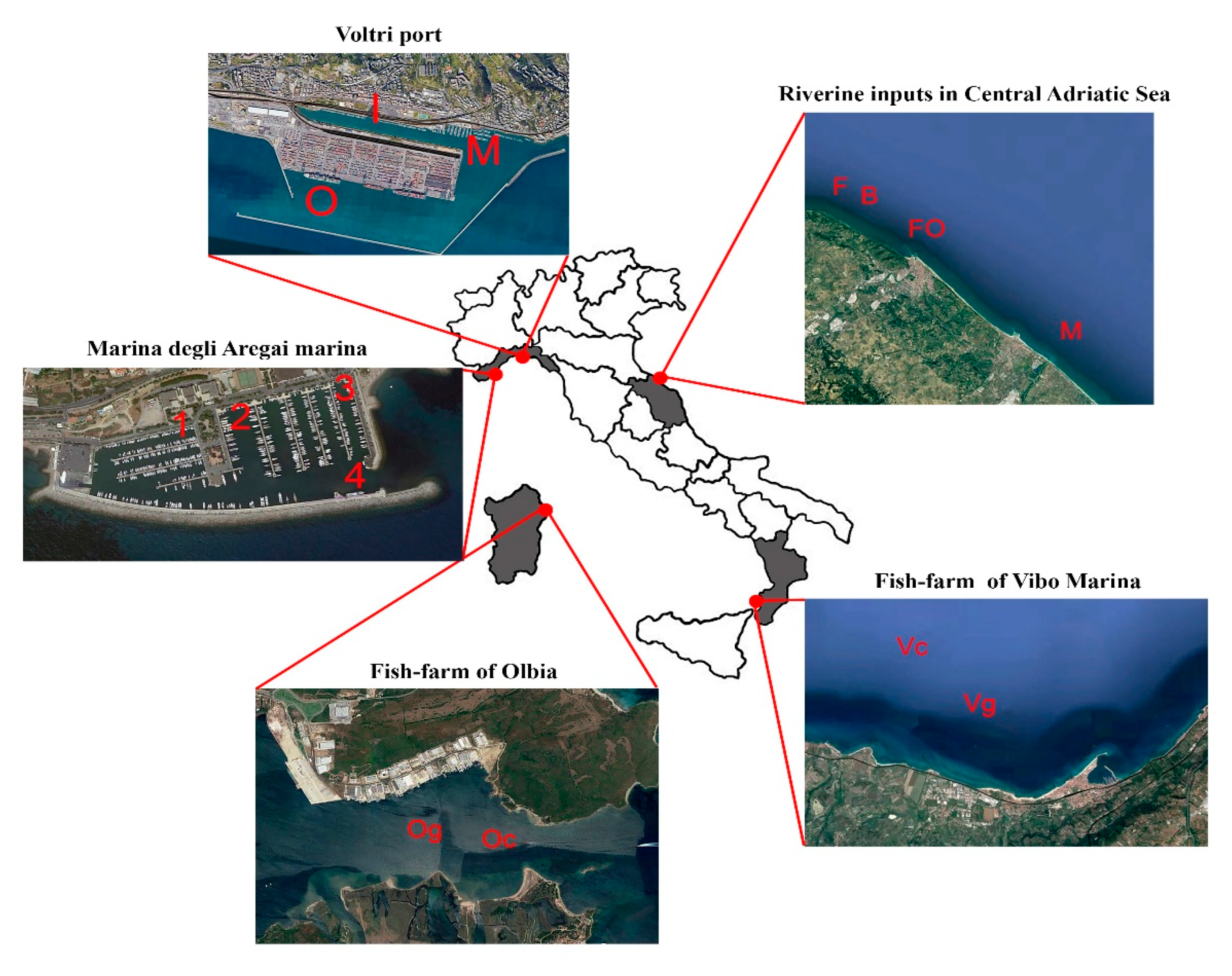
In the present study, the data sets of five study cases from the Mediterranean Sea were considered to demonstrate the efficiency of the combination of morpho-functional traits of nematodes. The coastal areas are exposed to different types of human activities/pressures: ports, marinas, fish farms, and discharge of urban rivers (Figure 1 and Table S1). Sediment samples (three replicates for each station) were collected by SCUBA divers or using modified Van Veen grabs. In this last case, the undisturbed aliquots of sediments were subsampled using plexiglass corers. The main information on sites, sampling routines, experimental designs, and sample processing techniques is hereafter summarized (Table S1).
2.1.1. Port-Contaminated Sediments from Ligurian Sea (NW Mediterranean)
The industrial port of Genoa-Voltri is an important container and oil terminal (total area ~34.5 ha). Samples were collected in four periods (i.e., June, July, and November 2002; and February 2003) at a mean depth of 9.5 m in three sampling stations selected on the basis of their position within the port: St. I (inner port area), St. M (central port area), and St. O (outer port area, close to the open sea). Substrata were mainly characterized by fine sands and muddy sediments. St. M resulted the most contaminated area (the highest concentrations of organic matter, lowest organic matter quality i.e., low PRT:CHO ratio, highest bacterial densities, and PAHs), followed by St. I [44].
Marina degli Aregai is a tourist marina where sediments (fine sands) were collected at a range from 3 to 12 m of depth. Sampling was carried out in two periods (i.e., July 2005 and January 2006) and at four stations from the inner (St. 1) to outer port part, close to the port mouth (St. 4). Fine sands dominated the substrata and contaminants showed higher concentrations at St. 4 (i.e., highest organic load, Pb, Cr, and PAH concentrations) [45].
2.1.2. Coastal Sediments Influenced by Riverine Inputs in Central Adriatic Sea
The study area was in the Central Adriatic Sea (Central Mediterranean), close to Pesaro. Most relevant pollution sources are the runoffs of the local rivers (i.e., Foglia, Metauro, and Tavollo) and, after severe rains, Po River plumes. A total of four positions were selected for the sampling: two within the Natural Regional Park of Monte San Bartolo (i.e., St. B, and St. FI) and two others in front of the mouths of Foglia and Metauro rivers (i.e., St. M and St. FO). Samples were collected ~5 m in depth from each station in two different periods (November 2011 and June 2012). Sediments of the study area are fine sands. Both the rivers receive wastewater from urban and industrial areas and livestock farms, and the Foglia River especially causes frequent eutrophication phenomena [46,47]. Furthermore, the occurrence of low-salinity waters and high levels of Chl-a in the FI station suggested the influence of the Po River plume also confirmed by clay mineralogy data [48].
2.1.3. Fish Farm Impact in the Tyrrhenian Sea
Two areas were under scrutiny for fish farm disturbance: the first one is in the sheltered Gulf of Olbia (Northern Sardinia, Tyrrhenian Sea) [7] and the second one is an open sea area at Vibo Marina (Calabria), SW Italy (W Mediterranean Sea) [49]. Many anthropogenic activities are present in the Gulf of Olbia: tourist and commercial harbors, mussel farms, fish farming, and river outflows contributed to affect the benthic communities [7]. Sampling was carried out in two periods (i.e., April 2006 and September 2006) in two stations (~6 m of depth): one under cages (Og) and the other, regarded as control, far from the farming (Oc). Sediments were mainly characterized by fine sand fraction, especially under the cages (i.e., at St. Og) where a high level of organic matter, heavy metals, and PAHs were found.
The bluefin tuna farming of Vibo Marina was located in open sea at ~46 m in depth. Samplings were carried out in October 2005 and May 2006 at four stations: two close to the floating cages (Vg) and two others (controls) far from the farming (Vc). Sediments were mainly characterized by sand with a fine fraction accounting for ~40–50% [49]. Despite the presence of the floating cages, a moderate amount of organic matter was recorded and was slightly higher at the control station. Globally, the parameters measured in this study did not point to a detectable impact of organic waste due to the fattening of Atlantic bluefin tuna (Thunnus thynnus). This was likely due to the ecological context (e.g., oligotrophy) and the exposed nature of the Vibo Marine area characterized by high water depth and a strong hydrodynamic regime [49].
2.2. Nematodes
For nematode analyses, details of sample treatment and laboratory analysis are given in the relative papers reported in Table S1. Once in the laboratory, sediment samples were firstly sieved through 500–38 µm mesh sieves, then the retained fraction was centrifuged three times in a gradient of Ludox HS 30 (density 1.18 g cm−3) as described by Danovaro et al. [50]. One hundred nematodes of those so obtained were randomly picked up and mounted on permanent slides according to Seinhorst [51]. Specimens were identified at the genus level with the aid of the pictorial keys of Platt and Warwick [52,53] and Warwick et al. [54], as well as the use of the NeMys website [19].
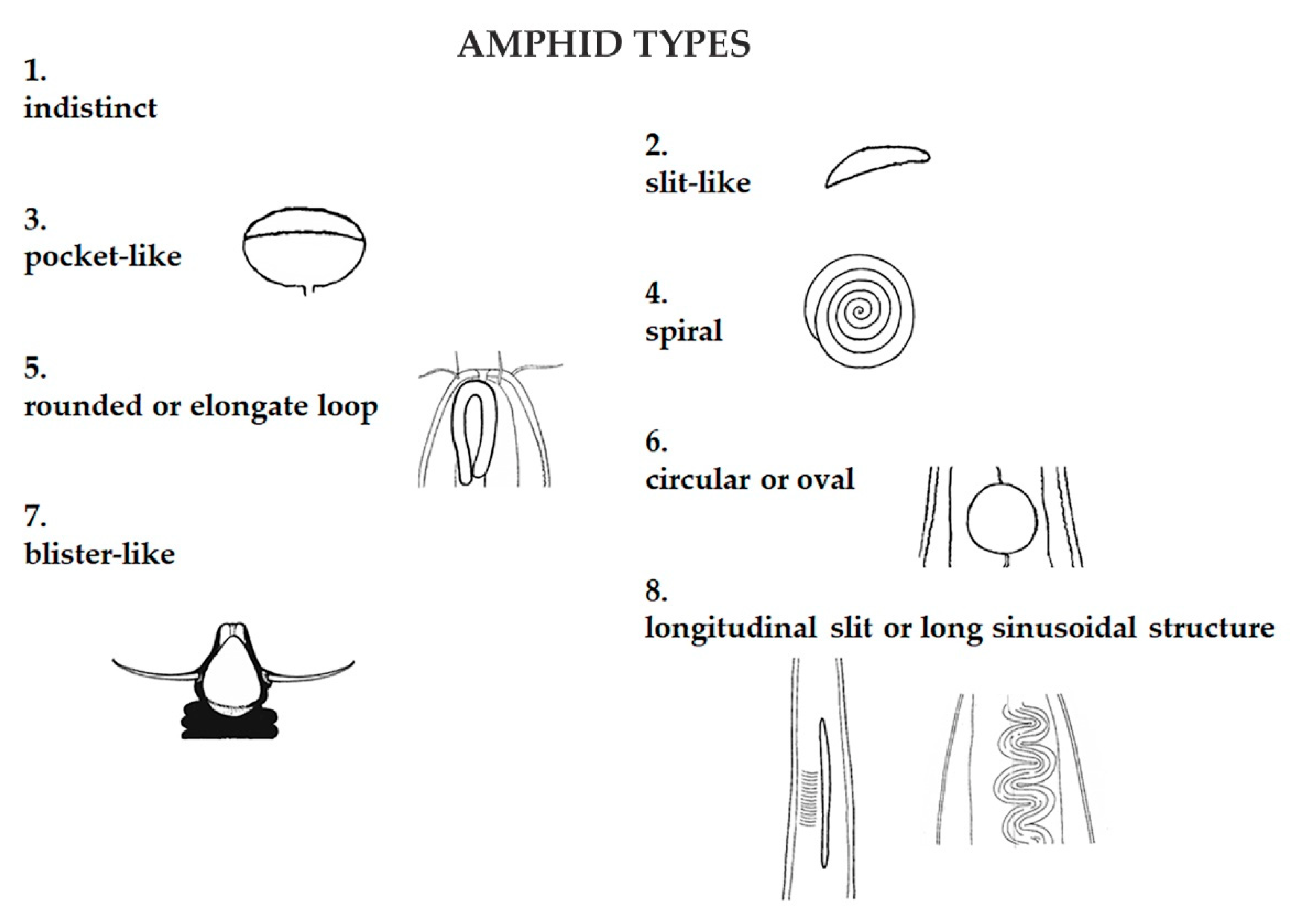
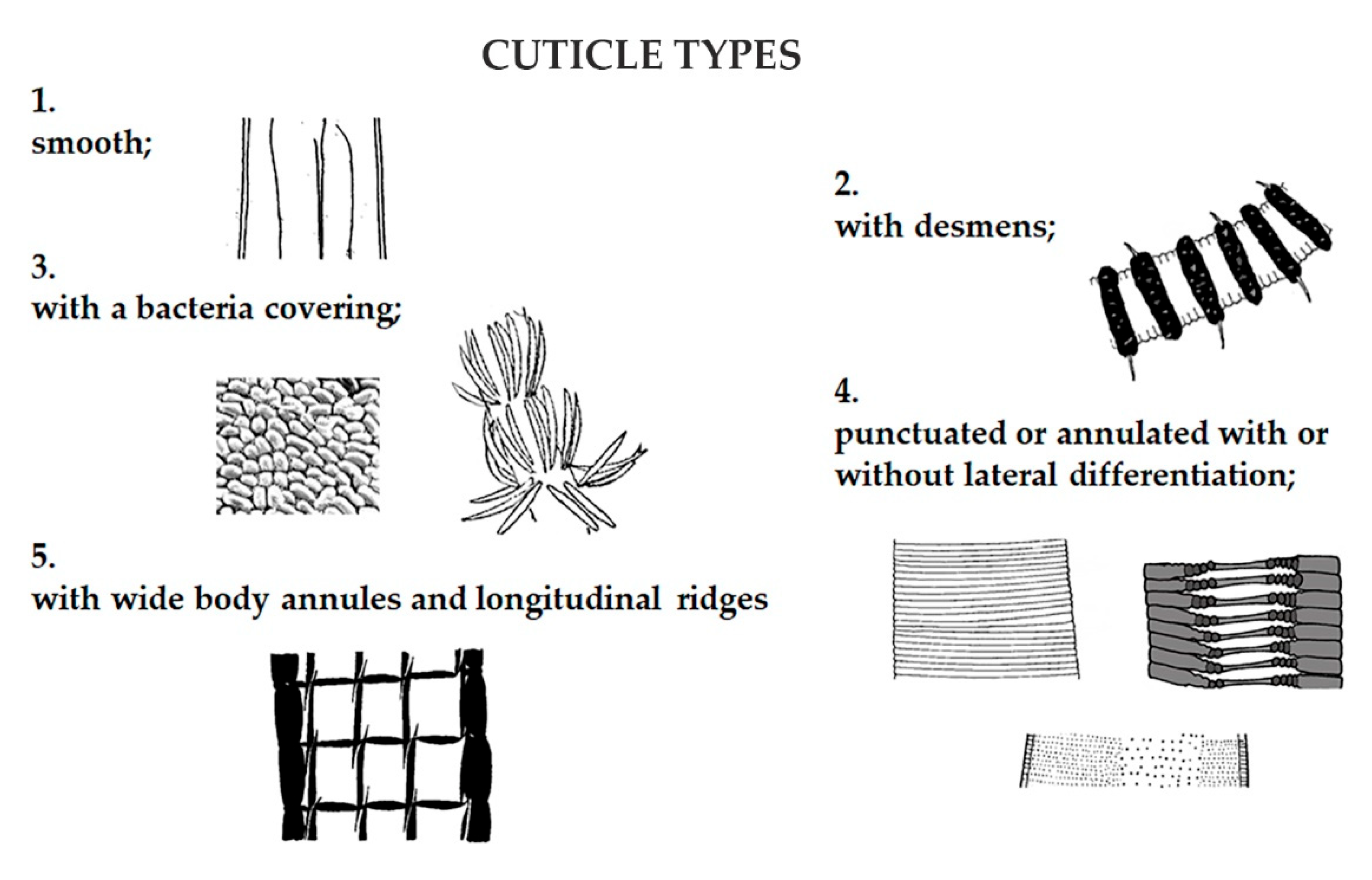
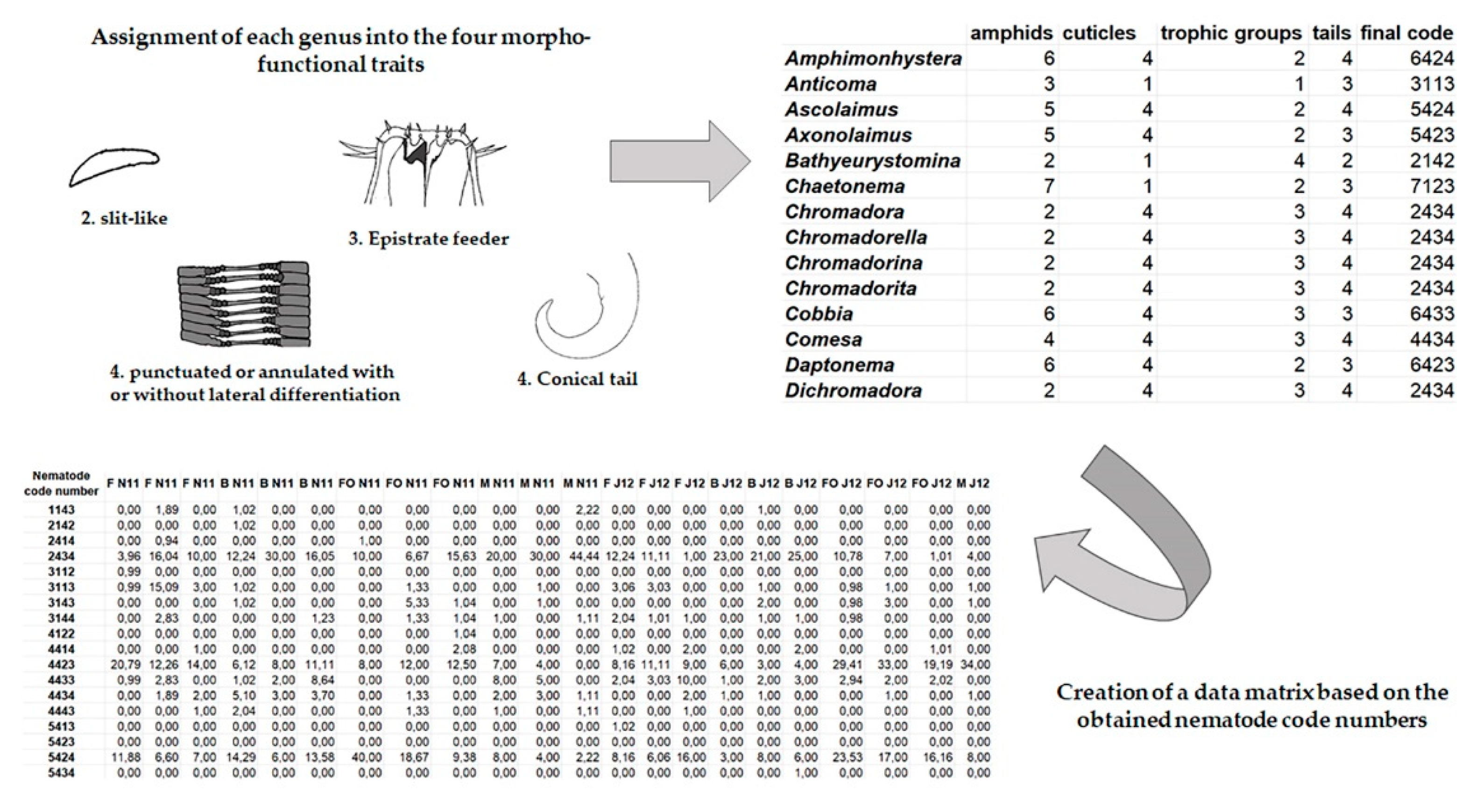
Furthermore, nematodes were characterized according to their morpho-functional traits. Amphid and cuticle structure were categorized according to the classification by Semprucci et al. [42]. Nematodes were assigned to eight categories based on amphid structure: indistinct, slit-like, pocket-like, spiral, rounded or elongate loop, circular-oval, blister-like, and longitudinal slit (Figure 2); and five based on cuticle structure: smooth, with desmens, with bacteria covering, punctuated or annulated with or without lateral differentiation, and with wide body annules and longitudinal ridges (Figure 3).
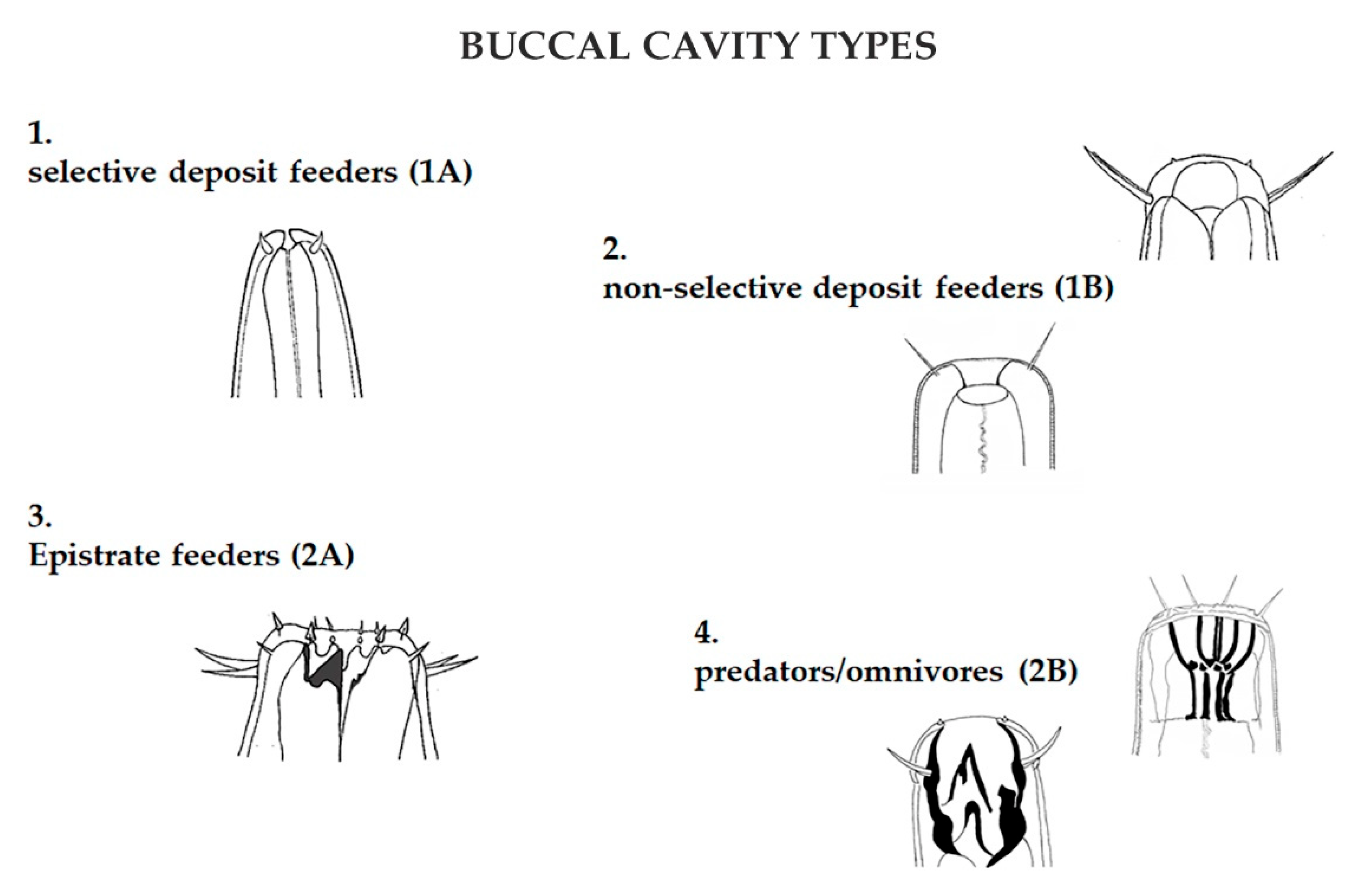
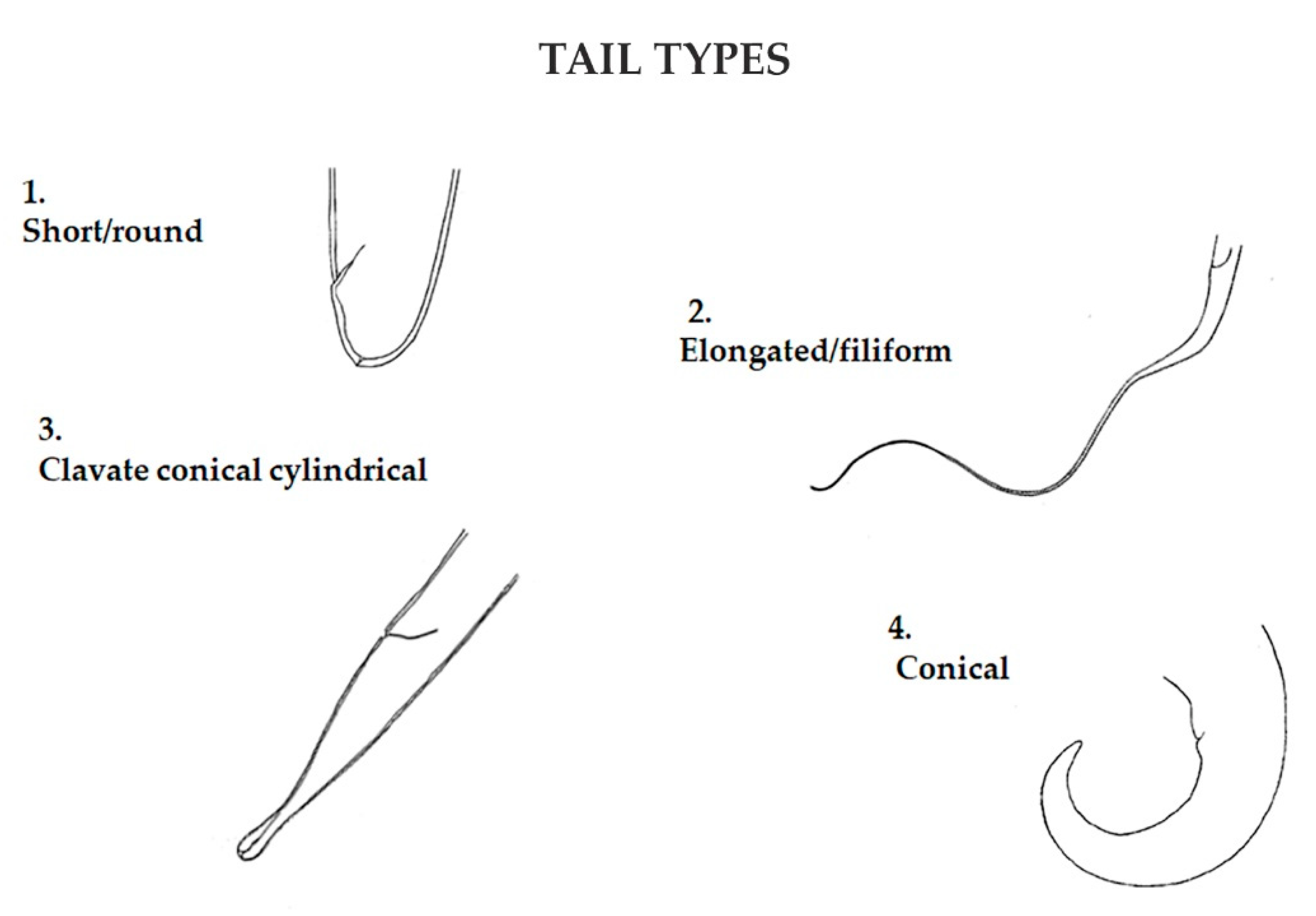
Specimens were then assigned to four trophic groups, according to Wieser [32], based on buccal morphology: selective (1A) and non-selective (1B) deposit feeders, epistratum feeders (2A), and predators/omnivores (2B) (Figure 4). As for the tail shape, specimens were assigned to one of four categories: short/round; elongated/filiform; conical and clavate/conical-cylindrical [35] (Figure 5). The four functional traits (buccal cavity, amphid, cuticle, tail) were assigned to each specimen found obtaining a four-digit number that identified each nematode (see Figure 6).
2.3. Data Analysis
All nematodes with the same code number were grouped together and utilized to produce a data matrix for each study case. Data matrices for the multivariate analyses were based on percentages of nematode genera and combinations of morpho-functional characteristics to compare changes in nematode community structure among sampling stations at each study area.
To visualize and compare the differences of the community structure (genus composition versus the combination of morpho-functional traits), non-Multidimensional scaling (nMDS) was applied to genera and trait codes after the Bray–Curtis similarity index computation. According to Schratzberger et al. [31], the matrix based on the combination of morpho-functional traits was not transformed and, to make comparison possible, neither were those on the genera. Spatial and temporal trends were further checked by means of Analysis of Similarities (ANOSIM) to assess the presence of significant differences between nematode communities at the stations. SIMPER test (cut-off, 90%, Bray–Curtis similarities) was applied to the genera and combination of functional traits to find the taxa or traits that mainly distinguished the disturbed stations. Shannon diversity (H’, log2) and Pielou evenness (J) indices were calculated for the functional and taxonomic structure of the nematode communities, respectively, in order to check whether the indices show the same result/trend in each case. The software package Primer v.6 [55] was utilized for all the multivariate analyses as well as index computation (i.e., H’ and J).
3. Results and Discussion
A total of 12,000 nematodes were analyzed, which could be assigned to 121 genera from 35 families and 47 different morpho-functional trait combinations (Table S2). As reported above, we posed three questions that are addressed in light of the outcomes from the analysis of genera composition and combination of the morpho-functional traits collected at five study case stations in the Mediterranean Sea.
3.1. Does the Combination of Morpho-Functional Traits Reflect the Taxonomic Composition of Nematode Fauna?
Many surveys on nematodes have highlighted in the past decades that the nematode assemblage structure is the first and most efficient tool to detect environmental changes and pollution effects [7]. The need to develop alternative indices (i.e., index of trophic diversity, maturity index, biological trait analysis) [56] or tools for ecological assessment is driven by the necessity to speed the analysis process and overcome the problems related to nematode taxonomic identification. Semprucci et al. [42] and Zullini and Semprucci [40] suggested that there are detectable morphological differences between free-living aquatic (both marine and freshwater) and soil nematodes strictly associated to the environmental variations and that might be used to document the changes in the structure composition.
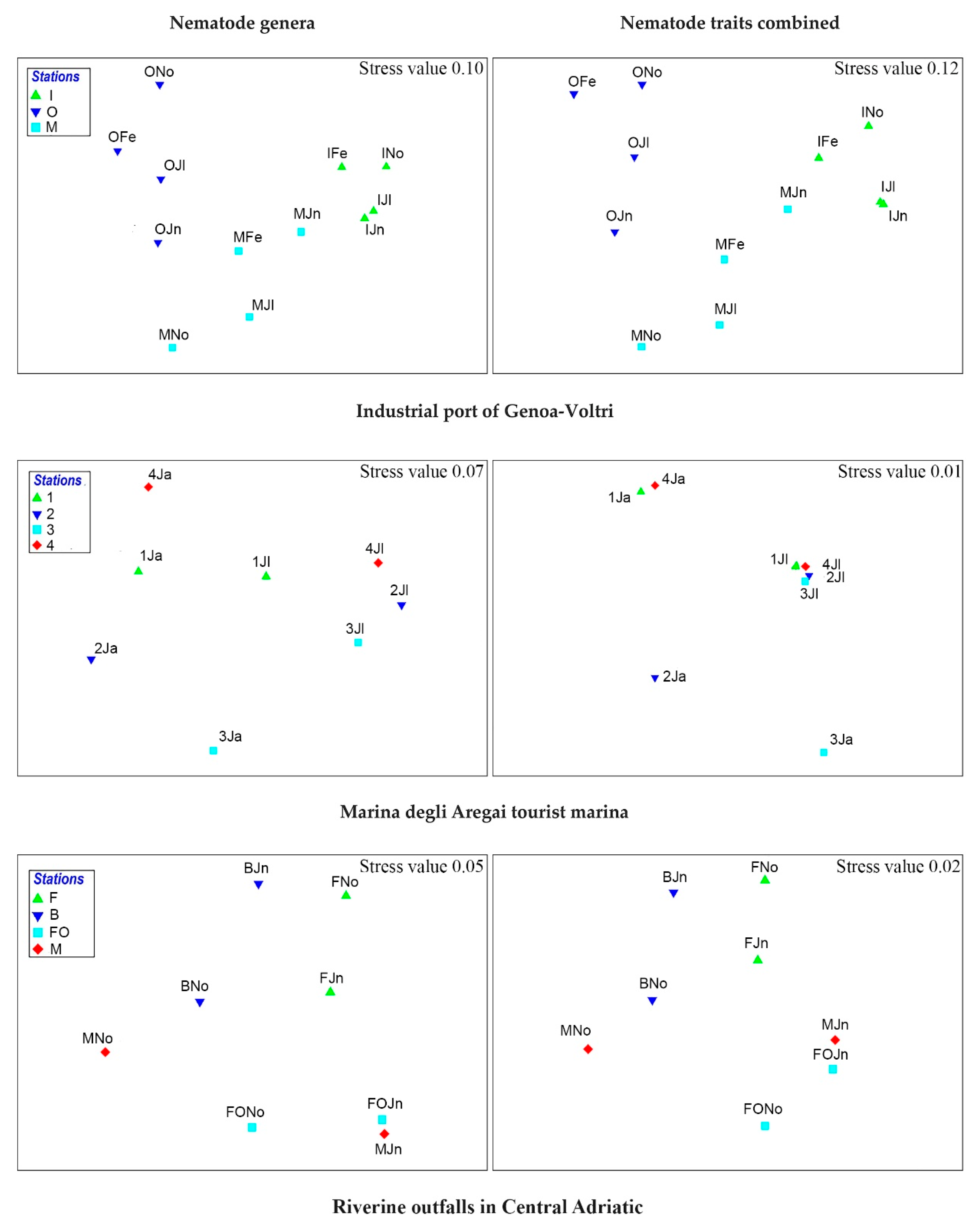
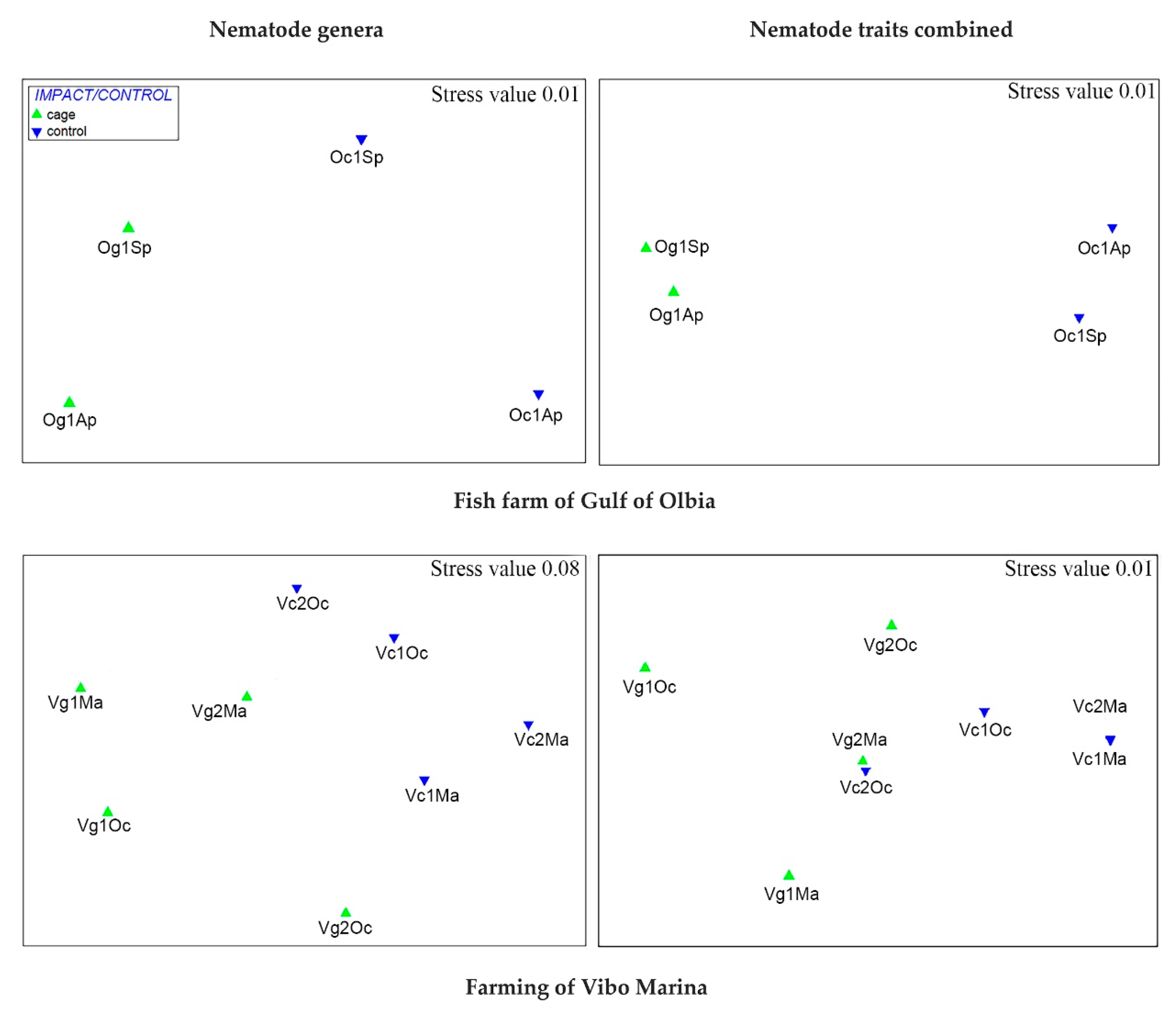
Multivariate statistical analysis such as nMDS revealed that the two data matrixes i.e., genera and combination of morpho-functional traits showed similar sub-divisions of the sampling stations in all the study cases considered (see Figure 7 and Figure 8). This was further evidenced by the analysis of similarities that detected approximately the same R ratio as well as level of significance (when occurred), especially in the comparisons between stations (see Table 1).
Thus, the finding that the functional trait code reflects the taxonomic structure of the nematode communities at the genus level in all cases studied suggests that this simple method could be a way to routinely use nematodes in biomonitoring programs at low financial cost and in a more time-efficient way.
The potentialities of this approach are further demonstrated by Armenteros et al. [57] that analyzed functional changes in the nematode assemblages across the wide span of habitats investigated in the Cuban Archipelago. They, in particular, reported an unclear pattern of amphidal fovea and cuticle types across the habitats analyzed and argued that different types of amphidal fovea or cuticle did not play an adaptive role in the behavior of nematodes, but likely reflect phylogenetic signatures from families/genera living in different habitat conditions. However, this statement is per se a crucial point that corroborates our idea. Most of the characteristics considered diagnostic in nematodes are functionally determined [58]. Therefore, the combination of these morpho-functional traits can take a snapshot of the taxonomic composition and identify possible assemblage changes.
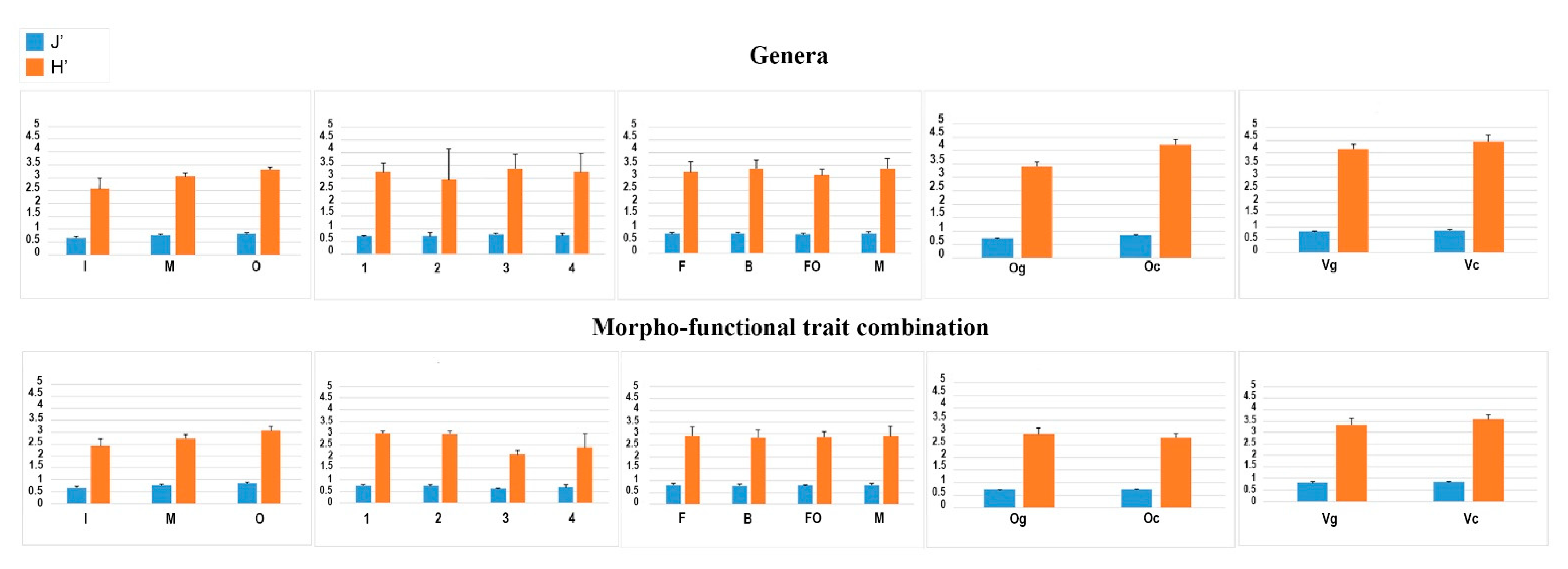
The diversity measures (i.e., H’ and J) calculated for morpho-functional and taxonomic nematode community structure revealed consistent results in the ports from the Ligurian Sea and in the Vibo fish farm, but not in the Olbia farm or Adriatic Sea (Figure 9). Therefore, more than the univariate diversity measures, the multivariate approach to trait combination is recommended.
3.2. Does the Combination of Morpho-Functional Traits Detect the Effect of a Different Human Pressure?
Spatial and temporal trends were checked by ANOSIM. The trait combination perfectly distinguished the pollution gradient of the port of Voltri from the inner station to the open sea station. St. M resulted a sampling point with all pollutants showing higher concentrations [44] along with a distinct morpho-functional trait combination represented mainly by 6423 (combination of circular or oval amphids, punctuated or annulated cuticle with or without lateral differentiation, 1B guild, clavate conical cylindrical tail) and 3143 (i.e., pocket-like amphid, smooth cuticle, 2B group, clavate conical cylindrical body end) (Table S3), which partially fits with the abundant taxa found by Moreno and co-authors in the original paper (i.e., Daptonema, Terschellingia, Paracomesoma and Metoncholaimus) [44] (Table S4).
In the Aregai tourist marina, more marked temporal rather than spatial differences were observed in the genus assemblage structure [45] as well as in the trait combination (Table 1) (Figure 7). However, a noticeably greater spatial heterogeneity of the communities (genus and functional trait combined) as well as the lowest diversity values were detected in January, likely in relation with the highest contaminant concentrations in the winter period [45].
The SIMPER analysis revealed that the most abundant functional trait combination in the most polluted station (i.e., St. 4) was again 6423 (circular or oval amphids, punctuated or annulated with or without lateral differentiation cuticle, 1B guild, clavate conical cylindrical tail) together with 2434 (slit-like amphid, punctuated or annulated cuticle with or without lateral differentiation, 2A guild, and conical tail) likely related to the higher abundance of Xyalidae, Linhomoeidae (e.g., 6423), and Chromadoridae (2434), respectively (Tables S3 and S4) [45].
The nMDS plots of the Adriatic Sea showed very comparable station clusters with both the approaches. This area is known for eutrophication phenomena mainly related to local river runoffs. Accordingly, the nematode assemblages of the Foglia and Metauro rivers (FO and M) showed the most marked differences from the other two stations (i.e., B and FI) as confirmed by ANOSIM. Indeed, FO and M stations did not show significant differences between them but proved to be significantly different from all other localities. Another station that revealed a high level of dissimilarity from the others was Fiorenzuola (St. F), but these differences were very much related to the effects of the Po River plume rather than to the influence of the local rivers [46,47,48] (Table 1). As in the Voltri port and Aregai tourist marina, 6423 was one of the combinations of traits more frequently found in the human disturbed stations, along with 4423 (i.e., spiral amphid, punctuated or annulated with or without lateral differentiation cuticles, 1B group, clavate conical cylindrical tail) and 5424 (rounded or elongate loop, punctuated or annulated cuticle with or without lateral differentiation, 1B, conical tail) (Table S3). This finding matches with the genera that mainly distinguished the stations mostly influenced by riverine discharges (i.e., Paramonohystera, Stylotheristus, Theristus, Sabatieria and Odontophora) [46].
The differences between the assemblages of fish farm cages and of controls are evident in both areas studied in the Tyrrhenian Sea even if they were more marked in Olbia (Sardinia). Indeed, SIMPER routine revealed a higher level of dissimilarity between Olbia control and cage (57%) than between Vibo control and cage (43%) (Table S3). Trait combination 6423 again characterized the stations under the cages in both fish farms (mainly due to the high abundance of Daptonema) together with trait combination 6413 (i.e., round or oval amphids, punctate or annulate cuticle with or without lateral differentiation, 1A trophic group, conical-cylindrical tail) in Olbia (represented by the high abundance of Terschellingia) and feature combination 2434 in Vibo (e.g., Ptycholaimellus) (Tables S3 and S4).
Instead, the diversity computed from the functional trait matrix was inconsistent and less helpful than the diversity based on genera in identifying areas with greater anthropogenic disturbance, so we suggest caution in its use (Figure 9).
3.3. Are There Recurrent Combinations of Morpho-Functional Traits in Nematode Communities in Polluted Sediments?
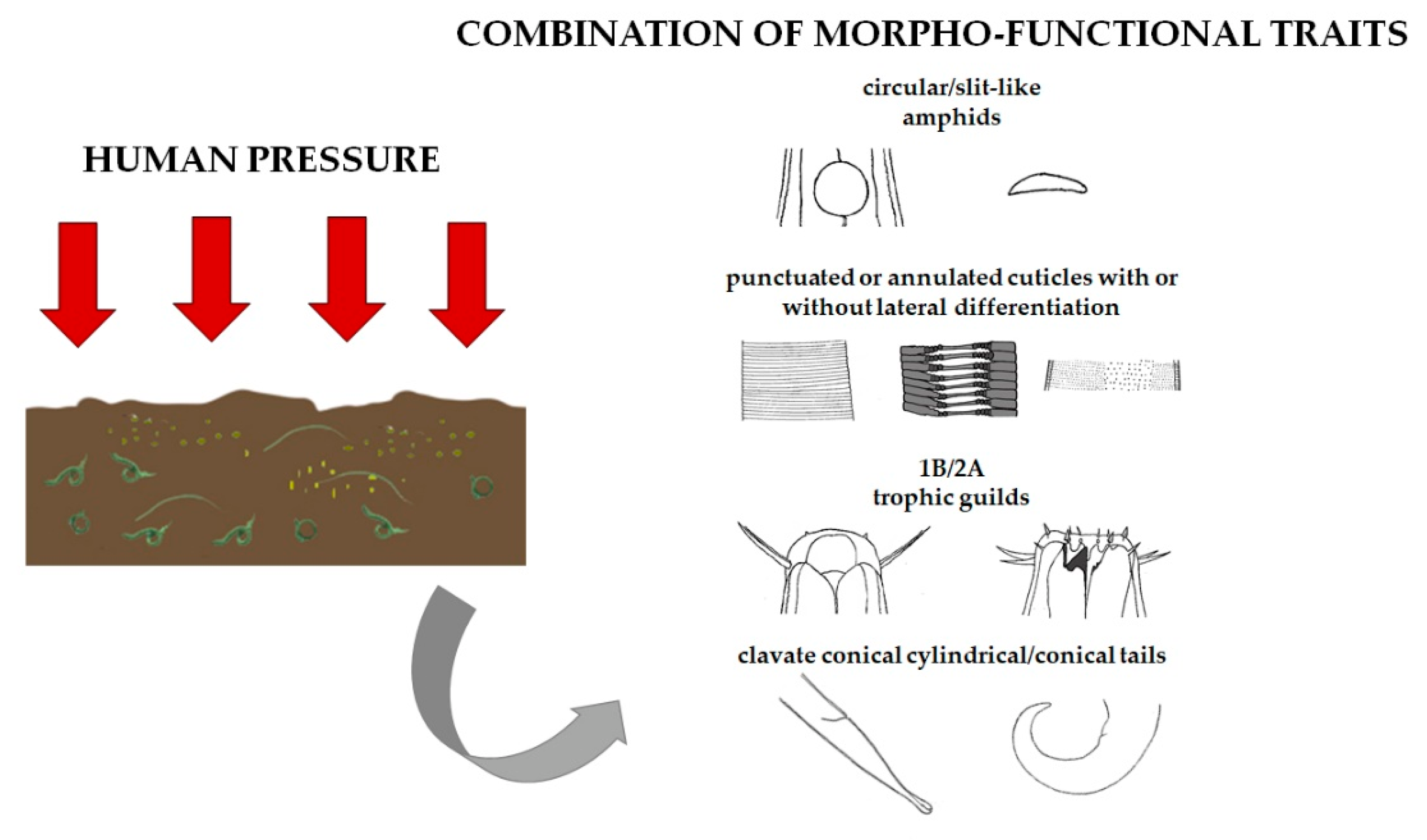
The high frequency of 6423 and 2434 combinations in the most impacted areas allows us to conclude that there are recurring trait associations in stressful conditions and are mainly related to the high abundance of genera belonging to Xyalidae, Linhomoeidae, and Chromadoridae genera. As reported in Figure 10, circular-oval and slit-like amphids, punctuated or annulated cuticles with or without lateral differentiation, 1B and 2A trophic guilds, and clavate conical cylindrical/conical tails are the prevalent traits.
The most anthropogenically disturbed stations favor opportunistic trophic guilds such as 1B and 2A that are able to take advantage of a wide range of food items from organic deposits and bacteria biofilm to microphytobenthos [59,60,61]. All of them are abundant in sediments under anthropogenic conditions [44,62]. In addition, 2A feeding mode (i.e., scraping food off surfaces, piercing it, and sucking out the content) allows nematodes to reduce the probability of ingesting xenobiotics, which is an advantage in contaminated sediments [60]. The occurrence of 2B group, e.g., in the commercial port of Voltri, is mainly restricted to the scavenging Oncholaimids. Scavengers still follow a tactical trophic strategy through which nematodes take advantage of the consumption of dead organisms (including plant material). They play a key role in the ecosystems and many species belonging to Oncholaimidae are successful extremophiles [61,63,64,65]: Oncholaimus campylocercoides shows a sulphide detoxification via formation of inclusions that allow it to tolerate sulphidic conditions and to benefit from the ‘sulphide niche’; Oncholaimus dyvae and Metoncholaimus albidus appear able to adapt to and be very abundant in extreme conditions due to their association with chemosynthetic micro-organisms.
Free-living nematodes evaluate the quality of the environment that they inhabit and food resource diversity mainly through the complex chemosensory system of the amphid [66,67]. In polluted sediments, nematodes exhibit a high frequency of circular amphids, which confirms the role of such a type and size of amphidal fovea in the detection and avoidance of xenobiotics [68]. Instead, small slit-like amphids belong to Chromadorids that, due to their trophic strategy (i.e., 2A), are already able to reduce their ingestion of xenobiotics. Furthermore, the occurrence of such a high percentage of small amphids is related to the fact that food is not a limiting factor in some of the disturbed sediments, especially for nematode species able to profit from a wide spectrum of trophic resources (microbes, organic material, microalgae).
Nematode cuticle is an extremely flexible and resistant exoskeleton essential for maintenance of body morphology and integrity and has a critical role in protection and locomotion via attachment to body-wall muscles [69]. Punctuated or annulated cuticle with or without lateral differentiation is a type of trait shared by numerous marine nematode families. For instance, nematodes such as Terschellingia longicaudata (Linhomoeidae) and Ptycholaimellus ponticus (Chromadoridae) belong to this category, and it might be related to oxygen consumption rate and locomotion [58,70]. Instead, clavate conical cylindrical/conical tails are probably strictly associated with the silt sediment fraction that characterized the sediments of the main part of the study areas [31,71].
4. Conclusions
In this paper, we suggested an approach based on a combination of morpho-functional traits (i.e., amphid, cuticle, buccal cavity, and tail shape) that might greatly speed the analyses of nematodes and might be used by unexperienced people in biomonitoring programs. We demonstrated that the simple combination of the four traits in a single code number may perfectly mirror the taxonomic structure of the nematode assemblage at the genus level and we, therefore, predict that similar results can be also obtained directly encoding nematode specimens with the four selected traits. This type of approach, if corroborated by additional studies, might also be adopted for other meiobenthic organisms that can be categorized in functional groups such as foraminifera or copepods, opening new perspectives in the assessment of many benthic taxa.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w14071114/s1, Table S1: Overview of the study cases considered in the present analysis; Table S2. List of families and genera from each sampling area and relative combination of traits. Table S3. Results of the SIMPER test carried out on the functional trait codes. Table S4. Results of the SIMPER test carried out on the nematode genera.
Author Contributions
Conceptualization, validation, formal analysis, writing—original draft preparation F.S.; data curation, E.G.; writing—original draft preparation, M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank Mariapaola Moreno, University of Genoa, a friend and colleague, who shared with us the data matrices of nematodes from Ligurian Sea and Tyrrhenian Sea.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Matsuyama, A.; Yano, S.; Taninaka, T.; Kindaichi, M.; Sonoda, I.; Tada, A.; Akagi, H. Chemical characteristics of dissolved mercury in the pore water of Minamata Bay sediments. Mar. Pollut. Bull. 2018, 129, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Losi, V.; Grassi, E.; Balsamo, M.; Rocchi, M.; Gaozza, L.; Semprucci, F. Changes in taxonomic structure and functional traits of nematodes as tools in the assessment of port impact. Estuar. Coast. Shelf Sci. 2021, 260, 107524. [Google Scholar] [CrossRef]
- Pawlowski, J.; Bruce, K.; Panksep, K.; Aguirre, F.I.; Amalfitano, S.; Apothéloz-Perret-Gentil, L.; Baussant, T.; Bouchez, A.; Carugati, L.; Cermakova, K.; et al. Environmental DNA metabarcoding for benthic monitoring: A review of sediment sampling and DNA extraction methods. Sci. Total Environ. 2021, 19, 151783. [Google Scholar] [CrossRef]
- Armynot du Châtelet, E.; Bout-Roumazeilles, V.; Coccioni, R.; Frontalini, F.; Francescangeli, F.; Margaritelli, G.; Rettori, R.; Spagnoli, F.; Semprucci, F.; Trentesaux, A.; et al. Environmental control on a land-sea transitional setting–Integrated microfaunal, sedimentological, and geochemical approaches. Environ. Earth Sci. 2016, 75, 123. [Google Scholar] [CrossRef]
- Gambi, C.; Dell’Anno, A.; Corinaldesi, C.; Lo Martire, M.; Musco, L.; Da Ros, Z.; Armiento, G.; Danovaro, R. Impact of historical contamination on meiofaunal assemblages: The case study of Bagnoli-Coroglio Bay (southern Tyrrhenian Sea). Mar. Environ. Res. 2020, 156, 104907. [Google Scholar] [CrossRef] [PubMed]
- Schratzberger, M.; Somerfield, P.J. Effects of widespread human disturbances in the marine environment suggest a new agenda for meiofauna research is needed. Sci. Total Environ. 2020, 728, 138435. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Semprucci, F.; Vezzulli, L.; Balsamo, M.; Fabiano, M.; Albertelli, G. The use of nematodes in assessing ecological quality status in the Mediterranean coastal ecosystems. Ecol. Indic. 2011, 11, 328–336. [Google Scholar] [CrossRef]
- Hong, J.H.; Semprucci, F.; Raehyuk, J.; Kim, K.; Lee, S.; Jeon, D.; Yoo, H.; Kim, J.; Kim, J.; Yeom, J.; et al. Meiobenthic nematodes in the assessment of the relative impact of human activities on coastal marine ecosystem. Environ. Monit. Assess. 2020, 192, 81. [Google Scholar] [CrossRef]
- Sahraeian, N.; Sahafi, H.H.; Mosallanejad, H.; Ingels, J.; Semprucci, F. Temporal and spatial variability of free-living nematodes in a beach system characterized by domestic and industrial impacts (Bandar Abbas, Persian Gulf, Iran). Ecol. Indic. 2020, 118, 106697. [Google Scholar] [CrossRef]
- Baldrighi, E.; Semprucci, F.; Franzo, A.; Cvitkovic, I.; Bogner, D.; Despalatovic, M.; Berto, D.; Malgorzata Formalewicz, M.; Scarpato, A.; Frapiccini, E.; et al. Meiofaunal communities in four Adriatic ports: Baseline data for risk assessment in ballast water management. Mar. Pollut. Bull. 2019, 147, 171–184. [Google Scholar] [CrossRef]
- Nelson, W.G. A quantitative assessment of organic carbon content as a regional sediment-condition indicator. Ecol. Indic. 2020, 114, 106318. [Google Scholar] [CrossRef] [PubMed]
- Kiddon, J.; Sullivan, H.; Nelson, W.; Pelletier, M.; Harwell, L.; Nord, M.; Paulsen, S. Lessons learned from 30 years of assessing U.S. coastal water. In Water Quality–Science, Assessments and Policy; Summers, K., Ed.; InTechOpen: Rijeka, Croatia, 2020; pp. 1–22. [Google Scholar]
- Guerra-García, J.M.; Navarro-Barranco, C.; Ros, M.; Sedano, F.; Espinar, R.; Fernández-Romero, A.; Martínez-Laiz, G.; Cuesta, J.A.; Giráldez, I.; Morales, E.; et al. Ecological quality assessement of marinas: An integrative approach combining, biological and environmental data. J. Environ. Manag. 2021, 15, 286–112237. [Google Scholar] [CrossRef]
- Diaz, R.J.; Solan, M.; Valente, R.M. A review of approaches for classifying benthic habitats and evaluating habitat quality. J. Environ. Manag. 2004, 73, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Borja, A.; Barbone, E.; Basset, A.; Borgersen, G.; Brkljacic, M.; Elliott, M.; Garmendia, J.M.; Marques, J.C.; Mazik, K.; Muxika, I.; et al. Response of single benthic metrics and multi-metric methods to anthropogenic pressure gradients, in five distinct European coastal and transitional ecosystems. Mar. Pollut. Bull. 2011, 62, 499–513. [Google Scholar] [CrossRef]
- Vanaverbeke, J.; Merckx, B.; Degraer, S.; Vincx, M. Sediment-related distribution patterns of nematodes and macrofauna: Two sides of the benthic coin? Mar. Environ. Res. 2011, 71, 31–40. [Google Scholar] [CrossRef]
- Appeltans, W.; Ahyong, S.T.; Anderson, G.; Angel, M.V.; Artois, T.; Bailly, N.; Costello, M.J. The Magnitude of Global Marine Species Diversity. Curr. Biol. 2012, 22, 2189–2202. [Google Scholar] [CrossRef] [Green Version]
- Jouili, S.; Essid, N.; Semprucci, F.; Boufahja, F.; Nasri, A.; Beyrem, H. Environmental quality assessment of El Bibane lagoon (Tunisia) using taxonomical and functional diversity of meiofauna and nematodes. J. Mar. Biol. Assoc. UK 2017, 97, 1593–1603. [Google Scholar] [CrossRef]
- Bezerra, T.N.; Decraemer, W.; Eisendle-Flöckner, U.; Hodda, M.; Holovachov, O.; Leduc, D.; Miljutin, D.; Mokievsky, V.; Peña Santiago, R.; Sharma, J.; et al. Nemys: World Database of Nematodes. Available online: http://nemys.ugent.be/ (accessed on 19 December 2021).
- Coull, B.C.; Chandler, G.T. Pollution and meiofauna: Field, laboratory and mesocosm studies. Oceanogr. Mar. Biol. 1992, 30, 191–271. [Google Scholar]
- Giere, O. Meiobenthology: The Microscopic Motile Fauna of Aquatic Sediments, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2009; p. 527. [Google Scholar]
- Schmid-Araya, J.M.; Schmid, P.E. Trophic relationships: Integrating meiofauna into a realistic benthic food web. Freshw. Biol. 2000, 44, 149–163. [Google Scholar] [CrossRef]
- Balsamo, M.; Semprucci, F.; Frontalini, F.; Coccioni, R. Meiofauna as a tool for marine ecosystem biomonitoring. Mar. Ecosyst. 2012, 4, 77–104. [Google Scholar]
- Chen, C.A.; Soo, C.L.; Balsamo, M.; Semprucci, F. An approach based on nematode descriptors for the ecological quality (EcoQ) classification of the Malaysian coasts. Mar. Biodivers. 2018, 48, 117–126. [Google Scholar] [CrossRef]
- Semprucci, F.; Balsamo, M.; Appolloni, L.; Sandulli, R. Assessment of ecological quality status along the Apulian coasts (Eastern Mediterranean Sea) based on meiobenthic and nematode assemblages. Mar. Biodivers. 2018, 48, 105–115. [Google Scholar] [CrossRef]
- Boufahja, F.; Semprucci, F.; Beyrem, H.; Bhadury, P. Marine nematodes taxonomy in Africa: Promising prospects against scarcity of information. J. Nematic. 2015, 47, 198–206. [Google Scholar]
- Derycke, S.; Maes, S.; Van den Bulcke, L.; Vanhollebeke, J.; Wittoeck, J.; Hillewaert, H.; Ampe, B.; Haegeman, A.; Hostens, K.; De Backer, A. Optimisation of metabarcoding for monitoring marine macrobenthos: Primer choice and morphological traits determine species detection in bulkDNA and eDNA from the ethanol preservative. Front. Mar. Sci. 2021, 8, 675. [Google Scholar]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Sechi, V.; De Goede, R.G.M.; Rutgers, M.; Brussaard, L.; Mulder, C. Functional diversity in nematode communities across terrestrial ecosystems. Basic Appl. Ecol. 2018, 30, 76–86. [Google Scholar] [CrossRef]
- Chalcraft, D.R.; Resetarits, W.J. Mapping functional similarity of predators on the basis of trait similarities. Am. Nat. 2003, 162, 390–402. [Google Scholar] [CrossRef] [Green Version]
- Schratzberger, M.; Warr, K.; Rogers, S.I. Functional diversity of nematode community in the south-western North Sea. Mar. Environ. Res. 2007, 63, 368–389. [Google Scholar] [CrossRef] [Green Version]
- Wieser, W. Die beziehung zwischen mundhöhlengestalt, ernährungsweise und vorkommen bei freilebenden marinen nematoden. Eine ökologisch-morphologische studie. Ark. Zool. 1953, 4, 439–484. [Google Scholar]
- Wieser, W. Free-living Marine Nematodes IV. General Part. Acta Univ. Lund. 1959, 55, 1–111. [Google Scholar]
- Thistle, D.; Sherman, K.M. The nematode fauna of a deep-sea site exposed to strong near-bottom currents. Deep Sea Res. 1985, 32, 1077–1088. [Google Scholar] [CrossRef]
- Thistle, D.; Lambshead, P.J.D.; Sherman, K.M. Nematode tail-shape groups respond to environmental differences in the deep sea. Life Environ. 1995, 45, 107–115. [Google Scholar]
- Vanaverbeke, J.; Steyaert, M.; Vanreusel, A.; Vincx, M. Nematode biomass spectra as descriptors of functional changes due to human and natural impact. Mar. Ecol. Prog. Ser. 2003, 249, 157–170. [Google Scholar] [CrossRef]
- Losi, V.; Moreno, M.; Gaozza, L.; Vezzulli, L.; Fabiano, M.; Albertelli, G. Nematode biomass and allometric attributes as indicators of environmental quality in a Mediterranean harbour (Ligurian Sea, Italy). Ecol. Indic. 2013, 30, 80–89. [Google Scholar] [CrossRef]
- Franzo, A.; Del Negro, P. Functional diversity of free-living nematodes in river lagoons: Can biological traits analysis (BTA) integrate traditional taxonomic-based approaches as a monitoring tool? Mar. Environ. Res. 2019, 145, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Thiele, T.R.; Faumont, S.; Lockery, S.R. The neural network for chemotaxis to tastants in Caenorhabditis elegans is specialized for temporal differentiation. J. Neurosci. 2009, 29, 11904–11911. [Google Scholar] [CrossRef] [Green Version]
- Zullini, A.; Semprucci, F. Morphological differences between free-living soil and freshwater nematodes in relation to their environments. Nematology 2020, 22, 125–132. [Google Scholar] [CrossRef]
- Raes, M.; De Troch, M.; Ndaro, S.G.M.; Muthumbi, A.; Guilini, K.; Vanreusel, A. The structuring role of microhabitat type in coral degradation zones: A case study with marine nematodes from Kenya and Zanzibar. Coral Reefs. 2007, 26, 113–126. [Google Scholar] [CrossRef]
- Semprucci, F.; Cesaroni, L.; Guidi, L.; Balsamo, M. Do the morphological and functional traits of free-living marine nematodes mirror taxonomical diversity? Mar. Environ. Res. 2018, 135, 114–122. [Google Scholar] [CrossRef]
- Ichiishi, K.; Ekino, T.; Kanzaki, N.; Shinya, R. Thick cuticles as an anti-predator defense in nematodes. Nematology 2021, 24, 11–20. [Google Scholar] [CrossRef]
- Moreno, M.; Ferrero, T.J.; Gallizia, I.; Vezzulli, L.; Albertelli, G.; Fabiano, M. An assessment of the spatial heterogeneity of environmental disturbance within an enclosed harbour through the analysis of meiofauna and nematode assemblages. Estuar. Coast. Shelf. Sci. 2008, 77, 565–576. [Google Scholar] [CrossRef]
- Moreno, M.; Albertelli, G.; Fabiano, M. Nematode response to metal, PAHs and organic enrichment in tourist marinas of the Mediterranean Sea. Mar. Poll. Bull. 2009, 58, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Semprucci, F.; Boi, P.; Manti, A.; Covazzi Harriague, A.; Rocchi, M.; Colantoni, P.; Papa, S.; Balsamo, M. Benthic communities along a littoral of the Central Adriatic Sea (Italy). Helgol. Mar. Res. 2010, 64, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Semprucci, F.; Sbrocca, C.; Rocchi, M.; Balsamo, M. Temporal changes of the meiofaunal assemblage as a tool for the assessment of the ecological quality status. J. Mar. Biolog. Assoc. UK 2015, 95, 247–254. [Google Scholar] [CrossRef]
- Semprucci, F.; Frontalini, F.; Sbrocca, C.; Armynot du Châtelet, E.; Bout-Roumazeilles, V.; Coccioni, R.; Balsamo, M. Meiobenthos and free-living nematodes as tools for biomonitoring environments affected by riverine impact. Environ. Monit. Assess. 2015, 187, 251. [Google Scholar] [CrossRef] [PubMed]
- Vezzulli, L.; Moreno, M.; Marin, V.; Pezzati, E.; Bartoli, M.; Fabiano, M. Organic waste impact of capture- based Atlantic bluefin tuna aquaculture at an exposed site in the Mediterranean Sea. Estuar. Coast. Shelf. Sci. 2008, 78, 369–384. [Google Scholar] [CrossRef]
- Danovaro, R.; Gambi, C.; Mirto, S.; Sandulli, R.; Ceccherelli, V.M. Mediterranean Marine Benthos: A Manual of Methods for Its Sampling and Study; Gambi, M.C., Dappiano, M., Eds.; S.I.B.M-Società Italiana di Biologia Marina: Genova, Italy, 2004; pp. 55–97. [Google Scholar]
- Seinhorst, J.W. A rapid method for the transfer of nematodes from fixative to unhydrous glycerine. Nematologica 1959, 4, 67–69. [Google Scholar] [CrossRef] [Green Version]
- Platt, H.M.; Warwick, R.M. Free-living Marine Nematodes. Part, I. British Enoplids. In Synopses of the British Fauna (New Series); Cambridge University Press: Cambridge, UK, 1983; Volume 28, p. 307. [Google Scholar]
- Platt, H.M.; Warwick, R.M. Free-living Marine Nematodes. Part II. British Chromadorids. In Synopses of the British Fauna (New Series); Brill: Leiden, The Netherlands, 1988; Volume 38, p. 502. [Google Scholar]
- Warwick, R.M.; Platt, H.M.; Somerfield, P.J. Free-living Marine Nematodes. Part III. British Monhysterids. In Synopses of the British Fauna (New Series); Field Studies Council: Shrewsbury, UK, 1998; Volume 53, p. 296. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, MA, USA, 2006; p. 192. [Google Scholar]
- Semprucci, F.; Losi, V.; Moreno, M. A review of Italian research on free-living marine nematodes and the future perspectives on their use as Ecological Indicators (EcoInds). Mediterr. Mar. Sci. 2015, 16, 352–365. [Google Scholar] [CrossRef] [Green Version]
- Armenteros, M.; Pérez-García, J.A.; Marzo-Pérez, D.; Rodríguez-García, P. The Influential Role of the Habitat on the Diversity Patterns of Free-Living Aquatic Nematode Assemblages in the Cuban Archipelago. Diversity 2019, 11, 166. [Google Scholar] [CrossRef] [Green Version]
- Inglis, W.G. The structure of the nematode cuticle. Proc. Zool. Soc. Lond. 1964, 143, 465–502. [Google Scholar] [CrossRef]
- Mirto, S.; La Rosa, T.; Gambi, C.; Danovaro, R.; Mazzola, A. Nematode community response to fish-farm impact in the western Mediterranean. Environ. Pollut. 2002, 116, 203–214. [Google Scholar] [CrossRef]
- Schratzberger, M.; Wall, C.M.; Reynolds, W.J.; Reed, J.; Waldock, M.J. Effects of paint-derived tributyltin on structure of estuarine nematode assemblages in experimental microcosms. J. Exp. Mar. Biol. Ecol. 2002, 272, 217–235. [Google Scholar] [CrossRef]
- Boufahja, F.; Semprucci, F. Stress-induced selection of a single species from an entire meiobenthic nematode assemblage: Is this possible using iron enrichment and does pre-exposure affect the ease of the process? Environ. Sci. Pollut. Res. 2015, 22, 1979–1998. [Google Scholar] [CrossRef]
- Cibic, T.; Franzo, A.; Nasi, F.; Auriemma, R.; Del Negro, P. The port of Trieste (northern Adriatic Sea)—A case study of the “ecosystem approach to management”. Front. Mar. Sci. 2017, 4, 336. [Google Scholar] [CrossRef] [Green Version]
- Bellec, L.; Cambon-Bonavita, M.-A.; Durand, L.; Aube, J.; Gayet, N.; Sandulli, R.; Brandily, C.; Zeppilli, D. Microbial communities of the shallow-water hydrothermal vent near Naples, Italy, and chemosynthetic symbionts associated With a free-living marine nematode. Front. Microbiol. 2020, 11, 2023. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, S.; Prein, M.; Valentin, C. Mass aggregations of the free-living marine nematode Pontonema vulgare (Oncholaimidae) in organically polluted fjords. Mar. Ecol. Prog. Ser. 1987, 37, 27–34. [Google Scholar] [CrossRef]
- Thiermann, F.; Vismann, B.; Giere, O. Sulphide tolerance of the marine nematode Oncholaimus campylocercoides-a result of internal sulphur formation? Mar. Ecol. Prog. Ser. 2000, 193, 251–259. [Google Scholar] [CrossRef]
- Bargmann, C.I. Chemosensation in C. elegans; WormBook, 2006; pp. 1–29. Available online: https://www.ncbi.nlm.nih.gov/books/NBK19746/ (accessed on 13 February 2022).
- Guden, R.M.; Derycke, S.; Moens, T. A Multi-Faceted Approach to Understand How Resource Diversity Can Mediate the Coexistence of Cryptic Marine Nematode Species. Front. Mar. Sci. 2021, 8, 777425. [Google Scholar] [CrossRef]
- Wakkaf, T.; Allouche, M.; Harrath, A.H.; Mansour, L.; Alwasel, S.; Mohamed Thameemul Ansari, K.G.; Beyrem, H.; Sellami, B.; Boufahja, F. The individual and combined effects of cadmium, polyvinyl chloride (PVC) microplastics and their polyalkylamines modified forms on meiobenthic features in a microcosm. Environ. Pollut. 2020, 266, 115263. [Google Scholar] [CrossRef]
- Page, A.P.; Johnstone, I.L. The cuticle. WormBook. 2007. Available online: http://wormbook.org/chapters/www_cuticle/cuticle.pdf (accessed on 13 February 2022).
- Boufahja, F.; Semprucci, F.; Beyrem, H. An experimental protocol to select nematode species from an entire community using progressive sedimentary enrichment. Ecol. Indic. 2016, 60, 292–309. [Google Scholar] [CrossRef]
- Armenteros, M.; Ruiz-Abierno, A.; Fernández-Garcés, R.; Pérez-García, J.A.; Díaz-Asencio, L.; Vincx, M.; Decraemer, W. Biodiversity patterns of free-living marine nematodes in a tropical bay: Cienfuegos, Caribbean Sea. Estuar. Coast. Shelf Sci. 2009, 85, 179–189. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Localities of the study cases considered in the present study. Port contaminated stations: Voltri (St. I: inner port area, St. M: central port area, and St. O: outer port area) and Marina degli Aregai (four stations from the inner, St. 1, to outer port part, St. 4); coastal sediments influenced by riverine inputs (St. B and St. FI: Natural Regional Park; St. M and St. FO: Foglia and Metauro rivers); fish farm of Olbia (Og: under cages, Oc: controls) and tuna farming of Vibo Marina (Vg: under cages, Vc: controls).
Figure 1.
Localities of the study cases considered in the present study. Port contaminated stations: Voltri (St. I: inner port area, St. M: central port area, and St. O: outer port area) and Marina degli Aregai (four stations from the inner, St. 1, to outer port part, St. 4); coastal sediments influenced by riverine inputs (St. B and St. FI: Natural Regional Park; St. M and St. FO: Foglia and Metauro rivers); fish farm of Olbia (Og: under cages, Oc: controls) and tuna farming of Vibo Marina (Vg: under cages, Vc: controls).

Figure 2.
The amphids, chemosensory organs located in the head region of nematodes, play a role in the search for food and partners. Eight categories were identified on the base of the amphidal fovea shape.
Figure 2.
The amphids, chemosensory organs located in the head region of nematodes, play a role in the search for food and partners. Eight categories were identified on the base of the amphidal fovea shape.

Figure 3.
Cuticle has a role in defense against predators, pollution, and hydrodynamic energy and in nematode locomotion. Five categories of cuticles were recognized on the base of their morphology and thickness.
Figure 3.
Cuticle has a role in defense against predators, pollution, and hydrodynamic energy and in nematode locomotion. Five categories of cuticles were recognized on the base of their morphology and thickness.

Figure 4.
Buccal cavity morphologies reflect the trophic role of the species in the ecosystem: selective (1A) and non-selective (1B) deposit feeders, epistratum feeders (2A), and predators/omnivores (2B).
Figure 4.
Buccal cavity morphologies reflect the trophic role of the species in the ecosystem: selective (1A) and non-selective (1B) deposit feeders, epistratum feeders (2A), and predators/omnivores (2B).

Figure 5.
Tail shape has an important role in nematode locomotion, feeding, and reproduction. Four categories were designed: short/round; elongated/filiform; conical; and clavate/conical-cylindrical.
Figure 5.
Tail shape has an important role in nematode locomotion, feeding, and reproduction. Four categories were designed: short/round; elongated/filiform; conical; and clavate/conical-cylindrical.

Figure 6.
Assignment of each genus in the four morpho-functional traits and creation of the data matrix based on the combination of morpho-functional traits.
Figure 6.
Assignment of each genus in the four morpho-functional traits and creation of the data matrix based on the combination of morpho-functional traits.

Figure 7.
nMDS carried out on untransformed data matrixes of both nematode genera and functional trait codes (Bray–Curtis similarity) from port seabeds (Ligurian Sea) and sediments under riverine outfall pressures (Adriatic Sea) (Ja = January; Fe = February; Jn = June; Jl = July; No = November).
Figure 7.
nMDS carried out on untransformed data matrixes of both nematode genera and functional trait codes (Bray–Curtis similarity) from port seabeds (Ligurian Sea) and sediments under riverine outfall pressures (Adriatic Sea) (Ja = January; Fe = February; Jn = June; Jl = July; No = November).

Figure 8.
nMDS carried out on untransformed data matrixes of both nematode genera and functional trait codes (Bray–Curtis similarity) from fish farm sediments (Tyrrhenian Sea) (Ap = April; Ma = May; Sp = September; Oc = October).
Figure 8.
nMDS carried out on untransformed data matrixes of both nematode genera and functional trait codes (Bray–Curtis similarity) from fish farm sediments (Tyrrhenian Sea) (Ap = April; Ma = May; Sp = September; Oc = October).

Figure 9.
Trends of the Shannon diversity (H’) and Pielou evenness (J’) computed on the matrixes based on genus and combination of morpho-functional traits for each study case.
Figure 9.
Trends of the Shannon diversity (H’) and Pielou evenness (J’) computed on the matrixes based on genus and combination of morpho-functional traits for each study case.

Figure 10.
Combination of morpho-functional traits in nematodes inhabiting sediments under human pressure.
Figure 10.
Combination of morpho-functional traits in nematodes inhabiting sediments under human pressure.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of Analysis of Similarities (ANOSIM) performed to detect the presence of significant differences among the stations and sampling periods of each study case (n.s. indicates when significant differences were not found).
Table 1.
Results of Analysis of Similarities (ANOSIM) performed to detect the presence of significant differences among the stations and sampling periods of each study case (n.s. indicates when significant differences were not found).
| Study Case | Structure at Genus Level | Structure at Morpho-Functional Level | ||
|---|---|---|---|---|
| Sample Statistic | Pairwise Tests | Sample Statistic | Pairwise Tests | |
| Genoa Voltri port | Stations R = 0.80; p = 0.001 | St. I vs. St. O: R = 0.99; p = 0.03 | Stations R = 0.78; p = 0.001 | St. I vs. St. O: R = 0,99; p = 0.03 |
| St. I vs. St. M: R = 0.65; p = 0.03 | St. I vs. St. M: R = 0.60; p = 0.03 | |||
| St. O vs. St. M: R = 0.75; p = 0.03 | St. O vs. St. M: R = 0.70; p = 0.03 | |||
| Periods p = n.s. | Periods p = n.s. | |||
| Aregai marina | Stations: p = n.s. | Stations p = n.s. | ||
| Periods R = 0.81; p = 0.029 | Periods p = n.s. | |||
| Riverine outfall areas | Stations R = 0.38; p = 0.001 | F vs. B: R = 0.26; p = 0.05 | Stations R = 0.35; p = 0.001 | F vs. B: R = 0.25; p = 0.05 |
| F vs. FO: R = 0.60; p = 0.002 | F vs. FO: R = 0.59; p = 0.004 | |||
| F vs. M: R = 0.48; p = 0.002 | F vs. M: R = 0.34; p = 0.004 | |||
| B vs. FO: R = 0.67; p = 0.002 | B vs. FO: R = 0.70; p = 0.002 | |||
| B vs. M: R = 0.30; p = 0.017 | B vs. M: R = 0.28; p = 0.017 | |||
| FO vs. M: n.s. | FO vs. M: n.s. | |||
| Periods R = 0.24; p = 0.001 | Periods R = 0.17; p = 0.016 | |||
| Olbia Fish-farm | Stations p = n.s. | Stations p = n.s. | ||
| Periods p = n.s. | Periods p = n.s. | |||
| Vibo Fish-farm | Stations p = n.s. | Stations p = n.s. | ||
| Periods p = n.s. | Periods p = n.s. | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Semprucci, F.; Grassi, E.; Balsamo, M. Simple Is the Best: An Alternative Method for the Analysis of Free-Living Nematode Assemblage Structure. Water 2022, 14, 1114. https://doi.org/10.3390/w14071114
AMA Style
Semprucci F, Grassi E, Balsamo M. Simple Is the Best: An Alternative Method for the Analysis of Free-Living Nematode Assemblage Structure. Water. 2022; 14(7):1114. https://doi.org/10.3390/w14071114
Chicago/Turabian StyleSemprucci, Federica, Eleonora Grassi, and Maria Balsamo. 2022. "Simple Is the Best: An Alternative Method for the Analysis of Free-Living Nematode Assemblage Structure" Water 14, no. 7: 1114. https://doi.org/10.3390/w14071114
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.