The Impacts of the Freezing–Thawing Process on Benthic Macroinvertebrate Communities in Riffles and Pools: A Case Study of China’s Glacier-Fed Stream

Abstract
:1. Introduction
2. Materials and Methods
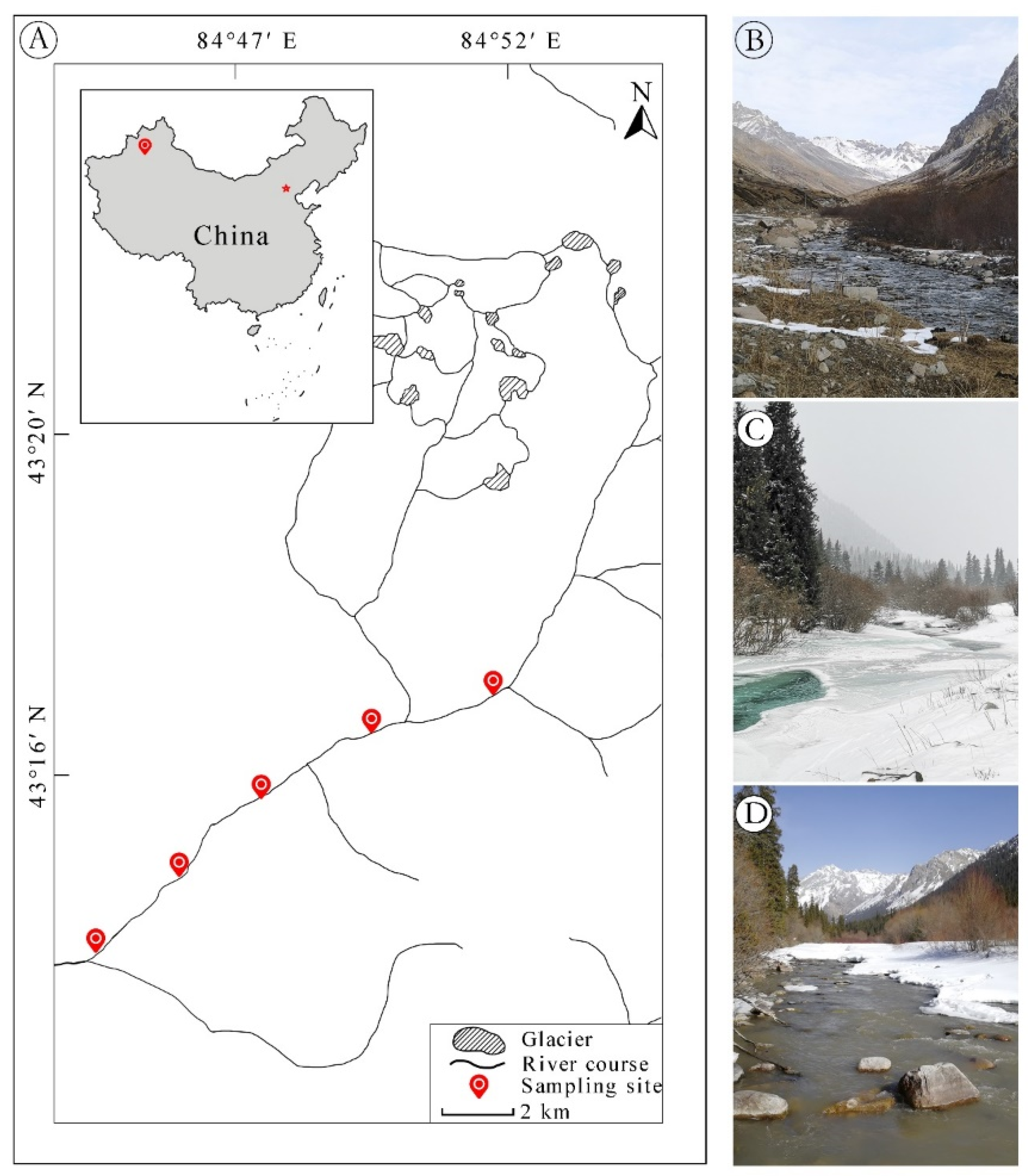
2.1. Study Area
2.2. Benthic Macroinvertebrate Collection
2.3. Environmental Characterization
2.4. Functional Traits
2.5. Data Analysis
3. Results
3.1. Environmental Characteristics
3.2. Indicator Taxa in the Riffle–Pool Habitat
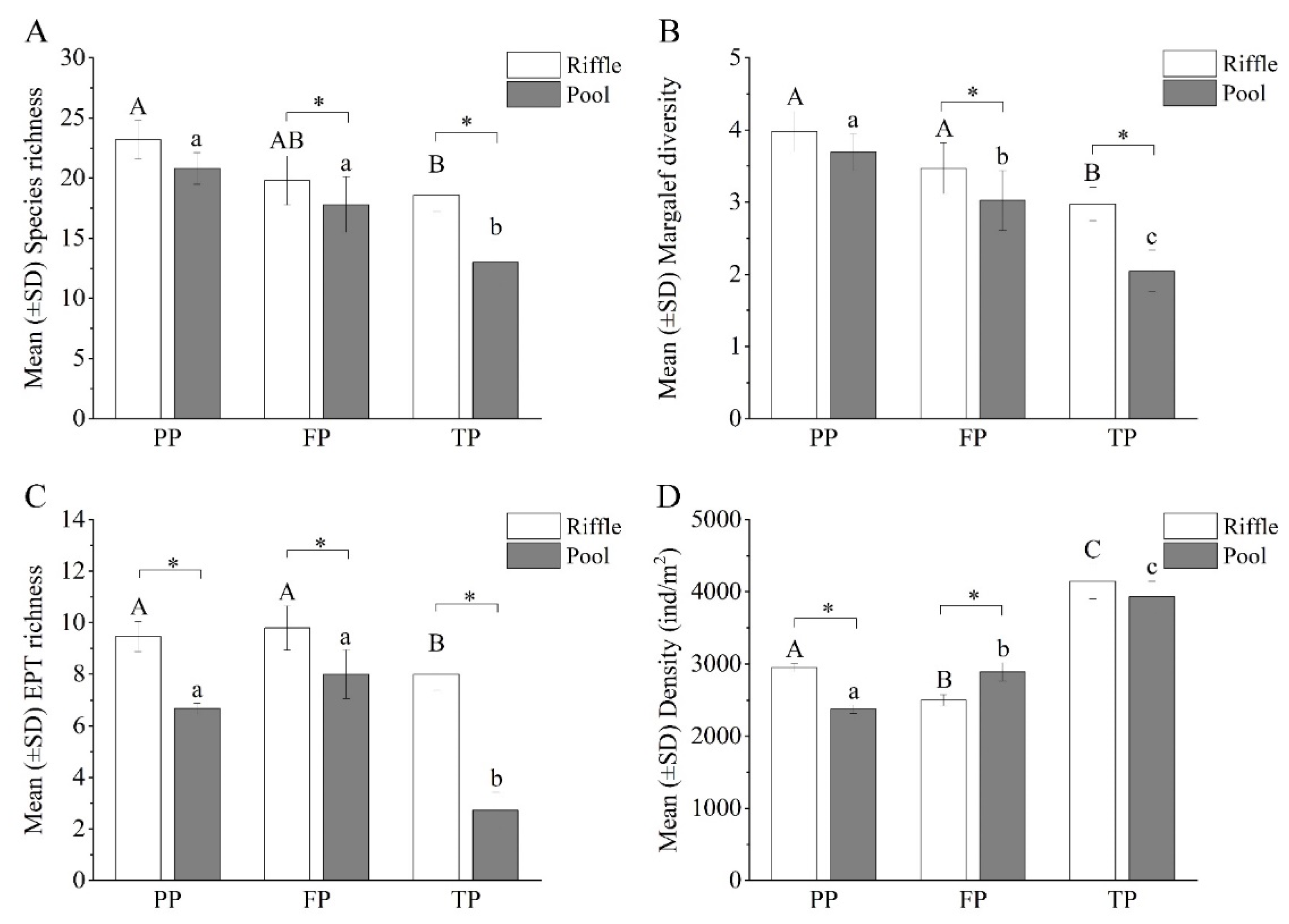
3.3. Benthic Macroinvertebrate Community Structure in the Riffle–Pool Habitat
3.4. Functional Traits of Benthic Macroinvertebrates
3.5. Relationships between Benthic Macroinvertebrate Communities and Environmental Factors
4. Discussion
4.1. Habitat Characteristics of Riffles and Pools during the Freezing–Thawing Process
4.2. Insights on Indicator Taxa
4.3. Benthic Macroinvertebrate Community Structure in Riffles and Pools during the Freezing–Thawing Process
4.4. Benthic Macroinvertebrate Functional Traits in Riffles and Pools during the Freezing–Thawing Process
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Trait State | Trait | Trait State |
|---|---|---|---|
| Life history | Ecology | ||
| Voltinism | Semi-voltine | Rheophily | Depositional only |
| Uni-voltine | Depositional and erosional | ||
| Bi- or multi-voltine | Erosional | ||
| Mobility | Thermal preference | Cool eurythermal | |
| Occurrence in drift | Rare | Cool/warm eurythermal | |
| Common | Warm eurythermal | ||
| Abundant | Habit | Burrower | |
| Swimming ability | None | Climber | |
| Weak | Sprawler | ||
| Strong | Clinger | ||
| Morphology | Swimmer | ||
| Attachment | None | Trophic habit | Collector-gatherer |
| Some | Collector-filterer | ||
| Both | Herbivore | ||
| Shape | Streamlined | Predator | |
| Not streamlined | Shredder | ||
| Size at maturity | Small (<9 mm) | ||
| Medium (9−16 mm) | |||
| Large (>16 mm) |
| Code | Taxa | Pre-Freezing Period | Freezing Period | Thawing Period | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Riffle | Pool | Riffle | Pool | Riffle | Pool | ||||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | mean | SD | Mean | SD | ||
| S1 | Planaria sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 4 |
| S2 | Naididae sp. | 10 | 2 | 11 | 7 | 12 | 5 | 16 | 7 | 2 | 2 | 27 | 15 |
| S3 | Anis sp. | 0 | 0 | 0 | 0 | 0 | 0 | 8 | 9 | 0 | 0 | 0 | 0 |
| S4 | Gammarus sp. | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 34 | 19 |
| S5 | Rhithrogena sp. | 384 | 27 | 87 | 17 | 127 | 34 | 47 | 4 | 24 | 6 | 0 | 0 |
| S6 | Iron sp. | 24 | 10 | 4 | 1 | 0 | 0 | 0 | 0 | 366 | 60 | 7 | 4 |
| S7 | Heptagenia sp. | 0 | 0 | 0 | 0 | 10 | 6 | 0 | 0 | 0 | 0 | 0 | 0 |
| S8 | Epeorus sp. | 0 | 0 | 0 | 0 | 23 | 5 | 7 | 1 | 0 | 0 | 0 | 0 |
| S9 | Cinygmula sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 41 | 23 | 0 | 0 |
| S10 | Baetis sp. | 387 | 25 | 84 | 15 | 656 | 32 | 232 | 20 | 1422 | 103 | 4 | 9 |
| S11 | Ameletus sp. | 40 | 8 | 33 | 13 | 185 | 22 | 23 | 8 | 339 | 57 | 8 | 4 |
| S12 | Isoperla sp. | 27 | 4 | 16 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S13 | Perlodes sp. | 0 | 0 | 0 | 0 | 22 | 2 | 7 | 6 | 0 | 0 | 0 | 0 |
| S14 | Starsolus sp. | 12 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S15 | Nemoura sp. | 13 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 693 | 27 | 13 | 9 |
| S16 | Amphinemura sp. | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 7 | 0 | 0 | 0 | 0 |
| S17 | Capnia sp. | 0 | 0 | 0 | 0 | 333 | 44 | 32 | 10 | 34 | 22 | 0 | 0 |
| S18 | Suwallia sp. | 0 | 0 | 0 | 0 | 79 | 13 | 0 | 0 | 0 | 0 | 0 | 0 |
| S19 | Haliplus sp. | 0 | 0 | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S20 | Homoplectra sp. | 5 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S21 | Glossosoma sp. | 220 | 15 | 143 | 25 | 17 | 10 | 0 | 0 | 0 | 0 | 0 | 0 |
| S22 | Theliopsyche sp. | 82 | 14 | 14 | 5 | 0 | 0 | 7 | 4 | 0 | 0 | 0 | 0 |
| S23 | Brachycentrus sp. | 345 | 23 | 96 | 25 | 12 | 5 | 12 | 4 | 74 | 24 | 7 | 6 |
| S24 | Amiocentrus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 16 | 7 | 0 | 0 | 0 | 0 |
| S25 | Rhyacophila sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 |
| S26 | Desmona sp. | 0 | 0 | 0 | 0 | 27 | 8 | 25 | 7 | 0 | 0 | 0 | 0 |
| S27 | Pseudostenophylax sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 15 | 15 | 0 | 0 |
| S28 | Dicranota sp. | 35 | 4 | 74 | 9 | 59 | 9 | 70 | 13 | 12 | 8 | 15 | 9 |
| S29 | Cheilotrichia sp. | 9 | 4 | 7 | 3 | 11 | 5 | 58 | 8 | 7 | 9 | 0 | 0 |
| S30 | Tipula (Arctotipula) sp. | 8 | 3 | 18 | 12 | 7 | 3 | 4 | 0 | 0 | 0 | 19 | 11 |
| S31 | Tipula (Sinotipula) sp. | 5 | 2 | 5 | 7 | 0 | 0 | 19 | 3 | 0 | 0 | 0 | 0 |
| S32 | Hexatoma sp. | 21 | 11 | 16 | 7 | 0 | 0 | 0 | 0 | 22 | 14 | 0 | 0 |
| S33 | Antocha sp. | 73 | 25 | 25 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S34 | Tanytarsini sp. | 216 | 24 | 1198 | 64 | 0 | 0 | 0 | 0 | 206 | 47 | 2526 | 139 |
| S35 | Chironomus sp. | 0 | 0 | 16 | 6 | 519 | 51 | 1454 | 39 | 22 | 15 | 550 | 61 |
| S36 | Orthocladiinae sp.1 | 26 | 7 | 21 | 9 | 246 | 35 | 776 | 72 | 295 | 31 | 59 | 8 |
| S37 | Orthocladiinae sp.2 | 12 | 1 | 219 | 23 | 0 | 0 | 0 | 0 | 72 | 21 | 603 | 66 |
| S38 | Orthocladiinae sp.3 | 257 | 22 | 90 | 17 | 0 | 0 | 0 | 0 | 438 | 66 | 24 | 12 |
| S39 | Tanypodinae sp. | 3 | 4 | 77 | 10 | 17 | 7 | 37 | 8 | 0 | 0 | 0 | 0 |
| S40 | Sphaeromias sp. | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 16 | 14 |
| S41 | Psychodidae sp. | 593 | 34 | 94 | 23 | 0 | 0 | 4 | 0 | 17 | 8 | 0 | 0 |
| S42 | Blefariceridae sp. | 61 | 9 | 8 | 7 | 10 | 6 | 8 | 9 | 0 | 0 | 0 | 0 |
| S43 | Tabanidae sp.1 | 79 | 18 | 13 | 4 | 20 | 5 | 23 | 8 | 17 | 4 | 7 | 5 |
| S44 | Tabanidae sp.2 | 0 | 0 | 0 | 0 | 8 | 9 | 0 | 0 | 7 | 8 | 4 | 1 |
| S45 | Blepharicera sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 24 | 11 | 0 | 0 |
| S46 | Hydrachnellae sp. | 0 | 0 | 0 | 0 | 99 | 23 | 10 | 3 | 0 | 0 | 0 | 0 |
References
- Milner, A.M.; Khamis, K.; Battin, T.J.; Brittain, J.E.; Barrand, N.E.; Füreder, L.; Cauvy-Fraunié, S.; Gíslason, G.M.; Jacobsen, D.; Hannah, D.M.; et al. Glacier shrinkage driving global changes in downstream systems. Proc. Natl. Acad. Sci. USA 2017, 114, 9770–9778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scotti, A.; Jacobsen, D.; Tappeiner, U.; Bottarin, R. Spatial and temporal variation of benthic macroinvertebrate assemblages during the glacial melt season in an Italian glacier-fed stream. Hydrobiologia 2019, 827, 123–139. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Pérez, V.; Dangles, O.; Ibarra, C.; Espinosa, R.; Andino, P.; Jacobsen, D.; Cauvy-Fraunié, S. Functional structure and diversity of invertebrate communities in a glacierised catchment of the tropical Andes. Freshw. Biol. 2020, 65, 1348–1362. [Google Scholar] [CrossRef]
- Jacobsen, D.; Dangles, O.; Andino, P.; Espinosa, R.; Hamerlík, L.; Cadier, E. Longitudinal zonation of macroinvertebrates in an Ecuadorian glacier–fed stream: Do tropical glacial systems fit the temperate model? Freshw. Biol. 2010, 55, 1234–1248. [Google Scholar] [CrossRef]
- Stahl, K.; Moore, R.D.; Shea, J.M.; Hutchinson, D.; Cannon, A.J. Coupled modeling of glacier and streamflow response to future climate scenarios. Water Resour. Res. 2008, 44, W02422. [Google Scholar] [CrossRef] [Green Version]
- Cauvy-Fraunié, S.; Dangles, O. A global synthesis of biodiversity responses to glacier retreat. Nat. Ecol. Evol. 2019, 3, 1675–1685. [Google Scholar] [CrossRef]
- Espinosa, R.; Andino, P.; Cauvy-Fraunié, S.; Dangles, O.; Jacobsen, D.; Crespo-Pérez, V. Diversity patterns of aquatic macroinvertebrates in a tropical high-Andean catchment. Rev. Biol. Trop. 2020, 68, S29–S53. [Google Scholar] [CrossRef]
- Shain, D.H.; Novis, P.M.; Cridge, A.G.; Zawierucha, K.; Geneva, A.J.; Dearden, P.K. Five animal phyla in glacier ice reveal unprecedented biodiversity in New Zealand’s Southern Alps. Sci. Rep. 2021, 11, 3898. [Google Scholar] [CrossRef]
- Sponseller, R.A.; Grimm, N.B.; Boulton, A.J.; Sabo, J.L. Responses of macroinvertebrate communities to long–term flow variability in a Sonoran Desert stream. Glob. Change Biol. 2010, 16, 2891–2900. [Google Scholar] [CrossRef] [Green Version]
- Cauvy–Fraunié, S.; Espinosa, R.; Andino, P.; Dangles, O.; Jacobsen, D. Relationships between stream macroinvertebrate communities and new flood–based indices of glacial influence. Freshw. Biol. 2014, 59, 1916–1925. [Google Scholar] [CrossRef]
- Chanut, P.C.; Datry, T.; Gabbud, C.; Robinson, C.T. Direct and indirect effects of flood regime on macroinvertebrate assemblages in a floodplain riverscape. Ecohydrology 2019, 12, 2095. [Google Scholar] [CrossRef]
- Danehy, R.J.; Bilby, R.E.; Justice, T.E.; Lester, G.T.; Jones, J.E.; Haddadi, S.S.; Merritt, G.D. Aquatic Biological Diversity Responses to Flood Disturbance and Forest Management in Small, Forested Watersheds. Water 2021, 13, 2793. [Google Scholar] [CrossRef]
- Wallace, J.B.; Webster, J.R. The role of macroinvertebrates in stream ecosystem function. Annu. Rev. Entomol. 1996, 41, 115–139. [Google Scholar] [CrossRef]
- Docherty, C.L.; Hannah, D.M.; Riis, T.; Lund, M.; Abermann, J.; Milner, A.M. Spatio-temporal dynamics of macroinvertebrate communities in northeast Greenlandic snowmelt streams. Ecohydrology 2018, 11, e1982. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.E.; Khamis, K.; Wilkes, M.; Blaen, P.; Brittain, J.E.; Carrivick, J.L.; Fell, S.; Friberg, N.; Füreder, L.; Gislason, G.M.; et al. Functional diversity and community assembly of river invertebrates show globally consistent responses to decreasing glacier cover. Nat. Ecol. Evol. 2018, 2, 325–333. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Functional diversity (FD), species richness and community composition. Ecol. Lett. 2002, 5, 402–411. [Google Scholar] [CrossRef]
- Usseglio-Polatera, P.; Bournaud, M.; Richoux, P.; Tachet, H. Biological and ecological traits of benthic freshwater macroinvertebrates: Relationships and definition of groups with similar traits. Freshw. Biol. 2000, 43, 175–205. [Google Scholar] [CrossRef]
- Poff, N.L.; Olden, J.D.; Vieira, N.K.M.; Finn, D.S.; Simmons, M.P.; Kondratieff, B.C. Functional trait niches of North American lotic insects: Traits-based ecological applications in light of phylogenetic relationships. J. N. Am. Benthol. Soc. 2006, 25, 730–755. [Google Scholar] [CrossRef] [Green Version]
- Buffagni, A.; Erba, S.; Cazzola, M.; Barca, E.; Belfiore, C. The ratio of lentic to lotic habitat features strongly affects macroinvertebrate metrics used in southern Europe for ecological status classification. Ecol. Indic. 2020, 117, 106563. [Google Scholar] [CrossRef]
- Hawkins, C.P.; Kershner, J.L.; Bisson, P.A.; Bryant, M.D.; Decker, L.M.; Gregory, S.V.; McCullough, D.A.; Overton, C.K.; Reeves, G.H.; Steedman, R.J.; et al. A hierarchical approach to classifying stream habitat features. Fisheries 1993, 18, 3–12. [Google Scholar] [CrossRef]
- Jowett, I.G. A method for objectively identifying pool, run, and riffle habitats from physical measurements. N. Z. J. Mar. Freshw. Res. 1993, 27, 241–248. [Google Scholar] [CrossRef]
- Bonada, N.; Rieradevall, M.; Prat, N.; Resh, V.H. Benthic macroinvertebrate assemblages and macrohabitat connectivity in Mediterranean–climate streams of northern California. J. N. Am. Benthol. Soc. 2006, 25, 32–43. [Google Scholar] [CrossRef]
- Herbst, D.B.; Cooper, S.D.; Medhurst, R.B.; Wiseman, S.W.; Hunsaker, C.T. A comparison of the taxonomic and trait structure of macroinvertebrate communities between the riffles and pools of montane headwater streams. Hydrobiologia 2018, 820, 115–133. [Google Scholar] [CrossRef]
- Zhu, P.H.; Pan, B.Z.; Li, Z.W.; Yuan, J.P.; Jiang, X.M.; Jiang, W.X.; Lu, Y. Responses of macroinvertebrate functional traits to riverbed structure of typical debris flow gullies in the upper reaches of the Yangtze River, China. Sci. Total Environ. 2020, 738, 138906. [Google Scholar] [CrossRef] [PubMed]
- Bogan, M.T.; Boersma, K.S.; Lytle, D.A. Resistance and resilience of invertebrate communities to seasonal and supraseasonal drought in aridland headwater streams. Freshw. Biol. 2015, 60, 2547–2558. [Google Scholar] [CrossRef]
- Liu, S.Y.; Guo, W.Q.; Xu, J.L. The Second Glacier Inventory Dataset of China (Version 1.0) (2006–2011); National Tibetan Plateau Data Center: Beijing, China, 2012. [Google Scholar] [CrossRef]
- Cummins, K.W. An evaluation of some techniques for the collection and analysis of benthic samples with special emphasis on lotic waters. Am. Midl. Nat. 1962, 67, 477–504. [Google Scholar] [CrossRef]
- Fu, L.; Jiang, Y.; Ding, J.; Liu, Q.; Peng, Q.Z.; Kang, M.Y. Impacts of land use and environmental factors on macroinvertebrate functional feeding groups in the Dongjiang River basin, southeast China. J. Freshw. Ecol. 2016, 31, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Moretti, M.; Dias, A.T.C.; Bello, D.F.; Altermatt, F.; Chown, S.L.; Azcárate, F.M.; Bell, J.R.; Fournier, B.; Hedde, M.; Hortal, J.; et al. Handbook of protocols for standardized measurement of terrestrial invertebrate functional traits. Funct. Ecol. 2017, 31, 558–567. [Google Scholar] [CrossRef]
- Dai, X.; Page, B.; Duffy, K.J. Indicator value analysis as a group prediction technique in community classification. S. Afr. J. Bot. 2006, 72, 589–596. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Yuan, X.Z.; Liu, H. Comparison study on macroinvertebrate assemblage of riffles and pools: A case study of Dong River in Kaixian County of Chongqing, China. Acta Ecol. Sin. 2012, 32, 6726–6736. [Google Scholar] [CrossRef] [Green Version]
- Carmignani, J.R.; Roy, A.H. Ecological impacts of winter water level drawdowns on lake littoral zones: A review. Aquat. Sci. 2017, 79, 803–824. [Google Scholar] [CrossRef] [Green Version]
- Füreder, L.; Niedrist, G.H. Glacial Stream Ecology: Structural and Functional Assets. Water 2020, 12, 376. [Google Scholar] [CrossRef] [Green Version]
- Feng, F.; Feng, Q.; Liu, X.D.; Li, Z.Q.; Liu, W.; Jin, S. A study of hydrochemical characteristics of meltwater runoff of the Urumqi glacier No. 1 Tianshan Mountains. J. Glaciol. Geocryol. 2014, 36, 183–191. [Google Scholar]
- Xu, M.; Zhao, N.; Zhou, X.; Pan, B.; Liu, W.; Tian, S.; Wang, Z. Macroinvertebrate Biodiversity Trends and Habitat Relationships within Headwater Rivers of the Qinghai–Tibet Plateau. Water 2018, 10, 1214. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.H.; Wang, Z.Y.; Cheng, D.S. Benthic macroinvertebrate communities and biodiversity in various stream substrata. Acta Ecol. Sin. 2007, 27, 1165–1172. [Google Scholar] [CrossRef]
- Gayraud, S.; Philippe, M. Does subsurface interstitial space influence general features and morphological traits of the benthic macroinvertebrate community in streams? Arch. Hydrobiol. 2001, 151, 667–686. [Google Scholar] [CrossRef] [Green Version]
- Lindenmayer, D.B.; Margules, C.R.; Botkin, D.B. Indicators of biodiversity for ecologically sustainable forest management. Conserv. Biol. 2000, 14, 941–950. [Google Scholar] [CrossRef]
- Milner, A.M.; Brittain, J.E.; Castella, E.; Petts, G.E. Trends of macroinvertebrate community structure in glacier-fed rivers in relation to environmental conditions: A synthesis. Freshw. Biol. 2001, 46, 1833–1847. [Google Scholar] [CrossRef]
- Chin, K.S.; Lento, J.; Culp, J.M.; Lacelle, D.; Kokelj, S.V. Permafrost thaw and intense thermokarst activity decreases abundance of stream benthic macroinvertebrates. Glob. Change Biol. 2016, 22, 2715–2728. [Google Scholar] [CrossRef]
- Lencioni, V. Glacial influence and stream macroinvertebrate biodiversity under climate change: Lessons from the Southern Alps. Sci. Total Environ. 2018, 622–623, 563–575. [Google Scholar] [CrossRef]
- Tanchet, H.; Richoux, P.; Bournaud, M.; Usseglio-Polatera, P. Invertébrés d’eau douce—Systématique, Biologie, écologie; Cnrs Editions: Paris, France, 2010; pp. 345–383. [Google Scholar]
- Hynes, H.B.N. Biogeography and origins of the North American stoneflies (Plecoptera). Mem. Entomol. Soc. Can. 1988, 144, 31–37. [Google Scholar] [CrossRef]
- Chen, Z.T. Two new species of Nemouridae (Insecta: Plecoptera) from China. Eur. J. Taxon. 2020, 651, 1–16. [Google Scholar] [CrossRef]
- Tian, L.X.; Yang, L.F.; Li, Y.W. Economic Entomology of China–Trichoptera (1): Hydroptilidae, Stenopsychidae, Hydropsychidae, Leptocerdae; Science Press: Beijing, China, 1996; pp. 15–38. [Google Scholar]
- Stevenson, R.J.; Bothwell, M.; Lowe, R.L. Algal Ecology: Freshwater Benthic Ecosystems. J. N. Am. Benthol. Soc. 1997, 16, 297–299. [Google Scholar] [CrossRef]
- Burmeister, E.G. Der Massenflug von Brach ycentrus subnubilus Curtis, 1834 in Passau im Mai 1990 (insecta, Trichoptera, Brachycentridae). Nachr. Bayer. Entomol. 1991, 40, 20–23. [Google Scholar]
- Burgherr, P.; Ward, J.V.; Robinson, C.T. Seasonal variation in zoobenthos across habitat gradients in an alpine glacial floodplain (Val Roseg, Swiss Alps). J. N. Am. Benthol. Soc. 2002, 21, 561–575. [Google Scholar] [CrossRef]
- Miserendino, M.L.; Brand, C.; Epele, L.B.; Prinzio, C.Y.D.; Omad, G.H.; Archangelsky, M.; Martínez, O.; Kutschker, A.M. Biotic diversity of benthic macroinvertebrates at contrasting glacier–fed systems in Patagonia Mountains: The role of environmental heterogeneity facing global warming. Sci. Total Environ. 2018, 622–623, 152–163. [Google Scholar] [CrossRef]
- Southwood, T.R.E.; Comins, H.N. A synoptic population model. J. Anim. Ecol. 1976, 45, 949–965. [Google Scholar] [CrossRef]
- Armitage, P.D.; MacHale, A.M.; Crisp, D.C. A survey of stream invertebrates in the Cow Green basin (Upper Teesdale) before inundation. Freshw. Biol. 1974, 4, 369–398. [Google Scholar] [CrossRef]
- Beisel, J.N.; Philippe, U.P.; Thomas, S.; Moreteau, J.C. Stream community structure in relation to spatial variation: The influence of mesohabitat characteristics. Hydrobiologia 1998, 389, 73–88. [Google Scholar] [CrossRef]
- Liu, X.J.; Ma, K.P. Advances in Functional Traits in Plants. Sci. China Life Sci. 2015, 45, 325–339. [Google Scholar]
- Menezes, S.; Baird, D.J.; Soares, A. Beyond taxonomy: A review of macroinvertebrate trait-based community descriptors as tools for freshwater biomonitoring. J. Appl. Ecol. 2010, 47, 711–719. [Google Scholar] [CrossRef]
- Cauvy-Fraunié, S.; Espinosa, R.; Andino, P.; Jacobsen, D.; Dangles, O. Invertebrate metacommunity structure and dynamics in an Andean glacial stream network facing climate change. PLoS ONE 2015, 10, e0136793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cauvy-Fraunié, S.; Andino, P.; Espinosa, R.; Calvez, R.; Jacobsen, D.; Dangles, O. Ecological responses to experimental glacier–runoff reduction in alpine rivers. Nat. Commun. 2016, 7, 12025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, E.J. Seasonal distribution and life histories of Ephemeroptera in a North umbrian river. Freshw. Biol. 1980, 10, 101–111. [Google Scholar] [CrossRef]
- Vannote, R.L.; Sweeney, B.W. Geographic analysis of thermal equilibria: A conceptual model for evaluating the effect of natural and modified thermal regimes on aquatic insect communities. Am. Nat. 1980, 115, 667–695. [Google Scholar] [CrossRef] [Green Version]
- Saito, V.S.; Soininen, J.; Fonseca-Gessner, A.A.; Siqueira, T. Dispersal traits drive the phylogenetic distance decay of similarity in Neotropical stream metacommunities. J. Biogeogr. 2015, 42, 2101–2111. [Google Scholar] [CrossRef]
- Hill, M.J.; Milner, V.S. Ponding in intermittent streams: A refuge for lotic taxa and a habitat for newly colonising taxa? Sci. Total Environ. 2018, 628–629, 1308–1316. [Google Scholar] [CrossRef]
- Wang, L.; Gao, Y.Z.; Han, B.P.; Fan, H.J.; Yang, H.J. The impacts of agriculture on macroinvertebrate communities: From structural changes to functional changes in Asia’s cold region streams. Sci. Total Environ. 2019, 676, 155–164. [Google Scholar] [CrossRef]
- Füreder, L. Life at the edge: Habitat condition and bottom fauna of alpine running waters. Int. Rev. Hydrobiol. 2007, 92, 491–513. [Google Scholar] [CrossRef]
- Cauvy-Fraunié, S.; Andino, P.; Espinosa, R.; Calvez, R.; Anthelme, F.; Jacobsen, D.; Dangles, O. Glacial flood pulse effects on benthic fauna in equatorial high-Andean streams. Hydrol. Processes 2013, 28, 3008–3017. [Google Scholar] [CrossRef]
- Bêche, L.A.; Mcelravy, E.P.; Resh, V.H. Long-term seasonal variation in the biological traits of benthic-macroinvertebrates in two Mediterranean-climate streams in California, U.S.A. Freshw. Biol. 2006, 51, 56–75. [Google Scholar] [CrossRef]
- O’Dowd, A.P.; Chin, A. Do bio–physical attributes of steps and pools differ in high–gradient mountain streams? Hydrobiologia 2016, 776, 67–83. [Google Scholar] [CrossRef]
- Cummins, K.W.; Wilzbach, M.A.; Gates, D.M.; Perry, J.B.; Taliaferro, W.B. Shredders and riparian vegetation. Bioscience 1989, 39, 24–30. [Google Scholar] [CrossRef]
- Jiang, W.X.; Cai, Q.H.; Tang, T.; Qu, X.D. The functional feeding group ecology of macroinvertebrate in Xiangxi River system. Acta Ecol. Sin. 2009, 29, 5207–5218. [Google Scholar] [CrossRef]
- Bogan, M.T.; Lytle, D.A. Seasonal flow variation allows ‘time–sharing’ by disparate aquatic invertebrate communities in montane desert streams. Freshw. Biol. 2007, 52, 290–304. [Google Scholar] [CrossRef]
- Rosi–Marshall, E.J.; Vallis, K.L.; Baxter, C.V.; Davis, J.M. Retesting a prediction of the River Coninuum Concept: Authochthonous versus allochthonous resources in the diets of invertebrates. Freshw. Sci. 2016, 35, 534–543. [Google Scholar] [CrossRef]
- Rosset, V.; Ruhi, A.; Bogan, M.T.; Datry, T. Do lentic and lotic communities respond similarly to drying? Ecosphere 2017, 8, e01809. [Google Scholar] [CrossRef] [Green Version]




| Parameters | Pre-Freezing Period | Freezing Period | Thawing Period | |||
|---|---|---|---|---|---|---|
| Riffles | Pools | Riffles | Pools | Riffles | Pools | |
| Velocity (m·s−1) | 0.35 ± 0.10 A | 0.20 ± 0.05 a | 0.20 ± 0.01 B | 0.18 ± 0.00 a | 0.50 ± 0.03 C | 0.30 ± 0.04 b |
| Depth (m) | 0.26 ± 0.03 A | 0.39 ± 0.05 a | 0.10 ± 0.03 B | 0.31 ± 0.02 b | 0.30 ± 0.05 A | 0.41 ± 0.04 a |
| Width (m) | 4.25 ± 0.30 A | 5.18 ± 0.60 ab | 4.19 ± 0.28 A | 5.01 ± 0.53 a | 5.48 ± 0.25 B | 5.71 ± 0.16 b |
| Temperature (°C) | 1.74 ± 0.04 A | 1.75 ± 0.04 a | 0.35 ± 0.11 B | 0.36 ± 0.12 b | 4.77 ± 0.09 C | 4.27 ± 0.05 c |
| Dissolved oxygen (mg·L−1) | 9.25 ± 0.03 A | 9.01 ± 0.09 a | 8.66 ± 0.12 B | 8.62 ± 0.10 b | 9.34 ± 0.03 C | 9.14 ± 0.04 c |
| Electrical conductivity (ms·cm−1) | 0.15 ± 0.05 A | 0.12 ± 0.04 a | 0.23 ± 0.02 B | 0.22 ± 0.01 b | 0.29 ± 0.03 C | 0.26 ± 0.02 c |
| pH | 7.83 ± 0.19 A | 7.73 ± 0.09 a | 7.86 ± 0.04 A | 7.86 ± 0.03 b | 8.18 ± 0.03 B | 8.04 ± 0.05 c |
| MSUBSTD | −4.14 ± 0.66 A | 1.26 ± 0.88 a | −4.16 ± 0.64 A | 1.26 ± 0.92 a | −4.16 ± 0.64 A | 1.26 ± 0.92 a |
| Taxa | Indicated Habitat | ||
|---|---|---|---|
| Pre-Freezing Period | Freezing Period | Thawing Period | |
| Turbellaria | |||
| Tricladida | |||
| Planariidae | |||
| Planaria sp. | Pools * | ||
| Oligochaeta | |||
| Haplotaxida | |||
| Naididae sp. | Pools * | ||
| Gastropoda | |||
| Basommatophora | |||
| Planorbidae | |||
| Anis sp. | Pools * | ||
| Malacostraca | |||
| Amphipoda | |||
| Gammaridae | |||
| Gammarus sp. | Pools * | ||
| Insect | |||
| Ephemeroptera | |||
| Heptageniidae | |||
| Rhithrogena sp. | Riffles * | Riffles * | Riffles * |
| Iron sp. | Riffles * | Riffles * | |
| Heptagenia sp. | Riffles * | ||
| Epeorus sp. | Riffles * | ||
| Cinygmula sp. | Riffles * | ||
| Baetidae | |||
| Baetis sp. | Riffles * | Riffles * | Riffles * |
| Ameletidae | |||
| Ameletus sp. | Riffles * | Riffles * | |
| Plecoptera | |||
| Perlodidae | |||
| Isoperla sp. | Riffles * | ||
| Perlodes sp. | Riffles * | ||
| Starsolus sp. | Riffles * | ||
| Nemouridae | |||
| Nemoura sp. | Riffles * | Riffles * | |
| Amphinemura sp. | Pools * | ||
| Capniidae | |||
| Capnia sp. | Riffles * | Riffles * | |
| Chloroperlidae | |||
| Suwallia sp. | Riffles * | ||
| Coleoptera | |||
| Haliplidae | |||
| Haliplus sp. | — | — | — |
| Trichoptera | |||
| Hydropsychidae | |||
| Homoplectra sp. | — | — | — |
| Glossosomatidae | |||
| Glossosoma sp. | Riffles * | ||
| Lepidostomatidae | |||
| Theliopsyche sp. | Riffles * | Pools * | |
| Brachycentridae | |||
| Brachycentrus sp. | Riffles * | Riffles * | |
| Amiocentrus sp. | Pools * | ||
| Rhyacophilidae | |||
| Rhyacophila sp. | Pools* | ||
| Limnephilidae | |||
| Desmona sp. | — | — | — |
| Pseudostenophylax sp. | — | — | — |
| Diptera | |||
| Tipulidae | |||
| Dicranota sp. | Pools * | Pools * | |
| Cheilotrichia sp. | Pools * | ||
| Tipula (Arctotipula) sp. | Pools * | Pools * | |
| Tipula (Sinotipula) sp. | Pools * | ||
| Hexatoma sp. | Riffles * | ||
| Antocha sp. | Riffles * | ||
| Chironomidae | |||
| Chironominae | |||
| Tanytarsini sp. | Pools * | Pools * | |
| Chironomus sp. | Pools * | Pools * | Pools * |
| Orthocladiinae sp.1 | Pools * | Riffles * | |
| Orthocladiinae sp.2 | Pools * | Pools * | |
| Orthocladiinae sp.3 | Riffles * | ||
| Tanypodinae sp. | Pools * | Pools * | |
| Ceratopogonidae | |||
| Sphaeromias sp. | Pools * | Pools * | |
| Psychodidae sp. | Riffles * | Pools * | Riffles * |
| Blefariceridae sp. | Riffles * | ||
| Tabanidae sp.1 | Riffles * | Riffles * | |
| Tabanidae sp.2 | Riffles * | ||
| Blephariceridae | |||
| Blepharicera sp. | Riffles * | ||
| Arachnida | |||
| Acariformes | |||
| Hydrachnellae sp. | Riffles * | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Y.; Liu, Y.; Gao, Y.; Cui, D.; Zhang, W.; Jiao, Z.; Yao, F.; Zhang, Z.; Yang, H. The Impacts of the Freezing–Thawing Process on Benthic Macroinvertebrate Communities in Riffles and Pools: A Case Study of China’s Glacier-Fed Stream. Water 2022, 14, 983. https://doi.org/10.3390/w14060983
Tian Y, Liu Y, Gao Y, Cui D, Zhang W, Jiao Z, Yao F, Zhang Z, Yang H. The Impacts of the Freezing–Thawing Process on Benthic Macroinvertebrate Communities in Riffles and Pools: A Case Study of China’s Glacier-Fed Stream. Water. 2022; 14(6):983. https://doi.org/10.3390/w14060983
Chicago/Turabian StyleTian, Yilin, Yuwei Liu, Yingzhi Gao, Dong Cui, Wei Zhang, Ziwei Jiao, Fulong Yao, Zhenxing Zhang, and Haijun Yang. 2022. "The Impacts of the Freezing–Thawing Process on Benthic Macroinvertebrate Communities in Riffles and Pools: A Case Study of China’s Glacier-Fed Stream" Water 14, no. 6: 983. https://doi.org/10.3390/w14060983