
Sunflower Photosynthetic Characteristics, Nitrogen Uptake, and Nitrogen Use Efficiency under Different Soil Salinity and Nitrogen Applications

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Field Experiments
2.3. Data Collection
2.3.1. Photosynthetic Characteristics
2.3.2. Plant Biomass and Seed Yield
2.3.3. Plant Nitrogen Uptake
2.4. Data Analysis
2.4.1. Stomatal Limitation Index
2.4.2. Nitrogen Production, Uptake, and Utilization Efficiency
2.4.3. Photosynthetic Nitrogen Use Efficiency
3. Results
3.1. Sunflower Photosynthetic Characteristics in Saline Fields
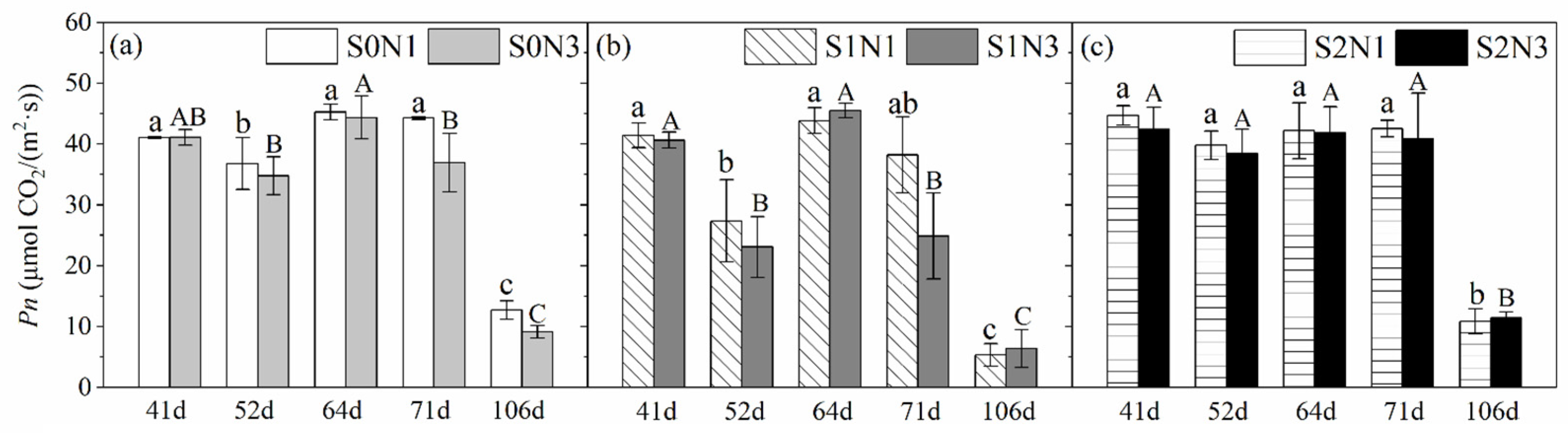
3.1.1. Net Photosynthetic Rate
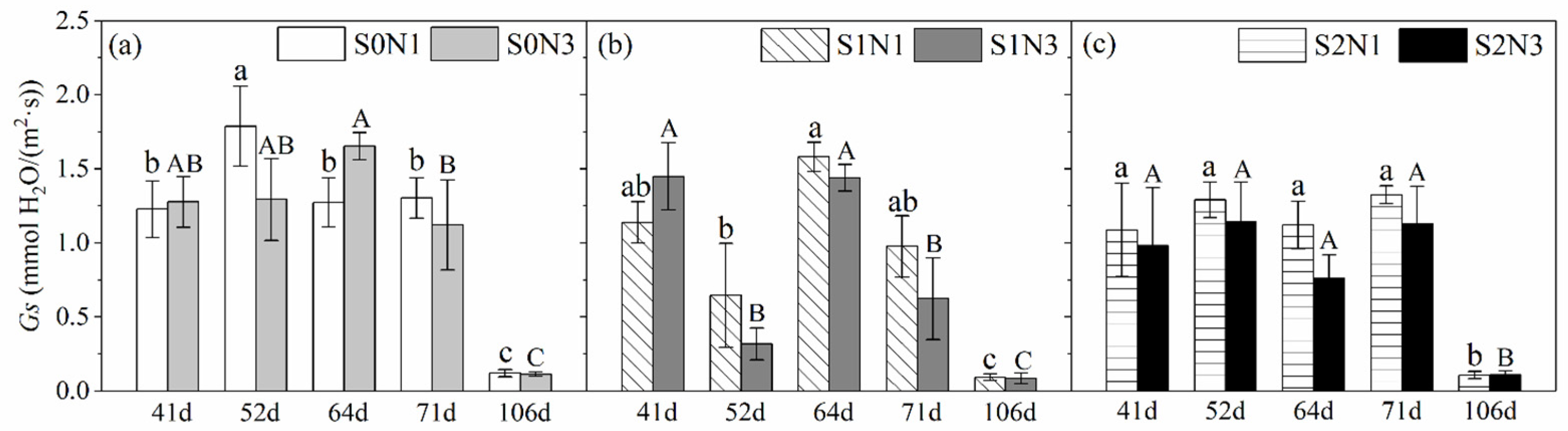
3.1.2. Stomatal Conductance
3.1.3. Intercellular CO2 Concentration and Stomatal Limitation Index
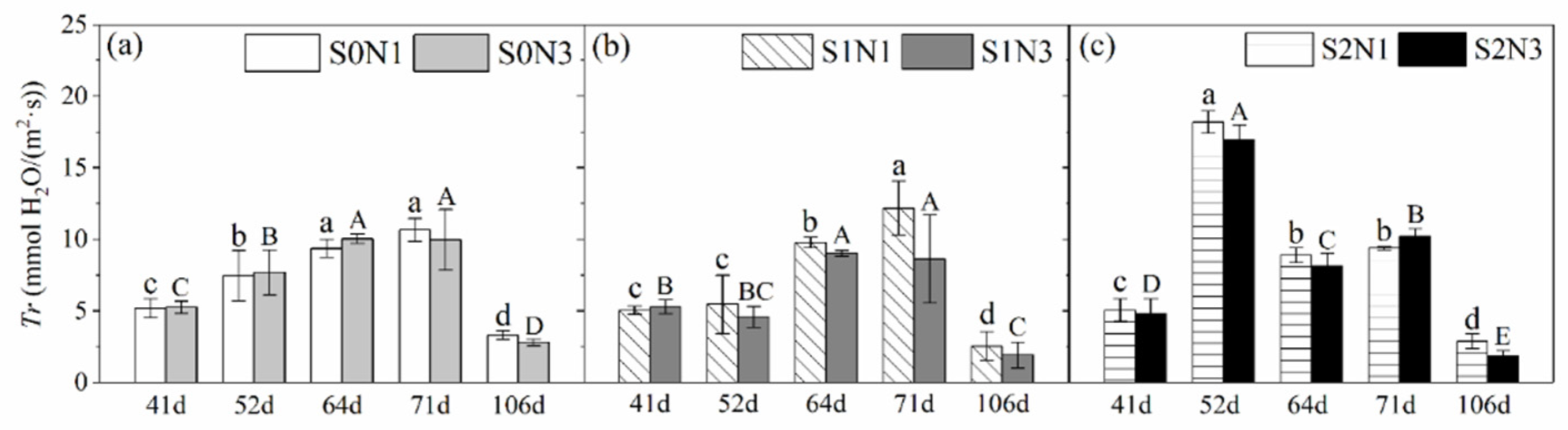
3.1.4. Leaf Transpiration Rate
3.2. Nitrogen Uptake and Utilization of Sunflowers in Saline Fields
3.2.1. Plant Nitrogen Uptake
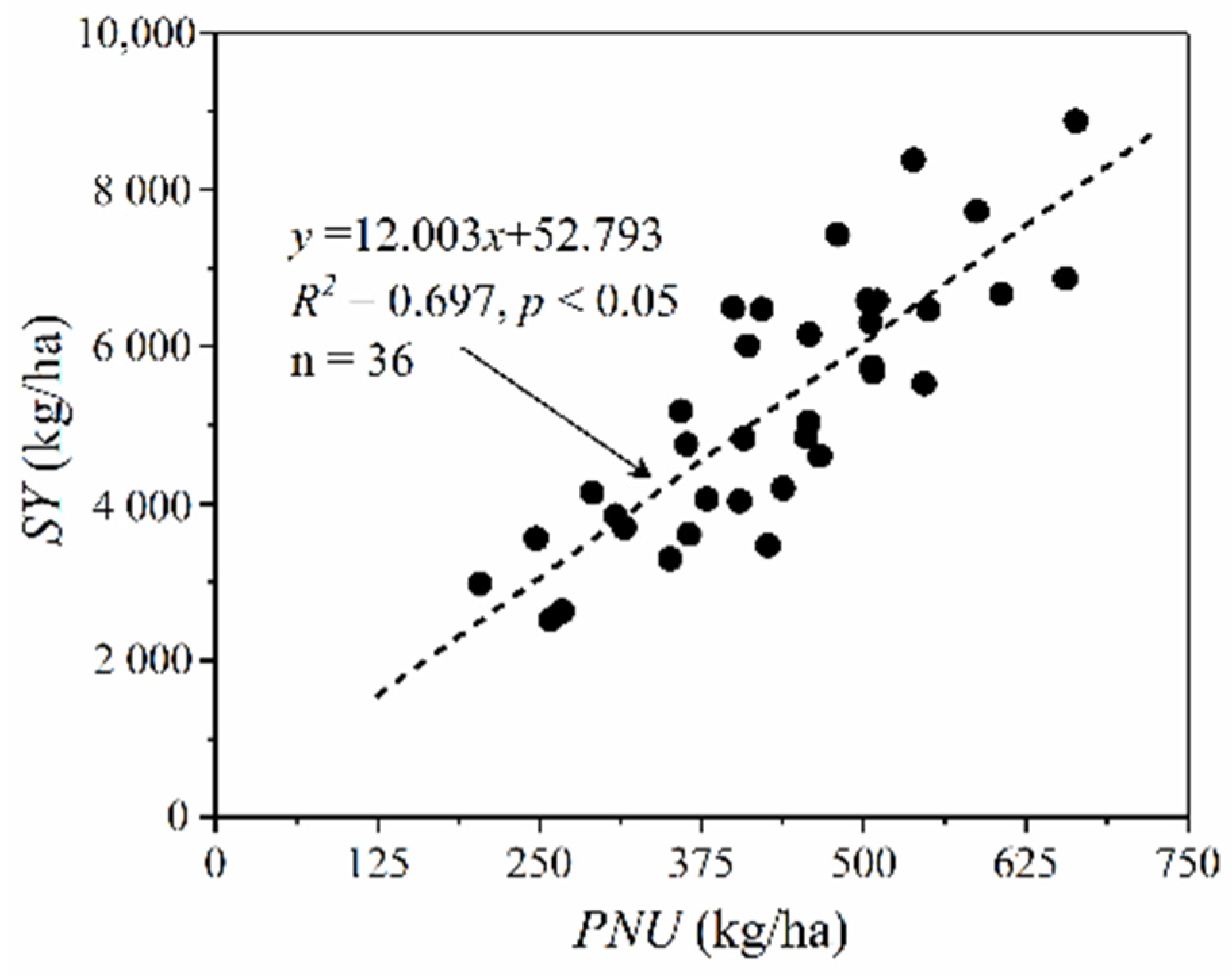
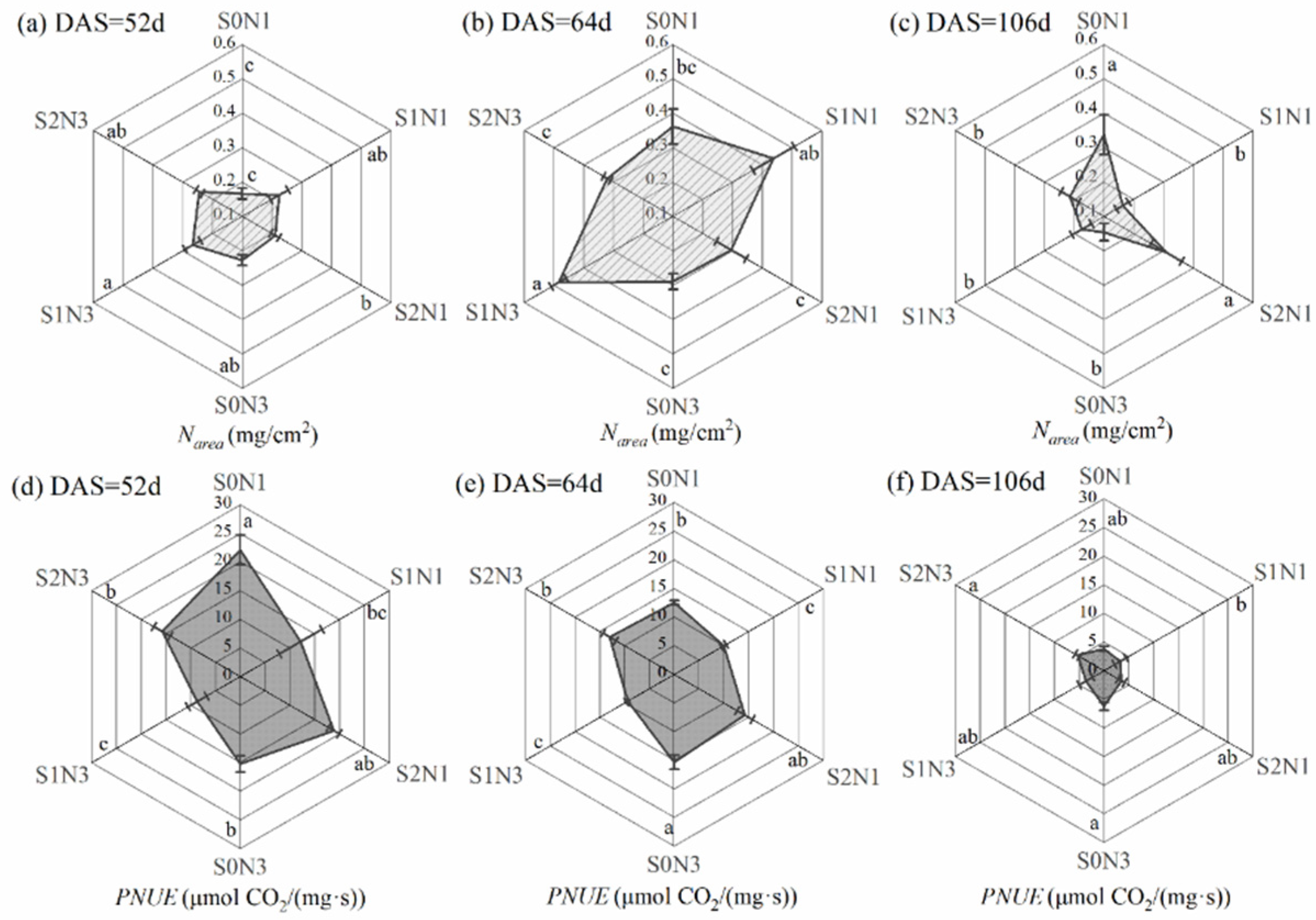
3.2.2. Photosynthetic Nitrogen Use Efficiency
3.2.3. Nitrogen Use Efficiency
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sinclair, T.R.; Rufty, T.W. Nitrogen and water resources commonly limit crop yield increases, not necessarily plant genetics. Glob. Food Secur. 2012, 1, 94–98. [Google Scholar] [CrossRef]
- Quan, Z.; Zhang, X.; Davidson, E.A.; Zhu, F.; Li, S.; Zhao, X.; Chen, X.; Zhang, L.M.; He, J.Z.; Wei, W.; et al. Fates and use efficiency of nitrogen fertilizer in maize cropping systems and their responses to technologies and management practices: A global analysis on field 15-N tracer studies. Earth’s Future 2021, 9, e2020EF001514. [Google Scholar] [CrossRef]
- Luo, L.; Zhang, Y.; Xu, G. How does nitrogen shape plant architecture? J. Exp. Bot. 2020, 71, 4415–4427. [Google Scholar] [CrossRef]
- Zörb, C.; Geilfus, C.M.; Dietz, K.J. Salinity and crop yield. Plant Biol. 2019, 21, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.Z.; Xu, C.; Wu, J.W.; Huang, J.S.; Ma, T. Effect of salinity on soil respiration and nitrogen dynamics. Ecol. Chem. Eng. S 2013, 20, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.Y.; Yang, J.S.; Yao, R.J.; Wang, X.P.; Liu, G.M.; Chen, Q. Effects of soil salinity on nitrogen transformation in Hetao Irrigation District of Inner Mongolia, China. Soils 2020, 52, 802–810, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Han, J.; Shi, J.; Zeng, L.; Xu, J.; Wu, L. Effects of nitrogen fertilization on the acidity and salinity of greenhouse soils. Environ. Sci. Pollut. Res. 2015, 22, 2976–2986. [Google Scholar] [CrossRef]
- Villa-Castorena, M.; Ulery, A.L.; Catalánvalencia, E.A.; Remmenga, M.D. Salinity and nitrogen rate effects on the growth and yield of chile pepper plants. Soil Sci. Soc. Am. J. 2003, 67, 1781–1789. [Google Scholar] [CrossRef]
- Ju, X.T.; Kou, C.L.; Christie, P.; Dou, Z.X.; Zhang, F.S. Changes in the soil environment from excessive application of fertilizers and manures to two contrasting intensive cropping systems on North China Plain. Environ. Pollut. 2007, 145, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Esmaili, E.; Kapourchal, A.; Malakouti, J.; Homaee, M. Interactive effect of salinity and two nitrogen fertilizers on growth and composition of sorghum. Plant Soil Environ. 2008, 54, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Hou, Z.; Wu, L.; Liang, Y.; Wei, C. Effects of salinity and nitrogen on cotton growth in arid environment. Plant Soil 2010, 326, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.Z.; Xu, C.; Huang, J.S.; Wu, J.W.; Ma, T. Emergence rate, yield, and nitrogen-use efficiency of sunflowers (Helianthus annuus) vary with soil salinity and amount of nitrogen applied. Commun. Soil Sci. Plant Anal. 2015, 46, 1006–1023. [Google Scholar] [CrossRef]
- Che, Z.; Wang, J.; Li, J. Effects of water quality, irrigation amount and nitrogen applied on soil salinity and cotton production under mulched drip irrigation in arid Northwest China. Agric. Water Manag. 2021, 247, 106738. [Google Scholar] [CrossRef]
- Mansour, M.F. Nitrogen containing compounds and adaptation of plants to salinity stress. Biol. Plant 2000, 43, 491–500. [Google Scholar] [CrossRef]
- Dong, H.Z. Technology and field management for controlling soil salinity effects on cotton. Aust. J. Crop Sci. 2012, 6, 333–341. [Google Scholar]
- Ma, T.; Zeng, W.Z.; Li, Q.; Yang, X.; Wu, J.W.; Huang, J.S. Shoot and root biomass allocation of sunflower varying with soil salinity and nitrogen applications. Agron. J. 2017, 109, 2545–2555. [Google Scholar] [CrossRef]
- Ma, T.; Zeng, W.Z.; Lei, G.Q.; Wu, J.W.; Huang, J.S. Predicting the rooting depth, dynamic root distribution and the yield of sunflower under different soil salinity and nitrogen applications. Ind. Crops Prod. 2021, 170, 113749. [Google Scholar] [CrossRef]
- Sharma, L.K.; Bali, S.K. A review of methods to improve nitrogen use efficiency in agriculture. Sustainability 2018, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Devkota, M.; Martius, C.; Lamers, J.P.A.; Sayre, K.D.; Devkota, K.P.; Vlek, P.L.G. Tillage and nitrogen fertilization effects on yield and nitrogen use efficiency of irrigated cotton. Soil Tillage Res. 2013, 134, 72–82. [Google Scholar] [CrossRef]
- Ashraf, M.; Shahzad, S.M.; Imtiaz, M.; Rizwan, M.S.; Arif, M.S.; Kausar, R. Nitrogen nutrition and adaptation of glycophytes to saline environment: A review. Arch. Agron. Soil Sci. 2018, 64, 1181–1206. [Google Scholar] [CrossRef]
- Zeng, W.Z.; Xu, C.; Wu, J.W.; Huang, J.S. Sunflower seed yield estimation under the interaction of soil salinity and nitrogen application. Field Crops Res. 2016, 198, 1–15. [Google Scholar] [CrossRef]
- Liu, G.H.; Zhou, B.B.; Hou, Y.L.; Duan, M.L.; Ning, S.R.; Wang, Q.J. Effects of nitrogen on winter wheat growth under different salt stress. J. Irrig. Drain. 2019, 38, 36–40, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Pei, C.Z.; Ma, T.; Liu, C.; Li, Q.; Fu, F.; Wu, J.W.; Huang, J.S. Influence of different nitrogen application rates on SPAD values of sunflower in saline field. Soil Fert. Sci. China 2016, 6, 111–115, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Zhang, J.F.; Wang, Z.H.; Zhang, J.Z.; Dou, Y.Q.; Hou, Y.S. The influences of different nitrogen and salt levels interactions on fluorescence characteristics, yield and quality of processed tomato under drip irrigation. Sci. Agric. Sin. 2020, 53, 990–1003, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Katerji, N.; Van Hoorn, J.W.; Hamdy, A.; Mastrorilli, M. Salt tolerance classification of crops according to soil salinity and to water stress day index. Agric. Water Manag. 2000, 43, 99–109. [Google Scholar] [CrossRef]
- Ma, T.; Zeng, W.Z.; Wu, J.W.; Ding, J.H.; Yu, S.E.; Huang, J.S. Sunflower canopy development, radiation absorption and use efficiency at different nitrogen application rates in saline fields. Trans. Chin. Soc. Agric. Mach. 2020, 51, 292–303, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Hachicha, M.; Mansour, M.; Rejeb, S.; Mougou, R.; Askrim, H.; Abdegawed, J. Applied research for the utilization of brackish/saline water in Central Tunisia: Water use, salinity evolution and crop response. In Proceedings of the International Salinity Forum, Riverside, CA, USA, 25–27 April 2005. [Google Scholar]
- Zeng, W.Z.; Xu, C.; Wu, J.W.; Huang, J.S.; Zhao, Q.; Wu, M.S. Impacts of salinity and nitrogen on the photosynthetic rate and growth of sunflowers (Helianthus annuus L.). Pedosphere 2014, 24, 635–644. [Google Scholar] [CrossRef]
- Schneiter, A.; Miller, J. Description of Sunflower Growth Stages. Crop Sci. 1981, 21, 901–903. [Google Scholar] [CrossRef]
- Pontes, F.; Carneiro, M.; Vaitsman, D.; da Rocha, G.; da Silva, L.; Neto, A.; Monteiro, M. A simplified version of the total Kjeldahl nitrogen method using an ammonia extraction ultrasound-assisted purge-and-trap system and ion chromatography for analyses of geological samples. Anal. Chim. Acta 2009, 632, 284–288. [Google Scholar] [CrossRef]
- Xu, D.Q. Some problems in stomatal limitation analysis of photosynthesis. Plant Physiol. Commun. 1997, 33, 241–244, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Wen, B.B.; Li, C.; Fu, X.L.; Li, D.M.; Li, L.; Chen, X.D.; Wu, H.Y.; Cui, X.W.; Zhang, X.H.; Shen, H.Y.; et al. Effects of nitrate deficiency on nitrate assimilation and chlorophyll synthesis of detached apple leaves. Plant Physiol. Biochem. 2019, 142, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Gaudinier, A.; Rodriguez-Medina, J.; Zhang, L.; Olson, A.; Liseron-Monfils, C.; Bågman, A.-M.; Foret, J.; Abbitt, S.; Tang, M.; Li, B.; et al. Transcriptional regulation of nitrogen-associated metabolism and growth. Nature 2018, 563, 259–264. [Google Scholar] [CrossRef]
- Jia, K.; Yan, C.; Yan, H.; Gao, J. Physiological responses of turnip (Brassica rapa L. subsp. rapa) seedlings to salt stress. HortScience 2020, 55, 1567–1574. [Google Scholar] [CrossRef]
- Hichem, H.; El Naceur, A.; Mounir, D. Effects of salt stress on photosynthesis, PSII photochemistry and thermal energy dissipation in leaves of two corn (Zea mays L.) varieties. Photosynthetica 2009, 47, 517–526. [Google Scholar] [CrossRef]
- Zhang, D.M.; Li, W.J.; Xin, C.S.; Tang, W.; Eneji, E.; Dong, H.Z. Lint yield and nitrogen use efficiency of field-grown cotton vary with soil salinity and nitrogen application rate. Field Crops Res. 2012, 138, 63–70. [Google Scholar] [CrossRef]
- Hong, M.H.; Zeng, W.Z.; Ma, T.; Lei, G.Q.; Zha, Y.Y.; Fang, Y.H.; Wu, J.W.; Huang, J.S. Determination of Growth Stage-Specific Crop Coefficients (Kc) of Sunflowers (Helianthus annuus L.) under Salt Stress. Water 2017, 9, 215. [Google Scholar] [CrossRef] [Green Version]
- Pei, L.; Wang, Z.H.; Zheng, X.R.; Li, W.H. Effects of soil salt content on water-salt dynamic and photosynthetic characteristics of drip irrigated silage maize in sequential cropping systems. Agric. Res. Arid Areas 2016, 34, 77–84, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Ma, T.; Zeng, W.Z.; Li, Q.; Wu, J.W.; Huang, J.S. Effects of water, salt and nitrogen stress on sunflower (Helianthus annuus L.) at different growth stages. J. Soil Sci. Plant Nutr. 2016, 16, 1024–1037. [Google Scholar] [CrossRef]
- Ciampitti, I.A.; Vyn, T.J. Physiological perspectives of changes over time in maize yield dependency on nitrogen uptake and associated nitrogen efficiencies: A review. Field Crops Res. 2012, 133, 48–67. [Google Scholar] [CrossRef]
- Wang, X.; Wang, G.; Turner, N.C.; Xing, Y.; Li, M.; Guo, T. Determining optimal mulching, planting density, and nitrogen application to increase maize grain yield and nitrogen translocation efficiency in Northwest China. BMC Plant Biol. 2020, 20, 282. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, M.; Li, Y.; Li, J.; Jing, Y.; Xiang, Y.; Yao, B.; Deng, Q. Linkage of crop productivity to soil nitrogen dynamics under biochar addition: A meta-analysis across field studies. Agronomy 2022, 12, 247. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, W.; Krienke, B.; Cao, Q.; Zhu, Y.; Cao, W.; Liu, X. In-season variable rate nitrogen recommendation for wheat precision production supported by fixed-wing UAV imagery. Precis. Agric. 2021, 22, 364–386. [Google Scholar] [CrossRef]
- Yan, F.L.; Zhang, F.C.; Fan, X.K.; Wang, Y.; Guo, J.J.; Zhang, C.Y. Effects of water and nitrogen fertilizer supply on yield and nitrogen absorption and utilization efficiency of spring maize in sandy soil area in Ningxia. Trans. Chin. Soc. Agric. Mach. 2020, 51, 283–293, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Zubillaga, M.; Aristi, P.; Lavado, S. Effect of phosphorus and nitrogen fertilization on sunflower (Helianthus annus L.) nitrogen uptake and yield. J. Agron. Crop Sci. 2002, 188, 267–274. [Google Scholar] [CrossRef]
- Murtaza, G.; Azooz, M.M.; Murtaza, B.; Usman, Y.; Saqib, M. Nitrogen-use-efficiency (NUE) in plants under nacl stress. In Salt Stress in Plants; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Onoda, Y.; Hikosaka, K.; Hirose, T. Allocation of nitrogen to cell walls decreases photosynthetic nitrogen-use efficiency. Funct. Ecol. 2004, 18, 419–425. [Google Scholar] [CrossRef]
- Hikosaka, K. Interspecific difference in the photosynthesis–nitrogen relationship: Patterns, physiological causes, and ecological importance. J. Plant Res. 2004, 117, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Nasar, J.; Khan, W.; Khan, M.Z.; Gitari, H.I.; Gbolayori, J.F.; Moussa, A.A.; Mandozai, A.; Rizwan, N.; Anwari, G.; Maroof, S.M. Photosynthetic activities and photosynthetic nitrogen use efficiency of maize crop under different planting patterns and nitrogen fertilization. J. Soil Sci. Plant Nutr. 2021, 21, 2274–2284. [Google Scholar] [CrossRef]
- Nandy, P.; Das, S.; Ghose, M.; Spooner, R. Effects of salinity on photosynthesis, leaf anatomy, ion accumulation and photosynthetic nitrogen use efficiency in five Indian mangroves. Wetlands Ecol. Manag. 2007, 15, 347–357. [Google Scholar] [CrossRef]
- Debez, A.; Saadaoui, D.; Ramani, B.; Ouerghi, Z.; Koyro, H.; Huchzermeyer, B.; Abdelly, C. Leaf H+-ATPase activity and photosynthetic capacity of Cakile maritima under increasing salinity. Environ. Exp. Bot. 2006, 57, 285–295. [Google Scholar] [CrossRef]
- Dinh, T.H.; Watanabe, K.; Takaragawa, H.; Nakabaru, M.; Kawamitsu, Y. Photosynthetic response and nitrogen use efficiency of sugarcane under drought stress conditions with different nitrogen application levels. Plant Prod. Sci. 2017, 20, 412–422. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | Treatments | S † | N Rate | Basal N Rate | Top-Dressed N Rate |
|---|---|---|---|---|---|
| dS/m | kg/ha | kg/ha | kg/ha | ||
| 2015 | S0N0 | 1.878 | 45 | 45 | 0 |
| S0N2 | 1.723 | 135 | 45 | 90 | |
| S1N0 | 5.017 | 45 | 45 | 0 | |
| S1N2 | 5.898 | 135 | 45 | 90 | |
| S2N0 | 8.157 | 45 | 45 | 0 | |
| S2N2 | 9.035 | 135 | 45 | 90 | |
| 2016 | S0N1 | 2.613 | 90 | 90 | 0 |
| S0N3 | 2.227 | 180 | 90 | 90 | |
| S1N1 | 4.731 | 90 | 90 | 0 | |
| S1N3 | 5.515 | 180 | 90 | 90 | |
| S2N1 | 6.847 | 90 | 90 | 0 | |
| S2N3 | 7.158 | 180 | 90 | 90 |
| Years | Treatments | N Rate | SY | NPE | NUPE | NUTE |
|---|---|---|---|---|---|---|
| kg/ha | kg/ha | kg/(kg N) | kg/kg | kg/(kg N) | ||
| 2015 | S0N0 ‡ | 45 | 6161.6 ± 172.5 ab † | 136.9 ± 3.3 a | 10.2 ± 1.1 a | 13.5 ± 1.1 b |
| S0N2 | 135 | 7436.3 ± 1119.7 a | 55.1 ± 7.0 c | 3.6 ± 0.4 c | 15.5 ± 0.1 a | |
| S1N0 | 45 | 4608.5 ± 1342.2 b | 102.4 ± 25.4 b | 10.4 ± 0.9 a | 9.9 ± 1.6 c | |
| S1N2 | 135 | 4853.4 ± 966.4 b | 36.0 ± 6.1 cd | 3.4 ± 0.4 c | 10.7 ± 0.6 c | |
| S2N0 | 45 | 3693.0 ± 1249.6 b | 82.1 ± 23.6 b | 7.0 ± 1.1 b | 11.7 ± 1.6 c | |
| S2N2 | 135 | 3558.5 ± 685.5 b | 26.4 ± 4.3 d | 1.8 ± 0.3 d | 14.4 ± 0.2 ab | |
| 2016 | S0N1 | 90 | 6589.9 ± 87.5 ab | 73.2 ± 1.0 a | 5.6 ± 1.1 a | 13.1 ± 2.6 a |
| S0N3 | 180 | 7734.9 ± 1148.5 a | 43.0 ± 6.4 bc | 3.3 ± 0.4 bc | 13.2 ± 0.2 a | |
| S1N1 | 90 | 5037.8 ± 1433.1 b | 56.0 ± 16.0 b | 5.1 ± 1.0 a | 11.0 ± 1.0 ab | |
| S1N3 | 180 | 5533.2 ± 1338.1 b | 30.7 ± 7.4 cd | 3.0 ± 0.6 bc | 10.1 ± 0.5 b | |
| S2N1 | 90 | 4061.3 ± 766.8 b | 45.1 ± 8.5 bc | 4.2 ± 0.3 ab | 10.7 ± 1.2 ab | |
| S2N3 | 180 | 3849.5 ± 1330 b | 21.4 ± 7.4 d | 1.7 ± 0.3 c | 12.5 ± 2.3 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, T.; Chen, K.; He, P.; Dai, Y.; Yin, Y.; Peng, S.; Ding, J.; Yu, S.; Huang, J. Sunflower Photosynthetic Characteristics, Nitrogen Uptake, and Nitrogen Use Efficiency under Different Soil Salinity and Nitrogen Applications. Water 2022, 14, 982. https://doi.org/10.3390/w14060982
Ma T, Chen K, He P, Dai Y, Yin Y, Peng S, Ding J, Yu S, Huang J. Sunflower Photosynthetic Characteristics, Nitrogen Uptake, and Nitrogen Use Efficiency under Different Soil Salinity and Nitrogen Applications. Water. 2022; 14(6):982. https://doi.org/10.3390/w14060982
Chicago/Turabian StyleMa, Tao, Kaiwen Chen, Pingru He, Yan Dai, Yiqun Yin, Suhan Peng, Jihui Ding, Shuang’en Yu, and Jiesheng Huang. 2022. "Sunflower Photosynthetic Characteristics, Nitrogen Uptake, and Nitrogen Use Efficiency under Different Soil Salinity and Nitrogen Applications" Water 14, no. 6: 982. https://doi.org/10.3390/w14060982