
Seawater with Added Monosodium Glutamate Residue (MSGR) Is a Promising Medium for the Cultivation of Two Commercial Marine Microalgae

1
School of Environmental Science and Engineering, Shandong University, Qingdao 266237, China
2
Shandong Provincial Engineering Center on Environmental Science and Technology, Jinan 250061, China
3
Department of Environmental Science and Engineering, Fudan University, Shanghai 200433, China
*
Author to whom correspondence should be addressed.
Water 2022, 14(6), 975; https://doi.org/10.3390/w14060975
Submission received: 11 February 2022
/
Revised: 8 March 2022
/
Accepted: 16 March 2022
/
Published: 19 March 2022
(This article belongs to the Section Wastewater Treatment and Reuse)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Phaeodactylum tricornutum and Nannochloropsis oceanica, with their satisfactory performance in accumulating lipids and other high-value products, have been successfully used for commercial production in recent years. However, costly chemicals in culture media greatly increase the price of the resulting bioproducts. To control the cultivation cost, this paper assessed the potential of seawater supplemented with monosodium glutamate residue wastewater at a ratio of 1/500 (S-MSGR) to serve as a growing medium for these two marine species. Compared with the standard chemical culture medium, Erdschreiber’s medium (EM), both the algal growth and metabolite accumulation of P. tricornutum and N. oceanica were greatly promoted in S-MSGR. The maximum biomass concentrations of P. tricornutum and N. oceanica reached 0.93 and 0.36 g/L, which were, respectively, 1.5 and 1.9 times higher than those in EM medium. For lipid accumulation, P. tricornutum exhibited an excellent lipid productivity of 22.9 mg/L/day in S-MSGR, a 64% increase compared to EM medium. Furthermore, the average yield coefficients indicated good performance of P. tricornutum and N. oceanica in transferring the nitrogen in S-MSGR to the biomass, at 74.8 and 174.8 mg/g of nitrogen. In addition, compared with EM, the costs of the medium for lipid production of P. tricornutum and N. oceanica cultured in S-MSGR were USD 2.3 and 5.8/(kg lipid), which saved 96.9% and 97.6%, respectively. Therefore, this paper demonstrates that S-MSGR is a suitable nutrient resource for P. tricornutum and N. oceanica, and it has a great potential to cut the cultivation cost during real commercial production.
1. Introduction
Microalgal biological resources can be widely used in various sectors, including energy, food, animal breeding industry, medicine, cosmetics and environment, giving them great commercial potential and value [1,2]. There are many species of microalgae, which can adapt to living in a variety of environmental conditions and are easy to cultivate. Microalgae characteristically have a shorter growth cycle and less demand for agricultural land than general higher plants [3]. Moreover, compared with lignocellulosic biomass biofuels, the main disadvantages of lignocellulosic biomass biofuels utilization are biological conversion processes with low reaction rates and thermochemical conversion technologies, such as the quality of bio-oil products and low energy efficiency, and low product yield [4,5]. At present, several microalgal species have been used commercially due to their satisfactory performance in biomass and lipid accumulation, such as Phaeodactylum tricornutum and Nannochloropsis oceanica.
P. tricornutum is a widely studied marine diatom and has been explored to produce products on a pilot scale, including biodiesel, fatty acids of nutraceutical value (eicosapentaenoic acid, EPA and docosahexaenoic acid, DHA) and fucoxanthin [6]. N. oceanica is a species of miniature marine microalga with high nutritional value and lipid content, which has drawn attention as a potential feedstock for biofuels and aquaculture [7]. However, the main choice of culture medium for these two marine algae continues to be the use of chemical reagents as the nutrient sources, with the result that the high cost of nutrient supply is still a severe challenge for large-scale commercialization of microalgal biomass [8].
Although microalgae can survive and thrive in wastewater, and simultaneously synthesize lots of valuable metabolites through their ability to assimilate nitrogen (N), phosphorous (P) and other nutrients in wastewater [9], wastewater-based technologies of microalgal cultivation face some challenges, such as the capital consumed in the cultivation process and the selection of wastewater [10]. Recent studies have shown that some kinds of wastewater were successfully used for growing microalgae, including textile wastewater, municipal wastewater, swine wastewater, coal seam gas industrial wastewater and pharmaceutical wastewater [11], which effectively reduces the nutrient cost required for the cultivation of microalgae. For example, Dębowski et al. [12] used digestate from anaerobic digestion of dairy wastewater as culture medium to cultivate microalgae, which obtained 3 g/L biomass, and the lipid content also reached 20%. In addition, recent studies have shown that coke-oven wastewater containing high ammonia nitrogen [13] and monosodium glutamate residue (MSGR) wastewater were also very potential sources of nutrients. From the viewpoint of wastewater selection, it is better for it to contain no heavy metals or other toxic substances to meet the needs of microalgal culture and application after harvest.
MSGR wastewater is a nitrogen- and phosphorus-rich residual wastewater that is generated after glutamate extraction and sterilization in the process of monosodium glutamate production in the food industry [14]. MSGR does not contain hazardous or pathogenic matter [15], which seems to make it an ideal nutrient source for culturing microalgae. Previous studies in our group have proven that MSGR is suitable for limnetic microalgae, including Chlorella vulgaris, Golenkinia sp. and Scenedesmus sp., whether in freshwater or seawater [14,16]. However, basic data to grow marine microalgae in MSGR are still lacking. Inexhaustible natural seawater supplemented with nutrient-rich MSGR may be an ideal medium for cultivation of marine microalgae and would achieve the goal of saving nutrient costs [17]. Therefore, it is necessary to conduct trials to verify the feasibility of culturing marine microalgae in natural seawater supplemented with MSGR (S-MSGR), especially for the two commercial marine microalgae, P. tricornutum and N. oceanica.
Based on the foregoing considerations, in this study, two common commercial marine microalgae, P. tricornutum and N. oceanica, were grown in seawater combined with MSGR to provide an economic and effective strategy for microalgal cultivation to further improve the diversified application of microalgae in natural seawater. The main aims of this study were to: (1) investigate the growth characteristics and nutrient assimilation of the two microalgae under the optimal addition ratio of MSGR; (2) ascertain the main metabolites of these two microalgae in S-MSGR; and (3) analyze the potential application of this technology for industrial production of marine microalgae.
2. Materials and Methods
2.1. Microalgal Strain and Culture
During the experiments, two marine microalgae species were used, Phaeodactylum tricornutum FACHB-863 and Nannochloropsis oceanica FACHB-926, which were purchased from the Freshwater Algae Culture Collection of the Institute of Hydrobiology (FACHB). Before the experiments, P. tricornutum and N. oceanica were cultivated in Erdschreiber’s medium (EM), consisting of 12 mL/L P-IV metallic stock solution, 200 mg/L NaNO3, 18 mg/L Na2HPO4·7H2O, 0.135 mg/L cyanocobalamin and 50 mL/L soil extract solution in filtered seawater. One liter of P-IV metallic stock solution contained 750 mg Na2EDTA·2H2O, 97 mg FeCl3·6H2O, 41 mg MnCl2·4H2O, 5 mg ZnCl2, 4 mg Na2MoO4·2H2O and 2 mg CoCl2·6H2O. To prepare the soil extract solution, 200 g of garden soil (without fertilizer) was put into a triangular bottle containing 1000 mL of distilled water. The mouth of the bottle was sealed with a porous plug, the bottle was heated by boiling water in a water bath for 3 h, and then, it was cooled and the contents allowed to precipitate for 24 h. This process was consecutively carried out 3 times. Then, the solution was filtered, and the supernatant was taken and sterilized in an autoclave. The microalgae were cultivated at 25 ± 1 °C under 45 μmol/m2/s continuous illumination.
2.2. Optimization of Dilution Ratio of MSGR
MSGR used for the experiments was supplied by Liangshan Linghua Gourmet Powder (Jining, China). MSGR was filtered through 8 layers of gauze to remove larger particles before being used in this study, and the nutrient characteristics of MSGR were as follows: 85.0 ± 5.6 g/L total nitrogen (TN), 2.8 ± 0.4 g/L total phosphorus (TP), 78.3 ± 4.5 g/L ammonia nitrogen (NH3-N), 108.3 ± 6.7 g/L CODCr and pH 3.2 ± 0.4.
In this study, based on the contents of nitrogen and phosphorus in the standard medium of these two species of algae, we designed three different dilution ratios (S-MSGR/seawater, 1/500, 1/1000, 1/2000, v/v). Under the same conditions (25 ± 1 °C and 45 μmol/m2/s continuous illumination), three parallel experiments were carried out at the same time, and the growth status of microalgae was characterized by the concentration of total chlorophyll.
2.3. Experimental Design
The media were prepared by diluting MSGR with seawater at the optimum dilution ratio. The two species of microalgae were cultivated in 1 L conical flasks with 0.8 L working volumes of the respective aforementioned seawater media under the condition of 25 ± 1 °C temperature and 45 μmol/m2/s continuous illumination. The initial inoculum concentration of microalgae in this experiment was about OD680 = 0.2. In addition, using standard medium (EM) as a control, three parallel experiments were carried out under the same conditions.
2.4. Algal Analysis
2.4.1. Determination of the Total Chlorophyll Content
The total chlorophyll (Total Chl) concentration of microalgae was measured by the method of methanol extraction [18]. Firstly, 1.5 mL of microalgal culture was transferred into a 2 mL centrifuge tube. After centrifugation, the supernatant was discarded, and the solids were resuspended to the same volume with methanol (99%). Next, the mixture was immersed at 45 °C for 24 h in the dark. After the dark treatment, the supernatant was collected after centrifugation, and the absorbances were measured at wavelengths of 652.4, 665.2 and 750 nm on a UV–visible spectrophotometer (UV-2450, Shimadzu, Japan). The total Chl concentration was then estimated by use of the following equation:
Total Chl (mg/L) = 1.44 (A665.2 − A750) + 24.93 (A652.4 − A750).
2.4.2. Measurement of Cell Biomass Concentration
The biomass concentration (X) was determined by weighing a certain volume of dry biomass [19]. In short, a 10 mL volume of microalgal solution was filtered through a 0.45 μm membrane, which was then dried to constant mass. The biomass concentration (g/L) was calculated as the dry mass per liter of algae culture solution. The OD680 (optical density at 680 nm) and corresponding biomass concentration of microalgae in different growth stages were measured, and the relation equations between OD680 and biomass concentration were fitted. Microalgal growth was monitored every day by measuring OD680, and the relationships between biomass concentration and the OD680 of the two microalgae were calculated following the equations
and
XP. tricornutum = 0.9773 OD680 + 0.0313, with R2 = 0.9964,
XN. oceanica = 0.2916 OD680 + 0.0254, with R2 = 0.9951.
The biomass productivity, Pb (g/L/day), was calculated from the variation of the biomass concentration within a specific cultivation time according to the following equation:
where ΔT (day) and ΔX (g/L) are the duration of the run and the variation of the biomass concentration, respectively.
Pb = ΔX/ΔT,
The specific growth rate, μ (day−1), of microalgae was calculated by the following equation:
where X1 and X2 (g/L) represent the dry biomass concentrations at times T1 and T2, respectively.
μ = [ln(X2) − ln(X1)]/(T2 − T1),
2.4.3. Determination of Metabolites in Biomass
The carbohydrate content of algae was measured by the anthrone–sulfuric acid method [20]. Specifically, the algae were harvested by centrifugation, and the collected algal biomass was freeze dried to constant weight at −50 °C in a lyophilizer (EYELA FDU-1200, Tokyo Rikakikai, Japan) to obtain dried algal powder. After that, the dried algal sample, 10 mL hydrochloric acid and 15 mL water were mixed in a 50 mL conical flask and heated in boiling water for 20 min. The hot, mixed system was then filtered and diluted to 100 mL. Next a 1 mL sample with 4 mL of anthrone (2% H2SO4 by mass) was transferred into a clean glass vial with a screw cap, and the bottom of the glass vial was immersed in boiling water to boil for exactly 10 min, then cooled to room temperature with ice water. Finally, the extracted carbohydrate was measured by colorimetric assay on a 96-well plate at a wavelength of 620 nm in a microplate photometer (Multiskan FC, Thermo, Waltham, MA, USA).
The protein content of algae was measured by the Bradford method [21]. An amount of 10 mL of distilled water was added to the dried algal powder, which was accurately weighed, and then, the algal cells were broken by ultrasonic crusher under ice-bath conditions. The supernatant was collected by centrifugation. Next, a 1 mL extract with 5 mL of Coomassie brilliant blue G-250 (0.1 g/L) was transferred into a clean glass vial with a screw cap and oscillated. The absorbance of the supernatant was measured by UV–visible spectrophotometer at 595 nm. Bovine serum albumin was used as the standard curve to calculate the protein content.
The lipid content of algae was measured by the chloroform–methanol method [22]. Specifically, the dried algal sample and chloroform–methanol solution (2:1, v/v) were mixed, followed by ultrasound for 10 min. After that, the supernatant was collected in a long test tube by centrifugation. The entire extraction process was repeated twice. Sodium chloride solution (0.9%) of one-fifth the volume of the lipid extract was added to the system. The mixed liquid was vortex vibrated for 1 min, then left to stand for 15 min. All the liquid in the organic phase was transferred through a 0.22 μm organic membrane to a clean, short test tube of known mass. Finally, the contents of the tube were dried by blowing nitrogen at 60 °C to obtain the lipid mass.
2.5. Nutrient Measurements
The total nitrogen (TN) and total phosphorus (TP) of filtered samples were determined by the alkaline potassium persulfate digestion UV spectrophotometric method (HJ 636-2012) and the ammonium molybdate spectrophotometric method (GB/T 11893-1989) [23], respectively. The assimilation efficiency, AE (%), and the average yield coefficient, AYC (mg/g), were calculated by use of the following equations [24]:
and
where C0 is the initial concentration of nutrients (TN and TP), ΔC (mg/L) is the reduction in concentration due to consumption of nutrients during cultivation, and ΔX (g/L) is the increase in the biomass concentration.
AE = ΔC/C0,
AYC = ΔC/ΔX,
2.6. Statistical Analysis
The results are presented in the mean values ± standard deviation (s.d.) from three independent experiments and analyzed using one-way analysis of variance in Duncan’s test. A value of p < 0.05 was considered statistically significant.
3. Results and Discussion
3.1. Effects of S-MSGR on Growth
According to the nutrients produced in standard medium by P. tricornutum and N. oceanica, three different dilutions of MSGR in natural seawater were specified to determine the optimal culture condition (Figure S1). The optimal dilution factor for P. tricornutum and N. oceanica was 500 times, which yielded total chlorophyll contents of 4.86 and 6.22 mg/L for the two respective algal species. Therefore, the condition of 1/500 was chosen in the following experiments.
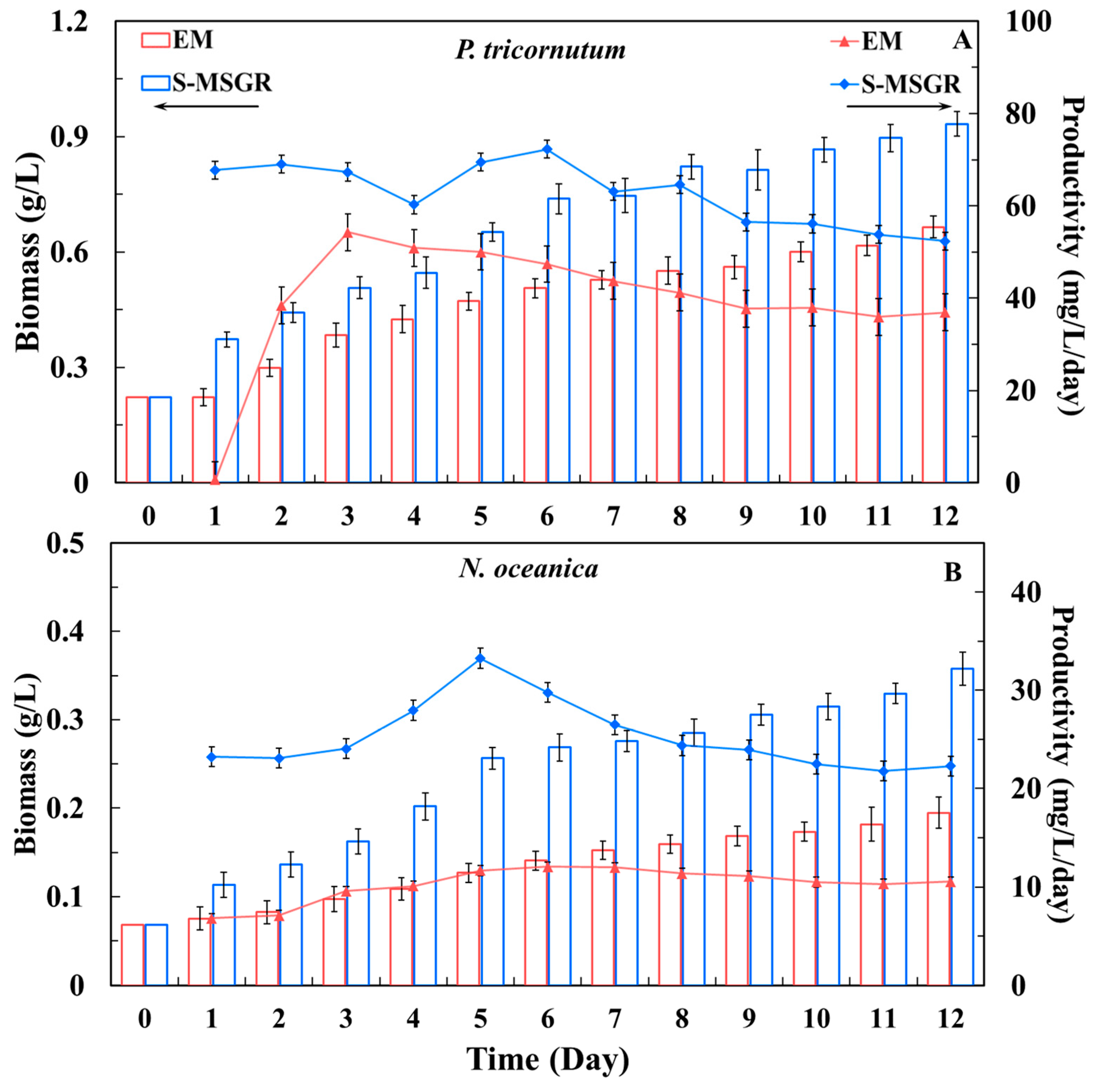
Figure 1 presents the variation in growth characteristics of the two microalgae. The biomass concentrations of P. tricornutum and N. oceanica reached 0.93 and 0.36 g/L in S-MSGR, which were 1.5 and 1.9 times higher than the respective values in standard medium. During the first week of culture, the biomass productivity of P. tricornutum in S-MSGR fluctuated around 67 mg/L/day, and the maximum value of 72.27 mg/L/day, obtained on the sixth day, was evidently higher than the 50.04 mg/L/day obtained in EM medium. Subsequently, with the consumption of nutrients, the biomass productivity gradually decreased to 52.34 mg/L/day at the harvest time. For N. oceanica, the biomass productivity at the beginning of the experiment increased rapidly to 33.26 mg/L/day (on the fifth day). Similarly, the biomass productivity gradually decreased due to the reduction in nutrients in the medium and finally stabilized at 22.28 mg/L/day. Figure 2 shows the specific growth rates of the two microalgae under EM and S-MSGR culture conditions. The average specific growth rates of P. tricornutum and N. oceanica cultured in S-MSGR were clearly different from those in EM medium, exhibiting increases of about 33.3% and 58.6%, respectively. The results indicate that using S-MSGR as the medium for growth of P. tricornutum and N. oceanica could effectively improve their biomass productivity, and thus promote the rapid accumulation of biomass.
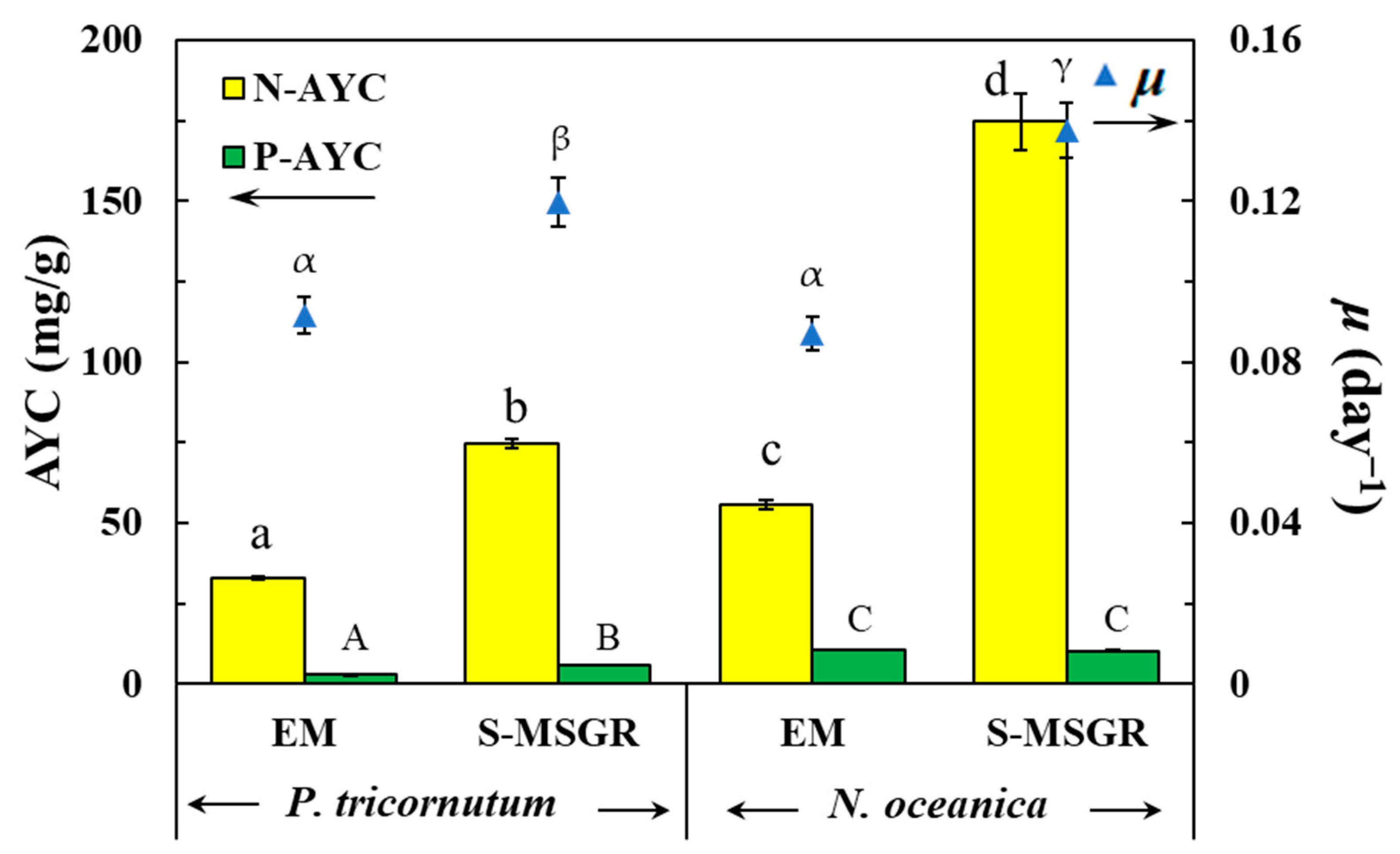
The nutrient assimilations of the two microalgae in the standard and S-MSGR media are shown in Figure 2 and Table S1. These two microalgae cultured with S-MSGR showed excellent capacities for TP assimilation, with P. tricornutum and N. oceanica achieving 97.8% and 66.9%, respectively. Compared with total phosphorus, the assimilation efficiency of total nitrogen (AEN) was slightly lower. The maximum AEN was only 38.9%, which was attained by P. tricornutum. This was mainly because the initial TN concentration in the MSGR medium was relatively high (179.4 mg/L), indicating that the growth of microalgae in this experiment was limited by the concentration of phosphorus. Therefore, this factor should be taken seriously in the following research. In this study, there was a high nutrient assimilation efficiency for P. tricornutum cultured in S-MSGR, which corresponded to its high biomass concentration, lipid, and protein productivity. Cultivating P. tricornutum in seawater with added MSGR not only enabled assimilation of N and P nutrients from MSGR, but also obtained higher lipid and protein productivity than in standard medium, indicating that S-MSGR, as the medium of growth for P. tricornutum, was fully in line with commercial demands. The average yield coefficient (AYC) was defined as the ratio of the amount of TN or TP assimilated by microalgae to the quantity of biomass produced; that is, the amount of nutrients used by the microalgae per gram of biomass produced [19]. N. oceanica manifested a maximum value of 174.8 mg/g for N-AYC (Figure 2), which showed an excellent ability to assimilate the nitrogen in MSGR.
3.2. Changes to Photosynthesis in S-MSGR
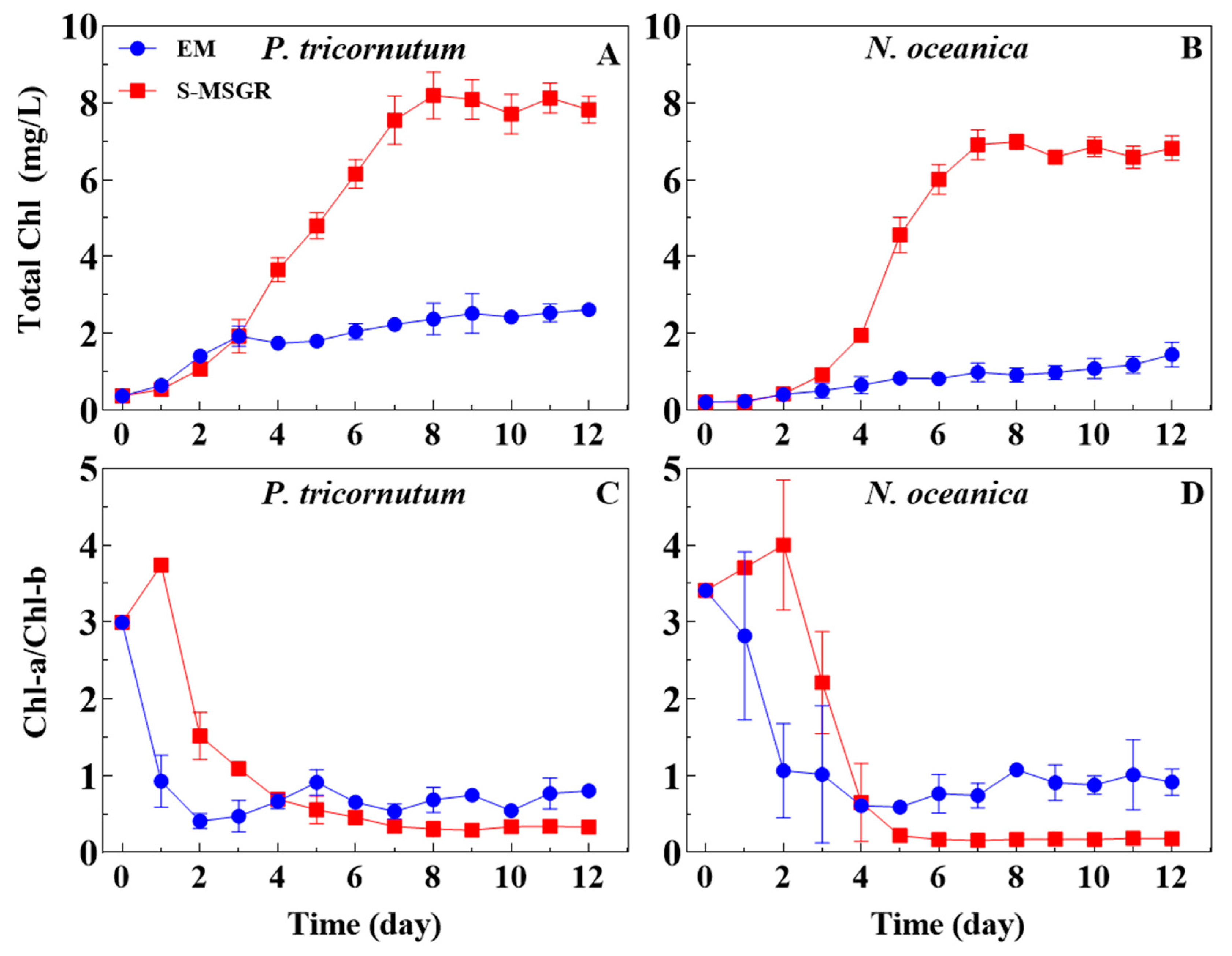
Chlorophyll is the main indicator in the photosynthetic system of algal cells, which can reflect the growth status of microalgae [25]. As shown in Figure 3, the total chlorophyll content of P. tricornutum and N. oceanica reached maximum values of 8.19 and 6.99 mg/L, respectively, within 7 days of being cultured in S-MSGR medium, while the total chlorophyll contents for EM medium were only 2.37 and 0.92 mg/L, respectively. The better chlorophyll synthesis of the two microalgae in S-MSGR demonstrated their higher photosynthetic efficiency in S-MSGR than in standard medium. It is generally considered that the ratio of Chl-a/Chl-b can reflect the ability to capture light energy related to the photochemical reaction rate of PS-II [26]. Great changes in Chl-a/Chl-b in the early stage of the two algae being cultured in EM or S-MSGR indicated the adaptation process in the new environment. On the fourth day, all the ratios tended to be stable, showing that the adaptation response of microalgae had finished. Finally, the Chl-a/Chl-b ratios for P. tricornutum stabilized at about 0.70 and 0.32 when cultured in EM and S-MSGR, respectively (Figure 3C). Similarly, the final ratio of N. oceanica cultured in S-MSGR was about 0.18, which was much lower than the 0.83 obtained with EM (Figure 3D). The results indicate that the activity of chloroplast photosynthetic phosphorylation increased in S-MSGR, thus improving the photosynthetic efficiency of microalgae.
3.3. Metabolite Accumulation
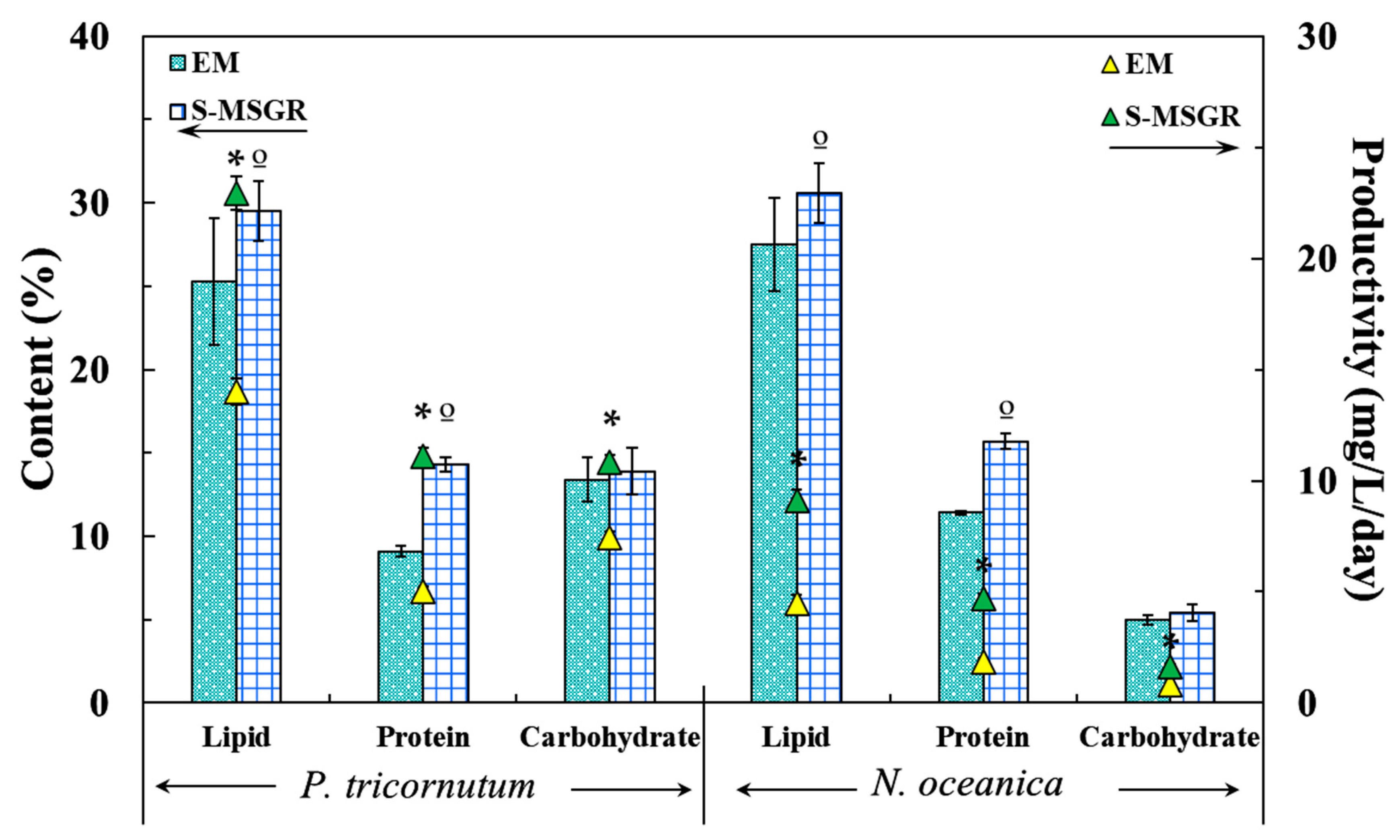
Figure 4 shows the accumulation of metabolites (lipid, protein and carbohydrate) by the two microalgae in EM medium and S-MSGR. It was found that the lipid content was the highest among the three metabolites synthesized by these two microalgae, exceeding one-quarter of the dry mass. The high salinity in EM medium and S-MSGR was the main stimulating factor for lipid synthesis.
For S-MSGR-cultured P. tricornutum, the lipid content and productivity reached 29.5% and 22.9 mg/L/day, which were 1.2 and 1.6 times higher than the values in standard medium (25.3% and 14.0 mg/L/day). The results indicate that P. tricornutum cultured in S-MSGR could potentially be used as a raw material for biofuels production. Although the content and productivity of protein synthesized by P. tricornutum in S-MSGR were not as high as those of lipids, the protein productivity (11.1 mg/L/day) was still doubled compared with that in EM medium (5.0 mg/L/day). The carbohydrate content in both cultures did not show an obvious difference but, considering the high biomass productivity in S-MSGR, the carbohydrate productivity in S-MSGR (10.8 mg/L/day) was 1.46 times higher than the 7.4 mg/L/day in EM medium.
Likewise, the main metabolite in N. oceanica was also lipid, accounting for one-third of the total dry biomass, indicating that lipid accumulation is a common approach to withstand the stimulation of high salinity, whether for limnetic or marine microalgae. The protein content and productivity of N. oceanica in S-MSGR also showed increases of 37.7% and 153% compared to those in EM medium, but carbohydrate synthesis was not affected by S-MSGR.
The main reason causing such phenomena is that P. tricornutum and N. oceanica demonstrated the capacity for heterotrophy, and the ample organic carbon in MSGR provided extra carbon sources apart from atmospheric CO2, which was conducive to promoting the accumulation of metabolites in microalgal cells [27]. Salinity was generally considered to be one of the triggers to changing the carbon partitioning from protein/carbohydrate synthesis to lipid synthesis, so that the lipid content was enhanced under exposure to high salinity [28]. However, in this study, both lipid and protein content of the two microalgae in S-MSGR increased, yet carbohydrate synthesis did not show a difference, indicating that there may be other metabolites in the cells providing carbon to lipid synthesis.
3.4. Benefit Analysis of S-MSGR for Culture of Marine Microalgae
A benefit analysis of S-MSGR for culture of P. tricornutum and N. oceanica was conducted herein, based on a comparison of biomass and lipid production performances found in the current work with those reported in the literature.
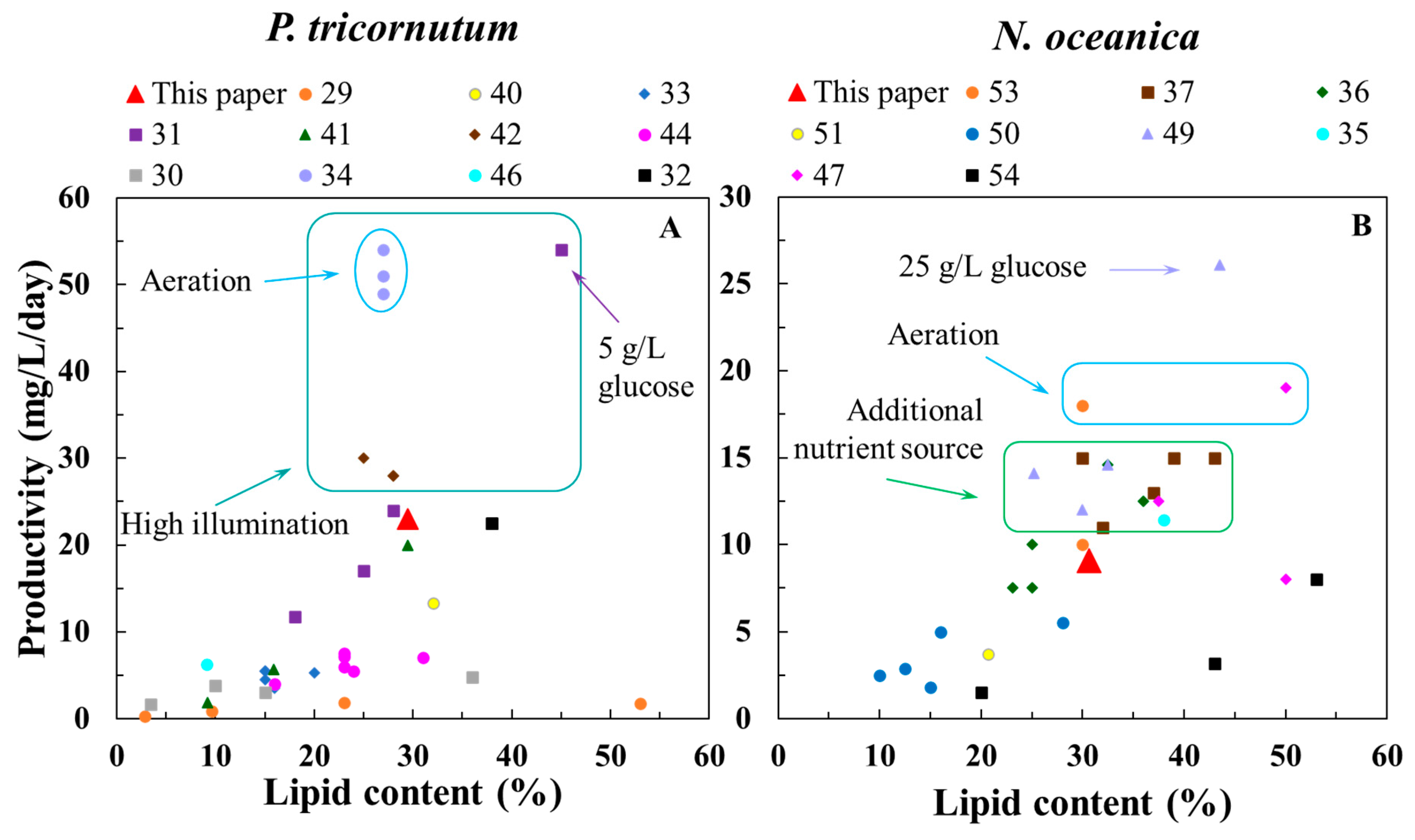
Figure 5 shows the lipid content and productivity reported by different studies on P. tricornutum and N. oceanica. At present, the culture medium for P. tricornutum was commonly f/2 medium or modified f/2 medium (Table S2). Yodsuwan et al. [29] added different concentrations of NaNO3 to f/2 medium for the cultivation of P. tricornutum, and the maximum lipid productivity was only 1.86 mg/L/day, which was far lower than that in S-MSGR. Notably, the lipid content in microalgal cells was low (2.8%) under the nitrogen-rich condition, and the corresponding biomass concentration was only 0.07 g/L. Similarly, Burch et al. [30] harvested the highest biomass concentration after culturing P. tricornutum in f/2 + 60% dairy manure wastewater, but the lipid content of algal cells was only 3.4%, which was far from meeting the demands of commercial application. Conversely, the total nitrogen content in S-MSGR was up to 179.4 mg/L, and the final lipid content of P. tricornutum increased to 22.9 mg/L/day, which suggested that S-MSGR as a nutrient source to cultivate P. tricornutum did not inhibit lipid synthesis in microalgae. Wang et al. [31] obtained the highest biomass concentration of P. tricornutum of 1.16 g/L with the help of high illumination and 5 g/L glucose, which increased the production cost. Simonazzi et al. [32] combined modified f/2 medium, effluent from anaerobic digestion and extra CO2 supplementation to cultivate P. tricornutum and obtained lipid productivity of 22.4 mg/L/day, which was close to one in this study. However, extra power consumption for the aeration device (0.6 L(CO2)/L/day) and high illumination (90–110 μmol/m2/s) increased the cultivation costs. Wang et al. [33] obtained 5.5 mg/L/day for the maximum lipid productivity of P. tricornutum in municipal wastewater, indicating that the municipal wastewater was not an ideal nutrient source. Su et al. [34] cultured P. tricornutum in wastewater from ultrafiltered digestate (UF) supplied with glycerol, which achieved 3.25 g/L and 51 mg/L/day of biomass concentration and lipid productivity, respectively. However, the extra aeration device and high light intensity (312 μmol/m2/s, versus 45 μmol/m2/s in our study) greatly increased the cost of microalgal culture.
Table S3 lists some studies on the biomass and lipid of N. oceanica. It can be found that there were few studies on cultivating N. oceanica with wastewater, and the medium for cultivating N. oceanica was mainly one of the standard media, such as f/2, BG11 or Conway medium. Compared with the experimental conditions in other studies, high illumination intensity and stable aeration may improve the biomass and lipid productivity of N. oceanica. Although the biomass and lipid productivity obtained by these studies [35,36,37] were close to those in the current study, the medium they all used was prepared with standard chemical reagents. In contrast, the S-MSGR medium greatly reduced the cost of nutrition during microalgal cultivation. In the present study, we used MSGR as a nutrient source to cultivate N. oceanica and obtained biomass and lipid productivities of 30 and 9.1 mg/L/day, respectively. The results in this study were not the highest among all studies, but the low illumination intensity, absence of an aeration device and cost-efficient nutrient resource greatly saved the culture cost, which will promote the commercial production of N. oceanica biomass.
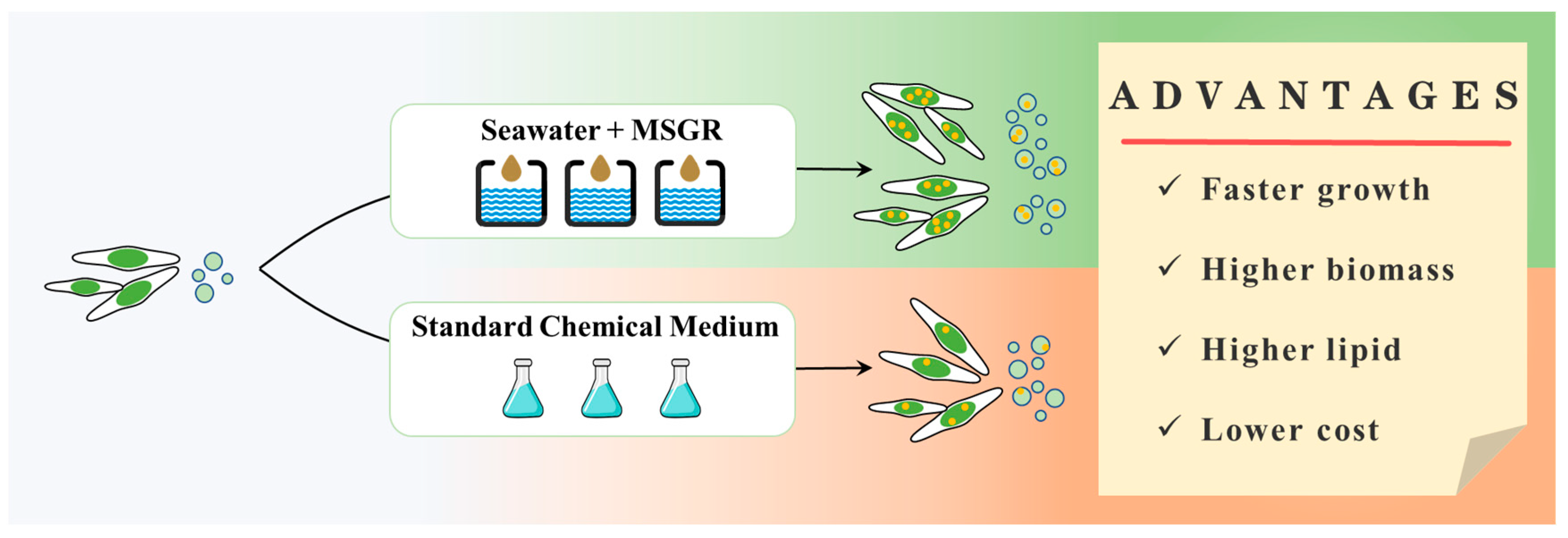
Figure 6 shows the advantages of biomass and biofuel production by P. tricornutum and N. oceanica in S-MSGR. S-MSGR was able to promote the fast growth of P. tricornutum and N. oceanica, and the biomass concentration trebled within the first five days, while in the standard medium (EM), it only doubled. P. tricornutum and N. oceanica cultured in S-MSGR obtained higher lipid productivities, which were, respectively, 1.6 times and 2 times higher than those obtained in EM.
To assess the economic benefits, the medium costs of biomass and lipid production of P. tricornutum and N. oceanica in EM and in S-MSGR were evaluated, without considering the cost of seawater needed for dilution of MSGR and preparation of EM. The cost of EM is mainly in chemical reagents, while the MSGR, as a waste nutrient source, has almost zero cost, so the main cost of S-MSGR medium was mainly from transportation. The medium costs of S-MSGR for biomass production of P. tricornutum and N. oceanica were USD 0.7 and 1.8/(kg biomass), which was merely 3.6% and 2.7% of the corresponding costs in EM. Similarly, the medium costs of S-MSGR for lipid production of P. tricornutum and N. oceanica were USD 2.3 and 5.8/(kg lipid), respectively. Compared with EM, 96.9% and 97.6% of the medium costs were saved for lipid production by P. tricornutum and N. oceanica cultured in S-MSGR. Hence, the S-MSGR medium was more economical for algal cultivation.
4. Conclusions
P. tricornutum and N. oceanica survived well in the seawater-based medium supplemented with MSGR at a ratio of 1/500. The increase in total chlorophyll content and the decrease of Chl-a/Chl-b showed that S-MSGR promoted photosynthesis in the microalgae. S-MSGR was more suitable for lipid production, and the lipid productivities of P. tricornutum and N. oceanica reached 22.9 and 9.1 mg/L/day, which was 1.6 and 2.0 times higher than those in EM. Both of the microalgae cultured in S-MSGR for biomass and lipid production saved more than 96% of the cost of standard culture medium. Hence, this study indicates that S-MSGR has a great promise as a medium for cultivating P. tricornutum and N. oceanica and bright prospects for use in commercial production.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w14060975/s1, Figure S1: The changes in total chlorophyll of (A) P. tricornutum and (B) N. oceanica, cultured in seawater supplemented with different levels of MSGR. Values are expressed as means ± s.d. (n = 3); Table S1: The nutritional assimilation of the two microalgae, P. tricornutum and N. oceanica, grown in EM and in S-MSGR; Table S2: Comparison of the biomass and lipid production performances of P. tricornutum cultured in S-MSGR with other results reported in the literature; Table S3: Comparison of the biomass and lipid production performances of N. oceanica cultured in S-MSGR with other results from the literature. References [38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54] are cited in the supplementary materials.
Author Contributions
Conceptualization, S.Y. and H.P.; methodology, S.Y.; software, S.Y.; validation, S.Y., Z.Y. and Q.H.; formal analysis, S.Y. and Z.Y.; investigation, S.Y.; resources, S.Y., Z.Y., Q.H. and H.P.; data curation, S.Y.; writing—original draft preparation, S.Y.; writing—review and editing, Z.Y. and H.P.; visualization, S.Y.; supervision, H.P.; project administration, H.P.; funding acquisition, H.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Department of Science and Technology of Shandong Province (Key R&D Program (Interna-tional Cooperation)), grant number: 2019GHZ030.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article or supplementary material.
Acknowledgments
This work was funded by the Department of Science and Technology of Shandong Province (Key R&D Program (International Cooperation)), grant number: 2019GHZ030. We would like to thank David Verrelli for kindly revising the grammar and language.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rizwan, M.; Mujtaba, G.; Memon, S.A.; Lee, K.; Rashid, N. Exploring the potential of microalgae for new biotechnology applications and beyond: A review. Renew. Sustain. Energy Rev. 2018, 92, 394–404. [Google Scholar] [CrossRef]
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.J.; Chang, J.S. Microalgae biorefinery: High value products perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Su, T.; Zhao, D.; Khodadadi, M.; Len, C. Lignocellulosic biomass for bioethanol: Recent advances, technology trends, and barriers to industrial development. Curr. Opin. Green Sustain. Chem. 2020, 24, 56–60. [Google Scholar] [CrossRef]
- Rodionova, M.V.; Bozieva, A.M.; Zharmukhamedov, S.K.; Leong, Y.K.; Lan, J.C.W.; Veziroglu, A.; Veziroglu, T.N.; Tomo, T.; Chang, J.S.; Allakhverdiev, S.I. A comprehensive review on lignocellulosic biomass biorefinery for sustainable biofuel production. Int. J. Hydrog. Energy 2021, 47, 1481–1498. [Google Scholar] [CrossRef]
- Butler, T.; Kapoore, R.V.; Vaidyanathan, S. Phaeodactylum tricornutum: A diatom cell factory. Trends Biotechnol. 2020, 38, 606–622. [Google Scholar] [CrossRef] [PubMed]
- Ashour, M.; Elshobary, M.E.; El-Shenody, R.; Kamil, A.W.; Abomohra, A.E.F. Evaluation of a native oleaginous marine microalga Nannochloropsis oceanica for dual use in biodiesel production and aquaculture feed. Biomass Bioenergy 2019, 120, 439–447. [Google Scholar] [CrossRef]
- Christenson, L.; Sims, R. Production and harvesting of microalgae for wastewater treatment, biofuels, and bioproducts. Biotechnol. Adv. 2011, 29, 686–702. [Google Scholar] [CrossRef]
- Shahid, A.; Malik, S.; Zhu, H.; Xu, J.; Nawaz, M.Z.; Nawaz, S.; Alam, A.; Mehmood, M.A. Cultivating microalgae in wastewater for biomass production, pollutant removal, and atmospheric carbon mitigation; a review. Sci. Total Environ. 2020, 704, 135303. [Google Scholar] [CrossRef]
- Mohsenpour, S.F.; Hennige, S.; Willoughby, N.; Adeloye, A.; Gutierrez, T. Integrating micro-algae into wastewater treatment: A review. Sci. Total Environ. 2021, 752, 142168. [Google Scholar] [CrossRef]
- Kumar, R.; Ghosh, A.K.; Pal, P. Synergy of biofuel production with waste remediation along with value-added co-products recovery through microalgae cultivation: A review of membrane-integrated green approach. Sci. Total Environ. 2020, 698, 134169. [Google Scholar] [CrossRef] [PubMed]
- Dębowski, M.; Zieliński, M.; Kisielewska, M.; Kazimierowicz, J.; Dudek, M.; Świca, I.; Rudnicka, A. The cultivation of lipid-rich microalgae biomass as anaerobic digestate valorization technology—A pilot-scale study. Processes 2020, 8, 517. [Google Scholar] [CrossRef]
- Kumar, R.; Chakrabortty, S.; Pal, P. Membrane-integrated physico-chemical treatment of coke-oven wastewater: Transport modelling and economic evaluation. Environ. Sci. Pollut. Res. 2015, 22, 6010–6023. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Sun, J.; Nie, C.; Li, Y.; Jenkins, J.; Pei, H. Filamentous cyanobacteria triples oil production in seawater-based medium supplemented with industrial waste: Monosodium glutamate residue. Biotechnol. Biofuels 2019, 12, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Rekha, P.D.; Arun, A.B.; Hameed, A.; Singh, S.; Shen, F.T.; Young, C.C. Glutamate wastewater as a culture medium for Azospirillum rugosum production and its impact on plant growth. Biol. Fertil. Soils 2011, 47, 419–426. [Google Scholar] [CrossRef]
- Ji, Y.; Hu, W.; Li, X.; Ma, G.; Song, M.; Pei, H. Mixotrophic growth and biochemical analysis of Chlorella vulgaris cultivated with diluted monosodium glutamate wastewater. Bioresour. Technol. 2014, 152, 471–476. [Google Scholar] [CrossRef]
- Correa, D.F.; Beyer, H.L.; Possingham, H.P.; Thomas-Hall, S.R.; Schenk, P.M. Global mapping of cost-effective microalgal biofuel production areas with minimal environmental impact. GCB Bioenergy 2019, 11, 914–929. [Google Scholar] [CrossRef] [Green Version]
- Pancha, I.; Chokshi, K.; George, B.; Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Nitrogen stress triggered biochemical and morphological changes in the microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2014, 156, 146–154. [Google Scholar] [CrossRef]
- Nie, C.; Jiang, L.; Yu, Z.; Yang, Z.; Hou, Q.; Pei, H. Campus sewage treatment by Golenkinia SDEC-16 and biofuel production under monochromic light. J. Chem. 2020, 2020, 5029535. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Pei, H.; Hu, W.; Ji, Y.; Han, L.; Ma, G. The feasibility of using complex wastewater from a monosodium glutamate factory to cultivate Spirulina subsalsa and accumulate biochemical composition. Bioresour. Technol. 2015, 180, 304–310. [Google Scholar] [CrossRef]
- Li, H.; Pei, H.; Xu, H.; Jin, Y.; Sun, J. Behavior of Cylindrospermopsis raciborskii during coagulation and sludge storage–higher potential risk of toxin release than Microcystis aeruginosa? J. Hazard. Mater. 2018, 347, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Bélanger-Lépine, F.; Tremblay, A.; Huot, Y.; Barnabé, S. Cultivation of an algae-bacteria consortium in wastewater from an industrial park: Effect of environmental stress and nutrient deficiency on lipid production. Bioresource technology. 2018, 267, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Pei, H.; Jiang, L.; Hou, Q.; Yu, Z. Toward facilitating microalgae cope with effluent from anaerobic digestion of kitchen waste: The art of agricultural phytohormones. Biotechnol. Biofuels 2017, 10, 76. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Song, M.; Pei, H.; Han, F.; Jiang, L.; Hou, Q. The growth characteristics and biodiesel production of ten algae strains cultivated in anaerobically digested effluent from kitchen waste. Algal Res. 2017, 24, 265–275. [Google Scholar] [CrossRef]
- Wang, X.W.; Liang, J.R.; Luo, C.S.; Chen, C.P.; Gao, Y.H. Biomass, total lipid production, and fatty acid composition of the marine diatom Chaetoceros muelleri in response to different CO2 levels. Bioresour. Technol. 2014, 161, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Demmig-Adams, B.; Adams, W.I. Chlorophyll and carotenoid composition in leaves of Euonymus kiautschovicus acclimated to different degrees of light stress in the field. Funct. Plant Biol. 1996, 23, 649–659. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Show, P.L.; Juan, J.C.; Chang, J.S.; Ling, T.C. Enhancing biomass and lipid productions of microalgae in palm oil mill effluent using carbon and nutrient supplementation. Energy Convers. Manag. 2018, 164, 188–197. [Google Scholar] [CrossRef]
- Ho, S.H.; Nakanishi, A.; Kato, Y.; Yamasaki, H.; Chang, J.S.; Misawa, N.; Hirose, Y.; Minagawa, J.; Hasunuma, T.; Kondo, A. Dynamic metabolic profiling together with transcription analysis reveals salinity-induced starch-to-lipid biosynthesis in alga Chlamydomonas sp. JSC4. Sci. Rep. 2017, 7, 45471. [Google Scholar] [CrossRef]
- Yodsuwan, N.; Sawayama, S.; Sirisansaneeyakul, S. Effect of nitrogen concentration on growth, lipid production and fatty acid profiles of the marine diatom Phaeodactylum tricornutum. Agric. Nat. Resour. 2017, 51, 190–197. [Google Scholar] [CrossRef]
- Burch, A.R.; Yothers, C.W.; Salemi, M.R.; Phinney, B.S.; Pandey, P.; Franz, A.K. Quantitative label-free proteomics and biochemical analysis of Phaeodactylum tricornutum cultivation on dairy manure wastewater. J. Appl. Phycol. 2021, 33, 2105–2121. [Google Scholar] [CrossRef]
- Wang, H.; Fu, R.; Pei, G. A study on lipid production of the mixotrophic microalgae Phaeodactylum tricornutum on various carbon sources. Afr. J. Microbiol. Res. 2012, 6, 1041–1047. [Google Scholar]
- Simonazzi, M.; Pezzolesi, L.; Guerrini, F.; Vanucci, S.; Samorì, C.; Pistocchi, R. Use of waste carbon dioxide and pre-treated liquid digestate from biogas process for Phaeodactylum tricornutum cultivation in photobioreactors and open ponds. Bioresour. Technol. 2019, 292, 121921. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Huang, L.; Ji, P.Y.; Chen, C.P.; Li, X.S.; Gao, Y.H.; Liang, J.R. Using a mixture of wastewater and seawater as the growth medium for wastewater treatment and lipid production by the marine diatom Phaeodactylum tricornutum. Bioresour. Technol. 2019, 289, 121681. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; D’Imporzano, G.; Veronesi, D.; Afric, S.; Adani, F. Phaeodactylum tricornutum cultivation under mixotrophic conditions with glycerol supplied with ultrafiltered digestate: A simple biorefinery approach recovering C and N. J. Biotechnol. 2020, 323, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Subasankari, K.; Thanappan, V.; Jeyapragash, D.; Anantharaman, P.; Sarangi, R.K. Growth promoting studies on co-culturing Nannochloropsis oceanica with Halomonas aquamarina actively enhance the algal biomass and lipid production. Biocatal. Agric. Biotechnol. 2020, 29, 101790. [Google Scholar]
- Mitra, M.; Patidar, S.K.; George, B.; Shah, F.; Mishra, S. A euryhaline Nannochloropsis gaditana with potential for nutraceutical (EPA) and biodiesel production. Algal Res. 2015, 8, 161–167. [Google Scholar] [CrossRef]
- Ma, X.; Liu, J.; Liu, B.; Chen, T.; Yang, B.; Chen, F. Physiological and biochemical changes reveal stress-associated photosynthetic carbon partitioning into triacylglycerol in the oleaginous marine alga Nannochloropsis oculata. Algal Res. 2016, 16, 28–35. [Google Scholar] [CrossRef]
- Pal, D.; Khozin-Goldberg, I.; Didi-Cohen, S.; Solovchenko, A.; Batushansky, A.; Kaye, Y.; Sikron, N.; Samani, T.; Fait, A.; Boussiba, S. Growth, lipid production and metabolic adjustments in the euryhaline eustigmatophyte Nannochloropsis oceanica CCALA 804 in response to osmotic downshift. Appl. Microbiol. Biotechnol. 2013, 97, 8291–8306. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Cheng, J.; Xu, X.; Lu, H.; Wang, Y.; Li, X.; Yang, W. Using polyethylene glycol to promote Nannochloropsis oceanica growth with 15 vol% CO2. Sci. Total Environ. 2020, 720, 137598. [Google Scholar] [CrossRef]
- Yang, Y.H.; Du, L.; Hosokawa, M.; Miyashita, K.; Kokubun, Y.; Arai, H.; Taroda, H. Fatty acid and lipid class composition of the microalga Phaeodactylum tricornutum. J. Oleo Sci. 2017, 66, 363–368. [Google Scholar] [CrossRef] [Green Version]
- KaiXian, Q.; Borowitzka, M.A. Light and nitrogen deficiency effects on the growth and composition of Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 1993, 38, 93–103. [Google Scholar] [CrossRef]
- Sharma, N.; Fleurent, G.; Awwad, F.; Cheng, M.; Meddeb-Mouelhi, F.; Budge, S.M.; Germain, H.; Desgagné-Penix, I. Red light variation an effective alternative to regulate biomass and lipid profiles in Phaeodactylum tricornutum. Appl. Sci. 2020, 10, 2531. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yin, W.; Ma, D.; Liu, X.; Xu, K.; Liu, J. Phytohormone supplementation significantly increases fatty acid content of Phaeodactylum tricornutum in two-phase culture. J. Appl. Phycol. 2021, 33, 13–23. [Google Scholar] [CrossRef]
- Wu, H.; Li, T.; Wang, G.; Dai, S.; He, H.; Xiang, W. A comparative analysis of fatty acid composition and fucoxanthin content in six Phaeodactylum tricornutum strains from different origins. Chin. J. Oceanol. Limnol. 2016, 34, 391–398. [Google Scholar] [CrossRef]
- Huete-Ortega, M.; Okurowska, K.; Kapoore, R.V.; Johnson, M.P.; Gilmour, D.J.; Vaidyanathan, S. Effect of ammonium and high light intensity on the accumulation of lipids in Nannochloropsis oceanica (CCAP 849/10) and Phaeodactylum tricornutum (CCAP 1055/1). Biotechnol. Biofuels 2018, 11, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branco-Vieira, M.; San Martin, S.; Agurto, C.; Santos, M.A.D.; Freitas, M.A.; Mata, T.M.; Martins, A.A.; Caetano, N.S. Potential of Phaeodactylum tricornutum for biodiesel production under natural conditions in Chile. Energies 2018, 11, 54. [Google Scholar] [CrossRef] [Green Version]
- Wan, C.; Bai, F.W.; Zhao, X.Q. Effects of nitrogen concentration and media replacement on cell growth and lipid production of oleaginous marine microalga Nannochloropsis oceanica DUT01. Biochem. Eng. J. 2013, 78, 32–38. [Google Scholar] [CrossRef]
- Wang, Z.; Cheng, J.; Li, K.; Zhu, Y.; Liu, J.; Yang, W.; Xu, J.; Park, J.Y. Comparison of photosynthetic carbon fixation of Nannochloropsis oceanica cultivated with carbon suppliers: CO2, NaHCO3 and CH3OH. J. CO2 Util. 2020, 41, 101235. [Google Scholar] [CrossRef]
- Mitra, M.; Mishra, S. Effect of glucose on growth and fatty acid composition of an euryhaline eustigmatophyte Nannochloropsis oceanica under mixotrophic culture condition. Bioresour. Technol. Rep. 2018, 3, 147–153. [Google Scholar] [CrossRef]
- Touliabah, H.E.S.; Almutairi, A.W. Effect of phytohormones supplementation under nitrogen depletion on biomass and lipid production of Nannochloropsis oceanica for integrated application in nutrition and biodiesel. Sustainability 2021, 13, 592. [Google Scholar] [CrossRef]
- Li, T.; Chen, Z.; Wu, J.; Wu, H.; Yang, B.; Dai, L.; Wu, H.; Xiang, W. The potential productivity of the microalga, Nannochloropsis oceanica SCS-1981, in a solar powered outdoor open pond as an aquaculture feed. Algal Res. 2020, 46, 101793. [Google Scholar] [CrossRef]
- Savvidou, M.G.; Sotiroudis, T.G.; Kolisis, F.N. Cell surface and cellular debris-associated heat-stable lipolytic enzyme activities of the marine alga Nannochloropsis oceanica. Biocatal. Biotransform. 2016, 34, 24–32. [Google Scholar] [CrossRef]
- Ra, C.H.; Sirisuk, P.; Jung, J.H.; Jeong, G.T.; Kim, S.K. Effects of light-emitting diode (LED) with a mixture of wavelengths on the growth and lipid content of microalgae. Bioprocess Biosyst. Eng. 2018, 41, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Ma, Y.; Wei, W.; Liu, W.; Ding, Y.; Balamurugan, S. Sequential treatment with bicarbonate and low-temperature to potentiate both biomass and lipid productivity in Nannochloropsis oceanica. J. Chem. Technol. Biotechnol. 2019, 94, 3413–3419. [Google Scholar] [CrossRef]
Figure 1.
Changes in biomass concentration and the biomass productivity of the two microalgae, P. tricornutum (A) and N. oceanica (B), in EM and in S-MSGR. Values are expressed as means ± s.d. (n = 3).
Figure 1.
Changes in biomass concentration and the biomass productivity of the two microalgae, P. tricornutum (A) and N. oceanica (B), in EM and in S-MSGR. Values are expressed as means ± s.d. (n = 3).

Figure 2.
The nutritional assimilation and the average specific growth rate (μ, day−1) of P. tricornutum and N. oceanica grown in standard medium (EM) and in S-MSGR. Values are expressed as means ± s.d. (n = 3). Values with the same letter (a, b, c, d; A, B, C or α, β, γ) show no significant difference of N-AYC, P-AYC or μ between groups, respectively (p > 0.05, Duncan’s test).
Figure 2.
The nutritional assimilation and the average specific growth rate (μ, day−1) of P. tricornutum and N. oceanica grown in standard medium (EM) and in S-MSGR. Values are expressed as means ± s.d. (n = 3). Values with the same letter (a, b, c, d; A, B, C or α, β, γ) show no significant difference of N-AYC, P-AYC or μ between groups, respectively (p > 0.05, Duncan’s test).

Figure 3.
Changes in total chlorophyll content (A,B) and ratios of Chl-a/Chl-b (C,D) of the two marine microalgae, P. tricornutum and N. oceanica, grown in EM and in S-MSGR. Values are expressed as means ± s.d. (n = 3).
Figure 3.
Changes in total chlorophyll content (A,B) and ratios of Chl-a/Chl-b (C,D) of the two marine microalgae, P. tricornutum and N. oceanica, grown in EM and in S-MSGR. Values are expressed as means ± s.d. (n = 3).

Figure 4.
The content and productivity of lipids, proteins and carbohydrates for P. tricornutum and N. oceanica, grown in EM and in S-MSGR. Values are expressed as means ± s.d. (n = 3). The symbol * indicates the significant difference in the productivity groups (p < 0.05), The symbol º indicates the significant difference in the content groups (p < 0.05).
Figure 4.
The content and productivity of lipids, proteins and carbohydrates for P. tricornutum and N. oceanica, grown in EM and in S-MSGR. Values are expressed as means ± s.d. (n = 3). The symbol * indicates the significant difference in the productivity groups (p < 0.05), The symbol º indicates the significant difference in the content groups (p < 0.05).

Figure 5.
Comparison of the lipid content and lipid production performances of (A) P. tricornutum and (B) N. oceanica cultured in S-MSGR with other results from the literature. The legend indicates the references for the data.
Figure 5.
Comparison of the lipid content and lipid production performances of (A) P. tricornutum and (B) N. oceanica cultured in S-MSGR with other results from the literature. The legend indicates the references for the data.

Figure 6.
The benefits of using S-MSGR as a medium for biomass and biofuel production with P. tricornutum and N. oceanica.
Figure 6.
The benefits of using S-MSGR as a medium for biomass and biofuel production with P. tricornutum and N. oceanica.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yu, S.; Yu, Z.; Hou, Q.; Pei, H. Seawater with Added Monosodium Glutamate Residue (MSGR) Is a Promising Medium for the Cultivation of Two Commercial Marine Microalgae. Water 2022, 14, 975. https://doi.org/10.3390/w14060975
AMA Style
Yu S, Yu Z, Hou Q, Pei H. Seawater with Added Monosodium Glutamate Residue (MSGR) Is a Promising Medium for the Cultivation of Two Commercial Marine Microalgae. Water. 2022; 14(6):975. https://doi.org/10.3390/w14060975
Chicago/Turabian StyleYu, Siteng, Ze Yu, Qingjie Hou, and Haiyan Pei. 2022. "Seawater with Added Monosodium Glutamate Residue (MSGR) Is a Promising Medium for the Cultivation of Two Commercial Marine Microalgae" Water 14, no. 6: 975. https://doi.org/10.3390/w14060975
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.