
Plankton under Pressure: How Water Conditions Alter the Phytoplankton–Zooplankton Link in Coastal Lagoons


1
Department of Integrative Marine Ecology, Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy
2
Ph.D. Program in Evolutionary Biology and Ecology, Department of Biology, University of Rome ‘Tor Vergata’, Via Della Ricerca Scientifica 1, 00133 Roma, Italy
3
Department of Biology and Evolution of Marine Organisms, Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy
*
Authors to whom correspondence should be addressed.
Water 2022, 14(6), 974; https://doi.org/10.3390/w14060974
Submission received: 23 December 2021
/
Revised: 14 March 2022
/
Accepted: 17 March 2022
/
Published: 19 March 2022
(This article belongs to the Special Issue The Phytoplankton-Zooplankton Link under Anthropogenic Pressures)
Abstract
:Transitional waters (TWs), such as coastal lagoons, are bodies of surface water at the transition between saline and freshwater domains. These environments play a vital role in guaranteeing ecosystem services, including provision of food, protection against meteorological events, as anthropogenic carbon sinks, and in filtering of pollutants. Due to the escalating overpopulation characterising coastlines worldwide, transitional systems are over-exploited, degraded, and reduced in their macroscopic features. However, information on the impact of anthropogenic pressures on planktonic organisms in these systems is still scanty and fragmented. Herein, we summarise the literature, with a special focus on coastal lagoons undergoing anthropogenic pressure. Specifically, we report on the implications of human impacts on the ecological state of plankton, i.e., a fundamental ecological component of aquatic ecosystems. Literature information indicates that human forces may alter ecosystem structures and functions in coastal lagoons, as in other TWs such as estuaries, hampering the phytoplankton–zooplankton link, i.e., the main trophic process occurring in those communities, and which sustains aquatic productivity. Changes in the dominance and lifestyle of key planktonic players, plus the invasion of ‘alien’ species, and consequent regime shifts, are among the most common outcomes of human disturbance.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Transitional waters (TWs) are bodies of surface water which are partly saline due to their proximity to coastal waters but which are substantially influenced by freshwater flows [1]. TWs offer exceptional services and are good providers of tangible, economic, and intangible values [2,3,4]. These systems are characterised by myriad habitats, from open waters to submerged aquatic vegetation, from tidal flats to fringing wetlands, which may act as nursery, feeding, and refuge grounds for several organisms, including terrestrial ones, with some also having high recreational and commercial value [5]. TWs are not only important to humans for fisheries and aquaculture, but are also used for electric power generation, biotechnology, transportation, and shipping, not to mention the recreational value of these ecosystems [2]. Moreover, TWs provide key chemo-physical functions in trapping, transforming, and filtering nutrients and wastes, thus strongly influencing the environmental status of surrounding areas [6].
One feature of TWs is almost unique among coastal systems, namely that terrestrial sources such as freshwater and land drainage can be more important than marine ones in providing them with nutrients [7]; this specific trait makes understanding the factors driving coastal ecosystem productivity extremely complex. Coastal lagoons are among the most productive TW ecosystems, since freshwater inputs stimulate fast biomass accumulation in photosynthetic organisms, thus fuelling the transfer of organic matter to animals, including megafauna [8]. Moreover, the ecological functioning of these systems is highly diverse even at the regional scale. The multifaceted geomorphological features that we can find at the land–sea interface determine different physical and chemical drivers, which lead, for example, to the asynchronous increase in primary production in geographical sites belonging to the same climatic region [9]. For instance, primary productivity at the local scale is further affected by the asymmetric impact of climate change and anthropogenic pressures, which act at different scales, from the ecosystem to species levels (and back) with high temporal and spatial irregularities [10,11].
TWs are highly diverse and can undergo different modifications in response to anthropogenic pressures and climate crises. For instance, precipitations are thought to be declining over the long term and this fact not only reduces land inflows, but also causes the progressive hyper-salinisation of TWs, the closure of periodically open systems and, consequently, dramatic ecological changes [12]. Weakly-flushed microtidal estuaries such as those in the Mediterranean are less ecologically robust than macrotidal estuaries of the Atlantic coast and this exacerbates the stress conditions induced in aquatic communities by pollution [13]. Model simulations predicted that Mediterranean semi-enclosed waterbodies would suffer from ecological crisis stemming from the combined effect of warmer waters, altered precipitation regimes, and unbalanced primary production cycles [14]. According to Pesce et al. (2018) [14], in the “Palude di Cona”, a water body in the Venice Lagoon, modelled riverine nutrient discharge shows a tendency to increase during winter and decrease during summer, due to, respectively, increased precipitation events and drought, along with an overall increase in temperature; this would provoke more frequent events of eutrophication—i.e., an increase in photo-autotrophic biomass due to nutrient enrichment—potentially altering the ecosystem state [15]. At the same time, these conditions could drive the rise to dominance of nitrogen-fixing cyanobacteria, in turn affecting trophic processes, by favouring microbial consumers over metazoans.
Although much of the research on ecological responses to environmental drivers in TWs has focused on Mediterranean and temperate systems [16], investigations of subtropical and tropical systems have also been carried out (e.g., [17,18]). Tarafdar et al., 2021 [17] observed that the dynamics of photo-autotrophic biomass in Chilika Lagoon, India, were mainly influenced by changes in parameters affected by anthropogenic activities such as salinity and temperature rather than by the seasons’ alternation. Similar responses were found in the subtropical Patos Lagoon in Brazil, where, from a long-term analysis, it was observed that freshwater discharge influenced by climate change was the main driver of primary productivity variability [18].
Given the ecological complexity of TWs, an understanding of the patterns of changes in the plankton community can only be achieved through a proper sampling program, which is able to capture changes in the different components of the community (bacterio-, phyto-, and zoo-plankton) at different spatial and temporal scales. On the other hand, even a proper sampling may be not enough to unveil the possible causative factors behind those changes. Climate forcing and anthropogenic pressures act on TW ecosystems in both directions, from land and from the sea, by modifying the quality and quantity of river discharge and by altering the physical-chemical conditions and coastal hydrodynamics. On top of all of this, climatic oscillations can affect both hydrology and hydrodynamics, modifying the portion of freshwater and marine waters in TWs, and thus their ecological state [19].
Building on the considerations above, in the present review, we provide a synthesis of the impact of anthropogenic threats on coastal lagoons and their effect on plankton therein, i.e., on those primary producers (i.e., phytoplankton) and consumers (i.e., protozoan and metazoan plankton) that play a pivotal role as main drivers of biogeochemical cycles in aquatic trophic webs, and which typically respond very quickly to perturbations due to physical (e.g., hydrodynamic, thermal, and chemical) stressors [20] (Figure 1). We discuss the potential of anthropogenic stressors to perturb the state of the phytoplankton–zooplankton link, which is fundamental in guarantying aquatic productivity.
2. Coastal Lagoons as Key Study Case for TWs
Lagoons are relatively small bodies of water that are semi-isolated from the sea due to the development of natural or human-made barriers. From a pure geoscience perspective, coastal lagoons may be considered as essentially ephemeral features within a continuum of coastal environments [21]. They are geologically young systems, mostly formed over the last 5000–7000 years, and are often short-lived in geological timescales because the same rivers that produce lagoons drive sedimentation therein [22].
Coastal lagoons cover only about 13% of the world’s coastlines [23]; nonetheless, they represent hotspots of biodiversity, productivity, and environmental changes, which may range from moderately to highly dynamic. Lagoons are generally characterised by very shallow waters (only a few meters deep), which often prevent water column stratification, and are distributed worldwide, showing a vast range of physical and biotic types. For example, the south-eastern coast of the United States can be viewed as a series of lagoons and estuaries, extending for ca. 4500 km along the coast, while the largest lagoon in the United Kingdom (the Fleet Lagoon in Dorset, England), has an extension of only 14 km.
The natural history of lagoons is intimately connected to the formation of the barriers enclosing them, which are, broadly speaking, of two types: enclosure of lagoons by offshore and longshore barriers, or bars [24]. Offshore bars are produced by wave action on shallow waters, causing a deposition of sands (e.g., the Laguna Madre, in Texas). Alternatively, barrier formation can extend out from the land, as for Vistula lagoon in Poland [24]. Although most of the lagoons worldwide are thought to originate from one of the mechanisms outlined above, other mechanism may have been locally important (see, e.g., [24]).
Therefore, considering the interplay between sea and land waters, costal lagoons may display a wide range of salinity regimes: from hyposaline (e.g., the Vistula lagoon), to hypersaline (e.g., the Laguna Madre), or, rarely, show salinity gradients even in a very small area (e.g., the Swanpool, Cornwall, UK) [24]. Along with salinity, the nutrient gradient may be considered the main characteristic that defines a lagoon. Both salinity and nutrients depend mostly on the proportion of freshwater to sea water [25,26].
As for the fauna, coastal lagoons are dominated by a few species of permanent residents, and the population abundance of individual species varies considerably from year to year, becoming very copious under favourable conditions [27]. Motile species migrate into lagoons to feed, reproduce, and use the protected waters as a nursery and refuge. Lagoons also represent nursery and reproduction areas for many marine species, thus performing a pivotal role in biodiversity maintenance and also in the productivity of marine and fluvial adjacent systems.
Finally, lagoons are natural filters through which continental organic matter and nutrients pass and are metabolised [28], thus providing a vast range of ecosystem services and resources with major roles in local economies and communities, such as in food provisioning (mainly fish and shellfish), freshwater storage, hydrological balance, climate regulation, flood protection, water purification, oxygen production, fertility, recreation, and ecotourism [29]. Historically, this fact promoted human settlement and urbanisation in nearby areas, which, along with the generally high residence time of lagoon waters, is one of the main causes of the high levels of environmental degradation therein.
Impact of Hydrodynamic Changes on Lagoon Plankton
Hydrodynamic changes are among the main driver of environmental stress in lagoon ecosystems [30,31,32,33,34,35,36,37,38,39,40,41,42]. These changes can be directly connected with anthropogenic drivers when human activities transform lagoons’ geomorphology with infrastructure. Land-use and building construction, inappropriate defence structures, or artificial lagoon openings may all cause beach erosion, such as in the lagoon-estuarine system of Isla del Carmen (Mexico) [30]. A large scale inland irrigation project caused an increase in freshwater inflow into the Kalametiya Lagoon, Sri Lanka; as a consequence, this lagoon has almost disappeared [31].
Hydrodynamic changes in coastal lagoon systems are also related to sea-level rise (SLR) driven by global warming, despite difficulties in converting future global SLR projections to strictly local events [32]. Morphological differences in coasts make it difficult to understand the response of TWs to SLR, and to set mitigation strategies. SLR has been studied in various TWs worldwide [33,34,35,36,37,38], but the difficulty of predicting the local influence of the relative SLR (RSLR) still persists, mainly due to the non-linearity of the coastal sedimentary processes [32], and to the different spatial scales of the coastal morphologies, ranging from bed forms (~cm to m), to full basins (~1–100 km) [39,40]. In the Venice Lagoon (Italy), anthropogenic structures built to protect the historical city from SLR, and ship traffic (propeller-wash erosion, keel ploughing), are rapidly altering the lagoon seafloor [41]. A complex array of large mobile barriers (the MoSE system) affected the hydrodynamics of the lagoon mainly by causing more intense current speeds and altering the micro-circulation [42]. It is worth noting that mechanisms of resistance and resilience to human-driven stressors are known to exist but have not been well studied [43]. Moreover, most of the studies above, regardless of the different regional and local conditions, identify a ‘defence’ system against RSLR, in which landward barriers retreat through continuous migration [32].
Hydrodynamic modifications can alter the abiotic environment in lagoons, i.e., by modifying water temperature and salinity, the absolute amount of inorganic nutrients present in the water, and ultimately planktonic assemblages and the overall ecosystem state, even though disentangling natural from anthropogenic drivers of ecological changes in lagoons is particularly challenging [44]. Among natural drivers, we highlight that even pristine coastal lagoons can differ in terms of water level, salinity, temperature, and the extent of nutrient loads. All these characteristics influence the residing ecological community dramatically. A comparative study on three North African coastal lagoons showed that salinity differences between lagoons is a main driving factor of the dominance of plankton species [45]. The seawater dominated lagoons Merja Zerga and Ghar El Melh showed a quasi-permanent predominance of marine diatoms and dinoflagellates, while in the freshwater Lake Manzala, Chlorophyceae contributed 39% of the total species. Similarly, zooplankton communities in both Ghar El Melh and Merja Zerga were dominated by marine copepods, while Lake Manzala was dominated by rotifers. A study conducted on the zooplankton of the Mossoró River Estuary (north-eastern coast of Brazil) along a horizontal salinity gradient and during both the dry and the rainy seasons, suggested the idea that the dominant species in such environment are plastic species that are able to cope with harsh spatial and seasonal variations [46]. The Coorong is a shallow saline coastal lagoon in Australia; herein, during high freshwater flow, phytoplankton–zooplankton interactions dominate the food web, while at low flow, bacteria, viruses, and nano/picoplankton interactions are more dominant [47]. However, it is of note that a study conducted in the Vistula Lagoon correlated changes in zooplankton abundance and biodiversity not with variation in a parameter such as salinity, but with more complex multi-parameter variations linked to the dry and rainy seasons [48].
In addition, the plankton composition in lagoon ecosystems changes greatly throughout the year, as a consequence of various physical factors such as water regime changes. For example, in the largest lagoon in Asia (Chilika, India), Bacillariophyta (138 species) are the most diverse and abundant phytoplankton group, but they are replaced by euglenoids under stronger freshwater discharge [17]. In the Mar Menor lagoon, the annual succession of phytoplankton is marked: the winter season displays a dominance of small phytoplankters, followed by a diatoms-dominated community during spring and summer seasons, with some monospecific diatom blooms, and a post-summer period dominated by dinoflagellates [49]. In the Mar Menor, copepods remain relatively constant through the year, but with a lower density from July to September: the autotrophs to heterotrophs biovolume ratio is constantly lower than 1, with the exception of phytoplankton bloom periods [49]. Thanks to the background knowledge on the seasonal variation of plankton composition in Mar Menor, long-term ecological changes have been related to anthropogenic variations in nutrient inputs from land [50]. Changes in zooplankton appeared less dramatic therein; however, an increase in the abundance of gelatinous zooplankton (such as the Appendicularia Oikopleura dioica and jellyfish species, in the years 1988 and 1997) was recorded [50].
Furthermore, to evaluate anthropogenic impacts on coastal lagoons it is necessary to take into consideration water level changes, which can alter the benthic-pelagic coupling, considering the shallowness of those systems. Indeed, the primary production of both benthos and plankton in lagoons is high, resulting in quantitatively important pelagic and benthic food chains, which support rich fisheries and aquaculture operations [51], from fish to seaweeds and to bivalve molluscs. A comparative study on three different coastal lagoons in Italy [52] demonstrated that consumer dependence on benthic and pelagic organic matter showed high variability. Another study on a shallow coastal lagoon (mean depth of the sampling sites = 0.7 m) on the Rhode Island (USA) coastal area demonstrated [53] that the benthos may consume up to 50% of the primary production. Interestingly, this percentage is not statistically too far from those seen in the somehow similar but deeper (ca. 10 m depth) Narragansett Bay [53]; however, in these waters, the phytoplankton in the water column are the main primary producers; conversely, in the case of shallow coastal lagoons, plankton, macroflora, and benthic epiflora are altogether the main actors.
The tight proximity of benthic and pelagic dominions in coastal lagoons exacerbates the effects of environmental changes on lagoon ecosystems, which can undergo marked ‘regime shifts’ [54]. Of note, changes in marine benthic macrophytic species have been used as a classification scheme to evaluate ecological status in Greek coastal lagoons [55]. Many factors may contribute to regime shift in lagoons. Nitrogen is often considered as the main limiting factor that controls the benthic vegetation, especially in temperate systems [56]. Eutrophication can favour the rapid growth and/or colonisation ability of seaweeds to exclude angiosperms, with the coexistence of the two at the transition from pristine to altered conditions [57]. Empirical and experimental data collected on the California lagoons system demonstrated that increased nitrogen loading caused a shift in producers from lowly abundant phytoplankton and microphytobenthos to blooms of opportunistic green macroalgae [58]. However, knowledge is also increasing regarding the contribution of planktonic organisms to regime shifts in lagoons, which must be evaluated whilst also taking into consideration the intrinsic variability of planktonic communities within those closed systems.
Coastal lagoons are frequently investigated using the principles of system ecology, in which biotic and abiotic observations are integrated in a common conceptual framework with the aim of establishing links between drivers of changes and their effects. Physical modification of the nearby wetlands, along with pollution, is causing degradation of the Sakumo Lagoon, Ghana [59]. Land-use changes have caused eutrophication and enhanced anoxic conditions in the Etoliko lagoon (Greece) [60]. In the Mar Menor lagoon in the Mediterranean Sea, the enlargement of the main canal connecting the system to open waters in 1972 resulted in decreased salinity, increased water renewal rate, and decreased extreme temperatures, enabling the lagoon’s colonisation by typically Mediterranean organisms [50], such as the jellyfishes Cotylorhiza tuberculata and Rhizostoma pulmo, which became pest species therein [61], and an influx of Caulerpa prolifera (a seaweed), which is now the dominant benthic species [50]. However, increased discharges from agriculture have more recently driven the Mar Menor towards an eutrophic state [62]. Interestingly, C. prolifera dominance in Mar Menor has likely delayed the eutrophication of the lagoon for some time, by preventing high phytoplankton densities [63].
3. Rewiring of Planktonic Trophic Networks in Impacted TWs
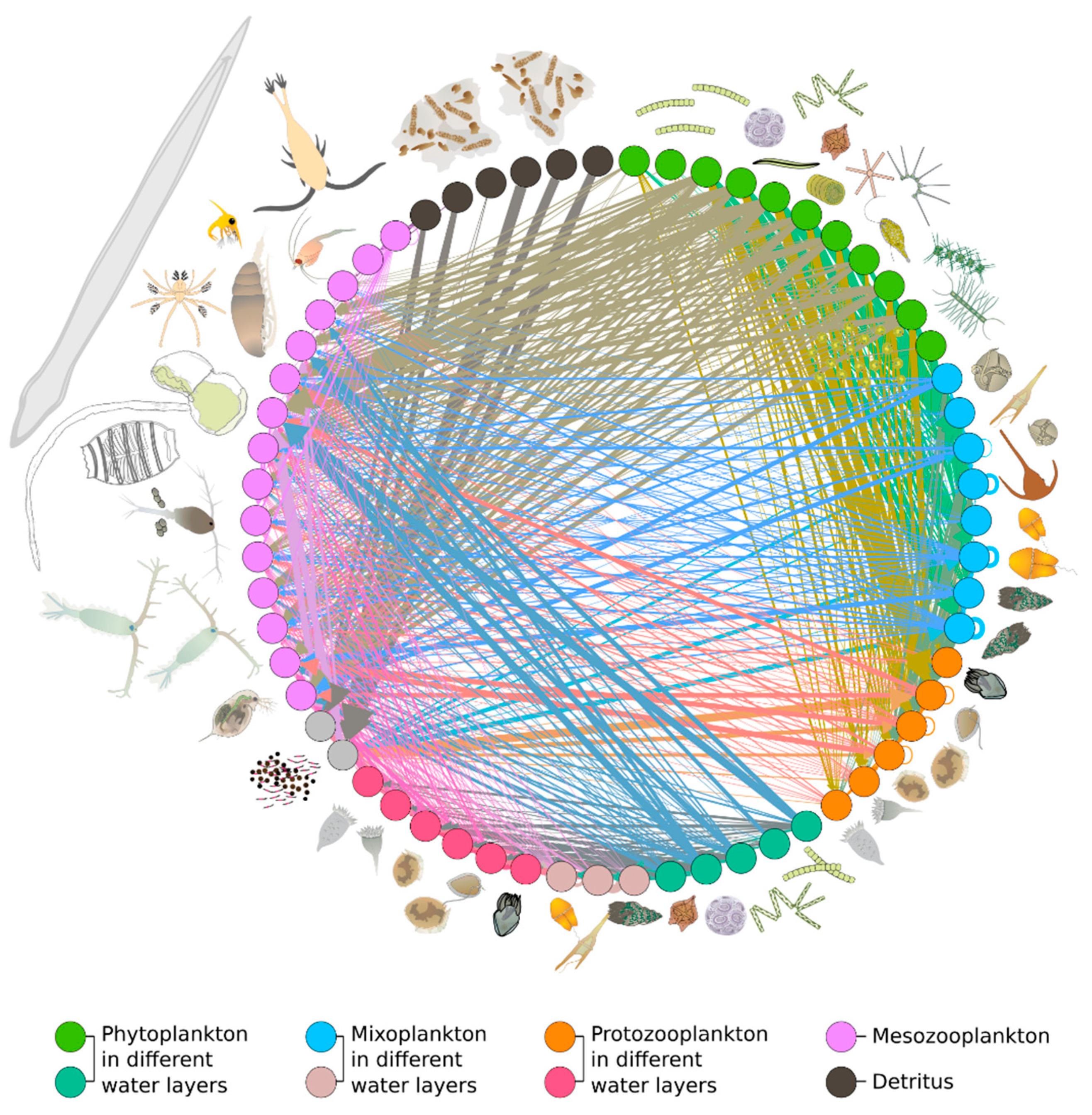
The overwhelming biological complexity of plankton communities gives rise to multiple and overlapping trophic links, which involve primary producers of different sizes and consumers that can be both unicellular and metazoan. The resulting trophic interaction networks are highly tangled and potentially regulated by indirect effects that can propagate from species to community levels [64,65] (Figure 2).
Recent research carried out in a coastal system (the Gulf of Naples) at the boundary between freshwater and offshore domains revealed that the organisation of plankton trophic networks can change quickly (i.e., at weekly time scales) in response to changes in the trophic state of the system [64,65]: for instance, eutrophic conditions—i.e., ‘green waters’ with lower salinity, higher nutrient inputs, and subsequent increases in phytoplankton blooms—favoured typical herbivorous pathways, with a straighter link between micro-phytoplankton and meso-zooplankton and a lower contribution of proto-zooplankton (i.e., heterotrophic protists) to the diet of planktonic metazoans; conversely, oligotrophic conditions—i.e., ‘blue waters’ with higher salinity, and with lower nutrient concentrations, phytoplankton biomass, and average phytoplankton size—favoured microbial grazing and were characterised by an increased contribution of heterotrophic protists to the mesozooplankton diet. Such trophic shifts are thought to stabilise planktonic networks against the short-term trophic shifts occurring along the coast due to intermittent runoff [64,65]. Indeed, it was estimated that seven-fold differences in phytoplankton biomass between blue-water and green-water states determined only two-fold differences in the flow of organic matter to mesozooplankton; i.e., thanks to microbial grazing, the trophic web was highly efficient even under condition of lower phytoplankton biomass.
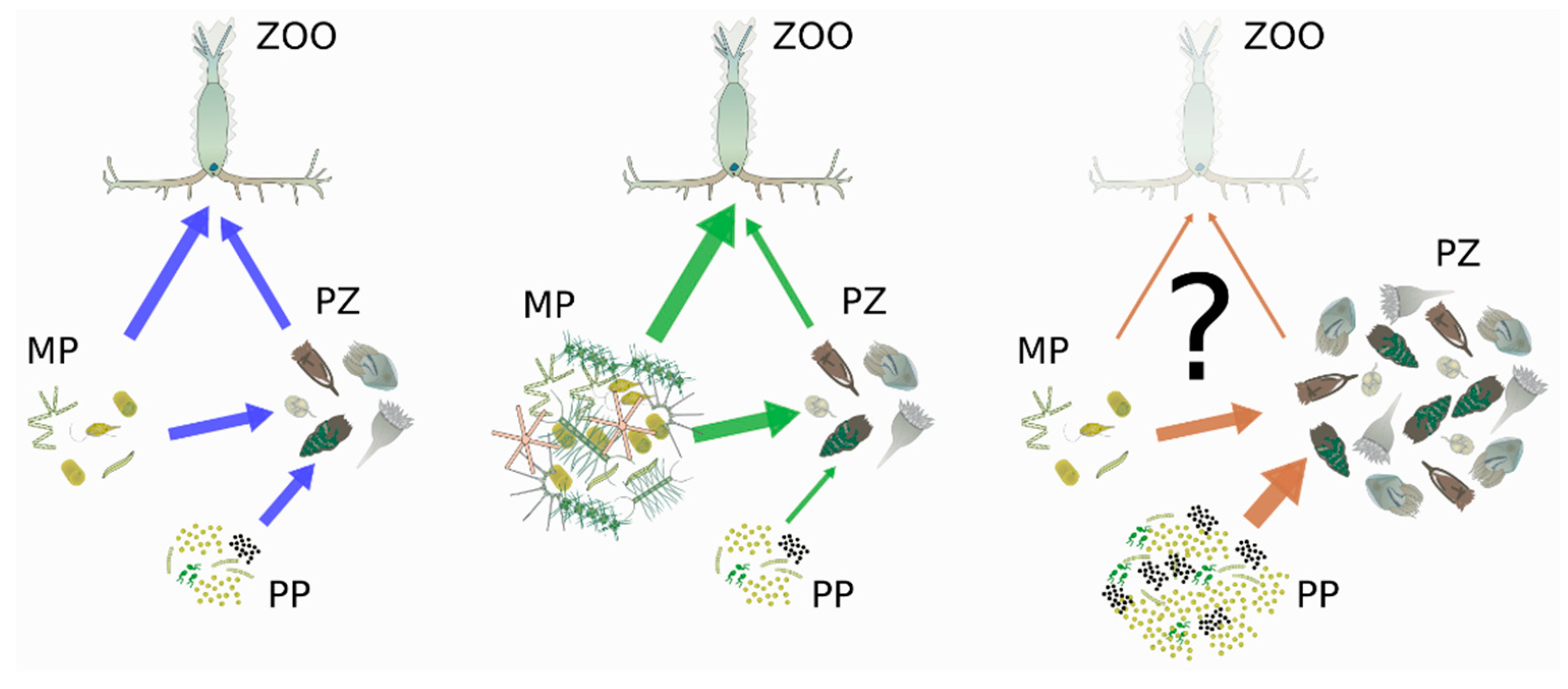
Similar trophic dynamics are likely to occur in coastal lagoons too, based on information available from those systems, where both natural and anthropogenic freshwater inputs impact the plankton community, with implications for trophic networks. Groundwater discharge was found to stimulate primary production through nutrient inputs in Avoca Lagoon, Australia [66]. Rainfall and nutrients run-off significantly increased phytoplankton biomass (up to 10 fold) and altered their composition, promoting herbivory and consequently increasing zooplankton abundance [67,68]. However, in the Jansen tropical lagoon (Brazil), the rainfall diluted nutrients and induced phosphorous limitation, changing the dominant primary producers from Cyclotella stelligera to Closterium setaceum [69], with impacts on trophic interactions due to the different forms (round vs. elongated, respectively) and sizes (<50 µm vs. 200–250 µm, respectively) of these diatoms. In addition, freshwater flow regulation appears capable, alone, of determining changes in trophic interactions within plankton, as shown in the course of a tentative plan to restore the degraded lagoons of Coorong (Australia): under a regime of high-flow from land—namely, under a ‘green’ trophic state—phytoplankton–zooplankton interactions where favoured; on the other hand, at low flows—under a ‘blue’ state—interactions between pico- and nano-plankton were promoted, the autotrophy/heterotrophy ratio decreased, and the decomposition of dead matter was enhanced [47] (Figure 3). On the one hand, increases in terrestrial inputs can influence the trophic state of transitional systems, e.g., by enhancing their productivity; on the other hand, the plasticity of response that is intrinsic to trophic interactions can help buffer external perturbations, thereby stabilising plankton networks.
The strong link between physical-chemical drivers and plankton trophic networks as shown in the previous paragraphs suggests that coastal lagoons often alternate eutrophic and oligotrophic states depending on the relative dominance of freshwater inputs and seawater in-flows. In this regard, coastal lagoons differ from other TWs, such as estuaries [70]: healthy lagoons more often support blooms of picoplankton (<2 μm) that may be eventually sustained for months to years, whereas riverine estuaries support massive, seasonal spring diatom blooms reaching higher biomass. Indeed, picoplankton blooms in coastal lagoons are often sustained by regenerated forms of nitrogen (ammonium, urea, or dissolved organic substrates), thanks to the higher surface:volume ratios and long residence times of these systems [70,71,72,73]. Long-term studies on the lagoon systems of the Florida Bay and the Maryland–Virginia Chincoteague Bay [70] indicate that the onset of ‘water greening’ in both systems depends on increased nutrient availability, and that nutrient enrichment in turn often depends on human factors; however, compared to river-dominated estuaries, lagoon productivity is more supported by regenerated nutrients that may be lower in concentration, but which are made available through higher rates of matter degradation and microbial processes.
In such a context, it is plausible that the transition from a healthy to a degraded state in lagoons co-occurs with the transition from a ‘blue-green’ state—i.e., increases in nutrient inputs, phytoplankton biomass, and phytoplankton average size (see previous paragraphs)—to a persistent brown state, characterised by high levels of freshwater and terrestrial organic carbon from land [74]. Similarly to estuaries, although a lesser increase in the inflow of terrestrial matter can drive a higher primary productivity sustained by micro-phytoplankton, above a certain threshold of discharge, water browning promotes pico-phytoplankton, which are more adapted to lower light regimes, and a shift from a mesozooplankton- to a protozooplankton-dominated community [75] (Figure 3). Detritus consumption by planktonic microbes has been shown to compensate for limited phytoplankton production in the highly turbid Guadalquivir estuary [76]. As for degraded bays, a nutrient increase can drive a shift from a classical copepods-dominated trophic web to a microbial-loop web, due to an increase in bacterial production stimulated by the higher availability of phytoplankton-derived dead matter; the higher activity of heterotrophic microbes, in turn, brings about a decreased concentration of oxygen and more intense and frequent episodes of hypoxia, further hampering metazoan plankton [77].
Freshwater flow not only raises the levels of nutrients (silicate, phosphates, nitrogen), but may also lower the salinity of lagoon waters [78], gravely affect water quality [79], and jeopardise their capability to recycle matter, a systemic property that makes aquatic trophic networks robust in lagoons [80]. In the latter respect, Meddeb et al. (2018) [80] employed a ‘recycling index’ to assess the degree of perturbation of a plankton trophic web in the degraded Bizerte coastal lagoon (Tunisia, Mediterranean Sea), that is, a site that underwent a strong increase in land-based anthropogenic pressures and a multiplication of aquaculture activities. By means of ecological network analysis fed by data from modelling approaches, these authors observed that the recycling capability of plankton halved in a decade, between 2000 and 2010.
Our synthesis suggests that anthropogenic pressures can impact not only single interactions but the whole trophic network architecture of the plankton community. All the changes mentioned above may project the effects of anthropogenic pressures from the species to the trophic web level, inducing a shift from a balanced intermittence between blue and green trophic webs (i.e., an herbivorous pathway sustained by a microbial loop, and higher transfer of organic matter towards higher trophic levels), to a brown trophic web (i.e., one dominated by the microbial loop and detritus links, and with a lower transfer of matter to higher trophic levels) (Figure 3). Such trophic dynamics should be investigated in future studies focusing on coastal lagoons undergoing brownification.
4. Research Perspectives
TWs are among the most studied ecosystems due to their socio-economic benefits, and indeed the study of these systems intersects many thematic areas pertaining to socio-ecology. Despite the body of scientific works on coastal lagoons, some aspects of their ecology and their potential responses to climate change and anthropogenic stress are still unclear. Indeed, research on TWs is particularly difficult for at least three reasons. First, in these coastal marine environments, the land influence is of primary importance. Land influence is not only restricted to the freshwater input but also includes atmospheric inputs and salt and heat balances that are different from the open sea. Second, it is difficult to make a clear distinction between pristine conditions and conditions influenced by human activities in TWs, given that the first are very rare. Third, TWs, in particular coastal lagoons, may be considered eco-complexes containing more than one ecosystem (wetlands, marshes, sea grass fields, intertidal flats, and pelagic systems).
It is therefore not easy to outline the future directions of research on TWs; herein, we will discuss only a few issues that may receive intensified attention in the future, specifically those concerning coastal lagoons. One of these issues concerns our understanding of plankton dynamics in impacted lagoons, because these organisms respond quickly to chemical perturbations and undergo strong changes in consequence of hydrodynamic modifications—e.g., they live in the water column, which is primarily expanded or contracted by sea level rise.
Considering the huge diversity of plankton communities, plankton dynamics during perturbation may be effectively assessed by employing meta-omics technologies (meta-barcoding, -genomics, -transcriptomics, -proteomics, and/or -metabolomics). These techniques represent exceptional tools to aid and improve our knowledge of changes in biological and functional diversity, and this information can be translated into knowledge of the state of the plankton trophic network [81,82]. To date, -omics have been used only rarely in the study of coastal lagoons, at least in a holistic approach—the same approach used, e.g., in oceanic waters [83]. While a comparative metagenomic study of two Spanish lagoons (Mar Menor and Albufera de Valencia) was carried out in 2012 [84], most of the research efforts based on -omics tools are even more restricted, focusing on single sites, species, or processes [85,86,87]. Therefore, it is possible to affirm that the power of omics tools in the study of TWs is still a long way from reaching its enormous potential.
In addition, if on the one hand we have a starting idea of the effect of wastewater discharge on lagoon ecosystems, we are still lacking clear models of how major threats such as SLR will impact them. This gap in knowledge cannot be entirely solved by TW studies alone, since it involves a much larger body of research. First, we need conceptual and statistical models of future (and past) coastal changes that are able to incorporate the role of human activity on coasts. This is of primary importance in the study of TWs, which are not only among the most anthropogenically impacted ecosystems on earth, but are also among the systems with the longest histories of human influence (see, e.g., the Venice Lagoon). Second, most of the studies so far have targeted linear, long shorelines. Future efforts should theoretically incorporate a wider range of spatial and morphological lagoon types, possibly within a single framework.
Furthermore, given the relative sensitivity of coastal lagoon ecosystems and their planktonic communities to human and natural stressors, it is of pivotal importance to perform long-term research studies on a high number of sites globally, or, at least, to compare data taken distantly in time in the best-studied sites. For example, a comparison of data collected from the Venice Lagoon during two time periods (1975–1980 and 1997–2002) revealed higher phytoplankton abundance in the most recent survey, likely linked to the higher concentration of orthosilicates and dissolved inorganic nitrogen in the sea water, and to increased resuspension processes induced by dredging for molluscs, which favoured the mobilisation of benthic diatoms from sediments, resulting in more pelagic conditions [88]. On the other hand, the standing stock of zooplankton, for reasons that remain unclear, diminished during the same time periods.
In addition to hydrodynamic changes, TW ecosystems can also be impacted by warming, which, acting directly on species metabolism, drives changes in trophic interaction networks. However, the effect of warming on coastal lagoon plankton has been rarely studied and results are divisive. Warming experiments on Mediterranean lagoon plankton showed that temperature rising can increase ciliate and phytoplankton abundance, decrease heterotrophic nanoflagellates, and induce changes in trophic interactions [89,90]. Thermal stress treatments on sediments from a coral reef lagoon area at Sesoko reef in Sesoko Island, Okinawa, Japan induced a shift from heterotrophy to autotrophy [91]. However, experiments conducted on sediments from the Munkeby Bay (Sweden), indicated that an initial, relatively brief shift to net heterotrophy occurring during warming was followed by a more prolonged autotrophic period [92].
The high human pressure on TWs is a threat that has become increasingly recognised, both within public opinion and among policy makers (see, for example, the US Clean Water Act, the European Water Framework Directive, and the National Water Act in South Africa), and positive actions to restore TW ecosystems have been initiated. Nonetheless, there remains a need for improved predictive approaches to evaluate the putative impact of human actions, from new infrastructures to restoration plans for TW systems.
Author Contributions
Conceptualisation, D.D. and L.C.; resources, D.D., L.C., L.R. and G.D.G.; writing—original draft preparation, D.D., L.C., L.R. and G.D.G.; writing—review and editing, D.D. and L.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the project PO FEAMP 2014/2020 (Misura 2.51) funded by Regione Campania, Italy.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Voulvoulis, N.; Arpon, K.D.; Giakoumis, T. The EU Water Framework Directive: From great expectations to problems with implementation. Sci. Total Environ. 2017, 575, 358–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagliapietra, D.; Povilanskas, R.; Razinkovas-Baziukas, A.; Tamin, J. Emerald growth: A new framework concept for managing ecological quality and ecosystem services of transitional waters. Water 2020, 12, 894. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, H.; Lillebø, A.I.; Culhane, F.; Robinson, L.; Trauner, D.; Borgwardt, F.; Kummerlen, M.; Barbosa, A.; McDonald, H.; Funk, A.; et al. Linking biodiversity to ecosystem services supply: Patterns across aquatic ecosystems. Sci. Total Environ. 2019, 657, 517–534. [Google Scholar] [CrossRef] [PubMed]
- D’Alelio, D.; Russo, L.; Hay Mele, B.; Pomati, F. Intersecting Ecosystem Services Across the Aquatic Continuum: From Global Change Impacts to Local, and Biologically Driven, Synergies and Trade-Offs. Front. Ecol. Evol. 2021, 9, 1–9. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Pérez-Ruzafa, I.M.; Newton, A.; Marcos, C. Coastal Lagoons: Environmental Variability, Ecosystem Complexity, and Goods and Services Uniformity. In Coasts and Estuaries; Elsevier: Amsterdam, The Netherlands, 2019; pp. 253–276. ISBN 9780128140048. [Google Scholar]
- Ibánhez, J.S.P.; Arévalo, E.; Kelly, T.; Papaspyrou, S.; Rocha, C.; Nicolaidou, A. Unraveling the dispersion and environmental impact of anthropogenic discharges in transitional water ecosystems. Estuar. Coast. Shelf Sci. 2019, 216, 204–217. [Google Scholar] [CrossRef]
- Fry, B.; Sherr, E.B. δ 13 C measurements as indicators of carbon flow in marine and freshwater ecosystems. In Stable Isotopes in Ecological Research; Springer: New York, NY, USA, 1989; Volume 47, pp. 196–229. [Google Scholar]
- Winder, M.; Sommer, U. Phytoplankton response to a changing climate. Hydrobiologia 2012, 698, 5–16. [Google Scholar] [CrossRef]
- Cloern, J.E.; Jassby, A.D. Complex seasonal patterns of primary producers at the land-sea interface. Ecol. Lett. 2008, 11, 1294–1303. [Google Scholar] [CrossRef]
- Rothenberger, M.B.; Swaffield, T.; Calomeni, A.J.; Cabrey, C.D. Multivariate analysis of water quality and plankton assemblages in an urban estuary. Estuaries Coasts 2014, 37, 695–711. [Google Scholar] [CrossRef]
- Araujo, A.V.; Dias, C.O.; Bonecker, S.L.C. Differences in the structure of copepod assemblages in four tropical estuaries: Importance of pollution and the estuary hydrodynamics. Mar. Pollut. Bull. 2017, 115, 412–420. [Google Scholar] [CrossRef]
- Hallett, C.S.; Hobday, A.J.; Tweedley, J.R.; Thompson, P.A.; McMahon, K.; Valesini, F.J. Observed and predicted impacts of climate change on the estuaries of south-western Australia, a Mediterranean climate region. Reg. Environ. Chang. 2018, 18, 1357–1373. [Google Scholar] [CrossRef]
- Warwick, R.M.; Tweedley, J.R.; Potter, I.C. Microtidal estuaries warrant special management measures that recognise their critical vulnerability to pollution and climate change. Mar. Pollut. Bull. 2018, 135, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Pesce, M.; Critto, A.; Torresan, S.; Giubilato, E.; Santini, M.; Zirino, A.; Ouyang, W.; Marcomini, A. Modelling climate change impacts on nutrients and primary production in coastal waters. Sci. Total Environ. 2018, 628, 919–937. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here? Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Ducrotoy, J.P.; Michael, E.; Cutts, N.D.; Franco, A.; Little, S.; Mazik, K.; Wilkinson, M. Temperate Estuaries: Their Ecology Under Future Environmental Changes. In Coasts and Estuaries; Elsevier: Amsterdam, The Netherlands, 2019; pp. 253–276. [Google Scholar] [CrossRef]
- Tarafdar, L.; Kim, J.Y.; Srichandan, S.; Mohapatra, M.; Muduli, P.R.; Kumar, A.; Mishra, D.R.; Rastogi, G. Responses of phytoplankton community structure and association to variability in environmental drivers in a tropical coastal lagoon. Sci. Total Environ. 2021, 783, 146873. [Google Scholar] [CrossRef]
- Haraguchi, L.; Carstensen, J.; Abreu, P.C.; Odebrecht, C. Long-term changes of the phytoplankton community and biomass in the subtropical shallow Patos Lagoon Estuary, Brazil. Estuar. Coast. Shelf Sci. 2015, 162, 76–87. [Google Scholar] [CrossRef]
- Jakimavičius, D.; Kriaučiūnienė, J.; Šarauskienė, D. Impact of climate change on the Curonian Lagoon water balance components, salinity and water temperature in the 21st century. Oceanologia 2018, 60, 378–389. [Google Scholar] [CrossRef]
- Morabito, G.; Mazzocchi, M.G.; Salmaso, N.; Zingone, A.; Bergami, C.; Flaim, G.; Accoroni, S.; Basset, A.; Bastianini, M.; Belmonte, G.; et al. Plankton dynamics across the freshwater, transitional and marine research sites of the LTER-Italy Network. Patterns, fluctuations, drivers. Sci. Total Environ. 2018, 627, 373–387. [Google Scholar] [CrossRef]
- Duck, R.W.; da Silva, J.F. Coastal lagoons and their evolution: A hydromorphological perspective. Estuar. Coast. Shelf Sci. 2012, 110, 2–14. [Google Scholar] [CrossRef]
- Martin, L.; Landim Dominguez, J.M. Geological History of Coastal Lagoons. In Elsevier Oceanography Series; Elsevier: Amsterdam, The Netherlands, 1994; Volume 60, pp. 41–68. [Google Scholar]
- Kennish, M.J.; Paerl, H.W.E. Coastal Lagoons: Critical Habitats of Environmental Change; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Barnes, R.S.K. Coastal Lagoons; Archive, C., Ed.; Cambridge Univ. Press: New York, NY, USA, 1980; Volume 1. [Google Scholar]
- Boyd, R.; Dalrymple, R.; Zaitlin, B.A. Classification of clastic coastal depositional environments. Sediment. Geol. 1992, 80, 139–150. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Sigovini, M.; Ghirardini, A.V. A review of terms and definitions to categorise estuaries, lagoons and associated environments. Mar. Freshw. Res. 2009, 60, 497–509. [Google Scholar] [CrossRef]
- Colombo, G. Lagoons; Barnes, R., Ed.; John Wiley and Sons: New York, NY, USA, 1977. [Google Scholar]
- McGlathery, K.J.; Sundbäck, K.; Anderson, I.C. Eutrophication in shallow coastal bays and lagoons: The role of plants in the coastal filter. Mar. Ecol. Prog. Ser. 2007, 348, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Newton, A.; Brito, A.C.; Icely, J.D.; Derolez, V.; Clara, I.; Angus, S.; Schernewski, G.; Inácio, M.; Lillebø, A.I.; Sousa, A.I.; et al. Assessing, quantifying and valuing the ecosystem services of coastal lagoons. J. Nat. Conserv. 2018, 44, 50–65. [Google Scholar] [CrossRef]
- Escudero, M.; Silva, R.; Mendoza, E. Beach erosion driven by natural and human activity at Isla del Carmen Barrier Island, Mexico. J. Coast. Res. 2014, 71, 62–74. [Google Scholar] [CrossRef]
- Madarasinghe, S.K.; Yapa, K.K.A.S.; Satyanarayana, B.; Udayakantha, P.M.P.; Kodikara, S.; Jayatissa, L.P. Inland Irrigation Project Causes Disappearance of Coastal Lagoon: The Trajectory of Kalametiya Lagoon, Sri Lanka from 1956 to 2016. Coast. Manag. 2020, 48, 188–209. [Google Scholar] [CrossRef]
- Carrasco, A.R.; Ferreira, O.; Roelvink, D. Coastal lagoons and rising sea level: A review. Earth-Sci. Rev. 2016, 154, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Troiani, B.T.; Simms, A.R.; Dellapenna, T.; Piper, E.; Yokoyama, Y. The importance of sea-level and climate change, including changing wind energy, on the evolution of a coastal estuary: Copano Bay, Texas. Mar. Geol. 2011, 280, 1–12. [Google Scholar] [CrossRef]
- Toldo, J.; Dillenburg, S.R.; Correa, I.C.S.; Almeida, L.E.S.B. Holocene sedimentation in Lagoa dos Pantos Lagoon, Rio Grande do Sul, Brazil. J. Coast. Res. 2000, 16, 816–822. [Google Scholar]
- Sloss, C.R.; Jones, B.G.; McClennen, C.E.; de Carli, J.; Price, D.M. The geomorphological evolution of a wave-dominated barrier estuary: Burrill Lake, New South Wales, Australia. Sediment. Geol. 2006, 187, 229–249. [Google Scholar] [CrossRef] [Green Version]
- Ferla, M.; Cordella, M.; Michielli, L.; Rusconi, A. Long-term variations on sea level and tidal regime in the lagoon of Venice. Estuar. Coast. Shelf Sci. 2007, 75, 214–222. [Google Scholar] [CrossRef]
- Navrotskaya, S.E.; Chubarenko, B.V. Trends in the variation of the sea level in the lagoons of the Southeastern Baltic. Oceanology 2013, 53, 13–23. [Google Scholar] [CrossRef]
- Frihy, O.E.; El-Sayed, M.K. Vulnerability risk assessment and adaptation to climate change induced sea level rise along the Mediterranean coast of Egypt. Mitig. Adapt. Strateg. Glob. Chang. 2013, 18, 1215–1237. [Google Scholar] [CrossRef]
- van de Plassche, O. Coastal Evolution: Late Quaternary Shoreline Morphodynamics; Cambridge University Press: Cambridge, England, 1994. [Google Scholar]
- Hibma, A.; Stive, M.J.F.; Wang, Z.B. Estuarine morphodynamics. Coast. Eng. 2004, 51, 765–778. [Google Scholar] [CrossRef]
- Madricardo, F.; Foglini, F.; Campiani, E.; Grande, V.; Catenacci, E.; Petrizzo, A.; Kruss, A.; Toso, C.; Trincardi, F. Assessing the human footprint on the sea-floor of coastal systems: The case of the Venice Lagoon, Italy. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ghezzo, M.; Guerzoni, S.; Cucco, A.; Umgiesser, G. Changes in Venice Lagoon dynamics due to construction of mobile barriers. Coast. Eng. 2010, 57, 694–708. [Google Scholar] [CrossRef] [Green Version]
- Basset, A.; Barbone, E.; Rosati, I.; Vignes, F.; Breber, P.; Specchiulli, A.; D′Adamo, R.; Renzi, M.; Focardi, S.; Ungaro, N.; et al. Resistance and resilience of ecosystem descriptors and properties to dystrophic events: A study case in a Mediterranean lagoon. Transit. Waters Bull. 2013, 7, 1–22. [Google Scholar] [CrossRef]
- Pitacco, V.; Reizopoulou, S.; Sfriso, A.; Sfriso, A.; Mistri, M.; Munari, C. The difficulty of disentangling natural from anthropogenic forcing factors makes the evaluation of ecological quality problematic: A case study from Adriatic lagoons. Mar. Environ. Res. 2019, 150, 104756. [Google Scholar] [CrossRef]
- Ramdani, M.; Elkhiati, N.; Flower, R.J.; Thompson, J.R.; Chouba, L.; Kraiem, M.M.; Ayache, F.; Ahmed, M.H. Environmental influences on the qualitative and quantitative composition of phytoplankton and zooplankton in North African coastal lagoons. Hydrobiologia 2009, 622, 113–131. [Google Scholar] [CrossRef]
- Silva, A.M.A.; Barbosa, J.E.; Medeiros, P.R.; Rocha, R.M.; Lucena-Filho, M.A.; Silva, D.F. Zooplankton (Cladocera and Rotifera) variations along a horizontal salinity gradient and during two seasons (dry and rainy) in a tropical inverse estuary (Northeast Brazil). Panam. J. Aquat. Sci. 2009, 4, 226–238. [Google Scholar]
- Hemraj, D.A.; Hossain, A.; Ye, Q.; Qin, J.G.; Leterme, S.C. Anthropogenic shift of planktonic food web structure in a coastal lagoon by freshwater flow regulation. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Paturej, E.; Gutkowska, A.; Koszałka, J.; Bowszys, M. Effect of physicochemical parameters on zooplankton in the brackish, coastal Vistula Lagoon. Oceanologia 2017, 59, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Gilabert, J. Seasonal plankton dynamics in a Mediterranean hypersaline coastal lagoon: The Mar Menor. J. Plankton Res. 2001, 23, 207–217. [Google Scholar] [CrossRef]
- Pérez Ruzafa, Á.; Marcos Diego, C.; Gilabert Cervera, F.J. The ecology of the Mar Menor coastal lagoon: A fast changing ecosystem under human pressure. In Coastal Lagoons. Ecosystem Processes and Modeling for Sustainable Use and Development; CRC Press: Boca Raton, FL, USA, 2005; pp. 392–422. [Google Scholar]
- Alongi, D.M. Coastal Ecosystem Processes; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Vizzini, S.; Mazzola, A. The fate of organic matter sources in coastal environments: A comparison of three Mediterranean lagoons. Hydrobiologia 2008, 611, 67–79. [Google Scholar] [CrossRef]
- Nowicki, B.; Nixon, S. Benthic community metabolism in a coastal lagoon ecosystem. Mar. Ecol. Prog. Ser. 1985, 22, 21–30. [Google Scholar] [CrossRef]
- Knowlton, N. Multiple “stable” states and the conservation of marine ecosystems. Prog. Oceanogr. 2004, 60, 387–396. [Google Scholar] [CrossRef]
- Orfanidis, S.; Panayotidis, P.; Stamatis, N. Ecological evaluation of transitional and coastal waters: A marine benthic macrophytes-based model. Mediterr. Mar. Sci. 2001, 2, 45–66. [Google Scholar] [CrossRef]
- Howarth, R.W.; Marino, R. Nitrogen as the limiting nutrient for eutrophication in coastal marine ecosystems: Evolving views over three decades. Limnol. Oceanogr. 2006, 51, 364–376. [Google Scholar] [CrossRef] [Green Version]
- Viaroli, P.; Bartoli, M.; Giordani, G.; Naldi, M.; Orfanidis, S.Z.J. Community shifts, alternative stable states, biogeochemical controlsand feedbacks in eutrophic coastal lagoons: A brief overview. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, S105–S117. [Google Scholar] [CrossRef]
- Fong, P.; Kennison, R.L. Phase shifts, alternative stable states, and the status of Southern California lagoons. In Coastal Lagoons: Critical Habitats of Environmental Change; Michael, J., Kennish, H.W.P., Eds.; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Odame Appiah, D.; Yankson, D. Anthropogenic Drivers of the Pressures on the Ramsar Site of Sakumo Lagoon in Ghana. Int. J. Technol. Manag. Res. 2020, 1, 48–56. [Google Scholar] [CrossRef]
- Koutsodendris, A.; Brauer, A.; Zacharias, I.; Putyrskaya, V.; Klemt, E.; Sangiorgi, F.; Pross, J. Ecosystem response to human- and climate-induced environmental stress on an anoxic coastal lagoon (Etoliko, Greece) since 1930 AD. J. Paleolimnol. 2015, 53, 255–270. [Google Scholar] [CrossRef]
- Pérez Ruzafa, A.; Munilla León, T. Pycnogoid ecology in the Mar Menor (Murcia, SW Mediterranean). Sci. Mar. 1992, 56, 21–28. [Google Scholar]
- Álvarez-Rogel, J.; Barberá, G.G.; Maxwell, B.; Guerrero-Brotons, M.; Díaz-García, C.; Martínez-Sánchez, J.J.; Sallent, A.; Martínez-Ródenas, J.; González-Alcaraz, M.N.; Jiménez-Cárceles, F.J.; et al. The case of Mar Menor eutrophication: State of the art and description of tested Nature-Based Solutions. Ecol. Eng. 2020, 158, 106086. [Google Scholar] [CrossRef]
- Lloret, J.; Marín, A.; Marín-Guirao, L. Is coastal lagoon eutrophication likely to be aggravated by global climate change? Estuar. Coast. Shelf Sci. 2008, 78, 403–412. [Google Scholar] [CrossRef]
- D’Alelio, D.; Libralato, S.; Wyatt, T.; Ribera D′Alcalà, M. Ecological-network models link diversity, structure and function in the plankton food-web. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Alelio, D.; Hay Mele, B.; Libralato, S.; Ribera d′Alcalà, M.; Jordán, F. Rewiring and indirect effects underpin modularity reshuffling in a marine food web under environmental shifts. Ecol. Evol. 2019, 9, 11631–11646. [Google Scholar] [CrossRef]
- Maher, D.T.; Call, M.; Macklin, P.; Webb, J.R.; Santos, I.R. Hydrological Versus Biological Drivers of Nutrient and Carbon Dioxide Dynamics in a Coastal Lagoon. Estuaries Coasts 2019, 42, 1015–1031. [Google Scholar] [CrossRef]
- Rissik, D.; Shon, E.H.; Newell, B.; Baird, M.E.; Suthers, I.M. Plankton dynamics due to rainfall, eutrophication, dilution, grazing and assimilation in an urbanized coastal lagoon. Estuar. Coast. Shelf Sci. 2009, 84, 99–107. [Google Scholar] [CrossRef]
- Shangguan, Y. Response of plankton communities in coastal laggons to changes in nutrient quality and quantity: Case study of Florida Bay. Ph.D. Thesis, University of Maryland, College Park, MD, USA, 2016. [Google Scholar]
- Cutrim, M.V.J.; Ferreira, F.S.; Duarte dos Santos, A.K.; Cavalcanti, L.F.; de Oliveira Araújo, B.; de Azevedo-Cutrim, A.C.G.; Furtado, J.A.; Oliveira, A.L.L. Trophic state of an urban coastal lagoon (northern Brazil), seasonal variation of the phytoplankton community and environmental variables. Estuar. Coast. Shelf Sci. 2019, 216, 98–109. [Google Scholar] [CrossRef]
- Glibert, P.M.; Boyer, J.; Heil, C.; Madden, C.; Sturgis, B.W.C. Blooms in lagoons: Different from those of river-dominated estuaries. In Coastal Lagoons: Critical Habitats of Environmental Change; Kennish, M.J., Paerl, H., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 91–114. [Google Scholar]
- Glibert, P.M.; Hinkle, D.C.; Sturgis, B.; Jesien, R.V. Eutrophication of a Maryland/Virginia Coastal Lagoon: A Tipping Point, Ecosystem Changes, and Potential Causes. Estuaries Coasts 2014, 37, 128–146. [Google Scholar] [CrossRef] [Green Version]
- Malone, T.C.; Conley, D.J.; Fisher, T.R.; Glibert, P.M.; Harding, L.W.; Sellner, K.G. Scales of Nutrient-Limited Phytoplankton Productivity in Chesapeake Bay. Estuaries 1996, 19, 371–385. [Google Scholar] [CrossRef]
- Legendre, L.; Rassoulzadegan, F. Plankton and nutrient dynamics in marine waters. Ophelia 1995, 41, 153–172. [Google Scholar] [CrossRef]
- Deininger, A.; Faithfull, C.L.; Lange, K.; Bayer, T.; Vidussi, F.; Liess, A. Simulated terrestrial runoff triggered a phytoplankton succession and changed seston stoichiometry in coastal lagoon mesocosms. Mar. Environ. Res. 2016, 119, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Paczkowska, J.; Brugel, S.; Rowe, O.; Lefébure, R.; Brutemark, A.; Andersson, A. Response of Coastal Phytoplankton to High Inflows of Terrestrial Matter. Front. Mar. Sci. 2020, 7, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Donázar-Aramendía, I.; Sánchez-Moyano, J.E.; García-Asencio, I.; Miró, J.M.; Megina, C.; García-Gómez, J.C. Human pressures on two estuaries of the Iberian Peninsula are reflected in food web structure. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Najjar, R.G.; Pyke, C.R.; Adams, M.B.; Breitburg, D.; Hershner, C.; Kemp, M.; Howarth, R.; Mulholland, M.R.; Paolisso, M.; Secor, D.; et al. Potential climate-change impacts on the Chesapeake Bay. Estuar. Coast. Shelf Sci. 2010, 86, 1–20. [Google Scholar] [CrossRef]
- Johannes, R.E.; Hearn, C.J. The effect of submarine groundwater discharge on nutrient and salinity regimes in a coastal lagoon off Perth, Western Australia. Estuar. Coast. Shelf Sci. 1985, 21, 789–800. [Google Scholar] [CrossRef]
- Cravo, A.; Barbosa, A.B.; Correia, C.; Matos, A.; Caetano, S.; Lima, M.J.; Jacob, J. Unravelling the effects of treated wastewater discharges on the water quality in a coastal lagoon system (Ria Formosa, South Portugal): Relevance of hydrodynamic conditions. Mar. Pollut. Bull. 2022, 174, 113296. [Google Scholar] [CrossRef]
- Meddeb, M.; Grami, B.; Chaalali, A.; Haraldsson, M.; Niquil, N.; Pringault, O.; Sakka Hlaili, A. Plankton food-web functioning in anthropogenically impacted coastal waters (SW Mediterranean Sea): An ecological network analysis. Prog. Oceanogr. 2018, 162, 66–82. [Google Scholar] [CrossRef] [Green Version]
- Stec, K.F.; Caputi, L.; Buttigieg, P.L.; D’Alelio, D.; Ibarbalz, F.M.; Sullivan, M.B.; Chaffron, S.; Bowler, C.; Ribera d′Alcalà, M.; Iudicone, D. Modelling plankton ecosystems in the meta-omics era. Are we ready? Mar. Genom. 2017, 32, 1–17. [Google Scholar] [CrossRef]
- D’Alelio, D.; Eveillard, D.; Coles, V.J.; Caputi, L.; Ribera d′Alcalà, M.; Iudicone, D. Modelling the complexity of plankton communities exploiting omics potential: From present challenges to an integrative pipeline. Curr. Opin. Syst. Biol. 2019, 13, 68–74. [Google Scholar] [CrossRef]
- Karsenti, E.; Acinas, S.G.; Bork, P.; Bowler, C.; De Vargas, C.; Raes, J. Tara Oceans Consortium A Holistic Approach to Marine Eco-Systems Biology. PLoS Biol. 2011, 9, e1001177. [Google Scholar] [CrossRef]
- Ghai, R.; Hernandez, C.M.; Picazo, A.; Mizuno, C.M.; Ininbergs, K.; Díez, B.; Valas, R.; Dupont, C.L.; McMahon, K.D.; Camacho, A.; et al. Metagenomes of mediterranean coastal lagoons. Sci. Rep. 2012, 2, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pinnell, L.J.; Turner, J.W. Shotgun metagenomics reveals the benthic microbial community response to plastic and bioplastic in a coastal marine environment. Front. Microbiol. 2019, 10, 1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, A.B.; Ghai, R.; Martin-Cuadrado, A.B.; Sánchez-Porro, C.; Rodriguez-Valera, F.; Ventosa, A. Prokaryotic taxonomic and metabolic diversity of an intermediate salinity hypersaline habitat assessed by metagenomics. FEMS Microbiol. Ecol. 2014, 88, 623–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milan, M.; Pauletto, M.; Boffo, L.; Carrer, C.; Sorrentino, F.; Ferrari, G.; Pavan, L.; Patarnello, T.; Bargelloni, L. Transcriptomic resources for environmental risk assessment: A case study in the Venice lagoon. Environ. Pollut. 2015, 197, 90–98. [Google Scholar] [CrossRef]
- Acri, F.; Aubry, F.B.; Berton, A.; Bianchi, F.; Boldrin, A.; Camatti, E.; Comaschi, A.; Rabitti, S.; Socal, G. Plankton communities and nutrients in the Venice Lagoon. Comparison between current and old data. J. Mar. Syst. 2004, 51, 321–329. [Google Scholar] [CrossRef]
- Pulina, S.; Suikkanen, S.; Padedda, B.M.; Brutemark, A.; Grubisic, L.M.; Satta, C.T.; Caddeo, T.; Farina, P.; Lugliè, A. Responses of a Mediterranean coastal lagoon plankton community to experimental warming. Mar. Biol. 2020, 167, 1–14. [Google Scholar] [CrossRef]
- Courboulès, J.; Vidussi, F.; Soulié, T.; Mas, S.; Pecqueur, D.; Mostajir, B. Effects of experimental warming on small phytoplankton, bacteria and viruses in autumn in the Mediterranean coastal Thau Lagoon. Aquat. Ecol. 2021, 55, 647–666. [Google Scholar] [CrossRef]
- Sultana, R.; Casareto, B.E.; Sohrin, R.; Suzuki, T.; Alam, M.S.; Fujimura, H.; Suzuki, Y. Response of subtropical coastal sediment systems of Okinawa, Japan, to experimental warming and high pCO2. Front. Mar. Sci. 2016, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Alsterberg, C.; Sundbäck, K.; Hulth, S. Functioning of a Shallow-Water Sediment System during Experimental Warming and Nutrient Enrichment. PLoS ONE 2012, 7, e51503. [Google Scholar] [CrossRef]
Figure 1.
Scheme of planktonic organisms. Drawings based on the open-source IAN symbol library (https://ian.umces.edu/media-library/symbols/, accessed on 1 November 2021).
Figure 1.
Scheme of planktonic organisms. Drawings based on the open-source IAN symbol library (https://ian.umces.edu/media-library/symbols/, accessed on 1 November 2021).

Figure 2.
Plankton trophic network. Based on D’Alelio et al. 2016 [64]. Arrows go from prey to predators. Plankton drawings based on IAN libraries (https://ian.umces.edu/media-library/symbols/, accessed on 1 November 2021).
Figure 2.
Plankton trophic network. Based on D’Alelio et al. 2016 [64]. Arrows go from prey to predators. Plankton drawings based on IAN libraries (https://ian.umces.edu/media-library/symbols/, accessed on 1 November 2021).

Figure 3.
Scheme of changes in matter fluxes within plankton as driven by changes in water quality (from blue to green to brown, i.e., at increasing levels of freshwater inputs). PP = picoplankton; MP = microplankton; PZ = protozooplankton; ZOO = mesozooplankton. Drawings based on the open-source IAN symbol library (https://ian.umces.edu/media-library/symbols/, accessed on 1 November 2021). Arrows indicate trophic links and go from prey to predators; arrow width is proportional to the amount of matter being transferred between prey and predator. Blue and green links are based on D’Alelio et al., 2016 [64]. The brown links are hypothetical.
Figure 3.
Scheme of changes in matter fluxes within plankton as driven by changes in water quality (from blue to green to brown, i.e., at increasing levels of freshwater inputs). PP = picoplankton; MP = microplankton; PZ = protozooplankton; ZOO = mesozooplankton. Drawings based on the open-source IAN symbol library (https://ian.umces.edu/media-library/symbols/, accessed on 1 November 2021). Arrows indicate trophic links and go from prey to predators; arrow width is proportional to the amount of matter being transferred between prey and predator. Blue and green links are based on D’Alelio et al., 2016 [64]. The brown links are hypothetical.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
D’Alelio, D.; Russo, L.; Del Gaizo, G.; Caputi, L. Plankton under Pressure: How Water Conditions Alter the Phytoplankton–Zooplankton Link in Coastal Lagoons. Water 2022, 14, 974. https://doi.org/10.3390/w14060974
AMA Style
D’Alelio D, Russo L, Del Gaizo G, Caputi L. Plankton under Pressure: How Water Conditions Alter the Phytoplankton–Zooplankton Link in Coastal Lagoons. Water. 2022; 14(6):974. https://doi.org/10.3390/w14060974
Chicago/Turabian StyleD’Alelio, Domenico, Luca Russo, Gabriele Del Gaizo, and Luigi Caputi. 2022. "Plankton under Pressure: How Water Conditions Alter the Phytoplankton–Zooplankton Link in Coastal Lagoons" Water 14, no. 6: 974. https://doi.org/10.3390/w14060974
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.