
Antimicrobial Resistance of Heterotrophic Bacteria in Drinking Water-Associated Biofilms


1
Laboratory Ecology of Pathogenic Bacteria, Department Infectious Microbiology, The Stephan Angeloff Institute of Microbiology, Bulgarian Academy of Sciences, 78 Nikola Gabrovski Str., 5002 Veliko Tarnovo, Bulgaria
2
Laboratory Bacterial Virulence, Resistance and New Antimicrobial Agents, Department Infectious Microbiology, The Stephan Angeloff Institute of Microbiology, Bulgarian Academy of Sciences, 26 Acad. G. Bonchev Str., 1113 Sofia, Bulgaria
*
Author to whom correspondence should be addressed.
Water 2022, 14(6), 944; https://doi.org/10.3390/w14060944
Submission received: 19 February 2022
/
Revised: 10 March 2022
/
Accepted: 14 March 2022
/
Published: 17 March 2022
(This article belongs to the Special Issue Antimicrobial Resistance in the Urban Water Cycle and Natural Aquatic Environments)
Abstract
:Antimicrobial resistance (AMR) is one of the major threats to human health and is becoming an environmental challenge for water resources too. Our study’s aim was: to assess the AMR of heterotrophic bacteria in drinking water-associated biofilms against six clinically important antibiotics; to compare the prevalence of antibiotic resistant bacteria (ARB) in drinking water and in the associated biofilms; to estimate biofilm formation ability of selected isolates. Culture-dependent methods were used in the population-based study of the biofilms and in assessment of the single-species biofilm formation ability and the AMR phenotype of the isolated strains. The population proportion of the bacteria resistant to each tested antibiotic significantly differed in the biofilms formed in drinking water from different sampling points. In all biofilms, the abundance of tetracycline- and ampicillin-resistant bacteria was low, and of streptomycin-resistant bacteria was high. An increased proportion of the bacteria resistant to ciprofloxacin, chloramphenicol and streptomycin was detected in the biofilms compared to those found in the drinking water. The prevalence of ARB in the biofilms implies an impact on the drinking water quality and an assessment of the attached and the planktonic bacteria is needed to clarify the prevalence of AMR in the drinking water distribution system.
1. Introduction
Antimicrobials set the beginning of a new era in healthcare, but nowadays, their effective action span has decreased dramatically, and pathogenic bacteria develop resistance to new antibiotics (ABs) quickly, sometimes for less than a year. The increasing AMR among clinically important pathogens is a serious health risk that, in recent years, has grown into an ecological challenge. There is increasing evidence that small amount of ABs (remaining undigested during treatment and excreted from the body) become emerging pollutants of wastewater and water resources, and that the aquatic microbiota represents an important reservoir of AMR against one or more classes of ABs [1,2,3]. Sharing different aquatic habitats and moving between different types of water—wastewater, natural and drinking water—determine the role of antibiotic resistant bacteria (ARB) as carriers or potential vectors of ABR to human microbiota [4].
In an aquatic environment, microorganisms are prone to attaching to the surfaces in contact with water, forming multispecies communities embedded in a self-produced matrix of extracellular polymeric substances, called biofilm [5]. Biofilms are generated in all types of water, being a common phenomenon in drinking water distribution systems (DWDS), where they comprise about 95% of overall active microbial biomass [6]. Biofilms provide more favorable growth conditions for the attached microorganisms in an oligotrophic environment, but also a shelter against disinfectants, protozoa predators, etc. [5]. Detachment of bacteria or sloughing of biofilm pieces into drinking water may increase the bacterial number and deteriorate bacteriological water quality [7].
Drinking water disinfection is the main strategy to ensure its microbiological quality and safety, but has insufficient effectiveness on biofilm biomass reduction [8,9,10]. Drinking water-associated biofilms manifest greater antimicrobial tolerance than free-living (planktonic) bacteria and require higher concentration of disinfectant and longer contact time to achieve the same degree of bacterial inactivation [8,11,12].
In fact, the biofilm formation is a preferred mode of life in different environments (water, soil, dental plaques, medical devices, implants, etc.) and an effective way for bacteria to survive in the presence of antimicrobials as disinfectants and ABs [5]. However, the effect of ABs mainly was studied on the biofilms of clinically important pathogenic bacteria, such as Pseudomonas aeruginosa, Staphylococcus aureus, Acinetobacter baumannii, etc. [13,14,15,16]. Some studies have reported for biofilm-specific resistance significantly higher than antibiotic resistance of suspended bacteria [15,16]. Contradicting results for quantitative correlations between biofilm formation capacity/antibiotic resistance and biofilm-specific resistance of some pathogenic bacteria have been found, too.
The three-dimensional structure of biofilms and their heterogeneity underlie the antimicrobial resistance [5,17]. It is considered that the increased AMR of biofilms is based on their specific spatial architecture, providing difficult penetration into the biofilm matrix, the higher bacterial density, the lower bacterial growth rate, conditions for acquiring resistant phenotypes and the presence of “persisters” [5,13,17,18,19]. For example, the penetration of positively charged aminoglycosides may be hampered by negatively charged exopolymers of the biofilm matrix [18,19]. The local pH and oxygen gradients created by the biofilm structure can affect the action of aminoglycosides, macrolides and beta-lactams, due to their reduced efficiency in anaerobic conditions [19]. The survival or growth of pathogenic bacteria in biofilms and the enhanced gene exchange, including the antibiotic resistance genes (ARGs), favored by high cell density and species diversity, may contribute to dissemination of AMR in potentially dangerous microorganisms. Similarly, there is evidence that ABs are involved as signaling molecules in the regulation of homeostasis of microbial communities [20] and in sub-inhibitory concentrations they can induce biofilm formation [21].
Despite the extensive research on biofilms in DWDS, few studies have been addressed to AMR of the attached microbial communities [22,23], while most of them have been focused on quantification of ARB and ARGs in water sources and purified drinking water [24,25,26,27]. An increased number of ARB and ARGs have been found in treated drinking water compared to the water source [24,26]. Ubiquitous bacteria from the genera Sphingomonas, Pseudomonas and Acinetobacter, capable of withstanding the action of various ABs have been also detected [25,28,29]. The ARB in drinking water, even when harmless, can increase the spread of AMR and be carriers or potential vectors of AMR to human-associated microbiota. Because of that, dissemination of ARB and ARGs in drinking water is considered an emerging health risk, in spite of the fact that some of the resistance phenotypes are intrinsic [4].
Our previous study on the prevalence of different types of ARB in drinking water of a municipal DWDS found significant population changes in the drinking water supply network. The populations’ density and the predominance of particular types of ARB varied depending on the drinking water sampling point [30]. Whether the prevalence of different ARB in water and biofilm follow the same levels and undergoes similar population changes in the pipeline network has not been sufficiently clarified. Because of that, the present study was addressed to conduct the following: (a) a population-based assessment of the AMR in biofilms formed in drinking water from a selected water supply system; (b) a comparative assessment of the prevalence of ARB in drinking water and water-associated biofilms, including an evaluation of the antibiotic resistance phenotype of bacterial isolates; (c) an evaluation of the ability of bacterial isolates with various level of AMR to form single-species biofilms.
The present study will be a basis for the application of a complex approach for AMR investigations of drinking water and drinking water-associated biofilms in other DWDSs in Bulgaria.
2. Materials and Methods
The object of the population-based study were the biofilms formed in drinking water samples for a certain period of time under defined conditions. The experimental procedure includes bacteriological analysis of the drinking water samples before starting the incubation period for biofilm production, biofilm formation at static conditions, and regular analyses of the generated biofilms for quantification of culturable heterotrophic bacteria (HPC) as total number and number of bacteria resistant to distinct antibiotics.
2.1. Enumeration of HPC Bacteria and ARB in Drinking Water and Drinking Water-Associated Biofilms
The total number of HPC bacteria in biofilm community or drinking water was determined by culturing on R2A agar (HiMedia, Mumbai, India) [31], while the number of bacteria resistant to each individual AB was assessed on R2A agar in which the tested AB substance was added at a minimum inhibitory concentration. Based on the HPC data pairs (with/without an AB), the relative proportion of the bacteria resistant to each of the tested ABs was calculated.
The population of heterotrophic bacteria resistant to six AB substances from different classes were quantified as follows: β-lactams (ampicillin sodium salt, AMP—32 mg/L); aminoglycosides (streptomycin sulphate, S—32 mg/L); tetracyclines (tetracycline hydrochloride, TE—8 mg/L); amphenicols (chloramphenicol, C—16 mg/L); fluoroquinolones (ciprofloxacin hydrochloride monohydrate, CIP—4 mg/L); antifolates (sulphamethoxazole, Sul—256 mg/L) (HiMedia, Mumbai, India).
2.2. Study Area and Drinking Water Sampling
Drinking water samples were collected from four sampling points of the municipal DWDS supplying drinking water to the inhabitants of the region of Veliko Tarnovo, Bulgaria (a scheme of the DWDS and the DW sampling points in Supplementary Materials). After the treatment and chlorination at the drinking water treatment plant, the drinking water is transported gravimetrically to many settlements in the surrounding area and to the water tanks in the town of Veliko Tarnovo, and then, by pumping, to the end consumers.
The tap water sampling sites—two residential properties (DW-1 and DW-4), one public building (DW-2) and one public fountain (DW-3)—were selected to cover sections of the distribution network in which drinking water differs in residual chlorine content. The sampling sites DW-1 and DW-4 were selected to collect low residual chlorine-containing tap water (0.32 ± 0.06 mg/L for DW-1 and 0.31 ± 0.03 mg/L for DW-4; n of 18), while the DW-2 and DW-3—for water samples with higher residual chlorine content—was as follows: 0.71 ± 0.13 mg/L for DW-2 and 0.74 ± 0.09 mg/L for DW-3; n of 22.
2.3. Biofilm Formation in Static Conditions
The biofilms were formed on the surface of sterile pipe test pieces made of polypropylene, material often used in plumbing. In brief, the procedure involved immersing six pipe test pieces (each with surface area of 50 cm2) in dechlorinated drinking water (900 mL) from each sampling location, and then culturing at 25 °C for 40 days. From each tested water sample, two test vessels were incubated. At regular intervals, one test piece was taken from each vessel and used for biofilm analysis. The biofilms formed on the test surfaces were analyzed in duplicates after 20, 30 and 40 days of incubation.
2.4. Biofilm Analysis
The biofilm formed on each test surface was washed with 20 mL of sterile saline. Next, the test surface was wiped with sterile cotton swab dipped in 50 mL physiological solution and treated for 5 min in ultrasonic bath (Apronecs; 40 kHz; Gabrovo, Bulgaria). The bacterial suspension of the removed biofilm biomass was serially diluted. Afterwards, the drinking water-associated biofilms were analyzed by plate surface spreading of aliquots (0.1 mL) from the last three decimal dilutions of the bacterial suspension. After 7 days incubation at a temperature of 25 °C, the colonies were counted, and the results were calculated as CFU/cm2.
2.5. Drinking Water Analysis
Each drinking water sample was analyzed in duplicates by membrane filtration method through 0.45 μm pore size, 47 mm diameter sterile membrane filters (Sartorious Stedim Biotech GmbH, Goettingen, Germany) and subsequent cultivation of the filters on R2A agar for 7 days at a temperature of 25 °C. By filtering certain volumes of drinking water (depending on the putative number of the analyzed type of ABR) and incubating the filters on standard R2A agar or on R2A agar with addition of AB, the total number of HPC bacteria and the number of bacteria resistant to each individual AB were determined in parallel.
2.6. Antibiotic Susceptibility Testing of Isolated Bacteria
Forty bacterial strains isolated from biofilm samples and 145 strains isolated from drinking water were analyzed for determining their antibiotic susceptibility by disc diffusion method. The susceptibility to the following ABs was tested: ampicillin, AMP 10 μg; cefotaxime, CTX 30 μg; streptomycin, S 10 μg; gentamycin, GEN 10 μg; ciprofloxacin, CIP 5 μg; tetracycline, TE 30 μg; chloramphenicol, C 30 μg; trimethoprim/sulfamethoxazole, SXT 1.25/23.75 μg.
In brief, the suspension of each tested strain (calibrated to 0.5 according MacFarland) was inoculated over the surface of Mueller–Hinton agar (HiMedia, Mumbai, India). Disks with the tested ABs (HiMedia, Mumbai, India) were placed on the inoculated agar surface. After 18 h of incubation at 35 °С, the inhibition zone diameter around each AB disk was measured (mm). The strains were classified as S (sensitive) or R (resistant). As there are no guidelines for breakpoints of environmental bacteria, the results for Gram-positive bacteria were interpreted according to CLSI breakpoints for Staphylococcus spp., and for Gram-negative bacteria according to breakpoints for family Enterobacteriaceae. The resistance towards at least one AB from at least three different classes was defined as multidrug resistance (MDR) [32,33].
2.7. Assessment of Biofilm Formation Ability of Bacteria Isolated from Drinking Water
Twenty bacterial strains with established MDR or ones representing the largest populations in drinking water were selected for assessment of their biofilm formation ability. The strains have been isolated and identified during our previous study on the prevalence of AMR among the aquatic microbiota of the same DWDS [30]. The isolates have been subjected to biochemical identification by MICROLATEST® tests (ERBA Lachema s.r.o., Brno, Czech Republic) or by BD PhoenixTM M50 Automated Microbiology System (Becton, Dickinson and Company, Franklin Lakes, NJ, USA). The instrument BD PhoenixTM M50 simultaneously identifies the isolates and assesses their antimicrobial susceptibility in combined panels. NMIC/ID-76 panels (Becton, Dickinson and Company, Franklin Lakes, NJ, USA) have been used for identification and antimicrobial susceptibility testing of Gram-negative bacteria, and PMIC/ID-88 have been used for Gram-positive bacteria [30]. Pure bacterial cultures were stored at −20 °C. The isolates were sub-cultured on Soyabean Casein Digest Agar (HiMedia, Mumbai, India).
The experimental procedure for assessing the ability of the isolated heterotrophic bacteria to form single-species biofilm is based on measurement of amount of the attached biomass in nutrient medium (R2A broth or minimal M63 medium with 0.1% glucose) by modified crystal violet assay. The experiments were performed in flat-bottom 24-well microtiter plates (Costar®, Corning Inc., Corning, NY, USA) with suspensions of one-day pure cultures of the tested bacterial strains in the nutrient medium, calibrated to optical density (OD620nm) of 0.1. One milliliter of each calibrated suspension was pipetted in four wells of two microtiter plates, and sterile medium was used as negative control. The inoculated microtiter plates were incubated for 2 days at 25 °C.
To assess the planktonic growth, the optical density (OD620nm) of the bacterial suspension taken from each well was measured. Then, the plate was washed three times with saline, dried, and the biofilm biomass in the individual wells was stained by 0.1% crystal violet (CV) solution as previously described [34,35,36]. The optical density (OD590nm) of the ethanol solution of CV dissolved from the dyed biofilm was measured. The biofilm-forming ability (BF) of the bacterial strains was classified as follow [34,36]:
(0) —non-biofilm forming bacteria, when BF ≤ ODc;
(+)—weak biofilm producer, when ODc < BF ≤ 2 ODc;
(++)—moderate biofilm producer, when 2 ODc < BF ≤ 4 ODc;
(+++)—strong biofilm producer, when BF ≥ 4 ODc.
This classification was based on the cutoff OD (ODc) value, defined as three standard deviation values above the mean OD590nm of the negative control.
For each bacterial strain, a ratio between the amount of the biofilm biomass and the plankton biomass, both measured as OD, was calculated [37]. The biofilm index (BI) presents the preferences of a particular bacterial strain for growth (sessile or free-living) in a particular nutrient medium. When BI is greater than 1, it is considered that the strain has a pronounced biofilm-forming potential.
2.8. Statistical Analysis of the Data
One-way analysis of variance (ANOVA) with the Tukey’s HSD procedure was performed to assess the significance of the differences between the sampling locations on the percentage of the bacteria resistant to the individual ABs within the biofilm samples as the dependent variable, and sampling location as the factor p < 0.05 was considered as statistically significant. The Tukey’s procedure facilitates pairwise comparisons within the ANOVA data detecting whether there is an overall difference between the sample means and establishing between which of the various pairs of means there is a significant difference.
A significance t-test for statistical comparison between the HPC values of two biofilms was used to check whether the difference between two values was significant. The truth of null hypothesis was tested and when the experimental value of |t| was greater than the critical value of t (at p = 0.05 and a degree of freedom n), the difference between the two compared results was considered significant.
3. Results
3.1. Total Number of HPC and ARB in Drinking Water and Water-Associated Biofilms
The data on the number of HPC bacteria in drinking water from all DWDS sampling points and the bacterial density of the associated biofilms are presented in Table 1.
The drinking water samples significantly differ in the number of HPC bacteria, which is not surprising, as it is known that bacteriological water quality may undergo changes in water supply network due to great number of factors. Some but not all are water chlorination, hydrodynamics and biological factors, as bacterial re-growth or biofilm impact in water supply network [5,38,39,40].
In the DWDSs, biofilms are dynamic structures in a pseudo-steady state [6,38]. The chosen 40-day period for biofilm formation presupposes reaching the plateau phase in the biofilm formation process [41]. The observed fluctuations in bacterial density are common in drinking water-associated biofilms and could be related to their inherent dynamic nature and characteristic spatial heterogeneity. No correlation was found between the HPCs values of the drinking water and the biofilm community depending on sampling point. Only the biofilms grown in DW-3 and DW-4 had similar HPC densities (|t| = 1.9 < t = 2.6 (p = 0.05; n = 10), but they were significantly different from the others.
Besides the shifts in the total number of HPC bacteria in the tap water, changes in populations of the bacteria resistant to various ABs have been reported for some studied DWDSs [24,27]. Our previous study on the prevalence of AMR among the drinking water microbiome of the same DWDS also found significant differences in the populations proportion of the bacteria resistant to nine ABs from seven classes. It was found that the populations of the different types of ARB varied in tap water depending on the sampling location and the type of AB to which they are resistant [30].
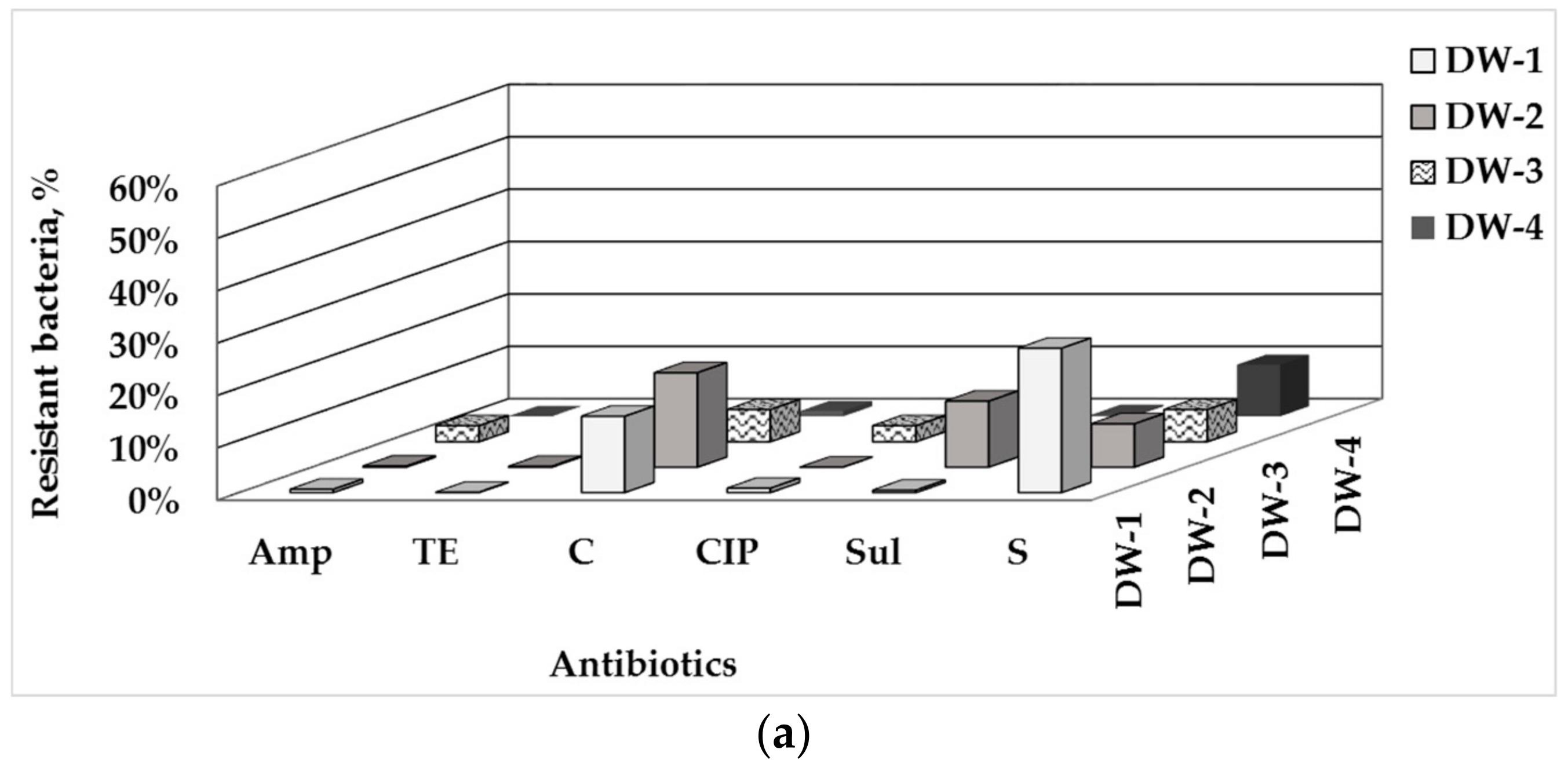
In all tap water samples used in this biofilm study, the population level of tetracycline-resistant bacteria was the lowest, followed by ampicillin-, ciprofloxacin- and sulfamethoxazole-resistant bacteria, then by the chloramphenicol-resistant bacteria with a moderate abundance and the highest level of streptomycin-resistant bacteria (Figure 1a).
The samples differed in the predominant type of ARB: in tap water DW-1, the chloramphenicol- and streptomycin-resistant bacteria were predominant, as well in DW-2, where a high level of sulfamethoxazole-resistant bacteria also was detected. In tap waters DW-3 and DW-4, the streptomycin-resistant bacteria prevailed, as well the chloramphenicol-resistant ones in TW-3 (detailed data in Supplementary Materials, Tables S1 and S2).
The data on the biofilms composition (Figure 1b) show similar trends as found for the drinking water: the lowest proportion of bacteria resistant to tetracycline (up to 0.06%) and ampicillin (up to 0.4%); predominantly low levels of AMR to ciprofloxacin (up to 1.8%, excepting DW-4), and the highest values of streptomycin-resistant bacteria. An apparent variation in the population levels of bacteria resistant to chloramphenicol (from 2.0% to 25.4%), sulfomethaxazole (from 0.2% to 8.6%) and streptomycin (from 23.8% to 54.9%) was found between the individual sampling points at the DWDS.
The populations proportion of the bacteria resistant to the individual ABs in the biofilm community can be ranged in the following ascending order: ТЕ < Amp < CIP < Sul < C < S. A few exceptions were found: in only the biofilms in DW-4, the population level of CIP-resistant bacteria was significantly higher and the proportion of Sul-resistant bacteria was many times lower than in the rest of the biofilms. Despite these general population trends, the biofilms formed in drinking water from all the DWDS-sampling points differed significantly (p < 0.05) in population proportion of bacteria resistant to the individual ABs. The data for the ciprofloxacin-, chloramphenicol- and streptomycin-resistant bacteria manifested in the best way the overall difference between the biofilms, depending on the sampling location.
The biofilm communities in the individual DW samples show some specific traits: similar abundance of the bacteria resistant to C and Sul in the biofilms in DW-1; a higher proportion of bacteria resistant to C in the biofilms in DW-2 and DW-3 than in the rest of the biofilms. The biofilms in DW-4 differed most significantly from the others with the highest population level of the bacteria resistant to CIP and S, and the lowest shares of the bacteria resistant to C and Sul.
3.2. Phenotype of AMR of Bacterial Isolates from Water and Biofilm
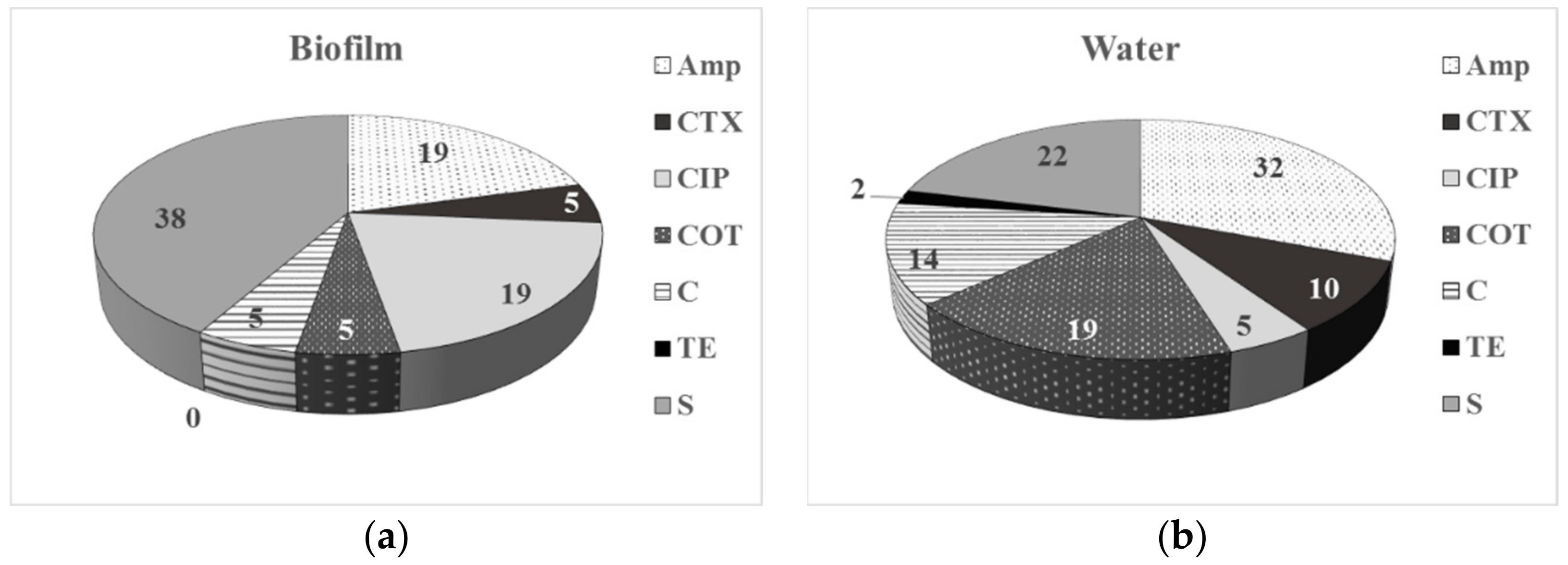
The comparative data on the AMR phenotype of the bacteria isolated from drinking water and associated biofilms (Figure 2) reveal a higher percentage of resistance to S and CIP among the biofilm bacteria comparing to the planktonic compartment. No tetracycline-resistant isolates were detected among the biofilm bacteria, compared to 2% of resistant planktonic bacteria.
Among the planktonic bacteria, a higher level of resistance to beta-lactams, C and COT was detected. Biofilm bacteria were resistant to one or two classes of ABs (52% and 24%, respectively), while the planktonic isolates also exhibited MDR to three classes of ABs (41%, 18%, and MDR-3 of 5%, respectively). These AMR phenotype data for a higher resistance to S (38%) and CIP (19%) among the biofilm bacterial isolates are in correspondence with the population-based biofilm data, especially for the biofilms formed in DW-4.
3.3. Single-Species Biofilm Formation of Bacterial Isolates
The differences in the populations of the studied types of ARB in drinking water and water-associated biofilms, and in the resistance phenotype of the planktonic and the attached bacteria, raise the question whether there is a relationship between AMR or MDR of the aquatic bacteria and their biofilm-forming ability. Therefore, the conducted study aimed to assess the biofilm-forming ability of bacterial strains with different levels of AMR, as well as representatives of the most common bacteria in drinking water.
Twenty bacterial strains of the dominant HPC bacteria in drinking water from the family Sphingomonadaceae and genera Methylobacterium, Pseudomonas, Staphylococcus and Bacillus were selected for assessment of their biofilm-forming ability (Table 2).
Several opportunistic pathogens as Sphingomonas paucimobilis, Embedobacter brevis and Stenotrophomonas maltophilia causing nosocomial infections or diseases in immuno-compromised patients were among the selected isolates. The AMR profiles and some parameters characterizing the biofilm-forming potential of the tested strains are presented in Table 2. Among the isolates, only Stenotrophomonas maltophilia was multidrug resistant to five classes of ABs (beta-lactams, aminoglycosides, FF, FM, TMP), and P. putida—to four classes (beta-lactams, C, CIP, FM). Twelve strains, including the opportunistic pathogens Sphingomonas paucimobilis and E. brevis were resistant to two antimicrobial classes (beta-lactams, FM). Pseudomonas strains exhibited diverse resistance toward two to four classes of ABs. Only Methylobacterium exorquens and Micrococcus luteus were fully sensitive to the tested ABs.
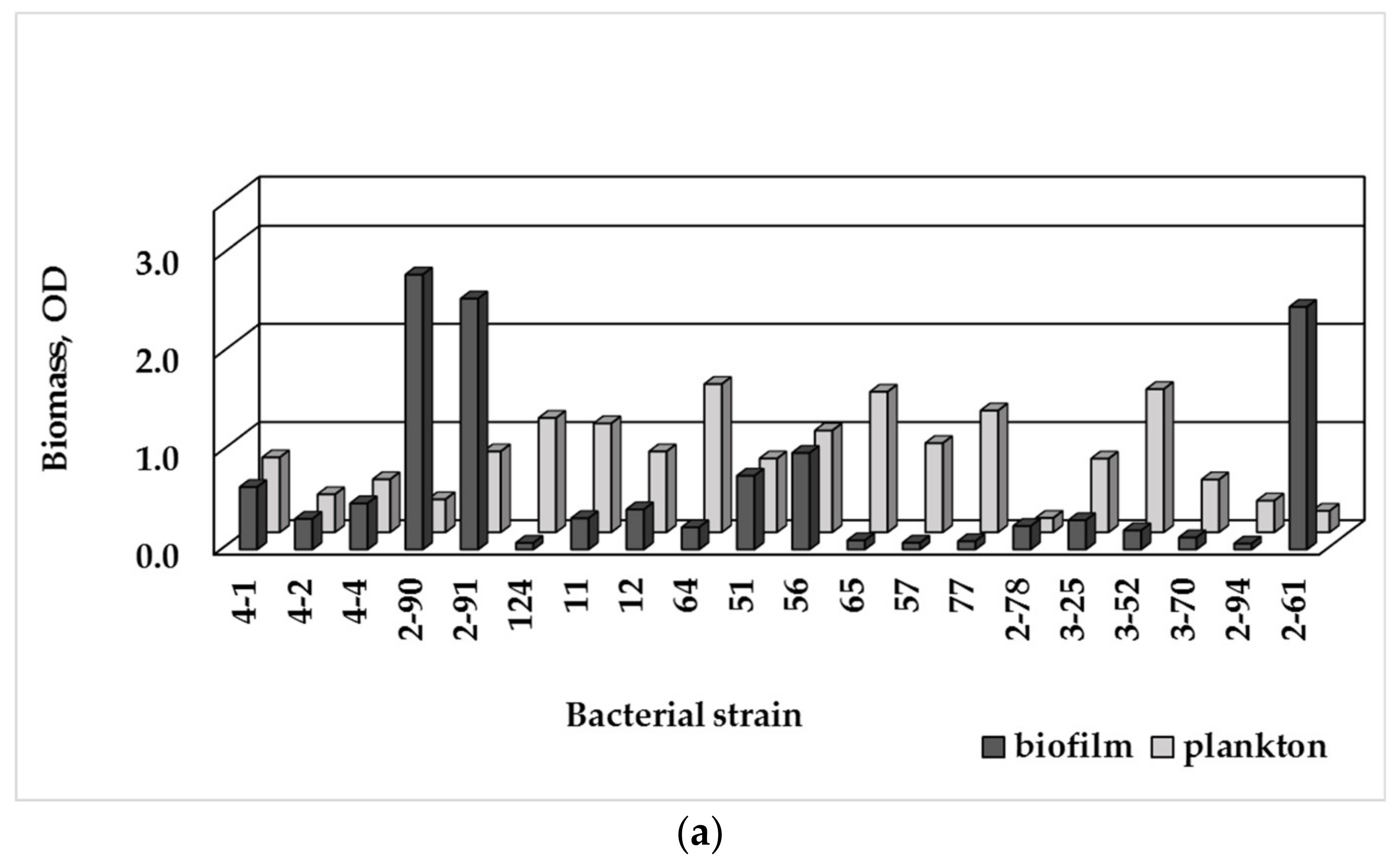
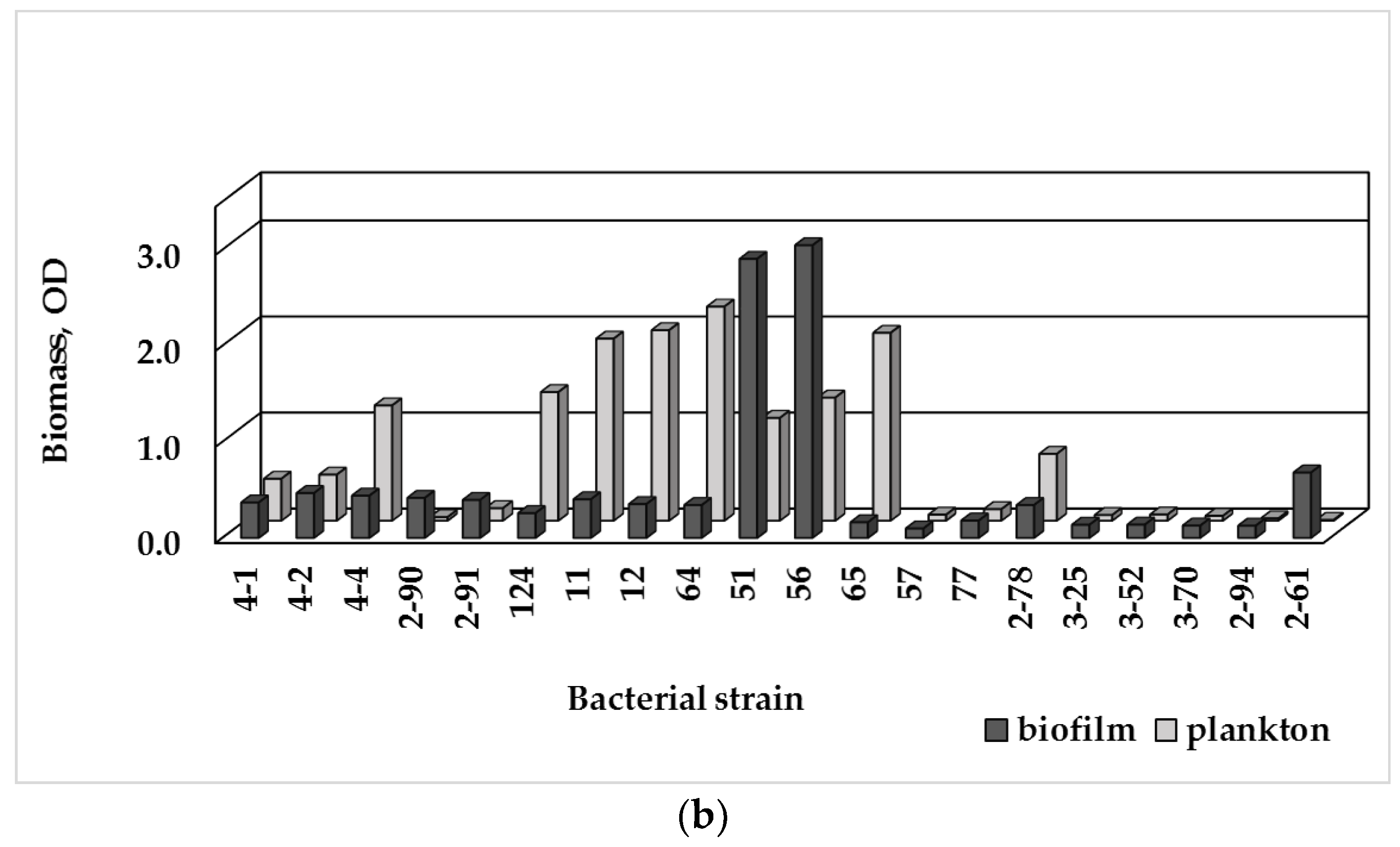
The tested strains formed biofilms in R2A broth (Figure 3a), with E. brevis (2-90; 2-91) producing the largest biomass amount, and Micrococcus luteus (2-61)—the second-strongest biofilm producer. Both strains P. fluorescens (51, 56) produced larger biofilm biomass amount than P. putida and Pseudomonas spp. In M63 medium (Figure 3b), P. fluorescens strains were producers of the largest biofilm amount, followed by Micrococcus luteus (2-61).
Based on the crystal violet assay of the biofilm formation ability (BF), the tested bacteria were classified as non-biofilm producing, weak, moderate or strong biofilm producers (Table 2). In R2A broth, 9 out of 20 isolates (45%) were considered as strong biofilm producers, despite the significant variations in the accumulated biofilm biomass. Six bacterial strains (30%) were moderate biofilm producers, while four other ones had weak biofilm-forming ability (20%), and only Sphingomonas paucimobilis was a non-biofilm former. In M63 medium, seven isolates were strong biofilm producers, while 10 were weak or moderate, and 3 isolates were non-biofilm formers.
Biofilm-forming ability is strain-specific characteristic of a bacterial species. Among the tested Sphingomonas strains, it varied from fully absence of adhesion to strong biofilm formation capacity. Diverse biofilm-formation ability was observed among the tested pseudomonads: P. fluorescens strains were strong biofilm producers, while P. putida and Pseudomonas spp. were moderate. The Methylobacterium strains possessed weak or moderate biofilm formation ability.
The planktonic and biofilm biomass data (plotted on Figure 3) manifesting the preferred growth mode of the tested aquatic bacteria, were used to calculate a ratio, called biofilm index, BI (Table 2). The BI index reveals the growth preferences of the bacterial strains under the conditions of either nutrient media. In oligotrophic R2A broth, under low but diverse level of nutrients, most isolates grew actively as bacterial suspension with BI ≤ 1; only 4 strains (E. brevis, Methylobacterium spp., Micrococcus luteus) manifested significant biofilm growth and BI > 1. In M63 medium, in the presence of glucose as an easily utilized carbon source, six Gram-negative strains and five Gram-positive bacteria preferably grew attached to surface (BI > 1). The tested Gram-positive bacteria exhibited various biofilm formation ability: from strong biofilm producing cocci Micrococcus luteus to weak biofilm- or non-biofilm producing bacilli.
3.4. Biofilm Formation and MDR
The incidence of multidrug-resistant bacteria with strong biofilm-formation ability in the biofilms may pose a hazard to the drinking water quality. Therefore, it is necessary to clarify whether MDR-strains have a stronger biofilm forming potential than the non-MDR ones.
In total, 25% of the tested strains were MDR to at least one AB from three different classes. The summarized data (Table 3) demonstrate the relation between the biofilm-formation ability (BF) or BI of the bacterial isolates and their overall multidrug resistance. In both media, the MDR-strains prevailingly were weak or moderate biofilm producers, while half among the non-MDR strains were strong biofilm producers.
It was found that in R2A only one out of 5 MDR-strains had a strong biofilm formation potential and none of them in M63. However, 8 out of 15 non-MDR strains (53%) have a strong biofilm potential in R2A, and 7 out of 15 (47%) in M63 medium. Besides, Stenotrophomonas maltophilia, the only multidrug resistant to 5 classes of ABs (MDR-5) strain was weak or moderate biofilm-producer, and P. putida with MDR-4 was moderate.
Comparing the MDR and non-MDR strains based on the BI index, no strong biofilm producer was observed among the MDR-strains in R2A broth, and only one was detected in M63 medium, while among the non-MDR bacteria 4 out of 15 (27%) had a strong biofilm forming ability in R2A, and 10 out of 15 (67%) in M63.
4. Discussion
4.1. Prevalence of ARB in Drinking Water-Associated Biofilms
In recent years, the presence of ARB and ARGs in drinking water is considered an emerging health risk. Moreover, the DWDSs serving as a reservoir and an incubator for ARB growth could play an important role for dissemination of AMR through drinking water to human-associated microbiome [4,24,26]. However, the studies were mainly focused on the prevalence of AMR among drinking water microbiome, and the contribution of water-associated biofilms to overall AMR of drinking water was not sufficiently clarified. The risk of ARB reaching the end-users is not only related to the planktonic bacteria, but also to biofilm community with whom they continuously interact. Therefore, the dissemination of AMR among the attached biofilm community is important and needs to be elucidated.
The conducted study assessed the prevalence of AMR among the heterotrophic bacteria in biofilm communities formed in drinking water from four sampling points at the municipal DWDS network of the town of Veliko Tarnovo. It was found that the biofilms associated with the drinking water from the different sampling points significantly differed in population proportion of bacteria resistant to six tested antibiotics (Figure 1b). Those population differences were well manifested by all types of ARB, especially the ciprofloxacin-, chloramphenicol- and streptomycin-resistant bacteria. The distinct biofilm communities exhibited some specific traits: the biofilms in DW-2 and DW-3 contained higher proportion of bacteria resistant to C and lower one of bacteria resistant to CIP, than the rest of the biofilms. The biofilms in DW-4 differed most significantly from all the other biofilms with the highest population level of bacteria resistant to CIP and S, and the lowest ones of bacteria resistant to C and Sul.
The population-based data for AMR of the planktonic and sessile communities are in agreement with the data on the AMR phenotype of the bacteria isolated from drinking water and associated biofilms (Figure 2). The higher number of isolates resistant to S and CIP among the biofilm bacteria comparing to the planktonic isolates is in correspondence to the ARB populations’ data, especially for the biofilms formed in DW-4. The high proportion of streptomycin-resistant bacteria in the DW-1 and the DW-4 biofilms is associated with the abundance of yellow pigment-producing bacteria from the family Sphingomonadaceae, which are known to be resistant to streptomycin. Similarly, some differences could be found among the biofilms formed in drinking waters, differing in chlorination degree. The biofilms in drinking water DW-1 and DW-4, containing 0.3 mg/L residual chlorine, manifested lower number of ARB to chloramphenicol compared to DW-2 and DW-3 (0.7 mg/L residual chlorine), and an opposite trend for bacteria resistant to CIP and S. The prolonged contact with chlorine is known to lead to changes in species diversity and cultivability of aquatic bacteria and the surviving bacteria may have increased innate resistance to chlorine and antimicrobials [24,26,40]. As a result of higher levels of water chlorination, the increased number of Gram-positive bacteria in the aquatic community of DW-2 and DW-3 [30] may affect their inclusion in biofilms and on the overall AMR of biofilm, respectively. Chlorine resistance is known to be strongly associated with the physiological state of bacterial cells, such as their ability to form spores or secrete extracellular polymeric substances [5,8]. However, Zhu et al. (2020) found small relation between the chlorine resistance of single-species biofilms and their biofilm formation potential. The single-species biofilm of the strongest biofilm formation producers was highly sensitive to disinfectant [42].
In DWDSs, biofilm community and free-living bacteria are in a continuous interaction [6,9,43]. Undoubtedly, these two-way interactions determine the level of AMR spreading among the aquatic microbiota at each particular location of a DWDS. The population data obtained for different types of ARB demonstrate such a relationship between the aquatic microbiota and the biofilm community. A comparison between the abundance of different ARB in the water and the biofilm depending on the DW sampling location shows a similar ranking of the population level of the distinct types of ARB. However, the population proportion of some ARB, especially chloramphenicol- and streptomycin-resistant bacteria, was higher in the biofilms comparing with the water. Apart from the microbiological composition of water, biofilm formation depends on physiological condition and motility of bacterial species, their ability to produce exopolymers, inter-species interactions, ability to repair their cellular damages due to the water chlorination, etc. [5,8]. The combination of these factors likely favors, to varying degrees, the attachment and inclusion of each bacterial species in the biofilm community, and thus leading to differences in the AMR level among the sessile and the planktonic populations in the individual DW sampling points.
The AMR differences between the biofilms depending on the sampling point suggest a different degree of their impact on the bacteriological composition of the drinking water, which is supported by the determined HPC values for the different ARB types. It is in agreement to the findings of Xi et al. (2009) for influence of biofilms on the dissemination of AMR through the resistant bacteria they emit in drinking water on its way from the DWTP to the consumers’ tap [24]. Being the predominant mode of life in DWDSs and due to the dispersal of bacteria in bulk water, the drinking water associated biofilms represent important potential source for drinking water contamination with ARB.
4.2. Relation between AMR and Biofilm Formation Ability of Bacterial Isolates
The differences between the populations of bacteria resistant to individual Abs in drinking water and water-associated biofilms suggest looking for a relationship between the AMR of aquatic bacteria and their biofilm-forming ability. Thus, the biofilm-forming ability of 20 bacterial strains with different levels of AMR was assessed. According to the AMR phenotype, the biofilm forming abilities of two groups of bacterial strains—non-MDR bacteria and MDR-bacteria, were compared.
The results show that the MDR-strains very rarely possessed strong biofilm-formation ability, unlike the non-MDR strains, about half of which were strong biofilm producers. Despite our study including various species of aquatic bacteria, the results are in agreement with the findings from studies on single species of clinically important pathogens, the sensitive and non-MDR strains, of which it has been reported to exhibited strong biofilm-forming potential, unlike that of the weak biofilms produced by the MDR-bacteria [14,16]. Qi et al. (2020) have found that Acinetobacter baumannii isolates with higher level of AMR tended to form weaker biofilms and that population exhibited more robust biofilm formation likely contained larger proportion of non-MDR bacteria. The authors considered that biofilm formation acts as a mechanism for a better survival of pathogenic bacteria with low or middle level of resistance, while the weak biofilms of isolates with high level of resistance could provide a similar level of protection [16]. However, the results, even for single species, have not been always unambiguous. For example, strong biofilm-forming clinical strains of Staphyloccocus epidermidis have been MDR to three or more antimicrobial classes, as the isolate with maximum biofilm-forming ability have been resistant to seven antimicrobial classes [44]. Eyoh et al. (2014) found insignificant difference in the percentage of MDR among the strong biofilm producing and weak biofilm- or non-biofilm-producing strains of Staphylococcus aureus [15]. Unlike the biofilms of clinically important bacterial pathogens, which provide protection from the therapeutic action of antibiotics, the biofilm formation in drinking water at the conditions of absent or extremely low content of residual ABs, and under the exposure to other environmental stress, is subjected to different survival strategies of the aquatic bacteria. Nevertheless, the predominant strong biofilm-forming ability of the aquatic bacteria with low antibiotic resistance levels (non-MDR bacteria) is of a particular importance due to the potential impact of biofilms on drinking water quality and the provision of favorable conditions for acquisition and transfer of AMR determinants. Moreover, resistance determinants not detected in drinking water could be proved in water-associated biofilms [23].
The simplified single-species biofilm test is an important tool to understand the contributions of individual species to community functions. However, the results for single-species biofilm formation in relation with its phenotype of AMR cannot be easily extrapolated to the complicated drinking water-associated biofilms. Numerous data have demonstrated that interspecies interactions in biofilms can radically change the response of community under the exposure to disinfectants and antimicrobials [45,46]. Burmolle et al. (2006) have determined increased AMR to tetracycline of four-species biofilm compared to single-species biofilm, while Lee et al. (2014) have found increased resistance to sodium dodecyl sulfate and tobramycin. The increased cells protection within the multispecies biofilm to disinfectants have suggested for an increased fitness provided by the partners in the consortium [37,46].
The conducted population studies of ARBs in the biofilms and the AMR phenotype and the biofilm-forming potential of isolated aquatic bacteria provide complementary information on the prevalence of AMR among the heterotrophic bacteria in drinking water-associated biofilms. The increased population share of some types of ARB in the biofilm could be due to the strong biofilm-forming ability exhibited by at least half of the tested bacterial isolates. The tested MDR-bacteria had a weak biofilm-forming ability, but in partnership with other species they could reside into the biofilm. Close cell-to-cell contact in biofilm community could favor transferring of genetic determinants and increasing of AMR. These data support the hypothesis that biofilm formation ability of bacteria is means for a better survival in hostile environment, especially of bacteria susceptible to antibiotics or with low level of antimicrobial resistance. Our next study will be focused on interspecies interactions in multispecies biofilms and the contribution of some of the isolated bacteria to the community response to some antimicrobials.
5. Conclusions
The study assessed the AMR of heterotrophic bacteria in the biofilms formed in drinking water from four sampling points of a municipal DWDS network. The population proportion of the ARB in the biofilms differed significantly depending on the DW sampling point and the type of AB, to which they were resistant.
The drinking water-associated biofilms show some general trends in the proportion of the distinct types of ARB, such as the smallest population of the bacteria resistant to tetracycline and ampicillin, and the largest one of bacteria resistant to streptomycin. The population share of some types of ARB in the biofilm community increased in comparison with the free-living aquatic bacteria. The population changes of some types of ARB in the biofilm, depending on the water chlorination effect on the aquatic microbiota, were evidenced. The increased population proportion of some types of ARB in the biofilms implies an increased impact on the bacterial composition of drinking water. The prevalence of ARB in the biofilm communities and their spreading into drinking water has a particular significance, considering the impact on human-associated microbiota. Since biofilm is the predominant mode of bacterial organization in DWDSs and due to the bacteria dispersal in water, the drinking water associated biofilms represent important potential source for contamination of drinking water with ARB.
The increased share of some types of ARB in the biofilms compared to drinking water show that the data on their occurrence in water do not provide sufficiently reliable and representative information on the abundance of ARBs in the biofilm community. Therefore, in order to clarify the antimicrobial resistance of the microbiome in a DWDS, a comprehensive assessment of both the attached and the planktonic populations is needed.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w14060944/s1; A scheme of the DWDS; Tables S1 and S2. Data plotted on Figure 1; Tables S3 and S4. Data plotted on Figure 3.
Author Contributions
Conceptualization, Z.T.; methodology, Z.T. and H.N.; investigation, Z.T. and I.T.; data curation, Z.T.; writing—original draft preparation, Z.T.; writing—review and editing, H.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Fund, Republic of Bulgaria (grant number: КП-06-Н31/20 from 17 December 2019.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in Supplementary Materials.
Acknowledgments
The scientific advices of V. Kussovski and the technical assistance of V. Slaveva are greatly appreciated.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AB | Antibiotic |
| AMR | Antimicrobial resistance |
| AMP | Ampicillin |
| ANOVA | One-way analysis of variance |
| ARB | Antibiotic resistant bacteria |
| ARGs | Antibiotic resistance genes |
| BF | Biofilm-formation ability |
| BI | Biofilm index |
| C | Chloramphenicol |
| CIP | Ciprofloxacin |
| CFU | Colony form unit |
| CTX | Cefotaxime |
| DW | Drinking water |
| DWDS | Drinking water distribution system |
| FF | Fosfomycin |
| FM | Nitrofurantoin |
| GEN | Gentamycin |
| HPC | Heterotrophic plate count |
| MDR | Multidrug resistance |
| OD | Optical density |
| ODc | Cutoff OD |
| P | Probability |
| S | Streptomycin |
| STX | Trimethoprim/sulfamethoxazole |
| Sul | sulphamethoxazole |
| TE | Tetracycline |
| TMP | Trimethoprim |
References
- Manaia, C.M.; Macedo, G.; Fatta-Kassinos, D.; Nunes, O.C. Antibiotic resistance in urban aquatic environments: Can it be controlled? Appl. Microbiol. Biotechnol. 2016, 100, 1543–1557. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marti, Е.; Jofre, J.; Balcazar, J.L. Prevalence of antibiotic resistance genes and bacterial community composition in a river influenced by a wastewater treatment plant. PLoS ONE 2013, 8, e78906. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Bacterial diversity and antibiotic resistance in water habitats: Searching the links with the human microbiome. FEMS Microb. Rev. 2014, 761–778. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, A. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Flemming, H.C.; Percival, S.I.; Walker, J.T. Contamination potential of biofilms in water distribution systems. Water Sci. Technol. 2002, 2, 271–280. [Google Scholar] [CrossRef]
- Wricke, B.; Korth, A.; Petzoldt, H.; Kruger, M. Change of bacterial water quality in drinking water distribution systems working with or without low chlorine residual. Water Sci. Technol. Water Supply 2002, 2, 275–281. [Google Scholar] [CrossRef]
- LeChevallier, M.W.; Cawthonm, C.D.; Leem, R.G. Inactivation of biofilm bacteria. Appl. Environ. Microbiol. 1988, 54, 2492–2499. [Google Scholar] [CrossRef] [Green Version]
- Codony, F.; Morato, J.; Mas, J. Role of discontinuous chlorination on microbial production by drinking water biofilms. Water Res. 2005, 39, 1896–1906. [Google Scholar] [CrossRef]
- Tsvetanova, Z. Quantification of the bacterial community of drinking water associated biofilms under different flow velocities and changing chlorination regimes. Appl. Water Sci. 2020, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef]
- Chen, X.; Stewart, P.S. Biofilm removal caused by chemical treatments. Water Res. 2000, 34, 4229–4233. [Google Scholar] [CrossRef]
- Ito, A.; Taniuchi, A.; May, T.; Kawata, K.; Okabe, S. Increased antibiotic resistance of Escherichia coli in mature biofilms. Appl. Environ. Microbil. 2009, 75, 4093–4100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moskowitz, S.M.; Foster, J.M.; Emerson, J.; Burns, J.L. Clinically feasible biofilm susceptibility assay for isolates of Pseudomonas aeruginosa from patients with cystic fibrosis. J. Clin. Microbiol. 2004, 42, 1915–1922. [Google Scholar] [CrossRef] [Green Version]
- Eyoh, A.B.; Toukam, M.; Atashili, J.; Fokunang, C.; Gonsu, H.; Lyonga, E.E.; Mandi, H.; Ikomey, G.; Mukwele, B.; Mesembe, M.; et al. Relationship between multiple drug resistance and biofilm formation in Staphylococcus aureus isolated from medical and non-medical personnel in Yaounde, Cameroon. Pan Afr. Med. J. 2014, 17, 186. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Li, H.; Zhang, C.; Liang, B.; Li, J.; Wang, L.; Du, X.; Liu, X.; Qiu, S.; Song, H. Relationship between antibiotic resistance, biofilm formation, and biofilm-specific resistance in Acinetobacter baumannii. Front. Microbiol. 2016, 7, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Balcázar, J.L.; Subirats, J.; Borrego, C.M. The role of biofilms as environmental reservoirs of antibiotic resistance. Front. Microbiol. 2015, 6, 1216. [Google Scholar] [CrossRef] [Green Version]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Linares, J.F.; Gustafsson, I.; Baquero, F.; Martinez, J.L. Antibiotics as intermicrobial signaling agents instead of weapons. Proc. Natl. Acad. Sci. USA 2006, 103, 19484–19489. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, J.B. Antibiotic-induced biofilm formation. Int. J. Artif. Organs 2011, 34, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Farkas, A.; Butiuc-Keul, A.; Ciatarâş, D.; Neamţu, C.; Crăciunaş, C.; Podar, D.; Drăgan-Bularda, M. Microbiological contamination and resistance genes in biofilms occurring during the drinking water treatment process. Sci. Total Environ. 2013, 443, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, T.; Kohnen, W.; Jansen, B.; Obst, U. Detection of antibiotic-resistant bacteria and their resistance genes in waste-water, surface water and drinking water biofilms. FEMS Microbiol. Ecol. 2003, 43, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Xi, C.; Zhang, Y.; Marrs, C.F.; Ye, W.; Simon, C.; Foxman, B.; Nriagu, J. Prevalence of antibiotic resistance in drinking water treatment and distribution systems. Appl. Environ. Microbiol. 2009, 75, 5714–5718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narciso-da-Rocha, C.; Vaz-Moreira, I.; Svensson-Stadler, L.; Moore, E.R.B.; Manaia, C.M. Diversity and antibiotic resistance of Acinetobacter spp. in water from the source to the tap. Appl. Microbiol. Biotechnol. 2013, 97, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Beattie, T.K.; Knapp, C.W. Relationship between antibiotic- and disinfectant-resistance profiles in bacteria harvested from tap water. Chemosphere 2016, 152, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Destiаni, R.; Templeton, M.R. The antibiotic resistance of heterotrophic bacteria in tap waters in London. Water Supply 2019, 19, 179–190. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Diversity and antibiotic resistance patterns of Sphingomonadaceae isolates from drinking water. Appl. Environ. Microbiol. 2011, 77, 5697–5706. [Google Scholar] [CrossRef] [Green Version]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Diversity and antibiotic resistance in Pseudomonas spp. from drinking water. Sci. Total Environ. 2012, 426, 366–374. [Google Scholar] [CrossRef]
- Tsvetanova, Z.; Dimitrov, D.; Najdenski, H. Prevalence of antimicrobial resistance in a Bulgarian drinking water supply system. Water Supply 2022, submitted.
- Reasoner, D.J.; Geldreich, E. A new medium for enumeration and subculture of bacteria from potable water. Appl. Environ. Microbiol. 1985, 49, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- EUCAST. European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation pf MICs and Zone Diameters. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_11.0_Breakpoint_Tables.pdf (accessed on 20 February 2022).
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 30, 2437. Available online: http://www.jove.com/details.php?id=2437 (accessed on 16 March 2022). [CrossRef]
- Simões, L.C.; Simões, M.; Vieira, M.J. Influence of the diversity of bacterial isolates from drinking water on resistance of biofilms to disinfection. Appl. Environ. Microbiol. 2010, 76, 6673–6679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burmølle, M.; Webb, J.S.; Rao, D.; Hansen, L.H.; Sørensen, S.J.; Kjelleberg, S. Enhanced biofilm formation and increased resistance to antimicrobial agents and bacterial invasion are caused by synergistic interactions in multispecies biofilms. Appl. Environ. Microbiol. 2006, 72, 3916–3923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boe-Hansen, R.; Albrechtsen, H.J.; Arvin, E.; Jørgensen, C. Dynamics of biofilm formation in a model drinking water distribution system. J. Water SRT Aqua 2002, 1, 399–406. [Google Scholar] [CrossRef]
- Kelly, J.J.; Minalt, N.; Culotti, A.; Pryor, M.; Packman, A. Temporal variations in the abundance and composition of biofilm communities colonizing drinking water distribution pipes. PLoS ONE 2014, 9, e98542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norton, C.D.; LeChevallier, M.W. A pilot study of bacteriological population changes through potable water treatment and distribution. Appl. Environ. Microbiol. 2000, 66, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Tsvetanova, Z.; Najdenski, H. Biofilm formation potential of enteropathogenic bacteria and their survival in drinking water-associated biofilms. Acta Microbiol. Bulg. 2018, 34, 153–159. [Google Scholar]
- Zhu, Z.; Shan, L.; Hu, F.; Li, Z.; Zhong, D.; Yuan, Y.; Zhang, J. Biofilm formation potential and chlorine resistance of typical bacteria isolated from drinking water distribution systems. RSC Adv. 2020, 10, 31295. [Google Scholar] [CrossRef]
- Douterelo, I.; Jackson, M.; Solomon, C.; Boxall, J. Microbial analysis of in situ biofilm formation in drinking water distribution systems: Implications for monitoring and control of drinking water quality. Appl. Microbiol. Biotechnol. 2016, 100, 3301–3311. [Google Scholar] [CrossRef] [Green Version]
- Sahal, G.; Bilkay, I.S. Multi drug resistance in strong biofilm forming clinical isolates of Staphylococcus epidermidis. Braz. J. Microbiol. 2014, 45, 539–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simões, M.; Simões, L.C.; Vieira, M.J. Species association increases biofilm resistance to chemical and mechanical treatment. Water Res. 2009, 43, 229–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.W.K.; Periasamy, S.; Mukherjee, M.; Xie, C.; Kjelleberg, S.; Rice, S.A. Biofilm development and enhanced stress resistance of a model mixed-species community biofilm. SME J. 2014, 8, 894–907. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Relative percentage share of the HPC bacteria resistant to the tested ABs in drinking water (a) and drinking water-associated biofilms (b) depending on sampling points at the DWDS.
Figure 1.
Relative percentage share of the HPC bacteria resistant to the tested ABs in drinking water (a) and drinking water-associated biofilms (b) depending on sampling points at the DWDS.


Figure 2.
Phenotype of antimicrobial resistance of bacteria isolated from biofilms (a) and drinking water (b), assessed by disk diffusion method.
Figure 2.
Phenotype of antimicrobial resistance of bacteria isolated from biofilms (a) and drinking water (b), assessed by disk diffusion method.

Figure 3.
Biomass amount of the single-species biofilms of the isolated bacterial strains measured as OD590nm value and the plankton biomass measured as OD620nm value: (a) in R2A broth; (b) in M63 medium with 0.1% glucose (Detailed data in Supplementary Materials, Tables S3 and S4).
Figure 3.
Biomass amount of the single-species biofilms of the isolated bacterial strains measured as OD590nm value and the plankton biomass measured as OD620nm value: (a) in R2A broth; (b) in M63 medium with 0.1% glucose (Detailed data in Supplementary Materials, Tables S3 and S4).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Number of HPC bacteria in drinking water and water-associated biofilms.
| Sampling Point | HPCs of Drinking Water, CFU/100 mL | HPC Density of the Biofilm Formed, CFU/cm2 |
|---|---|---|
| DW-1 | 2.9 (0.2) × 102 | 3.3 (2.4) × 104 |
| DW-2 | 7.9 (0.2) × 101 | 1.2 (1.2) × 104 |
| DW-3 | 1.3 (0.1) × 101 | 3.9 (2.1) × 105 |
| DW-4 | 1.6 (0.1) × 103 | 2.0 (0.9) × 105 |
Table 2.
AMR and biofilm formation ability of the bacterial strains isolated from drinking water.
| Strain № | Bacterial Taxon | AMR Phenotype | MDR, n | R2A | M63 | ||
|---|---|---|---|---|---|---|---|
| BI | BF | BI | BF | ||||
| 4-1 | Sphingomonas spp. | AMP, S | 2 | 0.8 | +++ | 0.8 | +++ |
| 4-2 | Sphingomonas spp. | AMP, S | 2 | 0.8 | +++ | 1.0 | +++ |
| 4-4 | Sphingomonas spp. | AMP, S | 2 | 0.9 | +++ | 0.4 | +++ |
| 124 | Sphingomonas paucimobilis | AMC, CZ, CN, CXM, FM | 2 | 0.1 | 0 | 0.2 | + |
| 2-90 | Embedobacter brevis | AMC, CZ, CN, CXM, FM | 2 | 7.9 | +++ | 11.3 | ++ |
| 2-91 | Embedobacter brevis | AMC, CZ, CN, CXM, FM | 2 | 3.1 | +++ | 3.0 | +++ |
| 11 | Pseudomonas spp. | AMP, AMC, CTX, CAZ, SXT, TE | 3 | 0.3 | ++ | 0.2 | ++ |
| 12 | Pseudomonas spp. | AMP, AMC, CTX, CAZ, SXT, TE | 3 | 0.5 | +++ | 0.2 | ++ |
| 64 | Pseudomonas spp. | AMP, CF, CTX, C, SXT | 3 | 0.1 | ++ | 0.2 | ++ |
| 56 | P. fluorescens | AMC, CN, CXM, CZ, FM | 2 | 0.9 | +++ | 2.4 | +++ |
| 51 | P. fluorescens | AMC, CN, CXM, CZ, FM | 2 | 1.0 | +++ | 2.7 | +++ |
| 65 | P. putida | AMP, AMC, CZ, CAZ, C, CIP, FM | 4 | 0.1 | ++ | 0.1 | ++ |
| 57 | Stenotrophomonas maltophilia | AMP, AMC, AN, ATM, CN, CTX, CXM, CZ, ETP, FF, FM, GEN, IPM, MEM, NN, TMP, TZP | 5 | 0.1 | + | 1.6 | ++ |
| 2-78 | Methylobacterium exorquens | sensitive | 0 | 1.6 | ++ | 0.5 | ++ |
| 77 | Methylobacterium spp. | SXT | 1 | 0.1 | + | 1.5 | + |
| 3-70 | Lysinibacillus sphaericus | S, NA | 2 | 0.2 | + | 2.6 | 0 |
| 3-25 | Bacillus spp. | Amp, CTX, CX, E | 2 | 0.4 | ++ | 2.3 | 0 |
| 3-52 | B. thuringiensis | AMP, AMC, CTX, CX, SXT | 2 | 0.1 | ++ | 2.2 | 0 |
| 2-94 | Staphylococcus cohnii | CIP, MXP | 2 | 0.2 | + | 5.4 | ++ |
| 2-61 | Micrococcus luteus | sensitive | 0 | 11.4 | +++ | 87.9 | +++ |
Legend: R2A and M63—nutrient media used in biofilm formation test; MDR—multidrug resistance to number of antibiotic classes, n; BI—biofilm index; BF—biofilm forming ability: (+++)—strong; (++)—moderate; (+)—weak; 0—non-biofilm producer; Antibiotics: AMP—ampicillin; AMC—amoxicillin/clavulanic acid; AN—amikacin; ATM—aztreonam; C—chloramphenicol; CAZ—ceftazidime; CN—cephalexin; CTX—cefotaxime; CZ—cefazolin; CF—cephalothin; CX—cefoxitin; CXM—cefuroxime; CIP—ciprofloxacin, E—erythromycin; ETP—ertapenem; FF—fosfomycin; FM—nitrofurantoin; GEN—gentamicin; IPM—imipenem; MEM—meropenem; MXP—moxifloxacin; NA—nalidixic acid; NN—tobramycin; S—streptomycin; STX—trimethoprim/sulfamethoxazole; TE—tetracycline; TMP—trimethoprim; TZP—piperacillin/tazobactam.
Table 3.
Summarized data on the biofilm formation ability of the tested MDR- and non-MDR bacterial stains.
Table 3.
Summarized data on the biofilm formation ability of the tested MDR- and non-MDR bacterial stains.
| BF or BI | Number of Strains in R2A (n) | Number of Strains in M63 (n) | ||
|---|---|---|---|---|
| MDR (5) | Non-MDR (15) | MDR (5) | Non-MDR (15) | |
| Strong (+++) | 1 | 8 | 0 | 7 |
| Weak (++; +) | 4 | 6 | 5 | 5 |
| Strong (BI > 1) | 0 | 4 | 1 | 10 |
| Weak (BI ≤ 1) | 5 | 11 | 4 | 5 |
Legend: BF—biofilm-forming ability; BI—biofilm index.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tsvetanova, Z.; Tsvetkova, I.; Najdenski, H. Antimicrobial Resistance of Heterotrophic Bacteria in Drinking Water-Associated Biofilms. Water 2022, 14, 944. https://doi.org/10.3390/w14060944
AMA Style
Tsvetanova Z, Tsvetkova I, Najdenski H. Antimicrobial Resistance of Heterotrophic Bacteria in Drinking Water-Associated Biofilms. Water. 2022; 14(6):944. https://doi.org/10.3390/w14060944
Chicago/Turabian StyleTsvetanova, Zvezdimira, Iva Tsvetkova, and Hristo Najdenski. 2022. "Antimicrobial Resistance of Heterotrophic Bacteria in Drinking Water-Associated Biofilms" Water 14, no. 6: 944. https://doi.org/10.3390/w14060944
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.