
Distribution and Transfer of Antibiotic Resistance Genes in Coastal Aquatic Ecosystems of Bohai Bay

1
Tianjin Fisheries Research Institute, Tianjin 300200, China
2
Southern Marine Science and Engineering Guangdong Laboratory-Zhanjiang, Fisheries College, Guangdong Ocean University, Zhanjiang 524000, China
*
Author to whom correspondence should be addressed.
Water 2022, 14(6), 938; https://doi.org/10.3390/w14060938
Submission received: 29 January 2022
/
Revised: 3 March 2022
/
Accepted: 14 March 2022
/
Published: 17 March 2022
(This article belongs to the Topic Aquatic Emerging Contaminants and Their Ecotoxicological Consequences)
Abstract
:Antibiotic resistance genes (ARGs) are abundant in diverse ecosystems and the resistome may constitute a health threat for humans and animals. It is necessary to uncover ARGs and the accumulation mechanisms from different environmental sources. Various habitats, such as soil, seawater and fish intestines, could overflow a considerable amount of ARGs and the horizontal transfer of ARGs may occur in these environments. Thus, we assessed the composition and abundance of ARGs in seawater, soil and intestinal tracts of Cynoglossus semilaevis collected from different sites in Bohai Bay (China), including a natural area and three fish farms, through a high-throughput qPCR array. In total, 243 ARGs were uncovered, governing the resistance to aminoglycoside, multidrug, beta-lactamase, macrolide lincosamide streptomycin B (MLSB), chloramphenicol, sulfonamide, tetracycline, vancomycin and other antibiotics. The action mechanisms of these ARGs were mainly antibiotic deactivation, efflux pump and cellular protection. Importantly, similar ARGs were detected in different samples but show dissimilar enrichment levels. ARGs were highly enriched in the fish farms compared to the natural sea area, with more genes detected, while some ARGs were detected only in the natural sea area samples, such as bacA-02, tetL-01 and ampC-06. Regarding sample types, water samples from all locations shared more ARGs in common and held the highest average level of ARGs detected than in the soil and fish samples. Mobile genetic elements (MGEs) were also detected in three sample types, in the same trend as ARGs. This is the first study comparing the resistome of different samples of seawater, soil and intestines of C. semilaevis. This study contributes to a better understanding of ARG dissemination in water sources and could facilitate the effective control of ARG contamination in the aquatic environment.
1. Introduction
Antibiotic resistance genes (ARGs) are genes governing the expression of enzymes involved in the degradation or in the chemical modification and subsequent inactivation of antibiotics [1]. The emergence of bacterial strains resistant to many antibiotics could be called into questions when using these antibiotics to fight against infectious diseases [2,3]. In the face of this public health problem, it is necessary to define the elements responsible for the emergence of these multiresistant bacteria.
It has now been established that the environment represents a reservoir of resistant bacteria or ARGs that could be transmissible to humans and animals [4]. The occurrence of ARGs in hospital settings, fish farms, organic waste products, wastewater treatment plants and soils/sediments has also been investigated [5,6,7,8,9,10,11]. Many organic waste products are used as fertilizer on agricultural soils [12] and treated wastewater or river water into which they are discharged can be used to irrigate crops [13]. Certain agricultural practices can, therefore, potentially lead to the environmental dissemination of ARGs and the contamination of animals, crops, soils and water resources [14,15]. For example, previous studies have reported that pond water in fish farms and shrimp farms contained ciprofloxacin, penicillin G, rifampicin and vancomycin-related ARGs [16]. The persistence of ARGs in the soils of fish farms has been strongly compared to the surrounding sediments [6].
Aquatic ecosystems constitute a big reservoir of ARGs. It was previously reported that aquaculture systems have a significant impact on the accumulation and spread of antibiotic-resistant bacteria (ARB) and ARGs in freshwater and sediment [16,17]. Fish may ingest ARB from the aquatic environment for their unique living environments, so the fish intestine would be an optimal niche for conjugal ARG transfer. In previous studies, it was reported that the guts of aquatic animals such as zebrafish contained a high abundance and diversity of bacteria, which indicated them as significant potential recipients of ARGs through horizontal gene transfer (HGT) and was beneficial to the spread of ARGs in water environments [18,19]. Moreover, the unique living environments are able to cause aquatic animals to ingest ARB in water easily [19]. There were some data regarding the abundance and the transfer of ARGs among water, sediment (soil) and organisms in aquatic systems [20,21], but the research topic needs an in-depth study. Here, we discuss the abundance of ARGs in aquaculture environments as well as fish ecosystems and the occurrence of the transfer of ARGs among these micro-systems.
The Chinese tongue sole (Cynoglossus semilaevis) is a marine fish species widely spread in the Bohai Sea and cultured in the area; however, not enough attention has been paid to the distributions of ARGs related to this fish till now. Based on the above, this study intends to investigate the different classes of ARGs present in the intestinal tract of C. semilaevis, culture water and soils in the same area. We also aimed to investigate the exchange and transfer mechanisms of ARGs between these ecosystems. This work hopes to provide a better understanding of the abundance and dissemination of ARGs in aquatic ecosystems.
2. Materials and Methods
2.1. Study Sites and Sample Collection
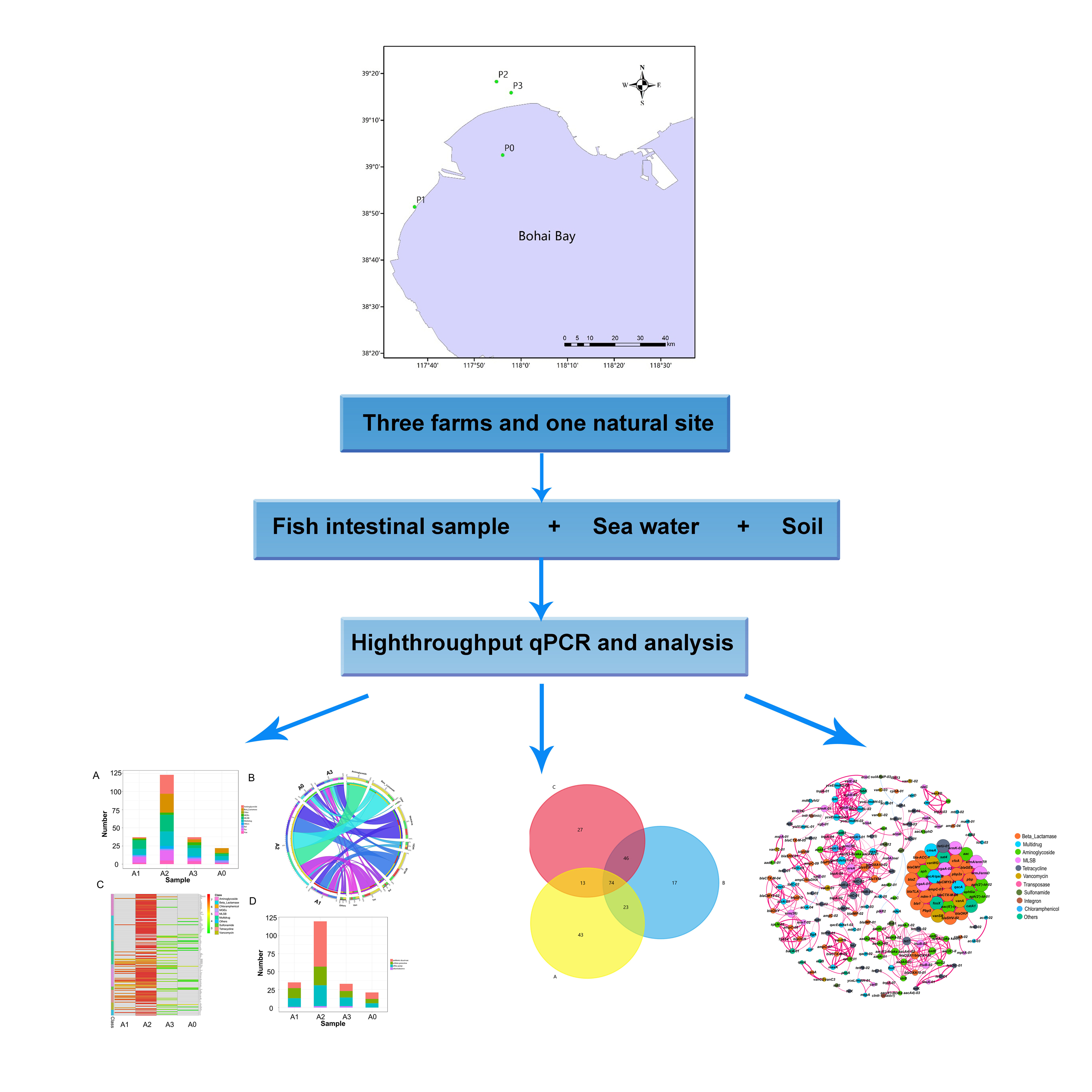
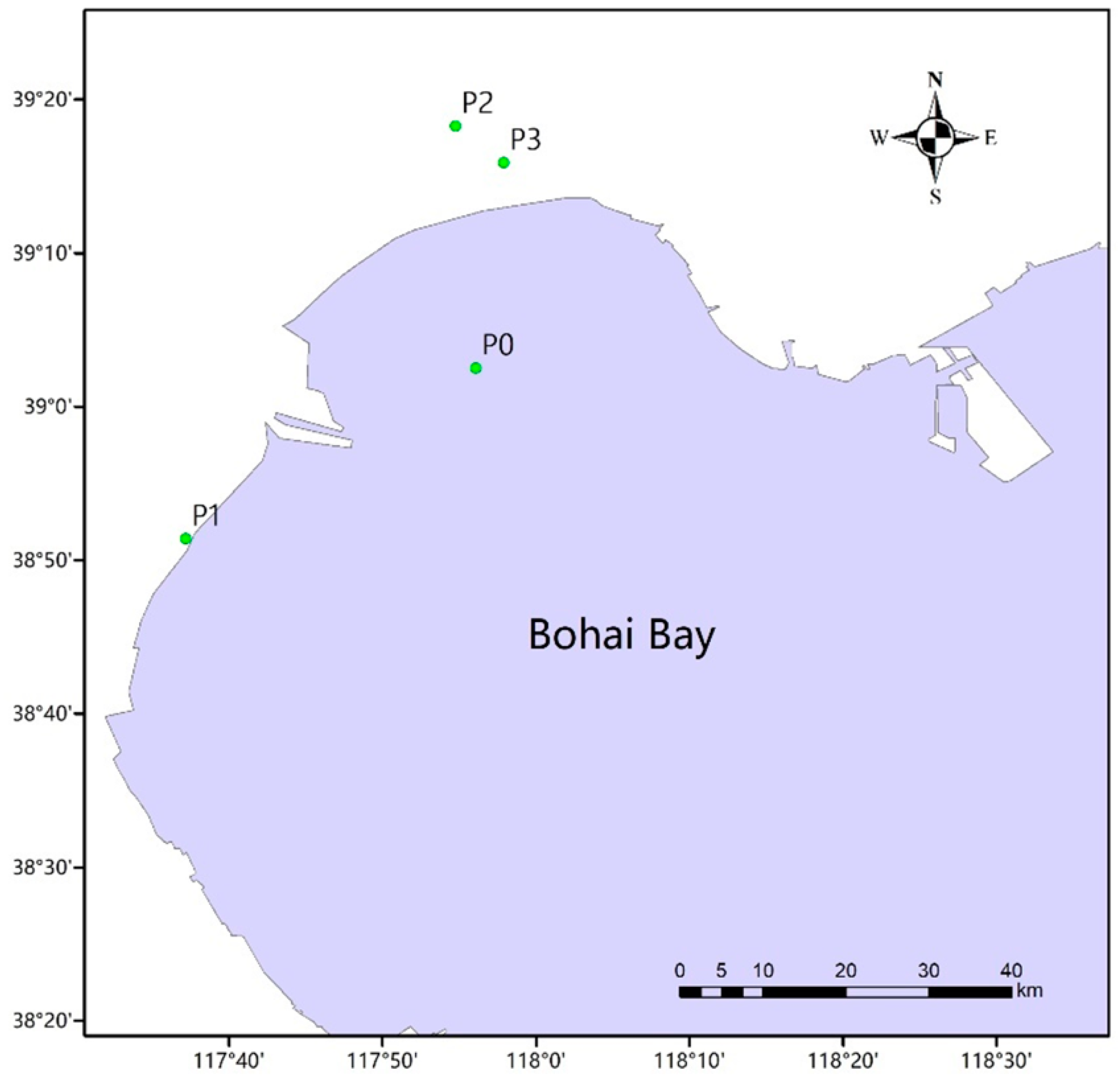
The sampling sites selected for this study are located in Bohai Bay, which is situated in the west of the Bohai Sea, in northeast China (Figure 1). The four sampling sites were as follows: P0 (117.935° E, 39.0415° N), P1 (117.626544° E, 38.852927° N), P2 (117.912634° E, 39.304451° N) and P3 (117.964958° E, 39.264878° N). There were three types of samples, including intestinal tracts of C. semilaevis (A), sea water (B) and soil (C), collected from the natural sea area or fish farms. Fish samples A0–A3 (C. semilaevis) were intestinal tracts collected using sterile plastic bags, followed by separation with sterile forceps and homogenization in a sterile mortar with liquid nitrogen. A volume of 5 L of water samples so-called B0–B3 were collected from 30 cm below the surface with a stainless steel sampler, and then transferred to sterile polyethylene bottles. For the extraction of DNA, 200 mL of each sample B was filtered with vacuum filtration apparatus with a sterile membrane filter (0.22 μm pore diameter), which was aseptically stored at −80 °C. About 100 g of dry soil samples C0–C3 was collected from a depth of 0–10 cm with the help of a grab sampler and gently mixed using sterile polyethylene bottles. For the fish farm area, soil at the drain outlet was sampled. Soil samples were then freeze-dried, ground with a mortar and sieved through a 100-mesh screen. All samples were collected in three replicates, rapidly stored in a portable ice box, which were then transported to the laboratory and the pretreatment was performed within 1 day (Table 1).
2.2. DNA Extraction and Purification
According to the manufacturer’s instructions and previous studies [4,22], total DNA was extracted from samples A, B and C using the DNeasy PowerWater Kit (QIAGEN Sciences, Germantown, MD, USA), the DNeasy PowerSoil Kit (QIAGEN Sciences, Germantown, MD, USA) and the QIAamp DNA Stool Mini Kit (QIAGEN Sciences, Germantown, MD, USA), respectively. A Nanodrop 1000™ spectrophotometer (Thermo Scientific, Wilmington, DE, USA) was used to analyze the DNA quality and concentration.
2.3. High-Throughput Quantitative PCR
Detection of ARGs was performed using the Wafergen Smartchip ultra-high-throughput fluorescent quantitative PCR system according to the method of Zhou [4], by controlling the total DNA concentration of bacteria in different sampling media identically. In this experiment, 296 pairs of primers were set up, including 285 pairs of primers for ARGs, 9 pairs of primers for MGEs, 1 pair of primers for the clinical class 1 integron-integrase gene and 1 pair of 16s rRNA internal reference primers to normalize the abundance of ARGs in different sample types [22]. The PCR mixture was introduced into a microwell chip using a nanoscale multi-sample spotter (MSND) 296 (assays) × 16 (samples) mode, followed by qPCR on the cycler. One no-template control (NTC) with three repetitions was set for each assay in each chip for each sample. The reaction system included 1 × LightCycler® 480 SYBR Green I Master, 500 nM of each primer and 2 ng/μL DNA template. The total reaction volume was 100 nL. PCR reaction conditions were as follows: predenaturation at 95 °C, 10 min, 40 cycles of 95 °C for 30 s; 60 °C for 30 s; dissolution curves. The qPCR results were analyzed through the qPCR software of the instrument. CT = 31 was set as the detection domain, and two or more of the three replicates were detected, with the deviation < 20%. All 16s rRNA internal reference genes of all samples with concentration ≥10 ng/µL in this project were detected, and NTC was not amplified, which proved that the experimental results were credible. The absolute copy number of each gene was calculated indirectly based on the relative copy number. The formula used for calculation was as follows: absolute copy number of target gene = 16 s rRNA gene absolute copy numbe × 2−ΔCT, which means the absolute abundance of each gene was recorded as the product of the relative copy number and the 16S rRNA’s absolute copy number, where in ΔCT = CT (gene)-CT (16 s). It is worth noting that CT (gene) and CT (16 s) were from the same sample.
2.4. Statistical Analysis
Distribution of ARG type in each sample was visualized via Circos graphs, which were carried out by Circos software (http://circos.ca/; accessed on 17 October 2021) online, while Heatmap graphs were produced using RStudio with pheatmap package. The 2−ΔCT abundance table was used to calculate the Pearson Correlation Coefficient and the Bray–Curtis distance. Adonis and Anosim analyses were then conducted using the Bray–Curtis distance matrix. Ordinary least square (OLS) graphs were drawn based on the 2−ΔCT abundance table. The correlation analysis among ARGs and MEGs was performed using the Hmisc package in R. One-way ANOVA test in SPSS version 26.0 was used to analyze the statistical differences of quantity of ARGs between fish farms and the adjacent natural sea area.
3. Results
3.1. Distribution, Classification and Abundance of ARGs in Different Environments Associated with C. semilaevis Farming Industry
The results of the high-throughput PCR indicated that a variety of ARGs could be found in different kinds of samples (Figure S1 in Supplementary Materials). The 243 detected ARGs included genes of eight distinct and predominant classes, conferring resistance to antibiotics such as aminoglycosides, multidrug, beta-lactamases, macrolide lincosamide streptomycin B (MLSB), chloramphenicol, sulfonamide, tetracycline and vancomycin, with a few of them related to other antibiotics (Figure S1 in Supplementary Materials). The proportion of the eight classes in each sample depended on both the sample type and location.
In C. semilaevis samples, ARGs were mostly abundant in A2 (P2), followed by A1 (P1) and A3 (P3). The lowest number of ARGs was found in A0 (P0). The copy number of almost all of the ARGs was higher in the A2 sample too (Figure S1A in Supplementary Materials). In A2, ARGs related to aminoglycosides (26 genes), beta-lactamases (26 genes), MLSB (23 genes), multidrug (20 genes) and tetracycline (14 genes) were the most abundant, among which the aph3ia (aminoglycoside), tetk (Tetracycline), ermK (MLSB) and bl3_cpha (beta-lactamase) genes were the ARGs with the highest copy numbers (Figure S1A in Supplementary Materials). ARGs responsible for resistance to chloramphenicol could not be detected in A1–A3, while ARGs related to aminoglycosides and tetracycline resistance were absent in A0. In general, in type A samples, the abundant ARGs were multidrug, MLSB, beta-lactamases, aminoglycoside and tetracycline-related ones.
In sea water samples, tetracycline, aminoglycosides and multidrug-related ARGs were the most abundant in B1 (P1), while tetracycline, aminoglycosides and MLSB in B2 (P2) (Figure S1B in Supplementary Materials). In B3, ARGs involved in the resistance to beta-lactamase, multidrug and tetracycline were the most abundant (Figure S1B in Supplementary Materials). The number of ARGs was lower in B0 (natural seawater collected from P0) compared to the culture water samples (Figure S1B in Supplementary Materials), among which multidrug, tetracycline and aminoglycosides-related ARGs were dominant. In type B samples, tetracycline, aminoglycoside, multidrug, beta-lactamases and MLSB-related ones were the main ARGs.
ARGs were detected in the soil samples C0-C3, among which multidrug, beta-lactamases, aminoglycoside, MLSB and tetracycline-related ARGs were in the majority (Figure S1C in Supplementary Materials). These ARGs were abundant in C1 (P1) and C2 (P2) compared to other soil samples (Figure S1 in Supplementary Materials).
3.2. Differential Distribution of ARGs between Fish Farms and the Adjacent Natural Sea Area
Based on the statistical analysis with the one-way ANOVA test in SPSS version 26.0, the comparison between samples from the fish farms (P1, P2 and P3) and the natural sea area (P0) indicated that ARGs were highly enriched in the fish farms, with more genes detected (Figure S1, Table S1 in Supplementary Materials). In addition, ARGs with a high abundance were recorded in P2. Some ARGs were detected only in the samples from the natural sea area, such as bacA-02, tetL-01 and ampC-06. Water samples from four locations shared more ARGs in common.
3.3. Mechanisms of ARGs in Different Type of Samples
The mechanisms of ARGs detected in all type of samples were antibiotic deactivation, cellular protection and efflux pump, with little other or unknown mechanisms (Figure S1 in Supplementary Materials). These ARG mechanisms were predominant in the A2 sample, but they were found in lesser extent in the other fish samples. In soil samples, antibiotic deactivation and efflux pump were the most representative mechanisms, especially in C1 and C2.
3.4. Transfer Potential of ARG Types between Fish, Soil and Water Samples
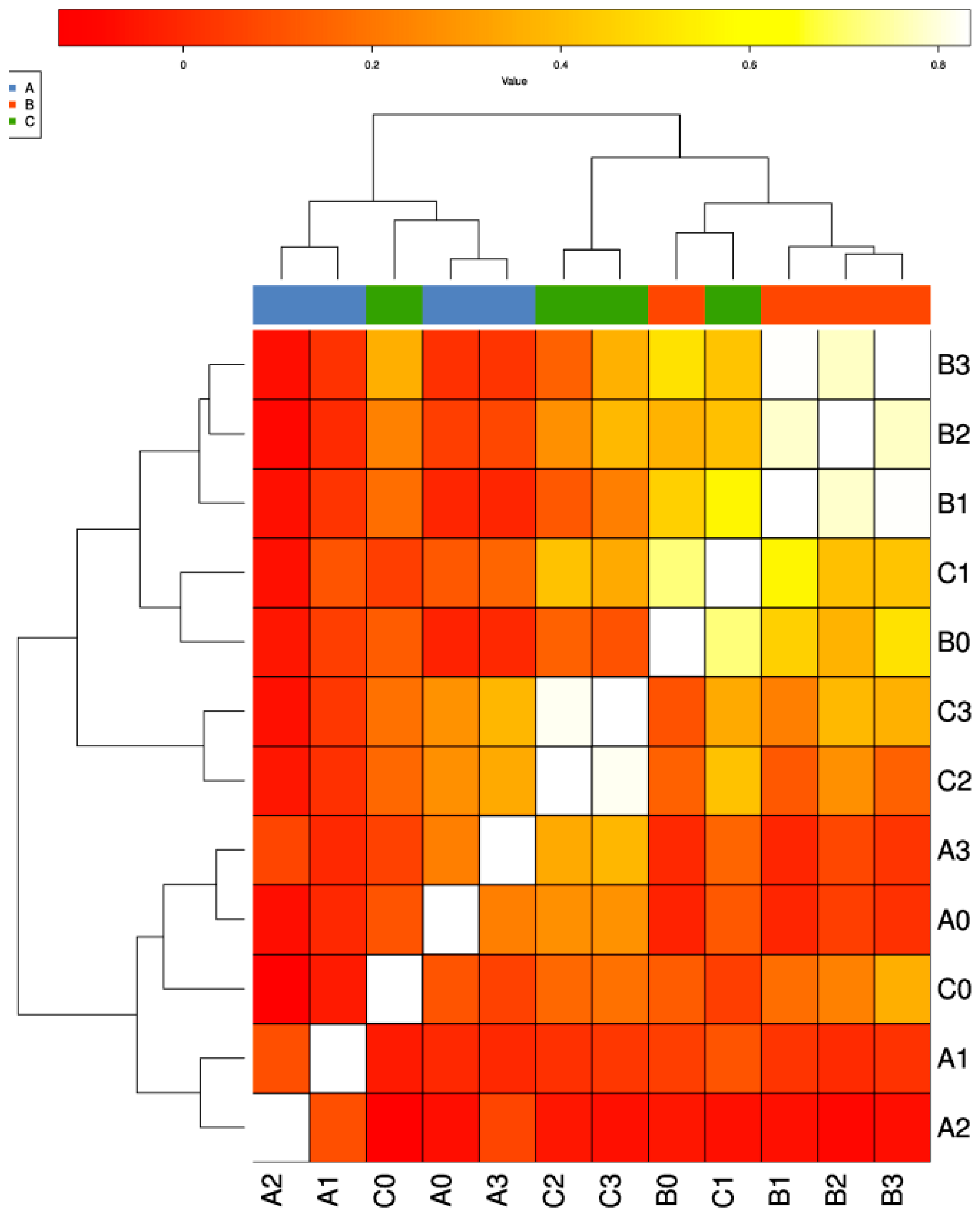
A comparative analysis was performed between samples from the gut of C. semilaevis (A0-A3), water (B0-B3) and soil (C0-C3) to evaluate the possibility of HGT between the different biomes. The correlation of ARGs detected from different samples showed a good cluster in the sample type (Figure 2), and samples from fish (A) were distinguished from water (B) and soil (C) (Adonis: p-value = 0.017; Anosim: p-value = 0.011). The results (Table S1 in Supplementary Materials) indicated that the average level of ARGs detected in fish samples was lower compared to the soil samples and the numbers of ARGs in soil samples were lower compared to the water samples. Relatively pristine environments (A0 and B0) revealed fewer ARGs than other fish farm (Table S1 in Supplementary Materials). Clustered A1 and A2 separated from A0; B1, 2 and 3 separated from B0 (Figure 2). The Venn diagram showing the distribution of ARGs among fish (A), water (B) and soil (C) samples was depicted (Figure 3). A total of 366 ARGs, all classes combined, was screened from the water samples, while 340 ARGs were screened from the soil samples and only 221 ARGs were obtained from the fish samples. Some individual ARGs followed this trend. For example, 62 ARGs encoding for resistances to aminoglycosides were found in water samples, while only 46 and 30 ARGs were recorded for the soil and fish samples, respectively. In addition, 10 ARGs encoding for resistance to chloramphenicol were obtained from water samples, while 6 and 1 from the soil and fish samples, respectively. The same trend was observed for ARGs encoding for tetracycline, as well as mobile genetic elements (MGEs). CIntI-1 (class1), IS613, Tp614, intI-1 (clinic), tnpA01, tnpA02, tnpA03, tnpA04, tnpA05 and tnpA07 were detected, and cIntI-1 (class1), intI-1 (clinic) tnpA04 and tnpA07 were the most common MGEs among the three sample types. Nevertheless, some disparities were observed. ARGs encoding for β-lactam, multidrug and sulfonamide were more abundant in soil samples than the water and fish samples, in this order. Additionally, the MLSB-associated ARGs were uniformly distributed in the water (32), soil (42) and fish (42) samples.
MGEs were in direct proportion with ARGs, and the integrase genes and transposases were in the same trend (Figure S2 in Supplementary Materials). The Spearman correlation analysis among ARGs indicated a significant correlation among all the ARGs across the studied samples (Figure S3 in Supplementary Materials). The correlation coefficient R was > 0.7, and the p-value was < 0.05. The integrase gene intI-1 (clinic) was significantly and positively correlated with ARGs related to multidrug, MLSB and tetracycline resistance., while the other integrase gene cIntI-1 (class1) was significantly and positively correlated with ARGs related to aminoglycosides and tnpA07. Transposases exhibited more correlations with ARGs. IS613 was significantly and positively correlated with vancomycin ARGs, while Tp614 with MLSB, beta-lactamase, aminoglycoside, multidrug and other ARGs were not. TnpA01 was significantly and positively correlated with MLSB, while tnpA02 mainly with tetracycline and beta-lactamase, tnpA03 with MLSB, multidrug, vancomycin and aminoglycosides and tnpA07 were significantly and positively correlated with aminoglycoside ARGs. TnpA04 and tnpA05 interacted more tightly with ARGs related to tetracycline and beta-lactamase.
4. Discussion
Studies have shown that approximately 30% to 90% of used antibiotics promote the production and spread of ARGs [23], but the abundance and dissemination of ARGs in the environment and their mechanisms are far from being completely elucidated. The present study aimed to explore the distribution of ARGs in aquatic microecosystems. Eight classes of ARGs, namely, aminoglycoside, multidrug, beta-lactamase, MLSB, chloramphenicol, sulfonamide, tetracycline and vancomycin ARGs, which are all common antibiotics, were those predominantly abundant in the aquatic environment. The abundance of these ARGs differed according to the microsystem and sampling site, mostly abundant in the soils, indicating an accumulation in the sediments. Regardless of the type of samples, the mechanisms adopted by the ARGs were the same and were represented by antibiotic deactivation, cellular protection and efflux pump. These results were the first look into the distribution and spread of ARGs related to the C. semilaevis breeding industry.
The results of the high-throughput analysis of the resistome revealed that a high level of ARGs could be found in the soil, water and intestinal tract of C. semilaevis. These results signposted that fish farms constitute a great reservoir of ARGs, conforming with previous research demonstrating that fish-rearing ponds, lakes, bays [24,25,26] and shrimp ponds [16,17] were all enriched in ARGs. In addition, the bacterial community structures also differed, which might affect the composition of ARGs [27,28,29]. The aminoglycoside-related ARGs were more abundant in the water samples, and this was corroborated by previous findings indicating that aminoglycoside ARGs are abundant and persistent in wastewater plants [9] and drinking waters [30]. The presence of aminoglycoside ARGs in the fish intestinal tract and soil samples was also supported by studies conveying the persistence of aminoglycoside ARGs in the gut of fish and zoo animals [31] and plenty of studies reporting the presence of aminoglycoside ARGs in the soils of diverse types [6,12,13,26,32]. Our findings also indicated that β-lactam ARGs were enriched in soil and water samples, but were found in a lesser extent in the fish intestinal tract. This observation is supported by previous works in the Tibetan environment, sewage sludge field application soils, agricultural soils and parks [9,30,33,34,35,36,37]. The abundance of chloramphenicol ARGs was relatively low in our samples, but its occurrence was in line with previous works which indicated the presence of chloramphenicol ARGs in feedlots, Baltic sea fish farm soils, water effluents and pristine remote Antarctic soils [6,8,32,38]. MLSB-related ARGs were uniformly distributed in the fish intestinal tract, water and soil samples. Studies reporting MLSB ARGs in fish intestinal tract are limited, but these ARGs have been reported in water effluents and soils treated with sewage sludge [8,26]. A particular enrichment of multidrug ARGs was recorded in the soil samples, followed by water and fish intestinal tract samples. These ARGs have been also reported in soil and water samples by other researchers [6,16,26,30,36,39]. Similarly, sulfonamide, tetracycline and vancomycin-resistant ARGs were all found in the three types of samples, especially tetracycline, which corroborated previous findings [6,8,30,40,41]. Our study revealed a diversity of ARGs with different degrees of abundance in water, the intestinal tract of C. semilaevis and soil in the aquatic settings and adjacent natural sea areas.
Disparities were found in ARGs detected from the same type of samples collected from diverse geographical locations. The level of ARGs was lower in the samples from natural sources. In daily production, antibiotics are usually applied through the splashing of the factory culture ponds in the fish farm, mainly florfenicol, quinolones (oxoquine), streptomycin, neomycin (neomycin sulfate and tetracycline), doxycycline hydrochloride, sulfonamide antibiotics (sulfadiazine and sulfamethoxazole) and enrofloxacin, related to the ARGs detected here. A frequent outside release of the culture water may decrease the concentration of antibiotics and ARB in it, which could explain the decreased level of ARGs in the water samples. The discharge of effluent water in the soil may also explain the increased accumulation of ARGs in soil samples. Indeed, previous studies indicated that the effluent water could be released into the natural world, infiltrating into the ground or being dumped into the sea, which leads to the accumulation of antibiotic-resistant bacteria in the soil; thus, causing the enrichment of soil in ARGs, which is equivalent to ARG transfer and enrichment from water to soil [29]. From the geographical map of our sampling, the three fish farms were all near the sea and the water for fish farming came from the sea, a large part of which would flow back in. Therefore, the transfer of ARGs may occur from the coastal fish farm to the sea (including natural seawater, sea mud and marine organisms such as fish species). Accordingly, inflow water was also proved to be a major source of trout farming contamination with salmonella and multidrug-resistant bacteria [27]. Thus, a cycle accumulation pattern seems to exist.
MGEs involved in the mobility of ARGs are an important factor [33,36,42,43] and were detected in all three types of samples, such as intI-1 (clinic) tnpA04 and tnpA07. Wild fish in a peri-urban river might be recipients and communicators of ARGs in the water environment and ARGs might transfer between fish and water using bacteria as a spreading medium, with nine ARGs and one MGE continuously shared in both sample types [44]. Notably, a differentially ecological niche of the host resulted in the various resistomes of the lower vertebrate, among which the resistomes of seawater fish shared more similarities and were characterized by a high abundance, distinct from other observed habitats, and ARGs in seawater fish were colocalized with mobile elements with a high frequency, suggesting that they were likely spread through HGT [45]. On the whole, these data revealed the significant diversity and heterogeneity of ARGs in lower vertebrates, indicating that these wild species potentially play an essential role in the global spread of ARGs [44,45].
The HGT of ARGs among environmental bacteria in different taxa is proved to be an important pathway that disseminates resistance and, subsequently, acquires resistance through human pathogens and commensals. Since transposition-related genes and integrases were found in the studied samples, HGT might occur between bacteria by transposition or integration in the aquatic environment. This threes in corroboration with previous reports, confirming that ARGs are introduced by DNA transposition into plasmid and subsequent drug selection in the aquaculture sites [46]. Further studies should be carried out to verify these HGT mechanisms.
In this study, mechanisms driven by ARGs were mainly antibiotic deactivation, cellular protection and efflux pump. Efflux pump is the major mechanism of multidrug resistance in bacteria and acts by interfering with the synthesis of nucleic acid or the inhibition of protein synthesis [6,13,17,47]. Antibiotic deactivation has also been reported in previous studies on urban park soils with reclaimed water irrigation [2,13]. Cellular protection has been proved as a mechanism driven by ARGs screened from farmed fish feces [6], which corroborated our results. Antibiotic deactivation was the most abundant ARG mechanism in the fish intestinal extract, water and soil samples, which indicated that the deactivation of antibiotics may be the main mechanism adopted by resistant bacteria in the aquatic environment. The three mechanisms were also reported by Muziasari [6]. Thus, finding ways to counteract this mechanism could be the key to overcome antibiotic resistance, especially for animals left in these environments.
This study investigated the distributions, diversity, abundance and potential transfer mechanism of ARGs in C. semilaevis-related samples; however, these results still need to be verified by a larger-scale sample test. This work also outlined the crisis of ARG transmission from fish farms to adjacent sea areas.
5. Conclusions
Our study indicated that there were abundant ARGs in the aquatic ecosystem and that there was a transfer of these ARGs into the water, which, subsequently, led to the serious enrichment of aquatic sediments in ARGs. This study allowed not only the knowledge of ARGs in the C. semilaevis-related niche, but also the understanding of ARG dissemination in aquatic ecosystems. Therefore, the investigation and in-depth study of the distribution and spread of ARGs could allow us to control the spread of ARGs in the environment and minimize the risk to animal and human health.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w14060938/s1. Figure S1: distribution and abundance of detected ARGs, Figure S2: correlation between trends of ARGs and MEGs in three types of samples, Figure S3: correlations among the detected ARGs and MEGs, Table S1: Abundance and distribution of ARG types in different samples. Table S2: The information of primers used in the study.
Author Contributions
Conceptualization, B.Z.; methodology, H.L.; validation, Q.D. and H.L.; formal analysis, L.J.; investigation, N.Z.; data curation, N.Z.; writing—review and editing, L.J.; project administration, C.Z.; funding acquisition, L.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from the Tianjin Natural Science Foundation (17JCQNJC15000), China Agriculture Research System of MOF and MARA (CARS-47-Z01) and the modern industrial technology system in Tianjin ITTMRS2022001.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board of Tianjin Fisheries Research Institute (protocol code 2019003 and date of approval is 2 April 2019).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors would like to acknowledge the support of the client project team and their collaborative effort with the wider project team, which included: the Tianjin Fisheries Research Institute; Guangdong Ocean University; Southern Marine Science and Engineering Guangdong Laboratory-Zhanjiang.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lin, H.; Zhang, J.; Chen, H.; Wang, J.; Sun, W.; Zhang, X.; Yang, Y.; Wang, Q.; Ma, J. Effect of temperature on sulfonamide antibiotics degradation, and on antibiotic resistance determinants and hosts in animal manures. Sci. Total Environ. 2017, 607–608, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Pontes, D.S.; de Araujo, R.S.A.; Dantas, N.; Scotti, L.; Scotti, M.T.; de Moura, R.O.; Mendonca-Junior, F.J.B. Genetic mechanisms of antibiotic resistance and the role of antibiotic adjuvants. Curr. Top. Med. Chem. 2018, 18, 42–74. [Google Scholar] [CrossRef] [PubMed]
- Loftie-Eaton, W.; Crabtree, A.; Perry, D.; Millstein, J.; Baytosh, J.; Stalder, T.; Robison, B.D.; Forney, L.J.; Top, E.M. Contagious Antibiotic Resistance: Plasmid Transfer among Bacterial Residents of the Zebrafish Gut. Appl. Environ. Microbiol. 2021, 87, e02735-20. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.C.; Feng, W.Q.; Han, Y.; Zheng, J.; Chen, T.; Wei, Y.Y.; Gillings, M.; Zhu, Y.G.; Chen, H. Prevalence and transmission of antibiotic resistance and microbiota between humans and water environments. Environ. Int. 2018, 121, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Brede, D.A.; Diep, D.B.; Nes, I.F.; Lotfipour, F.; Hojabri, Z. Efficient inactivation of multi-antibiotics resistant nosocomial enterococci by purified Hiracin bacteriocin. Adv. Pharm. Bull. 2015, 5, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Muziasari, W.I.; Pitkänen, L.K.; Sørum, H.; Stedtfeld, R.D.; Tiedje, J.M.; Virta, M. The resistome of farmed fish feces contributes to the enrichment of antibiotic resistance genes in sediments below Baltic Sea fish farms. Front. Microbiol. 2016, 7, 2137. [Google Scholar]
- Calero-Cáceres, W.; Méndez, J.; Martín-Díaz, J.; Muniesa, M. The occurrence of antibiotic resistance genes in a Mediterranean river and their persistence in the riverbed sediment. Environ. Pollut. 2017, 223, 384–394. [Google Scholar] [CrossRef]
- Szekeres, E.; Baricz, A.; Chiriac, C.M.; Farkas, A.; Opris, O.; Soran, M.L.; Andrei, A.S.; Rudi, K.; Balcázar, J.L.; Dragos, N.; et al. Abundance of antibiotics, antibiotic resistance genes and bacterial community composition in wastewater effluents from different Romanian hospitals. Environ. Pollut. 2017, 225, 304–315. [Google Scholar] [CrossRef]
- An, X.L.; Su, J.Q.; Li, B.; Ouyang, W.Y.; Zhao, Y.; Chen, Q.L.; Cui, L.; Chen, H.; Gillings, M.R.; Zhang, T.; et al. Tracking antibiotic resistome during wastewater treatment using high throughput quantitative PCR. Environ. Int. 2018, 117, 146–153. [Google Scholar] [CrossRef]
- Lerminiaux, N.A.; Cameron, A.D.S. Horizontal transfer of antibiotic resistance genes in clinical environments. Can. J. Microbiol. 2019, 65, 34–44. [Google Scholar] [CrossRef]
- Bouki, C.; Venieri, D.; Diamadopoulos, E. Detection and fate of antibiotic resistant bacteria in wastewater treatment plants: A review. Ecotoxicol. Environ. Saf. 2013, 91, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Jia, S.; He, X.; Zhang, X.; Ye, L. Different impacts of manure and chemical fertilizers on bacterial community structure and antibiotic resistance genes in arable soils. Chemosphere 2017, 188, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.H.; Qiao, M.; Su, J.Q.; Chen, Z.; Zhou, X.; Zhu, Y.G. High throughput profiling of antibiotic resistance genes in urban park soils with reclaimed water irrigation. Environ. Sci. Technol. 2014, 48, 9079–9085. [Google Scholar] [CrossRef] [PubMed]
- Larrañaga, O.; Brown-Jaque, M.; Quirós, P.; Gómez-Gómez, C.; Blanch, A.R.; Rodríguez-Rubio, L.; Muniesa, M. Phage particles harboring antibiotic resistance genes in fresh-cut vegetables and agricultural soil. Environ. Int. 2018, 115, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Muurinen, J.; Stedtfeld, R.; Karkman, A.; Pärnänen, K.; Tiedje, J.; Virta, M. Influence of manure application on the environmental resistome under finnish agricultural practice with restricted antibiotic use. Environ. Sci. Technol. 2017, 51, 5989–5999. [Google Scholar] [CrossRef]
- Pham, T.T.H.; Rossi, P.; Dinh, H.D.K.; Pham, N.T.A.; Tran, P.A.; Ho, T.; Dinh, Q.T.; De Alencastro, L.F. Analysis of antibiotic multi-resistant bacteria and resistance genes in the effluent of an intensive shrimp farm (Long An, Vietnam). J. Environ. Manag. 2018, 214, 149–156. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, X.X.; Zhao, Z.; Duan, C.; Chen, H.; Wang, M.; Ren, H.; Yin, Y.; Ye, L. Metagenomic analysis revealed the prevalence of antibiotic resistance genes in the gut and living environment of freshwater shrimp. J. Hazard Mater. 2018, 350, 10–18. [Google Scholar] [CrossRef]
- Almeida, A.R.; Tacão, M.; Soares, J.; Domingues, I.; Henriques, I. Tetracycline-resistant bacteria selected from water and zebrafish after antibiotic exposure. Int. J. Environ. Res. Public Health 2021, 18, 3218. [Google Scholar] [CrossRef]
- Fu, J.; Yang, D.; Jin, M.; Liu, W.; Zhao, X.; Li, C.; Zhao, T.; Wang, J.; Gao, Z.; Shen, Z.; et al. Aquatic animals promote antibiotic resistance gene dissemination in water via conjugation: Role of different regions within the zebra fish intestinal tract, and impact on fish intestinal microbiota. Mol. Ecol. 2017, 26, 5318–5333. [Google Scholar] [CrossRef]
- Wang, L.; Su, H.; Hu, X.; Xu, Y.; Xu, W.; Huang, X.; Li, Z.; Cao, Y.; Wen, G. Abundance and removal of antibiotic resistance genes (ARGs) in the rearing environments of intensive shrimp aquaculture in South China. J. Environ. Sci. Health B 2019, 54, 211–218. [Google Scholar] [CrossRef]
- Cycoń, M.; Mrozik, A.; Piotrowska-Seget, Z. Antibiotics in the soil environment-degradation and their impact on microbial activity and diversity. Front. Microbiol. 2019, 10, 338. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Zhao, Y.; Li, B.; Huang, C.L.; Zhang, S.Y.; Yu, S.; Chen, Y.S.; Zhang, T.; Gillings, M.R.; Su, J.Q. Continental-scale pollution of estuaries with antibiotic resistance genes. Nat. Microbiol. 2017, 2, 16270. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Cui, Y.; Li, A.; Zou, X.; Ma, C.; Chen, Z. Antibiotics and antibiotic resistance genes from wastewater treated in constructed wetlands. Ecol. Eng. 2022, 177, 106548. [Google Scholar] [CrossRef]
- Calero-Cáceres, W.; Melgarejo, A.; Colomer-Lluch, M.; Stoll, C.; Lucena, F.; Jofre, J.; Muniesa, M. Sludge as a potential important source of antibiotic resistance genes in both the bacterial and bacteriophage fractions. Environ. Sci. Technol. 2014, 48, 7602–7611. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewska, E.; Harnisz, M. Relationship between modification of activated sludge wastewater treatment and changes in antibiotic resistance of bacteria. Sci. Total Environ. 2018, 639, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.Y.; McGrath, S.P.; Su, J.Q.; Hirsch, P.R.; Clark, I.M.; Shen, Q.; Zhu, Y.G.; Zhao, F.J. Long-term impact of field applications of sewage sludge on soil antibiotic resistome. Environ. Sci. Technol. 2016, 50, 12602–12611. [Google Scholar] [CrossRef]
- Antunes, P.; Campos, J.; Mourão, J.; Pereira, J.; Novais, C.; Peixe, L. Inflow water is a major source of trout farming contamination with Salmonella and multidrug resistant bacteria. Sci. Total Environ. 2018, 642, 1163–1171. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Zhang, L.; Wang, X.; Lu, C.; Liu, J.; Liu, Y.; Li, L.; Peng, J.; Xue, M. High-throughput analysis of the effects of different fish culture methods on antibiotic resistance gene abundances in a lake. Environ. Sci. Pollut. Res. Int. 2019, 26, 5445–5453. [Google Scholar] [CrossRef]
- Yuan, J.; Ni, M.; Liu, M.; Zheng, Y.; Gu, Z. Occurrence of antibiotics and antibiotic resistance genes in a typical estuary aquaculture region of Hangzhou Bay, China. Mar. Pollut. Bull. 2019, 138, 376–384. [Google Scholar] [CrossRef]
- Ma, L.; Li, B.; Jiang, X.T.; Wang, Y.L.; Xia, Y.; Li, A.D.; Zhang, T. Catalogue of antibiotic resistome and host-tracking in drinking water deciphered by a large scale survey. Microbiome 2017, 5, 154. [Google Scholar] [CrossRef] [Green Version]
- Bello González Tde, J.; Zuidema, T.; Bor, G.; Smidt, H.; van Passel, M.W. Study of the aminoglycoside subsistence phenotype of bacteria residing in the gut of humans and zoo animals. Front. Microbiol. 2015, 6, 1550. [Google Scholar] [CrossRef] [PubMed]
- Van Goethem, M.W.; Pierneef, R.; Bezuidt, O.K.I.; Van De Peer, Y.; Cowan, D.A.; Makhalanyane, T.P. A reservoir of ’historical’ antibiotic resistance genes in remote pristine Antarctic soils. Microbiome 2018, 6, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Yuan, K.; Chen, X.; Yang, Y.; Zhang, T.; Wang, Y.; Luan, T.; Zou, S.; Li, X. Metagenomic analysis revealing antibiotic resistance genes (ARGs) and their genetic compartments in the Tibetan environment. Environ. Sci. Technol. 2016, 50, 6670–6679. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; An, X.; Li, H.; Su, J.; Ma, Y.; Zhu, Y.G. Long-term field application of sewage sludge increases the abundance of antibiotic resistance genes in soil. Environ. Int. 2016, 92–93, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.H.; Tang, X.Y.; Cui, J.F. Effect of long-term manure slurry application on the occurrence of antibiotic resistance genes in arable purple soil (entisol). Sci. Total Environ. 2019, 647, 853–861. [Google Scholar] [CrossRef]
- Gou, M.; Hu, H.W.; Zhang, Y.J.; Wang, J.T.; Hayden, H.; Tang, Y.Q.; He, J.Z. Aerobic composting reduces antibiotic resistance genes in cattle manure and the resistome dissemination in agricultural soils. Sci. Total Environ. 2018, 612, 1300–1310. [Google Scholar] [CrossRef]
- Han, X.M.; Hu, H.W.; Shi, X.Z.; Wang, J.T.; Han, L.L.; Chen, D.; He, J.Z. Impacts of reclaimed water irrigation on soil antibiotic resistome in urban parks of Victoria, Australia. Environ. Pollut. 2016, 211, 48–57. [Google Scholar] [CrossRef]
- He, L.Y.; Liu, Y.S.; Su, H.C.; Zhao, J.L.; Liu, S.S.; Chen, J.; Liu, W.R.; Ying, G.G. Dissemination of antibiotic resistance genes in representative broiler feedlots environments: Identification of indicator ARGs and correlations with environmental variables. Environ. Sci. Technol. 2014, 48, 13120–13129. [Google Scholar] [CrossRef]
- Kang, W.; Zhang, Y.J.; Shi, X.; He, J.Z.; Hu, H.W. Short-term copper exposure as a selection pressure for antibiotic resistance and metal resistance in an agricultural soil. Environ. Sci. Pollut. Res. Int. 2018, 25, 29314–29324. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, J.; Zhu, L.; Ge, W.; Wang, J. Environmental analysis of typical antibiotic-resistant bacteria and ARGs in farmland soil chronically fertilized with chicken manure. Sci. Total Environ. 2017, 593–594, 10–17. [Google Scholar] [CrossRef]
- Lin, H.; Chapman, S.J.; Freitag, T.E.; Kyle, C.; Ma, J.; Yang, Y.; Zhang, Z. Fate of tetracycline and sulfonamide resistance genes in a grassland soil amended with different organic fertilizers. Ecotoxicol. Environ. Saf. 2019, 170, 39–46. [Google Scholar] [CrossRef]
- Chen, Q.L.; An, X.L.; Zhu, Y.G.; Su, J.Q.; Gillings, M.R.; Ye, Z.L.; Cui, L. Application of struvite alters the antibiotic resistome in soil, rhizosphere, and phyllosphere. Environ. Sci. Technol. 2017, 51, 8149–8157. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.; Chen, Q.L.; Zhu, D.; An, X.L.; Yang, X.R.; Su, J.Q.; Qiao, M.; Zhu, Y.G. Spatial and temporal distribution of antibiotic resistomes in a peri-urban area is associated significantly with anthropogenic activities. Environ. Pollut. 2018, 235, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.C.; Lin, Z.J.; Shuai, X.Y.; Zheng, J.; Meng, L.X.; Zhu, L.; Sun, Y.J.; Shang, W.C.; Chen, H. Temporal variation and sharing of antibiotic resistance genes between water and wild fish gut in a peri-urban river. J. Environ. Sci. 2021, 103, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.M.; Holmes, E.C.; Chen, X.; Tian, J.H.; Lin, X.D.; Qin, X.C.; Gao, W.H.; Liu, J.; Wu, Z.D.; Zhang, Y.Z. Diverse and abundant resistome in terrestrial and aquatic vertebrates revealed by transcriptional analysis. Sci. Rep. 2020, 10, 18870. [Google Scholar] [CrossRef]
- Nonaka, L.; Maruyama, F.; Onishi, Y.; Kobayashi, T.; Ogura, Y.; Hayashi, T.; Suzuki, S.; Masuda, M. Various pAQU plasmids possibly contribute to disseminate tetracycline resistance gene tet(M) among marine bacterial community. Front. Microbiol. 2014, 5, 152. [Google Scholar] [CrossRef]
- Chen, B.; He, R.; Yuan, K.; Chen, E.; Lin, L.; Chen, X.; Sha, S.; Zhong, J.; Lin, L.; Yang, L.; et al. Polycyclic aromatic hydrocarbons (PAHs) enriching antibiotic resistance genes (ARGs) in the soils. Environ. Pollut. 2017, 220, 1005–1013. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Map of the sampling location and sampling sites: The four sampling sites were P0 (117.935° E, 39.0415° N), P1 (117.626544° E, 38.852927° N), P2 (117.912634° E, 39.304451° N) and P3 (117.964958° E, 39.264878° N) in the west of the Bohai Sea, in northeast China.
Figure 1.
Map of the sampling location and sampling sites: The four sampling sites were P0 (117.935° E, 39.0415° N), P1 (117.626544° E, 38.852927° N), P2 (117.912634° E, 39.304451° N) and P3 (117.964958° E, 39.264878° N) in the west of the Bohai Sea, in northeast China.

Figure 2.
Correlation of ARGs between different samples: detected correlation of ARGs between different samples from the gut of C. semilaevis (A0–A3), water samples (B0–B3) and soil samples (C0–C3) to evaluate the possibility of HGT between the different biomes.
Figure 2.
Correlation of ARGs between different samples: detected correlation of ARGs between different samples from the gut of C. semilaevis (A0–A3), water samples (B0–B3) and soil samples (C0–C3) to evaluate the possibility of HGT between the different biomes.

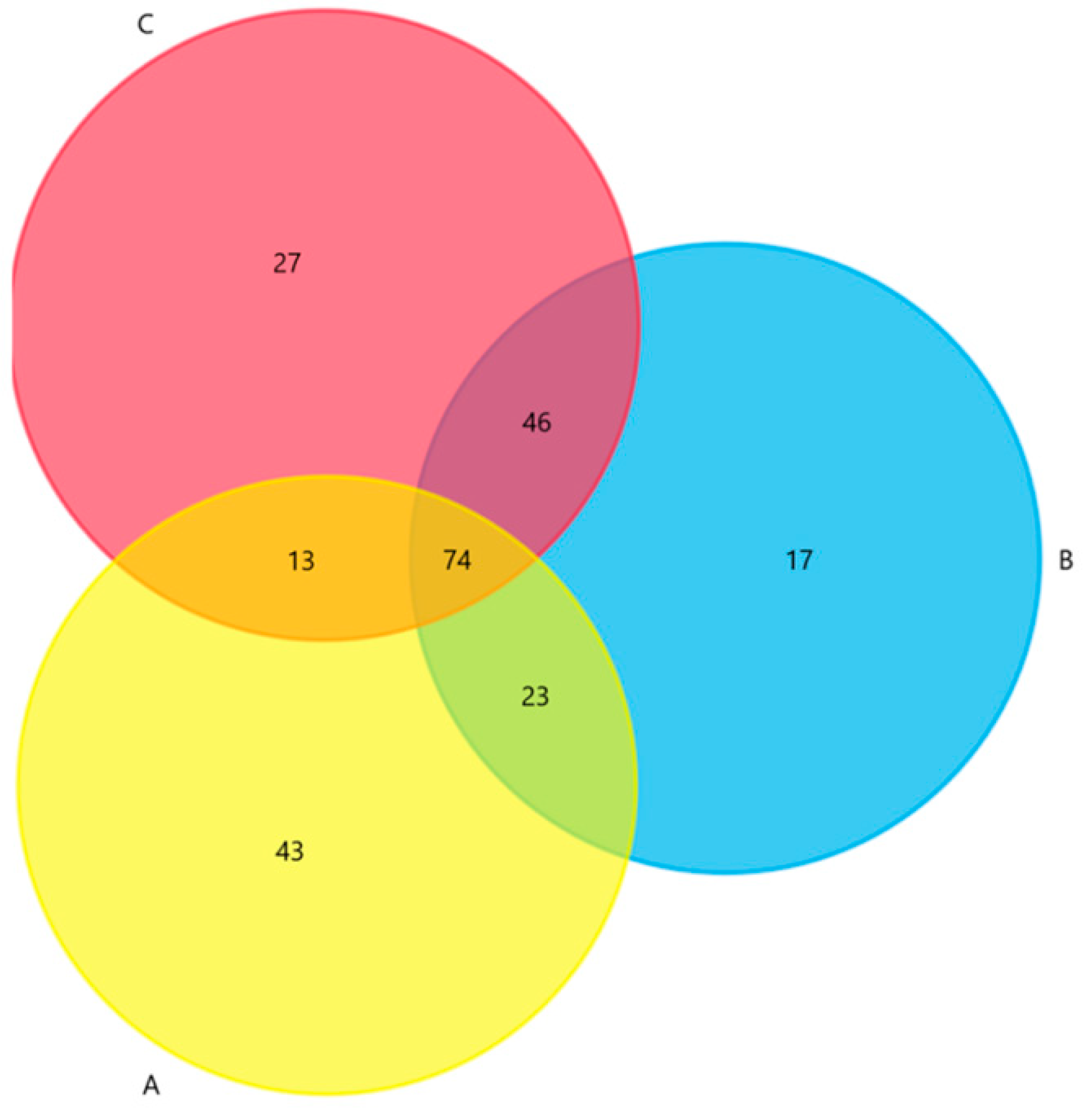
Figure 3.
Venn diagram indicating the abundance of shared and dissimilar ARGs in the three types of samples: Three types of samples were fish samples (A), water samples (B) and soil samples (C). There were 366 ARGs in total, 74 of them shared in the three types of samples, 23 of them shared between fish samples and water samples, 46 of them shared between water samples and soil samples and 13 of them shared between fish samples and soil samples.
Figure 3.
Venn diagram indicating the abundance of shared and dissimilar ARGs in the three types of samples: Three types of samples were fish samples (A), water samples (B) and soil samples (C). There were 366 ARGs in total, 74 of them shared in the three types of samples, 23 of them shared between fish samples and water samples, 46 of them shared between water samples and soil samples and 13 of them shared between fish samples and soil samples.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Samples from different sites.
| Site | Type | Sample | ||
|---|---|---|---|---|
| Intestinal Tracts of Fish | Sea Water | Soil | ||
| P0 | Natural sea area | A0 | B0 | C0 |
| P1 | Fish farm | A1 | B1 | C1 |
| P2 | Fish farm | A2 | B2 | C2 |
| P3 | Fish farm | A3 | B3 | C3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jia, L.; Liu, H.; Zhao, N.; Deng, Q.; Zhu, C.; Zhang, B. Distribution and Transfer of Antibiotic Resistance Genes in Coastal Aquatic Ecosystems of Bohai Bay. Water 2022, 14, 938. https://doi.org/10.3390/w14060938
AMA Style
Jia L, Liu H, Zhao N, Deng Q, Zhu C, Zhang B. Distribution and Transfer of Antibiotic Resistance Genes in Coastal Aquatic Ecosystems of Bohai Bay. Water. 2022; 14(6):938. https://doi.org/10.3390/w14060938
Chicago/Turabian StyleJia, Lei, Hao Liu, Na Zhao, Qiuxia Deng, Chunhua Zhu, and Bo Zhang. 2022. "Distribution and Transfer of Antibiotic Resistance Genes in Coastal Aquatic Ecosystems of Bohai Bay" Water 14, no. 6: 938. https://doi.org/10.3390/w14060938
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.