
Variation in the Structure and Composition of Bacterial Communities within Drinking Water Fountains in Melbourne, Australia

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Processing of Biofilm and Water Samples
2.2.1. Anion and Cation Determination
2.2.2. Coliform/Escherichia coli
2.2.3. Viable Counts
2.2.4. Total Cell Counts
2.2.5. DNA Extraction, PCR Amplification and Sequencing
2.3. Statistical Analysis
3. Results
3.1. Chemical and Microbiological Characteristics of Fountain Drinking Water
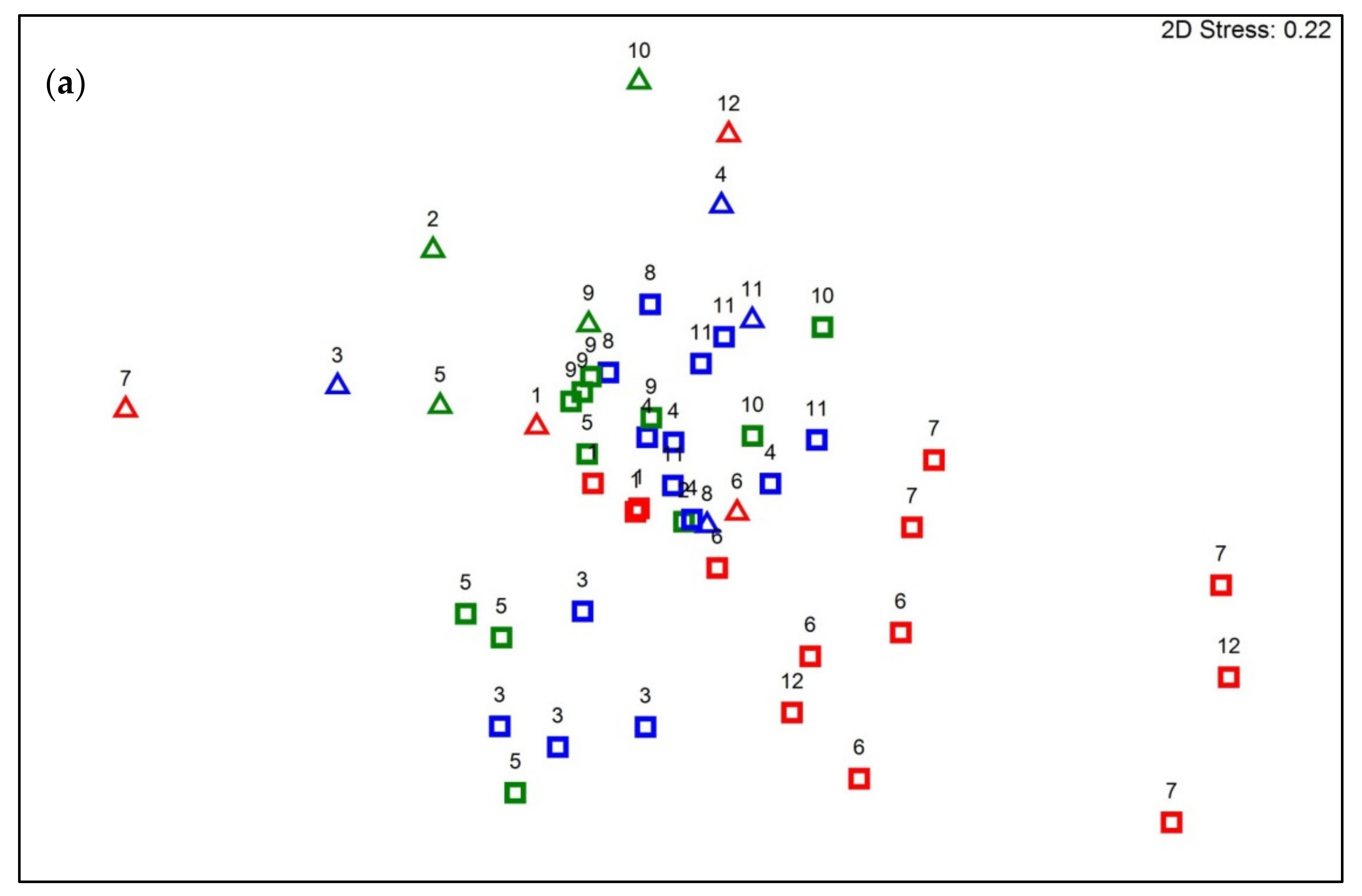
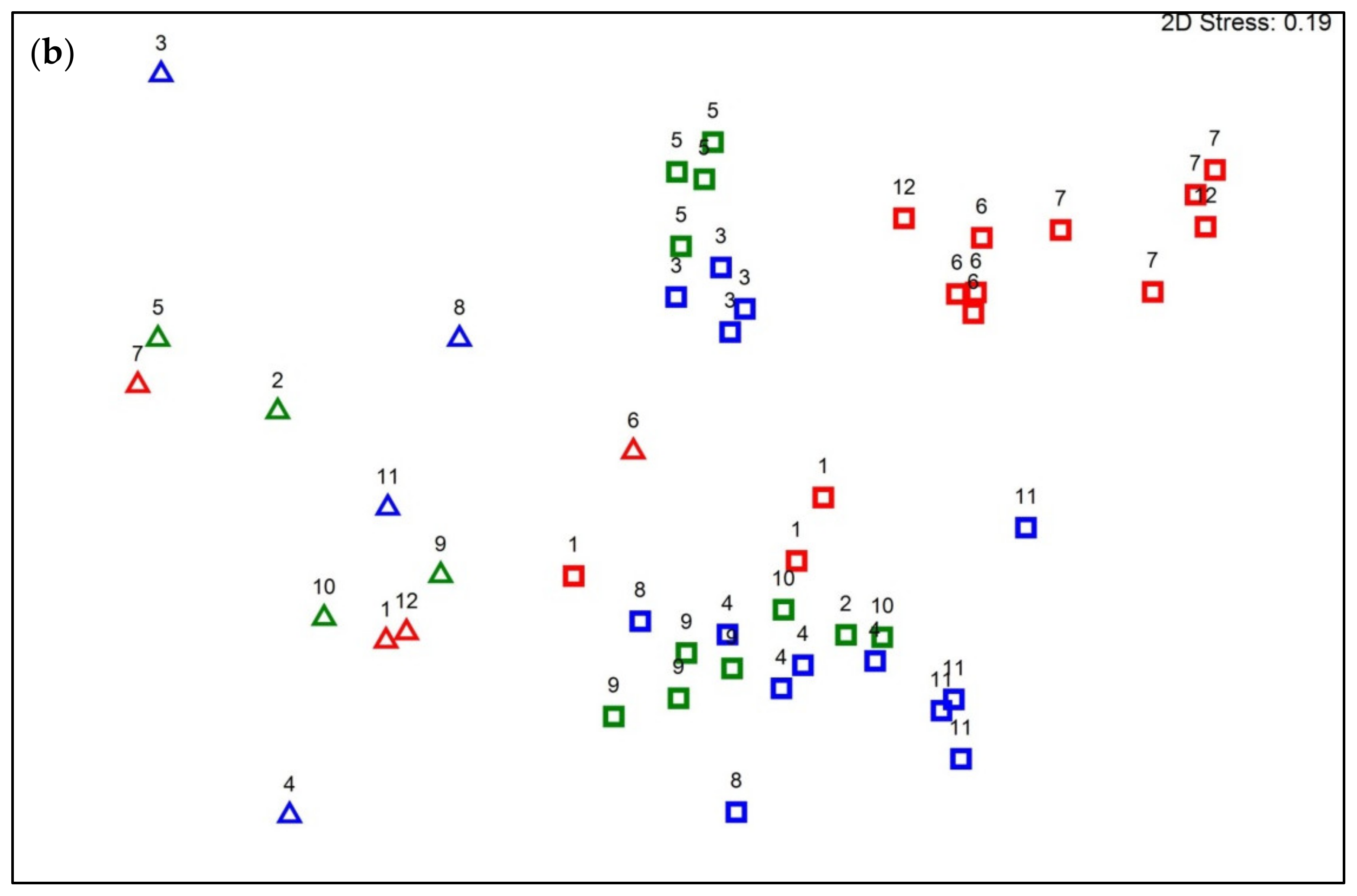
3.2. Diversity and Structure of Fountain Communities in Water and Biofilm
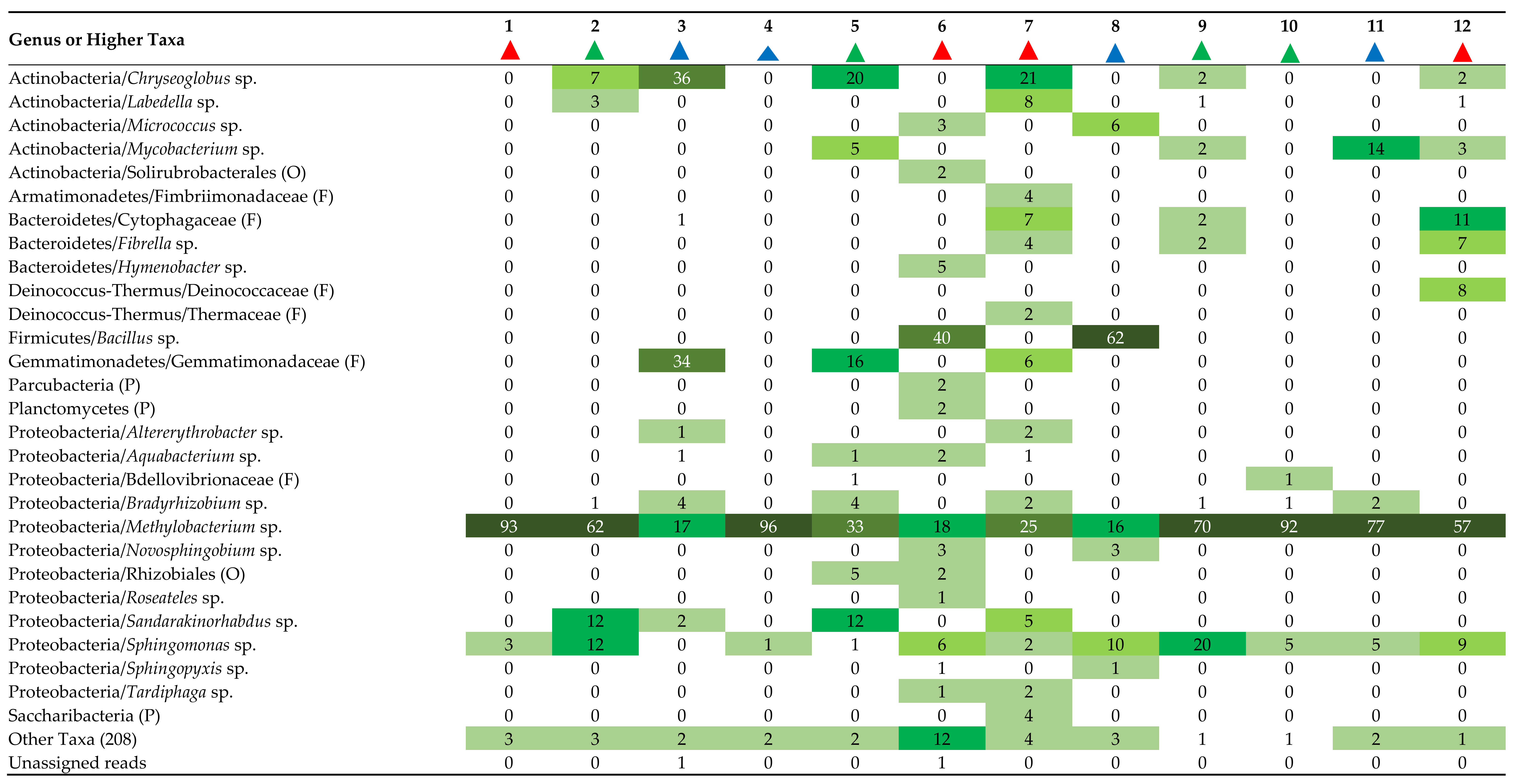
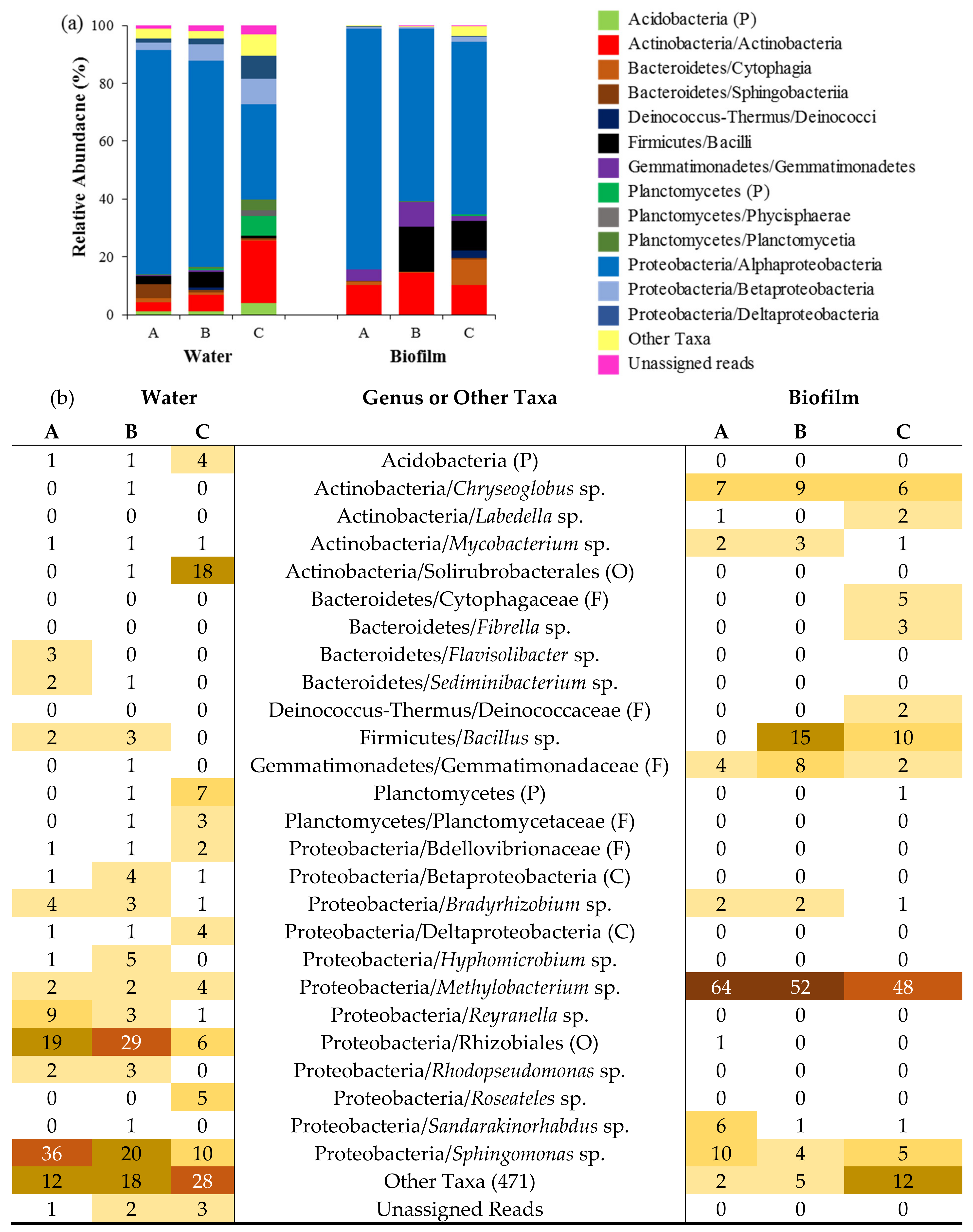
3.3. Composition of Fountain Drinking Water and Biofilm Communities
3.4. Variation in Bacterial Community Composition between Different Fountain Types
4. Discussion
4.1. Diversity and Structure of Fountain Communities in Water and Biofilm
4.2. Composition of Fountain Drinking Water and Biofilm Communities
4.3. Variation in Bacterial Community Composition between Different Fountain Types
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Husband, P.S.; Boxall, J.B. Asset deterioration and discolouration in water distribution systems. Water Res. 2011, 45, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.J.; Schroeder, J.; Lunn, M.; Sloan, W.; Raskin, L. Spatial-Temporal Survey and Occupancy-Abundance Modeling to Predict Bacterial Community Dynamics in the Drinking Water. Microbiome 2014, 5, e01135-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potgieter, S.; Pinto, A.; Sigudu, M.; du Preez, H.; Ncube, E.; Venter, S. Long-term spatial and temporal microbial community dynamics in a large-scale drinking water distribution system with multiple disinfectant regimes. Water Res. 2018, 139, 406–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douterelo, I.; Sharpe, R.L.; Boxall, J.B. Influence of hydraulic regimes on bacterial community structure and composition in an experimental drinking water distribution system. Water Res. 2013, 47, 503–516. [Google Scholar] [CrossRef]
- Sekar, R.; Deines, P.; Machell, J.; Osborn, A.M.; Biggs, C.A.; Boxall, J.B. Bacterial water quality and network hydraulic characteristics: A field study of a small, looped water distribution system using culture-independent molecular methods. J. Appl. Microbiol. 2012, 112, 1220–1234. [Google Scholar] [CrossRef]
- Liu, R.; Zhu, J.; Yu, Z.; Joshi, D.; Zhang, H.; Lin, W.; Yang, M. Molecular analysis of long-term biofilm formation on PVC and cast iron surfaces in drinking water distribution system. J. Environ. Sci. 2014, 26, 865–874. [Google Scholar] [CrossRef]
- Ren, H.; Wang, W.; Liu, Y.; Liu, S.; Lou, L.; Cheng, D.; He, X.; Zhou, S.; Fu, L.; Liu, J.; et al. Pyrosequencing analysis of bacterial communities in biofilms from different pipe materials in a city drinking water distribution system of East China. Appl. Microbiol. Biotechnol. 2015, 99, 10713–10724. [Google Scholar] [CrossRef]
- Asghari, F.B.; Jaafari, J.; Yousefi, M.; Mohammadi, A.A.; Dehghanzadeh, R. Evaluation of water corrosion, scaling extent and heterotrophic plate count bacteria in asbestos and polyethylene pipes in drinking water distribution system. Hum. Ecol. Risk Assess. Int. J. 2018, 24, 1138–1149. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Percival, S.L.; Walker, J.T. Contamination potential of biofilms in water distribution systems. Water Sci. Technol. Water Supply 2002, 2, 271–280. [Google Scholar] [CrossRef]
- Wingender, J.; Neu, T.; Flemming, H.-C. (Eds.) What are Bacterial extracellular polymer substances? In Bacterial Extracellular Polymer Substances; Springer: Berlin/Heidelberg, Germany, 1999; pp. 1–19. [Google Scholar]
- Fish, K.E.; Osborn, A.M.; Boxall, J.B. Biofilm structures (EPS and bacterial communities) in drinking water distribution systems are conditioned by hydraulics and influence discolouration. Sci. Total Environ. 2017, 593–594, 571–580. [Google Scholar] [CrossRef]
- Douterelo, I.; Jackson, M.; Solomon, C.; Boxall, J. Spatial and temporal analogies in microbial communities in natural drinking water biofilms. Sci. Total Environ. 2017, 581–582, 277–288. [Google Scholar] [CrossRef]
- Davies, D.G. Biofilm Dispersion. In Biofilm Highlights; Flemming, H.-C., Wingender, J., Szewzyk, U., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–28. [Google Scholar]
- Homonnay, Z.G.; Torok, G.; Makk, J.; Brumbauer, A.; Major, E.; Marialigeti, K.; Toth, E. Bacterial communities in the collection and chlorinated distribution sections of a drinking water system in Budapest, Hungary. J. Basic Microbiol. 2014, 54, 729–738. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Ubiquitous and persistent Proteobacteria and other Gram-negative bacteria in drinking water. Sci. Total Environ. 2017, 586, 1141–1149. [Google Scholar] [CrossRef]
- Prest, E.I.; El-Chakhtoura, J.; Hammes, F.; Saikaly, P.E.; van Loosdrecht, M.C.; Vrouwenvelder, J.S. Combining flow cytometry and 16S rRNA gene pyrosequencing: A promising approach for drinking water monitoring and characterization. Water Res. 2014, 63, 179–189. [Google Scholar] [CrossRef]
- Lautenschlager, K.; Hwang, C.; Liu, W.-T.; Boon, N.; Koster, O.; Vrouwenvelder, H.; Hammes, F. A microbiology-based multi-parametric approach towards assessing biological stability in drinking water distribution networks. Water Res. 2013, 47, 3015–3025. [Google Scholar] [CrossRef] [Green Version]
- Ji, P.; Parks, J.; Edwards, M.A.; Pruden, A. Impact of Water Chemistry, Pipe Material and Stagnation on the Building Plumbing Microbiome. PLoS ONE 2015, 10, e0141087. [Google Scholar] [CrossRef] [Green Version]
- Mi, Z.; Dai, Y.; Xie, S.; Chen, C.; Zhang, X. Impact of disinfection on drinking water biofilm bacterial community. J. Environ. Sci. 2015, 37, 200–205. [Google Scholar] [CrossRef]
- Li, C.; Ling, F.; Zhang, M.; Liu, W.T.; Li, Y.; Liu, W. Characterization of bacterial community dynamics in a full-scale drinking water treatment plant. J. Environ. Sci. 2017, 51, 21–30. [Google Scholar] [CrossRef]
- Hou, L.; Zhou, Q.; Wu, Q.; Gu, Q.; Sun, M.; Zhang, J. Spatiotemporal changes in bacterial community and microbial activity in a full-scale drinking water treatment plant. Sci. Total Environ. 2018, 625, 449–459. [Google Scholar] [CrossRef]
- Oh, S.; Hammes, F.; Liu, W.T. Metagenomic characterization of biofilter microbial communities in a full-scale drinking water treatment plant. Water Res. 2018, 128, 278–285. [Google Scholar] [CrossRef]
- Choi, Y.; Cha, Y.; Kim, B. Characteristics of Bacterial Communities in Biological Filters of Full-Scale Drinking Water Treatment Plants. J. Microbiol. Biotechnol. 2019, 29, 91–104. [Google Scholar] [CrossRef]
- Hong, P.-Y.; Hwang, C.; Ling, F.; Andersen, G.L.; LeChevallier, M.W.; Liu, W.-T. Pyrosequencing Analysis of Bacterial Biofilm Communities in Water Meters of a Drinking Water Distribution System. Appl. Environ. Microbiol. 2010, 76, 5631–5635. [Google Scholar] [CrossRef] [Green Version]
- Ling, F.; Hwang, C.; Le Chevallier, M.S.; Anderson, G.K.; Liu, W.-T. Core-satellite populations and seasonality of water meter biofilms in a metropolitan drinking water distribution system. Int. Soc. Microb. Ecol. 2016, 10, 582–595. [Google Scholar] [CrossRef] [Green Version]
- Kelly, J.J.; Minalt, N.; Culotti, A.; Pryor, M.; Packman, A. Temporal variations in the abundance and composition of biofilm communities colonizing drinking water distribution pipes. PLoS ONE 2014, 9, e98542. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Tang, W.; Ma, J.; Wang, H. Comparison of microbial community shifts in two parallel multi-step drinking water treatment processes. Appl. Microbiol. Biotechnol. 2017, 101, 5531–5541. [Google Scholar] [CrossRef]
- Jaravani, F.D.; Durrheim, D.; Byleveld, P.; Oelgemoeller, M.; Judd, J. Drinking water safety in recreational parks in northern New South Wales, Australia. Australas. J. Environ. Manag. 2015, 22, 432–445. [Google Scholar] [CrossRef]
- Kaestli, M.; O’Donnell, M.O.; Rose, M.; Webb, J.R.; Mayo, M.; Currie, B.J.; Gibb, K. Opportunistic pathogens and large microbial diversity detected in source-to-distribution drinking water of three remote communities in Northern Australia. PLoS Negl. Trop. Dis. 2019, 13, e0007672. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, W.; Staley, C.; Hamilton, K.A.; Beale, D.J.; Sadowsky, M.J.; Toze, S.; Haas, C.N. Amplicon-based taxonomic characterization of bacteria in urban and peri-urban roof-harvested rainwater stored in tanks. Sci. Total Environ. 2017, 576, 326–334. [Google Scholar] [CrossRef]
- Chubaka, C.E.; Whiley, H.; Edwards, J.W.; Ross, K.E. Microbiological Values of Rainwater Harvested in Adelaide. Pathogens 2018, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Hampton, J.; Spencer, P.B.S.; Elliot, A.D.; Thompson, R.C.A. Prevalence of Zoonotic Pathogens from Feral Pigs in Major Public Drinking Water Catchments in Western Australia. EcoHealth 2006, 3, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Koehler, A.V.; Haydon, S.R.; Jex, A.R.; Gasser, R.B. Cryptosporidium and Giardia taxa in faecal samples from animals in catchments supplying the city of Melbourne with drinking water (2011 to 2015). Parasites Vectors 2016, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahedi, A.; Odgers, T.; Ball, A.; Watkinson, A.; Robertson, I.; Ryan, U. Longitudinal analysis of Giardia duodenalis assemblages in animals inhabiting drinking water catchments in New South Wales and Queensland–Australia (2013–2015). Sci. Total Environ. 2020, 718, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Blyton, M.D.; Gordon, D.M. Genetic Attributes of E. coli Isolates from Chlorinated Drinking Water. PLoS ONE 2017, 12, e0169445. [Google Scholar] [CrossRef] [PubMed]
- Nolan, M.J.; Jex, A.R.; Koehler, A.V.; Haydon, S.R.; Stevens, M.A.; Gasser, R.B. Molecular-based investigation of Cryptosporidium and Giardia from animals in water catchments in southeastern Australia. Water Res. 2013, 47, 1726–1740. [Google Scholar] [CrossRef]
- Marshall, H.M.; Carter, R.; Torbey, M.J.; Minion, S.; Tolson, C.; Sidjabat, H.E.; Thomson, R.M. Mycobacterium lentiflavum in drinking water supplies, Australia. Emerg. Infect. Dis. 2011, 17, 395–402. [Google Scholar] [CrossRef]
- Thomson, R.M.; Carter, R.; Tolson, C.; Coulter, C.; Huygens, F.; Hargreaves, M. Factors associated with the isolation of Nontuberculous mycobacteria (NTM) from a large municipal water system in Brisbane, Australia. BMC Microbiol. 2013, 13, 89. [Google Scholar] [CrossRef] [Green Version]
- Thomson, R.; Tolson, C.; Huygens, F.; Hargreaves, M. Strain variation amongst clinical and potable water isolates of M. kansasii using automated repetitive unit PCR. Int. J. Med. Microbiol. 2014, 304, 484–489. [Google Scholar] [CrossRef]
- City of Melbourne. Explore Melbourne/Interactive Maps. 2018. Available online: http://maps.melbourne.vic.gov.au/?_ga=2.185820137.1309982183.1538480420–1070169282.1522717709 (accessed on 24 July 2018).
- Hoy, M. (Cartographer). Fountains in Zurich. 2016. Available online: http://skyplan.ch/maps/zurich-water/ (accessed on 17 April 2016).
- CWW (City West Water). Drinking Water Quality Report 2019. City West Water. 2019. Available online: https://www.citywestwater.com.au/about_us/reports_publications/drinking_water_quality_report.aspx (accessed on 7 January 2020).
- City of Melbourne. Melbourne, Australia. Drinking Water Fountain Instillation Information. 2018. (Personal correspondence via telephone on 15 February 2019).
- CWW (City West Water). Drinking Water Quality Report 2017. City West Water. 2017. Available online: https://www.citywestwater.com.au/sites/default/files/attachments/drinking_water_quality_report_2017.pdf (accessed on 17 February 2022).
- WHO. Heterotrophic Plate Counts and Drinking-Water Safety. 2003. Available online: https://www.who.int/water_sanitation_health/dwq/HPCFull.pdf (accessed on 16 February 2022).
- ADWG. Australian Drinking Water Guidelines 2018 Version 3.5. 2019. Available online: https://www.nhmrc.gov.au/about-us/publications/australian-drinking-water-guidelines#block-views-block-file-attachments-content-block-1 (accessed on 11 December 2019).
- Woese, C.R.; Fox, G.E. Phylogenetic structure of the prokaryotic domain: The primary kingdoms. Evolution 1977, 74, 5088–5090. [Google Scholar] [CrossRef] [Green Version]
- Treibel, T.A.; Manisty, C.; Burton, M.; McKnight, A.; Lambourne, J.; Augusto, J.B.; Couta-Parada, X.; Cutino-Moguel, T.; Noursadeghi, M.; Moon, J.C. COVID-19: PCR screening of asymptomatic healthcare workers at London hospital. Lancet 2020, 395, 1608–1610. [Google Scholar] [CrossRef]
- Utsunomiya, Y.T.; Milanesi, M.; Utsunomiya, A.T.; Ajmone-Marsan, P.; Garcia, J.F. GHap: An R package for genome-wide haplotyping. Bioinformatics 2016, 32, 2861–2862. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R.; Wang, W.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Chao, A. Nonparametric Estimation of the Number of Classes in a Population. Scandinavian J. Stat. 1984, 11, 265–270. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Pielou, E.C. The Measurement of Diversity in Different Types of Biological Collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Clarke, K.; Warwick, R. Primer-6 Computer Program. Natural Environment Research Council, Plymouth; Quest Research Limited: Auckland, New Zealand, 2015. [Google Scholar]
- Schloss, P.D.; Handelsman, J. Introducing DOTUR, a computer program for defining operational taxonomic units and estimating species richness. Appl. Environ. Microbiol. 2005, 71, 1501–1506. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Li, W.; Li, Y.; Zhang, J.; Chen, J.; Zhang, W.; Wu, X. Molecular analysis of bacterial community in the tap water with different water ages of a drinking water distribution system. Front. Environ. Sci. Eng. 2018, 12, 1–10. [Google Scholar] [CrossRef]
- Inkinen, J.; Jayaprakash, B.; Santo Domingo, J.W.; Keinanen-Toivola, M.M.; Ryu, H.; Pitkanen, T. Diversity of ribosomal 16S DNA- and RNA-based bacterial community in an office building drinking water system. J. Appl. Microbiol. 2016, 120, 1723–1738. [Google Scholar] [CrossRef]
- Revetta, R.P.; Gomez-Alvarez, V.; Gerke, T.L.; Santo Domingo, J.W.; Ashbolt, N.J. Changes in bacterial composition of biofilm in a metropolitan drinking water distribution system. J. Appl. Microbiol. 2016, 121, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Bautista-de Los Santos, Q.M.; Schroeder, J.L.; Sevillano-Rivera, M.C.; Sungthong, R.; Ijaz, U.Z.; Sloan, W.T.; Pinto, A.J. Emerging investigators series: Microbial communities in full-scale drinking water distribution systems–a meta-analysis. Environ. Sci. Water Res. Technol. 2016, 2, 611–634. [Google Scholar] [CrossRef]
- Shi, P.; Jia, S.; Zhang, X.X.; Zhang, T.; Cheng, S.; Li, A. Metagenomic insights into chlorination effects on microbial antibiotic resistance in drinking water. Water Res. 2013, 47, 111–120. [Google Scholar] [CrossRef]
- Huang, K.; Zhang, X.X.; Shi, P.; Wu, B.; Ren, H. A comprehensive insight into bacterial virulence in drinking water using 454 pyrosequencing and Illumina high-throughput sequencing. Ecotoxicol. Environ. Saf. 2014, 109, 15–21. [Google Scholar] [CrossRef]
- Gomez-Alvarez, V.; Humrighouse, B.W.; Revetta, R.P.; Santo Domingo, J.W. Bacterial composition in a metropolitan drinking water distribution system utilizing different source waters. J. Water Health 2015, 13, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Liu, W.; Nie, X.; Li, C.; Gu, J.; Zhang, C. Molecular Analysis of Bacterial Communities in Biofilms of a Drinking Water Clearwell. Microbes Environ. 2012, 27, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Douterelo, I.; Fish, K.E.; Boxall, J.B. Succession of bacterial and fungal communities within biofilms of a chlorinated drinking water distribution system. Water Res. 2018, 141, 74–85. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, J.; Mi, Z.; Xie, S.; Chen, C.; Zhang, X. Biofilm bacterial communities in urban drinking water distribution systems transporting waters with different purification strategies. Appl. Microbiol. Biotechnol. 2015, 99, 1947–1955. [Google Scholar] [CrossRef]
- Gerrity, D.; Arnold, M.; Dickenson, E.; Moser, D.; Sackett, J.D.; Wert, E.C. Microbial community characterization of ozone-biofiltration systems in drinking water and potable reuse applications. Water Res. 2018, 135, 207–219. [Google Scholar] [CrossRef]
- Li, X.; Wang, H.; Hu, C.; Yang, M.; Hu, H.; Niu, J. Characteristics of biofilms and iron corrosion scales with ground and surface waters in drinking water distribution systems. Corros. Sci. 2015, 90, 331–339. [Google Scholar] [CrossRef]
- Koskinen, R.; Ali-Vehmas, T.; KaÈmpfer, P.; Laurikkala, M.; Tsitko, I.; Kostya, E.; Atroshi, F.; Salkinoja-Salonen, M. Characterization of Sphingomonas isolates from Finnish and Swedish drinking water distribution systems. J. Appl. Microbiol. 2020, 89, 687–696. [Google Scholar] [CrossRef]
- Gusman, V.; Medic, D.; Jelesic, Z.; Mihajlovic-Ukropina, M. Sphingomonas paucimobilis as a Biofilm Producer. Arch. Biol. Sci. 2012, 64, 1327–1331. [Google Scholar] [CrossRef]
- Sun, H.; Shi, B.; Bai, Y.; Wang, D. Bacterial community of biofilms developed under different water supply conditions in a distribution system. Sci. Total Environ. 2014, 472, 99–107. [Google Scholar] [CrossRef]
- Tsagkari, E.; Keating, C.; Couto, J.M.; Sloan, W.T. A Keystone Methylobacterium Strain in BiofilmFormation in Drinking Water. Water 2017, 9, 778. [Google Scholar] [CrossRef]
- Baik, K.S.; Park, S.C.; Kim, H.J.; Lee, K.H.; Seong, C.N. Chryseoglobus frigidaquae gen. nov., sp. nov., a novel member of the family Microbacteriaceae. Int. J. Syst. Evol. Microbiol. 2010, 60, 1311–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Vanzin, G.; Cupples, A.M.; Strathmann, T.J. Influence of terminal electron-accepting conditions on the soil microbial community and degradation of organic contaminants of emerging concern. Sci. Total Environ. 2020, 706, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Hui, C.; Jiang, H.; Liu, B.; Wei, R.; Zhang, Y.; Zhang, Q.; Zhao, Y. Chitin degradation and the temporary response of bacterial chitinolytic communities to chitin amendment in soil under different fertilization regimes. Sci. Total Environ. 2020, 705. [Google Scholar] [CrossRef]
- Jia, S.; Wu, J.; Ye, L.; Zhao, F.; Li, T.; Zhang, X.-X. Metagenomic assembly provides a deep insight into the antibiotic resistome alteration induced by drinking water chlorination and its correlations with bacterial host changes. J. Hazard. Mater. 2019, 379, 1–9. [Google Scholar] [CrossRef]
- Holinger, E.P.; Ross, K.A.; Robertson, C.E.; Stevens, M.J.; Harris, J.K.; Pace, N.R. Molecular analysis of point-of-use municipal drinking water microbiology. Water Res. 2014, 49, 225–235. [Google Scholar] [CrossRef]
- Gomez-Smith, C.K.; LaPara, T.M.; Hozalski, R.M. Sulfate Reducing Bacteria and Mycobacteria Dominate the Biofilm Communities in a Chloraminated Drinking Water Distribution System. Environ. Sci. Technol. 2015, 49, 8432–8440. [Google Scholar] [CrossRef]
- Vaerewijck, M.J.; Huys, G.; Palomino, J.C.; Swings, J.; Portaels, F. Mycobacteria in drinking water distribution systems: Ecology and significance for human health. FEMS Microbiol. Rev. 2005, 29, 911–934. [Google Scholar] [CrossRef]
- Furuhata, K.; Kato, Y.; Goto, K.; Saitou, K.; Sugiyama, J.-I.; Hara, M.; Fukutama, M. Identification of Yellow-Pigmented Bacteria Isolated from Hospital Tap Water in Japan and their Chlorine Resistance. Biocontrol Sci. 2007, 12, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Cheung, C.Y.; Cheng, N.H.Y.; Ting, W.M.; Chak, W.L. Mycobacterium paragordonae: A rare cause of peritonitis in a peritoneal dialysis patient. Clin. Nephrol. 2017, 88, 371–372. [Google Scholar] [CrossRef]
- Corsaro, D.; Pages, G.S.; Catalan, V.; Loret, J.F.; Greub, G. Biodiversity of amoebae and amoeba-associated bacteria in water treatment plants. Int. J. Hyg. Environ. Health 2010, 213, 158–166. [Google Scholar] [CrossRef]
- Lu, J.; Struewing, I.; Vereen, E.; Kirby, A.E.; Levy, K.; Moe, C.; Ashbolt, N. Molecular Detection of Legionella spp. and their associations with Mycobacterium spp., Pseudomonas aeruginosa and amoeba hosts in a drinking water distribution system. J. Appl. Microbiol. 2015, 120, 509–521. [Google Scholar] [CrossRef]
- Delafont, V.; Mougari, F.; Cambau, E.; Joyeux, M.; Bouchon, D.; Hechard, Y.; Moulin, L. First evidence of amoebae-mycobacteria association in drinking water network. Environ. Sci. Technol. 2014, 48, 11872–11882. [Google Scholar] [CrossRef]
- Li, Q.; Yu, S.; Li, L.; Liu, G.; Gu, Z.; Liu, M.; Ye, Y.; Xia, Q.; Ren, L.; Li, Q.; et al. Microbial Communities Shaped by Treatment Processes in a Drinking Water Treatment Plant and Their Contribution and Threat to Drinking Water Safety. Front. Microbiol. 2017, 8, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ohta, H.; Hattori, R.; Ushiba, Y.; Mitsui, H.; Ito, M.; Watanabe, H.; Tonosaki, A.; Hattori, T.; Ohta, H.; Hattori, R.; et al. Sphingomonas oligophenolica sp. nov., a halo-and organo-sensitive oligotrophic bacterium from paddy soil that degrades phenolic acids at low concentrations. Int. J. Syst. Evol. Microbiol. 2004, 5, 2185–2190. [Google Scholar] [CrossRef]
- Nikitin, D.I.; Tashtemirova, M.A.; Pitriuk, I.A.; Oranskaia, M.S.; Nikitin, L.E. High resistance of some oligotrophic bacteria to ionizing radiation. Mikrobiologiya 1993, 62, 1064–1071. [Google Scholar]
- Slabova, O.I.; Nikitin, D.I. Influence of the incubation temperature on the reaction of oligotrophic bacteria to stress. Microbiology 2003, 73, 758–762. [Google Scholar] [CrossRef]
- Furuhata, K.; Banzai, A.U.; Kawakami, Y.; Ishizaki, N. Genotyping and Chlorine-Resistance of Methylobacterium aquaticum Isolated from Water Samples in Japan. Biocontrol 2011, 16, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-K.; Kim, Y.-K.; Oh, Y.-S.; Choi, S.-C. Growth kinetics and chlorine resistance of heterotrophic bacteria isolated from young biofilms formed on a model drinking water distribution system. Korean J. Microbiol. 2015, 51, 355–363. [Google Scholar] [CrossRef]
- Gulati, P.; Ghosh, M. Biofilm forming ability of Sphingomonas paucimobilis isolated from community drinking water systems on plumbing materials used in water distribution. J. Water Health 2017, 15, 942–954. [Google Scholar] [CrossRef] [Green Version]
- NCBI. Taxonomy Browser. Solirubrobacterales. 2019. Available online: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi (accessed on 18 February 2020).
- Wiegand, S.; Jogler, M.; Boedeker, C.; Pinto, D.; Vollmers, J.; Rivas-Marin, E.; Kohn, T.; Peeters, S.H.; Heuer, A.; Rast, P.; et al. Cultivation and functional characterization of 79 planctomycetes uncovers their unique biology. Nat. Microbiol. 2020, 5, 126–140. [Google Scholar] [CrossRef]
- Wiegand, S.; Jogler, M.; Jogler, C. On the maverick Planctomycetes. FEMS Microbiol. Rev. 2018, 42, 739–760. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Xing, X.; Hu, C.; Wang, H.; Lyu, L. Effect of sequential UV/free chlorine disinfection on opportunistic pathogens and microbial community structure in simulated drinking water distribution systems. Chemosphere 2019, 219, 971–980. [Google Scholar] [CrossRef]
- Perrin, Y.; Bouchon, D.; Delafont, V.; Moulin, L.; Hechard, Y. Microbiome of drinking water: A full-scale spatio-temporal study to monitor water quality in the Paris distribution system. Water Res. 2019, 149, 375–385. [Google Scholar] [CrossRef]
- Douterelo, I.; Sharpe, R.L.; Boxall, J.B. Bacterial community dynamics during the early stages of biofilm formation in a chlorinated experimental drinking water distribution system: Implications for drinking water discolouration. J. Appl. Microbiol. 2014, 117, 286–301. [Google Scholar] [CrossRef] [Green Version]
- Lautenschlager, K.; Boon, N.; Wang, Y.; Egli, T.; Hammes, F. Overnight stagnation of drinking water in household taps induces microbial growth and changes in community composition. Water Res. 2010, 44, 4868–4877. [Google Scholar] [CrossRef]
- Lipphaus, P.; Hammes, F.; Kotzsch, S.; Green, J.; Gillespie, S.; Nocker, A. Microbiological tap water profile of a medium-sized building and effect of water stagnation. Environ. Technol. 2014, 35, 620–628. [Google Scholar] [CrossRef]
- Ling, F.; Whitaker, R.; Le Chevallier, M.W.; Liu, W.T. Drinking water microbiome assembly induced by water stagnation. ISME J. 2018, 12, 1520–1531. [Google Scholar] [CrossRef] [Green Version]
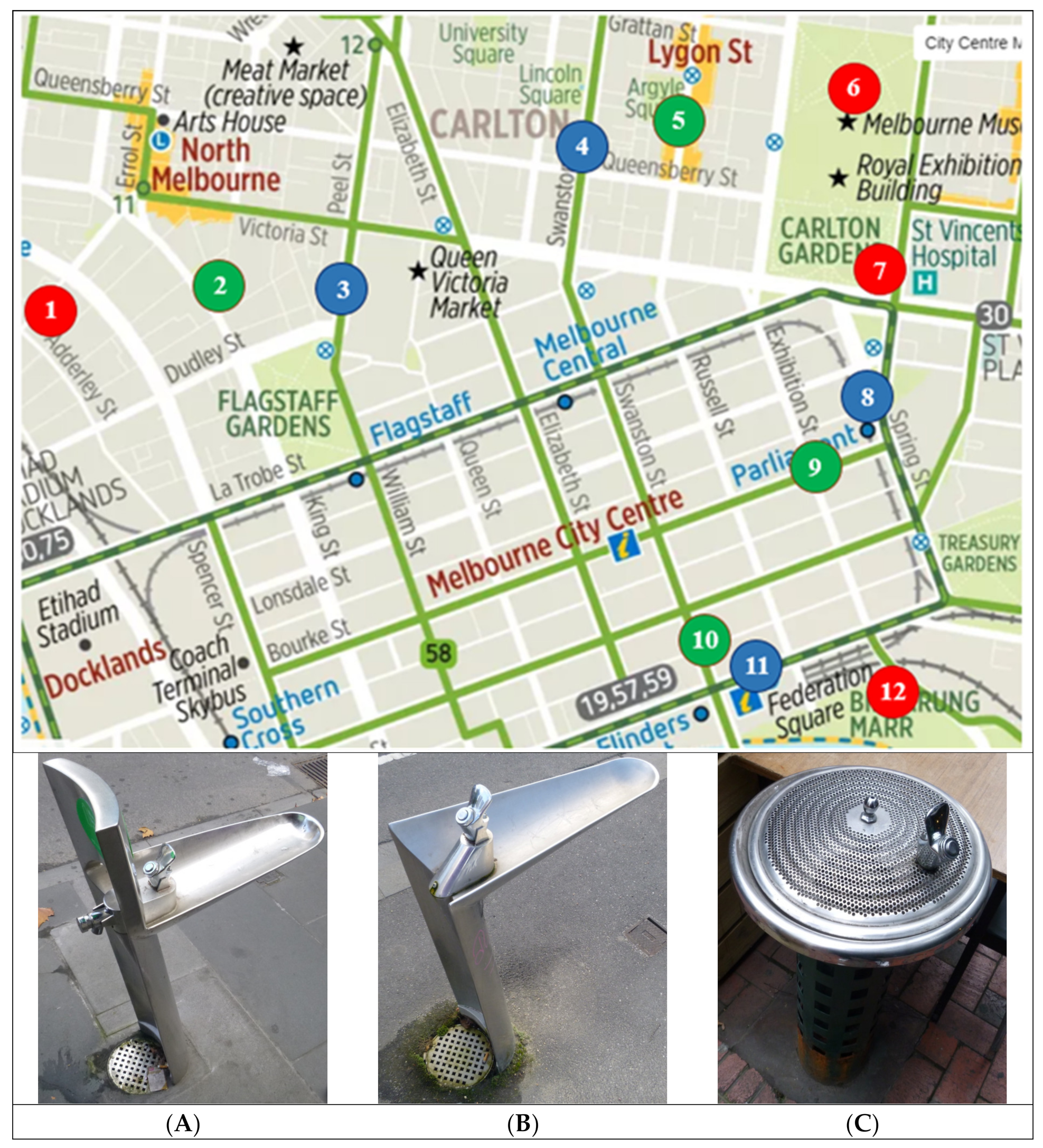
 ), (B) (
), (B) (  ) and (C) (
) and (C) (  ) [40] and images of the three fountain types (A), (B) and (C) (left–right).
), (B) ( ) and (C) ( ) [40] and images of the three fountain types (A), (B) and (C) (left–right).
) [40] and images of the three fountain types (A), (B) and (C) (left–right).
), (B) ( ) and (C) ( ) [40] and images of the three fountain types (A), (B) and (C) (left–right).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Measurement | Guideline Value [45] |
|---|---|---|
| HPC at 22 °C (CFU mL−1) | 56 ± 10 | 1000 |
| HPC at 37 °C (CFU mL−1) | 34 ± 7 | 1000 |
| Total cell counts (CFU mL−1) | 6.1 × 104 ± 2.0 × 104 | NA |
| Coliforms (cells 100 mL−1) | 92.5 ± 54.7 | NA |
| Escherichia coli (cells 100 mL−1) | nd | %100 at zero cells mL−1 |
| pH | 6.85 ± 0.15 | 6.5–8.5 |
| Conductivity (µS cm−1) | 109.4 ± 4.4 | ~900 µS cm−1 |
| Temperature (°C) | 21.46 ± 0.42 | NA |
| Dissolved oxygen (mg L−1) | 6.60 ± 0.44 | NA |
| Sodium (mg L−1) | 10.96 ± 0.50 | 180 mg L−1 |
| Potassium (mg L−1) | 1.67 ± 0.09 | NA |
| Magnesium (mg L−1) | 2.52 ± 0.11 | NA |
| Calcium (mg L−1) | 7.52 ± 0.32 | NA |
| Fluoride (mg L−1) | 0.98 ± 0.01 | 1.5 mg L−1 |
| Chloride (mg L−1) | 22.43 ± 0.22 | 250 mg L−1 |
| Nitrate (mg L−1) | 2.44 ± 0.04 | 50 mg L−1 |
| Sulfate (mg L−1) | 9.59 ± 0.38 | 250 mg L−1 |
| Hardness as CaCO3 (mg L−1) | 18.78 ± 0.80 | 200 mg L−1 |
| Fountain Type | Sample Type | Reads | Observed OTUs | Chao1 Estimates | Shannon Diversity Index |
|---|---|---|---|---|---|
| A (■) | Water | 52,253 ± 10,243 | 139 ± 19 | 193 ± 23 | 2.23 ± 0.18 |
| B (■) | Water | 46,318 ± 7269 | 234 ± 49 | 288 ± 55 | 2.64 ± 0.26 |
| C (■) | Water | 85,097 ± 18,477 | 782 ± 116 | 862 ± 124 | 3.62 ± 0.23 |
| Mean | 61,303 ± 9945 | 394 ± 64 | 457 ± 74 | 2.86 ± 0.16 | |
| A (▲) | Biofilm | 97,961 ± 15,886 | 100 ± 14 | 197 ± 39 | 1.46 ± 0.34 |
| B (▲) | Biofilm | 62,159 ± 13,745 | 82 ±2 2 | 147 ± 27 | 1.07 ± 0.26 |
| C (▲) | Biofilm | 52,476 ± 13,423 | 157 ± 28 | 206 ± 31 | 1.97 ± 0.46 |
| Mean | 70,865 ± 10,047 | 113 ± 16 | 183 ± 20 | 1.50 ± 0.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blyth, W.E.; Shahsavari, E.; Aburto-Medina, A.; Ball, A.S.; Osborn, A.M. Variation in the Structure and Composition of Bacterial Communities within Drinking Water Fountains in Melbourne, Australia. Water 2022, 14, 908. https://doi.org/10.3390/w14060908
Blyth WE, Shahsavari E, Aburto-Medina A, Ball AS, Osborn AM. Variation in the Structure and Composition of Bacterial Communities within Drinking Water Fountains in Melbourne, Australia. Water. 2022; 14(6):908. https://doi.org/10.3390/w14060908
Chicago/Turabian StyleBlyth, Warren Evan, Esmaeil Shahsavari, Arturo Aburto-Medina, Andrew S. Ball, and Andrew Mark Osborn. 2022. "Variation in the Structure and Composition of Bacterial Communities within Drinking Water Fountains in Melbourne, Australia" Water 14, no. 6: 908. https://doi.org/10.3390/w14060908