
Study on the Algae Lysis Method of White Rot Fungi Algae Control System
by

Jun Wang
1,2,†, 
Guoming Zeng
2,†,
Fei Wang
2,
Xin Huang
2,
Yan Li
3,
Dong Liang
2,
Maolan Zhang
2 and
Da Sun
4,*
1
CCTEG Chongqing Engineering (Group) Co., Ltd., Chongqing 401331, China
2
School of Architecture and Engineering, Chongqing University of Science and Technology, Chongqing 401331, China
3
School of Pharmacy, Taizhou Polytechnic College, Taizhou 225300, China
4
Biomedical Collaborative Innovation Center of Zhejiang Province, Institute of Life Sciences, Wenzhou University, Wenzhou 325035, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Water 2022, 14(6), 903; https://doi.org/10.3390/w14060903
Submission received: 11 February 2022
/
Revised: 2 March 2022
/
Accepted: 10 March 2022
/
Published: 14 March 2022
(This article belongs to the Topic Advances in Environmental Biotechnology (AEB))
Abstract
:At present, there are few studies on known bacterial species and even less on fungi in biological algae control technology. In this paper, the green eutrophic shallow water lake Scenedesmus quadricauda (Turpin) was used as the research object, and white rot fungi, which has a high removal effect on water pollutants, algae and biological toxins, was used for algae control. The extent of the removal effect, physiological characteristics and the internal functional groups of the algae cells in the white rot fungi solution, the supernatant of the white rot fungi solution after centrifugation and the sterilized white rot fungi solution were investigated. The results showed that the best algae solubilization effect of the algae control system was achieved at 250 mg/L, with 8 mg/L of dissolved oxygen and a hydraulic retention time of 48 h. The average algae lysis rate was 85.48%, the average dehydrogenase activity reduction rate was 59.23%, the average soluble protein reduction rate was 65.16% and the average malondialdehyde content increased to 0.128 umol/L. After treatment with the white rot algae control system, the spatial structure of the Turpin cells was severely disrupted, and significant lysis occurred within the algal cells, forcing the release of intracellularly soluble substances, and reducing the dehydrogenase activity of the Turpin cells, thus inhibiting the growth activity of the algae cells. A further reduction in the soluble protein content reduces the nutrients required for the growth of Turpin, exacerbating the rate of reduction in the physiological metabolic activity of the Turpin cells and, ultimately, the inhibition or killing of the algal cells. The results of this research may provide theoretical guidance for the microbial control of water eutrophication; however, whether there will be secondary pollution after the algae dissolution of this process is worthy of further study.
1. Introduction
With the influence of human and industrial activities, algal blooms, induced by water eutrophication, have become a serious threat to water ecology and human health around the world [1,2]. Due to the extreme regeneration ability of algae, and their protection mechanism that adapts to the environment, it is very difficult for people to solve the problem of algae-containing water bodies. The common methods of algae control are currently divided into physical, chemical and biological methods. Although physical and chemical methods are effective in controlling algae, they cannot fundamentally address the problem of algal recurrence, and are slightly less economically and environmentally sound than biological methods, with the risk of secondary pollution. It is, therefore, particularly important to explore stable, efficient and safe biological algae control techniques.
Biological algae control is a process that uses the principle of the food chain and the combined growth activities of aquatic plants, aquatic animals and micro-organisms to adsorb, predate and degrade algae in the water column [3]. Due to the constraints of the geographical environment, and the fact that the treatment cycle is long, it is difficult to implement common biological methods that utilize aquatic animals, aquatic plants and their combined techniques to control algae and sufficiently assess the algae removal effect, to meet the needs of the actual project. Microbial algae lysis, as a new type of green water treatment technology, without secondary pollution, plays a role in degrading inorganic salts in water, improving the quality of the water environment and inhibiting the growth of harmful algal blooms [4], which is incomparable to that of aquatic animals and plants, and is simple, efficient and ecologically compatible.
Microbial algae lysis refers to the inhibition of the growth of algae cells, or killing algae cells, through direct or indirect algae lysis methods, and its mode of action is divided into two main types. The first is direct algae lysis [5,6,7], that is, direct contact with algal cells, or invasion into the interior of algal cells, for algae lysis; the second is indirect algae lysis [8,9,10], that is, competition with algae for limited nutrients, or microbial secretion of extracellular substances, inducing algae lysis. At present, there have been more extensive studies on the action mode and mechanism of algolytic bacteria, at home and abroad [11,12,13,14,15], while the mode of action and mechanism of algae lysis by white rot fungi have been studied less.
Consequently, so as to thoroughly and comprehensively explore the mechanism of algae lysis, this paper selected Turpin as the algae species to be studied, and first investigated the following three bacteria solution addition modes of white rot fungi: white rot fungi solution, the supernatant of white rot fungi solution after centrifugation and the sterilized white rot fungi solution. The extent of the removal effect, physiological characteristics and internal functional groups of the algae cells were studied. The aim was to explore, in depth, the effects of changes in the internal and surface structures of algal cells, in the process of algae lysis by the white rot fungi algae control system, on the dominant algae species of eutrophic water bodies, i.e., Turpin, in an attempt to further analyze and determine the mechanism of algae lysis in the treatment of eutrophic water bodies by white rot fungi at the microscopic level.
2. Materials and Methods
2.1. Algal Cultures
The algal species used for the test were Turpin FACHB-507, which was purchased from the Algae Collection Center, Institute of Hydrobiology, Chinese Academy of Sciences. The formula of the medium was provided by the Algae Collection Center, Institute of Hydrobiology, Chinese Academy of Sciences, and the detailed formula is shown in Table 1.
Culture conditions were as follows: temperature (25 ± 1) °C, light intensity 2000 lx–2500 lx, light–dark ratio of 12 h–12 h, and static culture, shaken 3 times a day, so that the growth of algal cells in the same medium tended to reach the same stage. The experiment was carried out once the cells were normal, by microscopy.
2.2. Fungal Strains and Culture
Strain: purchased from China Microbial Strain Network, CGMCC5.776-white rot fungi (Phanerochaete chrysosporium), frozen and stored at −40 °C by the research group after purchase, inoculated into a PDA medium before use, incubated at 25–35 °C for 5–7 days and stored at 4 °C for later use.
PDA solid medium: potato 200 g, glucose 20 g, KH2PO43 3 g, MgSO4·7H2O 1.5 g, vitamin Bl 0.01 g, and agar 20 g.
PDA liquid medium: potato 200 g, glucose 20 g, KH2PO43 3 g, MgSO4.7H2O 1.5 g, and vitamin Bl 0.01 g.
Nutrient solution: 0.4 g glucose, 0.1 g ammonium tartrate, 0.5 g sodium acetate, and 10 mL nutrient salts (3.0 g/L KH2PO43, 4.0 g/L MgSO4-7H2O, 1.0 g/L CaCl2, 0.5 g/L NH4Cl, and 10.0 mg vitamin B1) per 1 liter of mixed water medium, to be sterilized for use.
2.3. Experimental Device
In this experiment, strains were screened in 500 mL conical flasks. The white rot fungi algae control test device consists of plastic barrels, the reactor height is 40 cm, the inner diameter is 30 cm, the effective volume is 20 L, and the hanging film density is 30%. The reactor is equipped with a heating constant temperature device and an immobilized carrier filler. A constant temperature heater is used to control the temperature of the reactor between 15 and 30 °C, and the sequence batch operation mode of “water inlet–reaction–drainage” is adopted. The test setup is shown in Figure 1.
2.4. Experimental Methods
The white rot fungus liquid, the supernatant after centrifugation of the white rot fungus liquid, and the sterilized white rot fungus liquid were added to 250 mL of algae liquid; a sterilized culture medium was added for the control group, and placed in a light box. After 7 days, the algae-dissolving effect and physiological characteristics of the control group and the experimental group were analyzed, respectively.
2.5. Experimental Measurement Index
2.5.1. Measurement of Chlorophyll-a
Chlorophyll-a(ug/L) = [11.64 × (D663 − D750)-2.16 × (D645 − D750) + 0.10 × (D630 − D750)] V1/V/δ.
2.5.2. Determination of Dehydrogenase Activity in Algal Cells
The triphenyl tetrazolium chloride (TTC)–dehydrogenase reduction method [17] was used to determine the dehydrogenase activity of algal cells. The measurement steps were as follows:
- (1)
- Firstly, 15 mL of the solution was taken to be tested and was filtered through a 0.25 μm microporous filter membrane. After suction filtration, the filter membrane was placed into a centrifuge tube, and 2 mL of Tris-HCl (pH = 7.5) and 1 mL of 0.8% (w/v) TTC were successively added until the algal cells were thoroughly mixed. Finally, the sample was placed in a dark place and the reaction was carried out in a constant-temperature water bath at 35 ± 1 °C;
- (2)
- The reaction was terminated after 60 min by adding 3–6 drops of 40% (w/w) formaldehyde solution to the centrifuge tube;
- (3)
- The triphenyl formazone (TPF) was extracted by adding 4 mL of acetone and 5 mL of petroleum ether, and shaken with a vortex mixer for 3 min. All the red extract was placed on the upper layer, and the absorbance of the petroleum ether extract was measured at 492 nm (using petroleum ether as a reference). The dehydrogenase activity (DHA) of algal cells was calculated according to the TPF value. DHA was represented by the TPF produced within 1 h in 1 mL of the test solution (μg TPF/mL.h). All experiments were repeated 3 times.
2.5.3. Determination of Soluble Protein Content
The soluble protein content was determined by the Coomassie brilliant blue G-250 method, using bovine serum protein as a standard.
Determination steps: 5 mL of Coomassie brilliant blue G-250 protein reagent was added to 1 mL of the extracted enzyme solution (the control group was 1 mL of distilled water), mixed well and left for 5 min. The absorbance value of the reaction solution was measured at a wavelength of 595 nm. The content (μg) of soluble protein in the extracted enzyme solution was checked by the standard curve. The preparation method of the required medicine was as follows:
- (1)
- Preparation of 100 mg/L Coomassie brilliant blue reagent: Firstly, 100 mg of G-250 Coomassie brilliant blue was accurately weighed and dissolved in 50 mL of 95% ethanol solution, then 100 mL of 85% (w/v) H3PO4 solution was added, using distilled water to make the total volume of solution 1 L.
- (2)
- Preparation of 0.1 mg/mL standard protein: 10 mg of bovine serum albumin was accurately weighed, then dissolved in 100 mL of distilled water to prepare a 0.1 mg/mL standard protein solution, and stored at 4 °C for later use.
- (3)
- Preparation of 0.05 mol/L phosphate buffer solution with pH value of 7.8: 1.52 g of KH2PO4·2H2O and 14.33 g of K2HPO4·12H2O were accurately weighed, dissolved in distilled water, and made up to 1 L.
2.5.4. Determination of Malondialdehyde Content
According to the determination of the thiobarbituric acid (TBA) method [18], the malondialdehyde content (MDA) reacts with TBA under high-temperature and acidic conditions to form a colored substance with maximum absorbance at a wavelength of 532 nm. The specific measurement steps were as follows:
- (1)
- Firstly, 2 mL of crude enzyme solution was drawn (2 mL of distilled water for the control group), 2 mL of 0.5% (w/v) trichloroacetic acid solution was added, and then 2 mL of 0.67% (w/v) TBA solution was added. This solution was shaken well;
- (2)
- The solution was put in a boiling water bath and boiled for 15 min to make it fully react;
- (3)
- After 15 min., the solution was immediately taken out of the test tube and put into cold water for cooling. After cooling, the solution was centrifuged at 4000 r/min for 20 min. The supernatant absorbance was measured at 600 nm, 532 nm and 450 nm wavelengths, respectively. The MDA concentration and content were calculated according to Equations (2) and (3), as follows:
In the formula, A450, A532 and A600 represent the absorbance values at 450 nm, 532 nm and 600 nm, respectively.
The preparation method of the drugs required for the experiment was as follows:
Preparation of 0.67% TBA solution: 0.67 g of TBA was firstly weighed, then a small amount of 1 mol/L sodium hydroxide was added to dissolve, and the solution was finally settled with 5% trichloroacetic acid to a volume of 100 mL, and dissolved in a constant temperature water bath at 65 °C.
Preparation of 0.5% trichloroacetic acid (TCA) solution: 0.5 g of trichloroacetic acid was accurately weighed and diluted to 100 mL with distilled water.
2.5.5. Fourier Transform Infrared Absorption Spectrometer Detection
Firstly, 10 mL of the fungi liquid, treated in three different ways, was added to 250 mL of algae solution, with 10 mL of sterilized medium added to the control group; appropriate amounts of water samples were taken from the control group and the experimental group after 7 days, and the low-temperature freeze-drying mechanism was used to obtain dry algae powder. Then, KBr was used to make tablets, and the infrared absorption spectra of the dry algae powder in the experimental group and the control group were measured with the aid of a Fourier infrared spectrometer. The Fourier transform infrared absorption spectrometer (FTIR) scanning range was 400 cm−1 to 4000 cm−1.
2.6. Statistical Analysis Methods
The data obtained in the experiment were analyzed and processed using the Origin 8.0 software, OriginLab, USA processing system, and Windows Excel and Word (2003, 2010 editions) office software, Microsoft, USA. All determinations were carried out in triplicate and the mean values were presented. The data are reported as the averages of three separate experiments ± SD (n = 3).
3. Results
3.1. Effect of Three Different Ways of Adding White Rot Fungi on the Treatment of Turpin
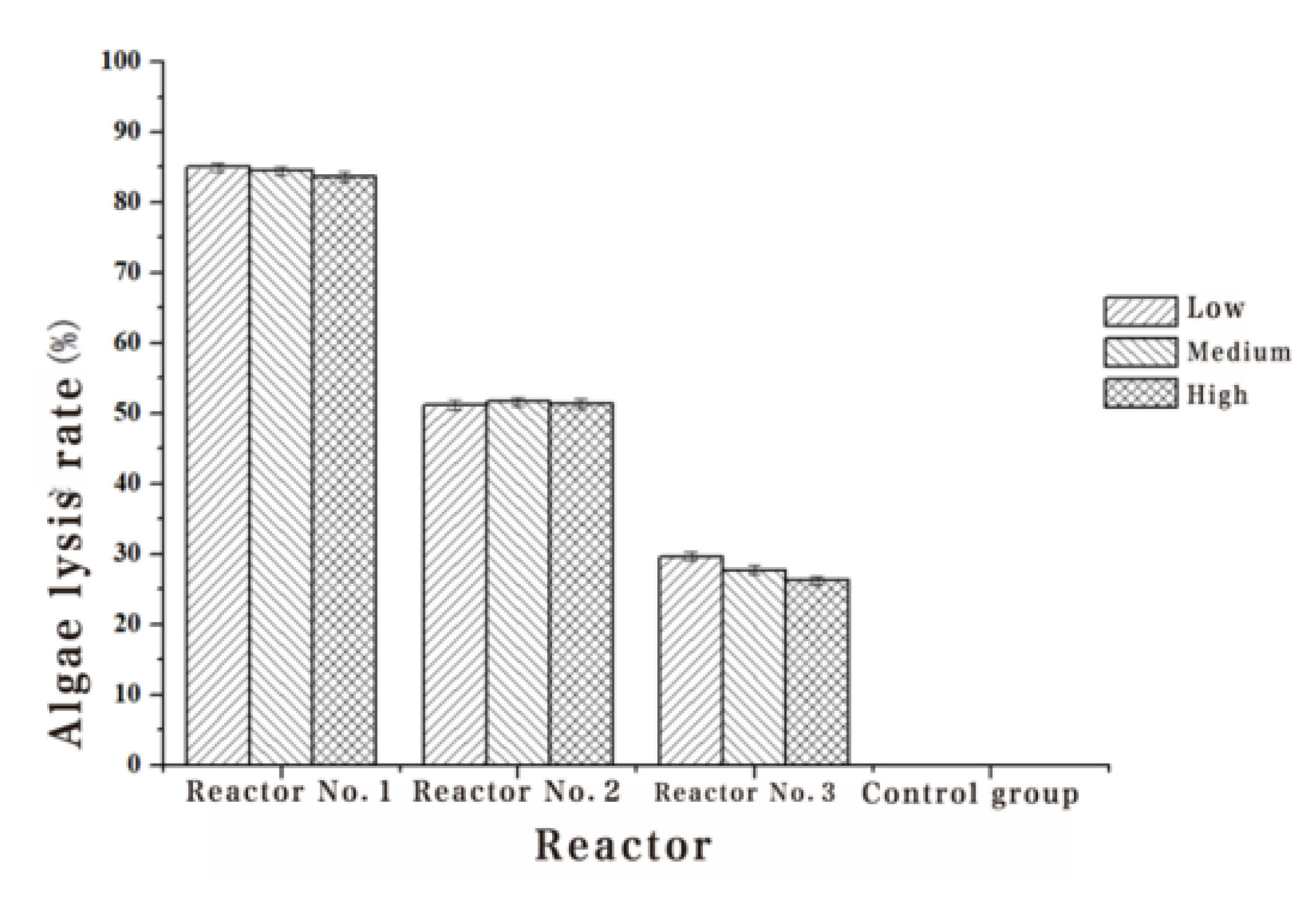
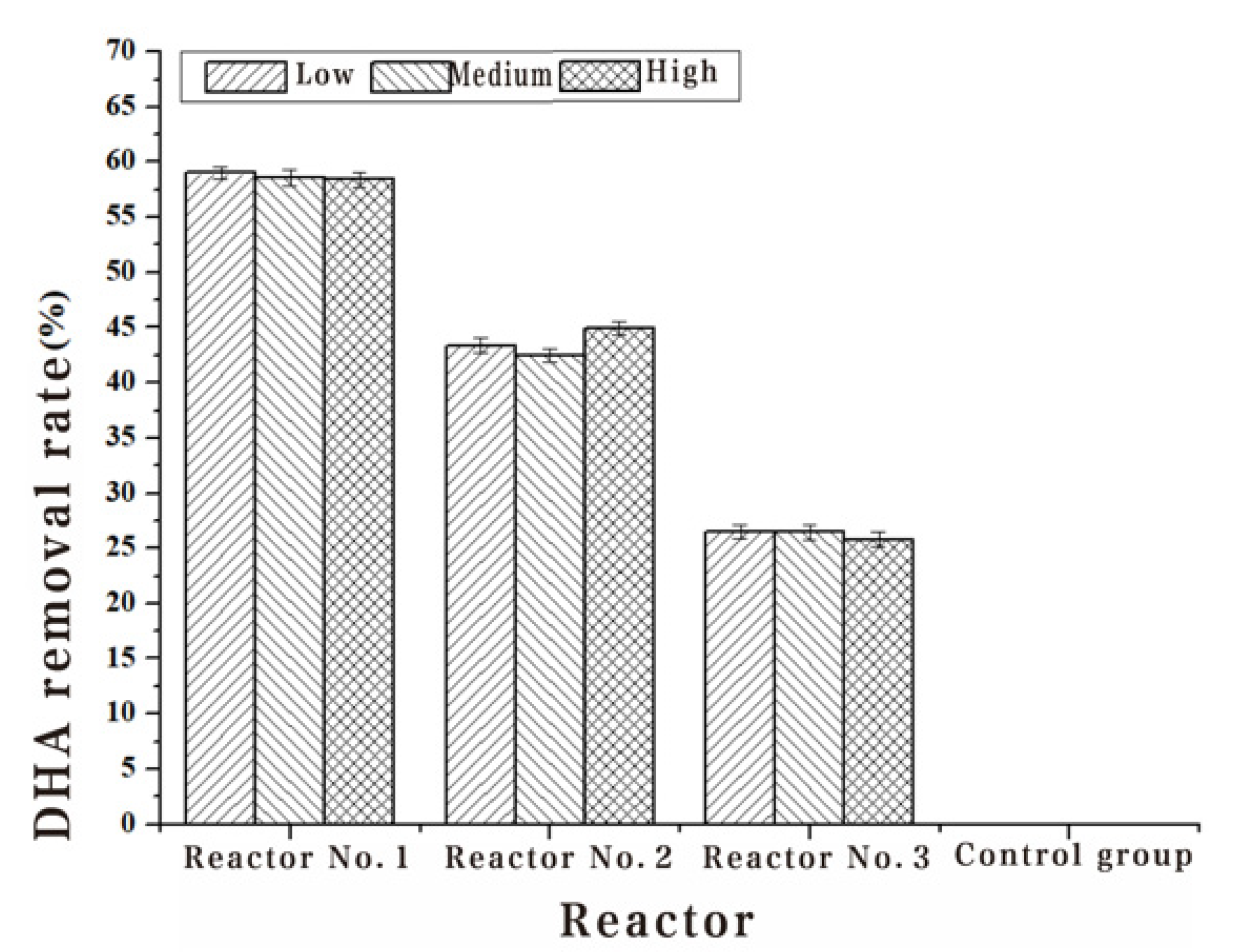
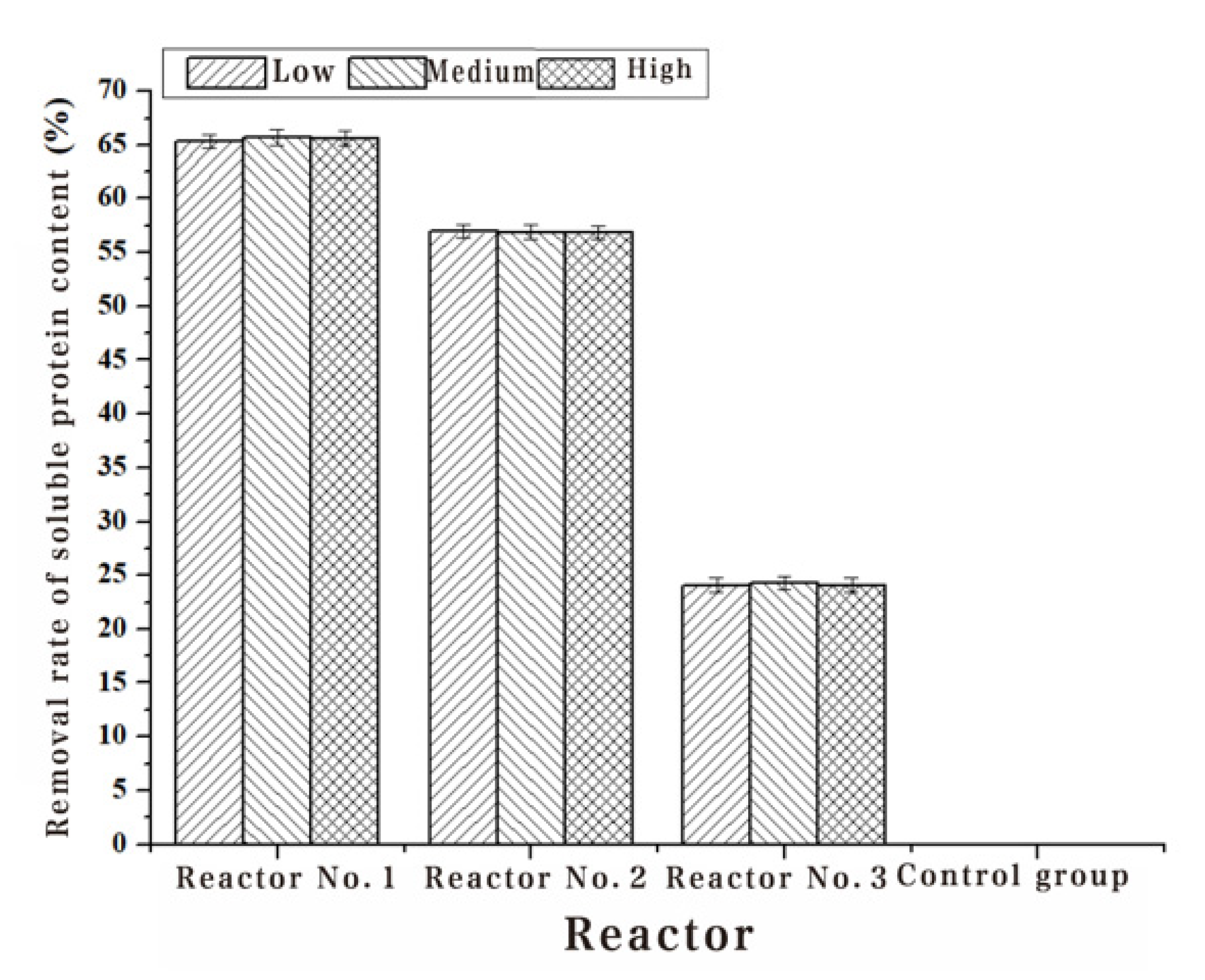
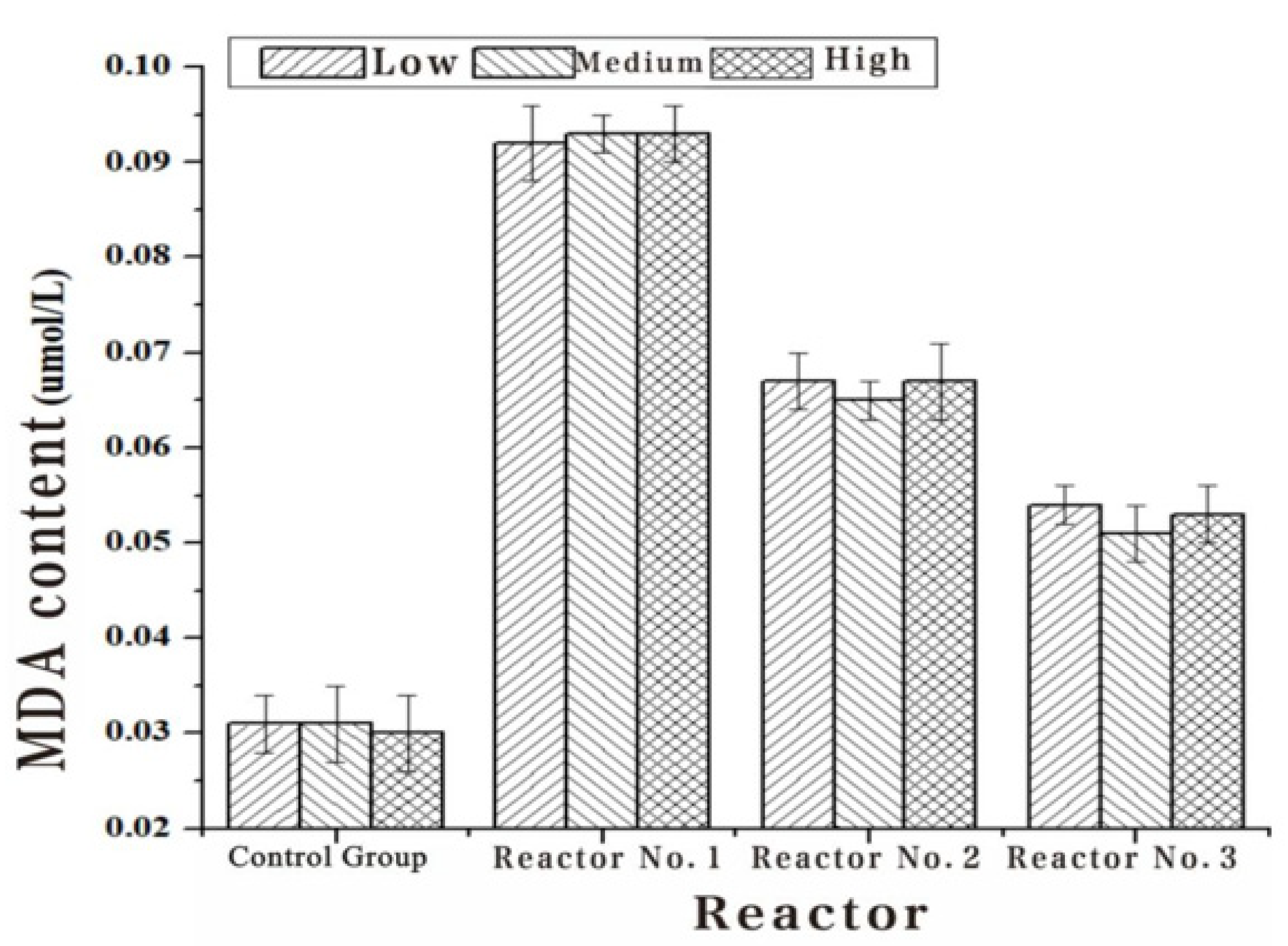
The Chl-a, algae dehydrogenase activity and soluble protein content of Turpin algae cells were significantly decreased after three different methods of adding white rot fungi, while the MDA content increased manifestly in Table 2, Table 3, Table 4 and Table 5. Through the detection results of the Chl-a, dehydrogenase activity, soluble protein content and MDA content of Turpin algae cells in Figure 2, Figure 3, Figure 4 and Figure 5, it was found that reactor 1 has an obvious algae yellow bloom phenomenon, reactor 2 also has a certain yellow bloom algae-dissolving effect, but reactor 3 has no obvious yellow bloom effect in Figure 6. The results demonstrate that the algolytic effect of the white rot fungi liquid on Turpin is significantly better than that of the supernatant after centrifugation and the sterilized fungi liquid. The best algae solubilization effect of the algae control system was achieved with Phanerochaete chrysosporium 250 mg/L, dissolved oxygen 8 mg/L and a hydraulic retention time of 48 h. The average algae lysis rate was 85.48%, the average dehydrogenase activity reduction rate was 59.23%, the average soluble protein reduction rate was 65.16% and the average malondialdehyde content increased to 0.128 umol/L.
3.2. Changes in Algal Cell Functional Groups before and after Treatment with Three Different Addition Methods of White Rot Fungi
Changes in FTIR of Turpin Cells Before and After Treatment with Three Different Addition Methods of White Rot Fungi
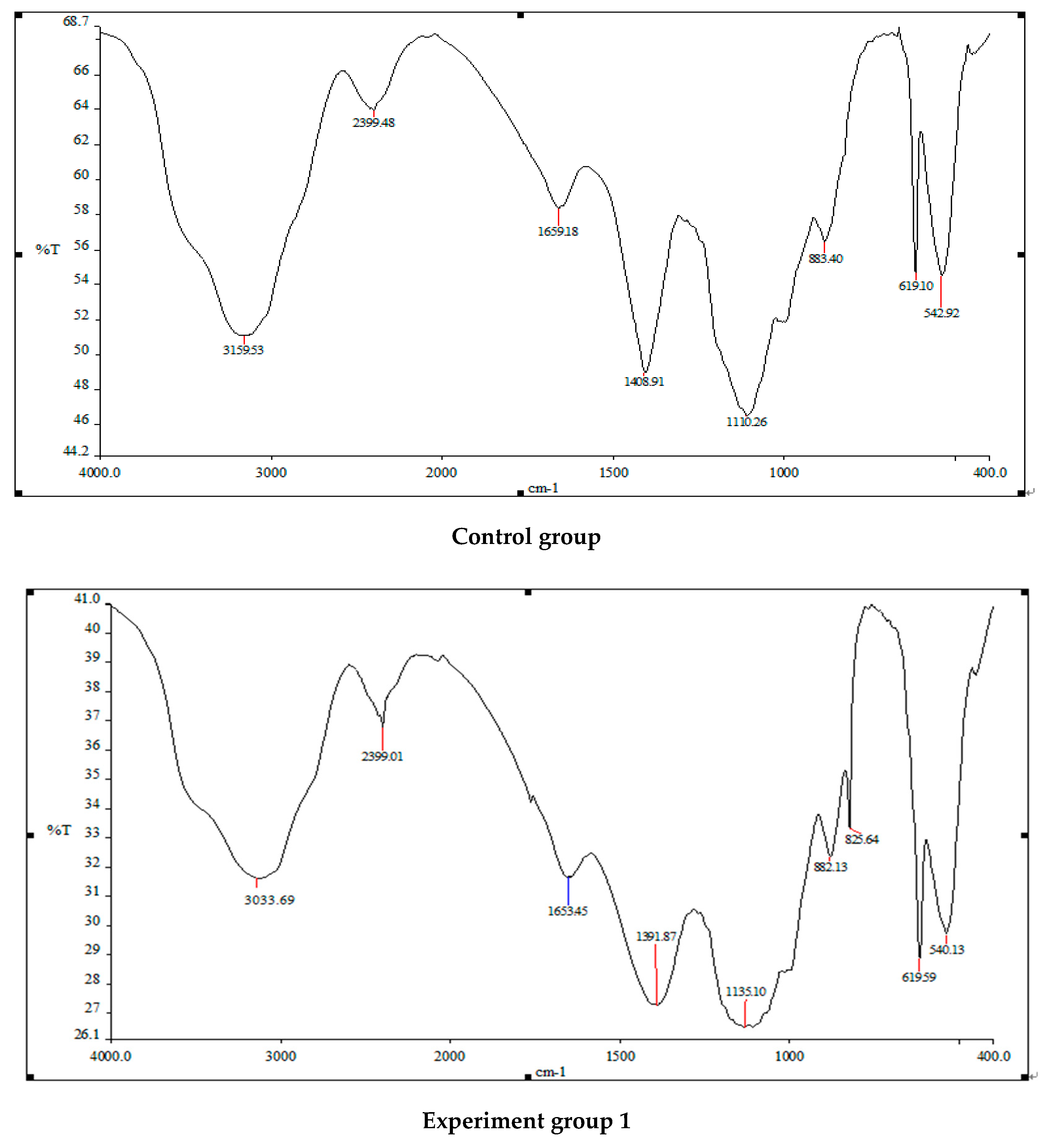
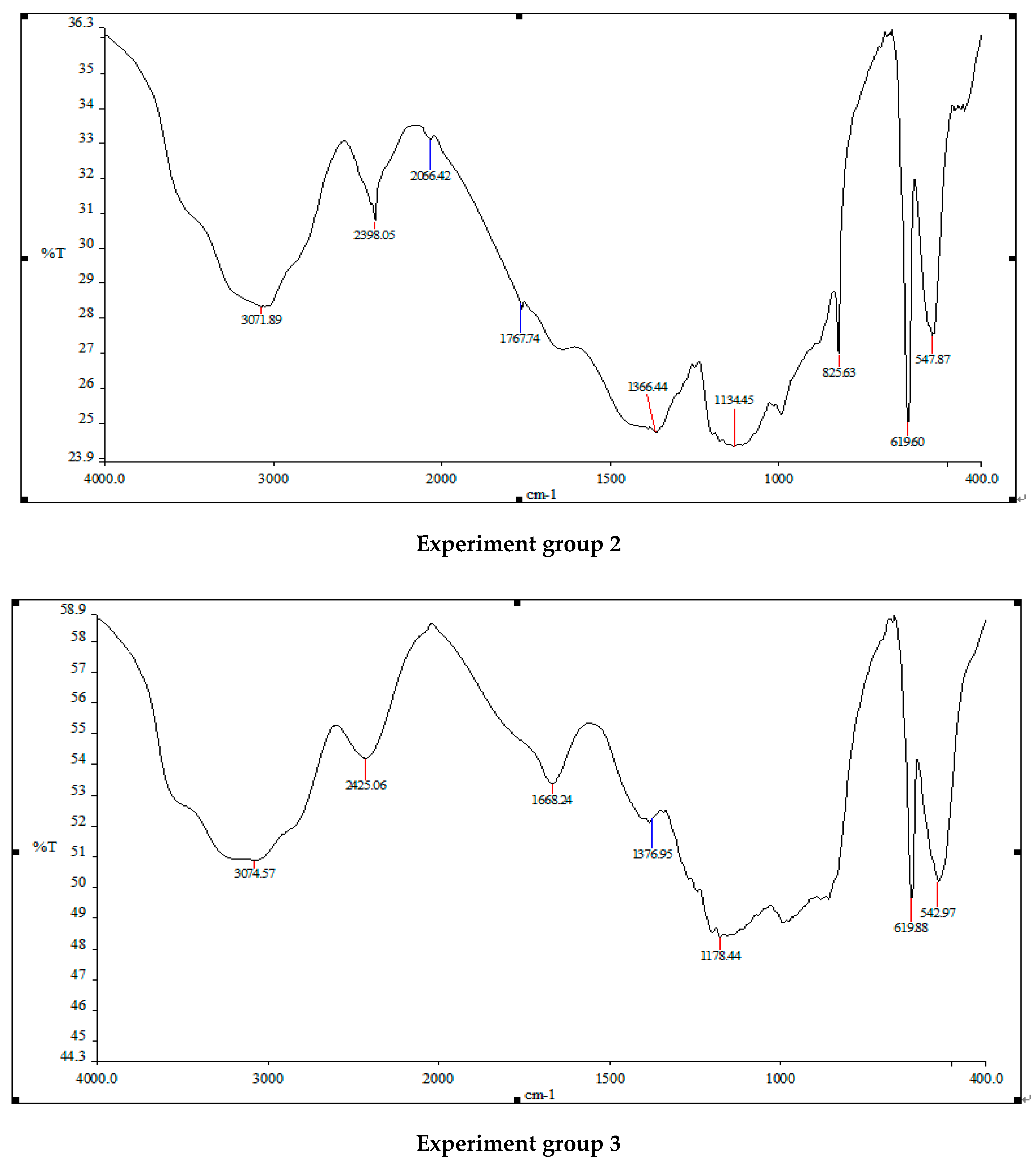
An infrared spectrometer was used to analyze the components of normal Turpin cells and those treated using three different addition methods of white rot fungi in Figure 7. The normal Turpin cells and the main absorption peak positions and waveform shapes of the infrared spectrum of Turpin treated with white rot fungi liquid, white rot fungi supernatant and sterilized white rot fungi liquid are roughly the same, but the relative intensities of the peaks are quite different. The absorption peak intensity of Turpin cells treated using the three methods is obviously weaker than that of the original Turpin cells. After the action of white rot fungi liquid, white rot fungi supernatant and sterilized white rot fungi liquid, Turpin achieves a sharper absorption peak at 3000–3500 cm−1, and a stronger O-H stretching vibration. The intensity of the zone decreased significantly, changing from absorption peaks of 3159.33 to 3033.69, 3071.89 and 3074.57. It is preliminarily speculated that this may be related to the destruction of the O-H bonds in the polysaccharide components in the Turpin cells, and the protein components of the Turpin cells, by white rot fungi; 2300–2900 cm−1 is the stretching vibration zone of Turpin cells saturated with C-H. The absorption peak intensity of Turpin after the three methods of treatment is also significantly weakened. It is inferred that this may be related to the fact that the carbon–hydrogen bonds in the protein structure of the algal cells was degraded by white rot fungi; 1300–1700 cm−1 is the intensity of the C=O of Turpin cell amides. After three methods of treatment, the intensity of the absorption peak of Turpin is also significantly weakened, changing from an absorption peak of 1408.91 to 1391.87, 1364.44 and 1376.95, indicating that the amide bond of the protein in the Turpin cells has been damaged, to a certain extent. There are many small, sharp peaks in the range of 600–1000 cm−1. The above results indicate that the structure of Turpin has been significantly damaged, and the nutrients in the algae cells are released in large quantities. Finally, the Turpin cells’ physiological metabolic activity and photosynthesis of the algae cells are reduced, which is consistent with the significant changes in the functional groups in the algae cells after the action of the white rot fungi reported in the previous period.
4. Discussion
4.1. Effects of Three Addition Methods of White Rot Fungi on the Physiological Metabolism of Algae
Algolytic microorganisms (algicidal bacteria) can inhibit algal growth, or kill algal cells, by direct or indirect algal lysis. In the natural ecosystem, as an important component of the structure and function of biological populations in aquatic ecosystems, microorganisms play an important role in maintaining the balance of algal biomass [18,19,20,21,22,23,24,25,26,27,28]. Based on the experimental results, it can be concluded that the algae lysis effect of the white rot fungi liquid is significantly better than that of the supernatant after centrifugation and the sterilized liquid, illustrating that the algae lysis method of white rot fungi may be a combination of direct and indirect algae lysis, with direct algae lysis being the main mode, and indirect algae lysis being an assist. According to the above-mentioned experimental phenomena and results, it is speculated that the algae lysis method of white rot fungi may initially be that white rot fungi directly lyses algae. After the white rot fungi physically adsorbs to the surface of the algae cell, it begins to damage the cell membrane of the algae cell and the inside nutrients of the phagocytic algae cell. When the nutrients are not enough to support the growth and reproduction of white rot fungi, the white rot fungi begin to initiate indirect algae lysis and secrete a series of algae-lysing substances. In addition, the white rot fungi supernatant and the sterilized white rot fungi have a certain algolytic effect, while the latter has a significantly lower algolytic effect than the former. This indicates that the algae-lysing substance secreted by the white rot fungi mainly contains protein substances, presumably manganese peroxidase, catalase and laccase, but still has a small algae-lysing effect after high-temperature sterilization, which indicates that the secreted algae-lysing substance also contains a small amount of polysaccharides or lipids.
4.2. The Effect of Three Addition Methods of White Rot Fungi on the Functional Groups of Algae
Fourier infrared spectroscopy finds that the amide bonds, O-H bonds and C-H bonds in the algae cells treated by the three methods of adding white rot fungi were significantly weaker than the original algae cells that were not treated by white rot fungi. The algae lysis rate of the algae cells treated in this way is also significantly better than that of the original algae cells. The reason for the analysis results is that the photosynthetic pigment, or chlorophyll a imidazole ring, in the chlorophyll of the algae cells has undergone major changes after treatment with white rot fungi, which ultimately makes the rate of cell algae lysis increase. Consequently, it is speculated that the algae lysis mechanism of white rot fungi is as follows: after the white rot fungi interacts with the algae cells, the O-H and C-H bonds in the algae cells are severely destroyed, which affects photosynthesis and a series of physiological metabolic activities within the algae cells. Ultimately, the growth and metabolic activities of the algae cells are affected.
4.3. Discussion on the Algae Lysis Mechanism of White Rot Fungi
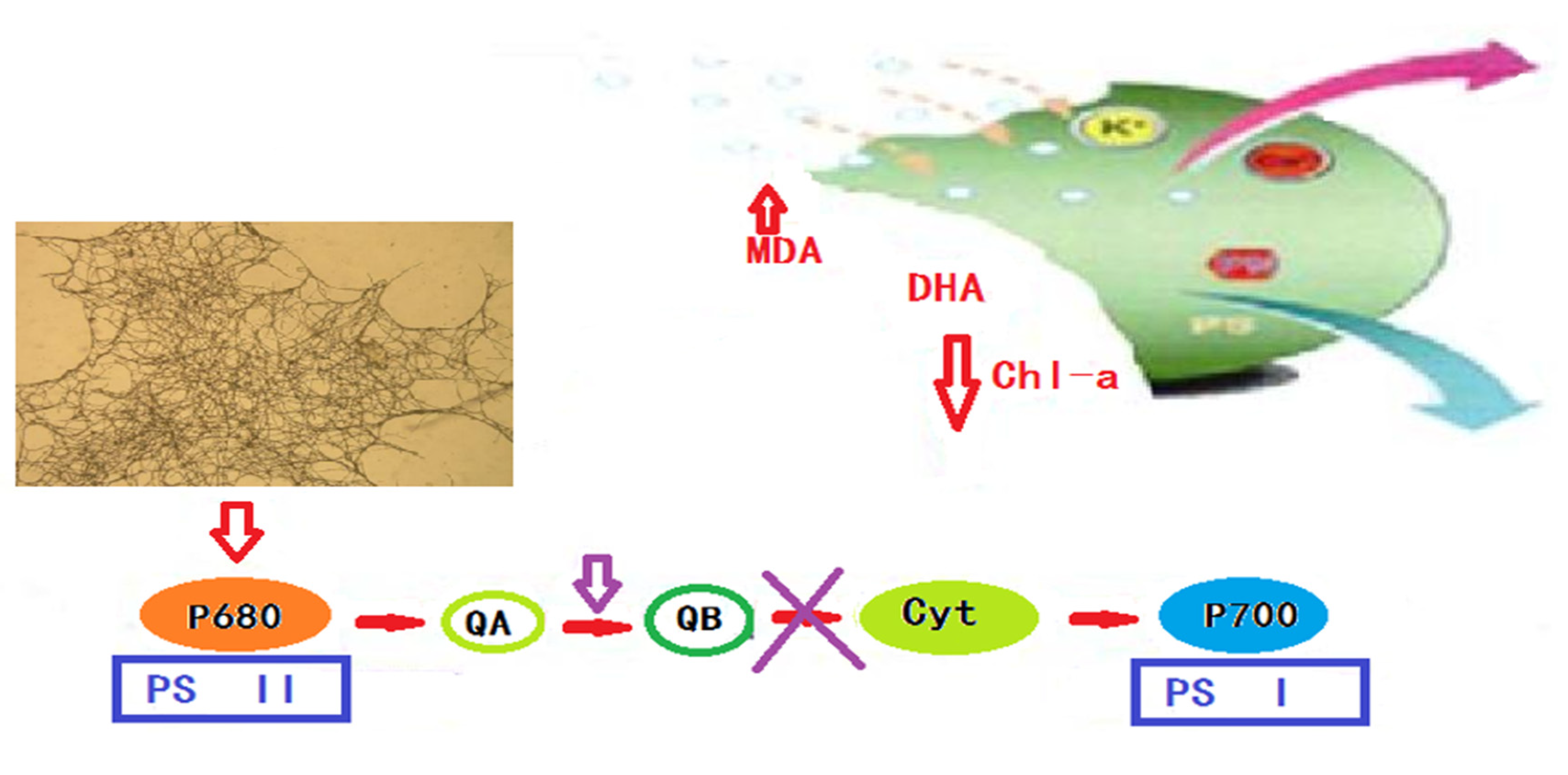
Based on the physiological and biochemical indicators of algae cells, such as the chlorophyll a content, algae cell dehydrogenase activity, soluble protein content, malondialdehyde content, and detection methods, and the results of Fourier infrared spectroscopy, it is speculated that the algae lysis mechanism of white rot fungi is mainly divided into the following three steps in Figure 8: The first step is physical adsorption—after the algae cells are adsorbed onto the carrier filler, the white rot fungi grown on the filler begin to make contact with the algae cells and slowly envelop them, gradually destroying the integrity of the algae cell membrane structure. The second step is the secretion of algae-lysing substances by white rot fungi, such as manganese peroxidase, catalase, laccase, etc., and a small amount of lipid or polysaccharide substances, through the damaged cell membrane, which begin to destroy the photosynthetic pigments in the algae cells, blocking normal photosynthesis and physiological and biochemical metabolic activities in the algae cells. The third step is killing the algae cells. The continuous attack of white rot fungi not only effectively inhibits the normal performance of the physiological and metabolic functions in the algae cells, but also effectively destroys the nucleic acid material of the algae cells, so that the DNA is also damaged, and this ultimately inhibits, or kills, the algae cells.
5. Conclusions
The cell biological characteristics of algal cells and the changes in the algal cell morphological structure during the algae lysis process of Turpin treated by white rot fungi were studied. The main conclusions are as follows:
- (1)
- Based on the above physiological and biochemical indicators, and the results of the Fourier infrared spectroscopy experiment, it is found that the method of algae lysis for white rot fungi may mainly occur through direct algae lysis, supplemented by indirect algae lysis, and the algae-lysing substance contains a large amount of protein substances, and, at the same time, contains a small amount of polysaccharides or lipids.
- (2)
- The algae-dissolving mechanism of the white rot fungi algae control system is as follows: after the white rot fungi make contact with the algal cells, they begin to directly destroy the integrity of the algal cell membrane, and then the algae-lysing substances secreted by the white rot fungi pass through the damaged cell membrane and gradually destroy the algal body. A large number of photosynthetic pigments inhibit the normal physiological metabolism of the algal cells and the occurrence of photosynthesis. Finally, under the continuous action of white rot fungi, the nucleic acid substances, nutrients and genetic materials in algal cells are also severely damaged, which inhibits or kills the algal cells.
Author Contributions
Conceptualization, J.W and G.Z.; methodology, D.L.; software, F.W.; validation, D.S., J.W. and X.H.; formal analysis, Y.L.; investigation, D.L.; resources, D.S.; data curation, F.W.; writing—original draft preparation, J.W.; writing—review and editing, D.S.; visualization, G.Z.; supervision, J.W.; project administration, M.Z.; funding acquisition, D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China, grant number 52103156, 51901160; and Chongqing Science and Technology Commission Project, grant numbercstc2021jcyj-msxmX0663, cstc2020jcyj-msxmX0928, cstc2021jcyj-msxmX0901, cstc2019jcyj-msxmX0562; and Taizhou Science and Technology Sup-port (Agriculture) Project, grant number TN202012, SNY20210008; and Basic scientific re-search project of Wenzhou, grant number S2020 005; and China Postdoctoral Science Foundation, grant number2020M673112;and Provincial and Ministerial Co-constructive of Collaborative Innovation Center for MSW Comprehensive Utilization, grant number shljzyh2021-23; and Chongqing Bayu Scholars Young Scholars Project, grant numberYS2021089; and College Students Innovation Training Program, grant number 2021198, 202211551007; and CCTEG CHONGQING ENGINEERING(GROUP)CO, LTD Science and Technology Innovation Fund, grant number2022-04-12.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
MDPI Research Data Policies at https://www.mdpi.com/ethics (accessed on 9 March 2022).
Conflicts of Interest
No conflict of interest.
References
- Marsalek, B.; Jancula, D.; Marsalkova, E. Multimodal Action and Selective Toxicity of Zerovalent Iron Nanoparticles against Cyanobacteria. Environ. Sci. Technol. 2012, 46, 2316–2323. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.R.; Hong, Y.; Xie, X. Research progress on prevention and control methods of algal blooms and new technologies for inactivation and capture. Environ. Sci. Technol. 2021, 44, 171–185. [Google Scholar]
- Xie, S.L.; Wang, J.; Liu, Q. Research Progress of Plant Allelopathic Algae Control. J. Shanxi Univ. Nat. Sci. Ed. 2017, 40, 652–660. [Google Scholar]
- Ye, X.; Yi, C.; Li, T.L.; Yang, L.; Zhuang, X.C. Current status of research on the application of microbial agents in the water environment. Sichuan Environ. 2021, 40, 240–245. [Google Scholar]
- Yang, L.L.; Pan, W.B.; Li, Y. Composition and algal lysis mechanism of an active algae solubilizing substance from a strain of algae solubilizing bacteria. Environ. Sci. Technol. 2010, 33, 72–75. [Google Scholar]
- He, J.Y.; Pan, W.B.; Lin, M. Effects of algicidal bacteria on algal community structure in eutrophic water. Environ. Pollut. Prev. 2008, 30, 70–74. [Google Scholar]
- Mu, R.; Fan, Z.; Pei, H. Isolation and algae-lysing characteristics of the algicidal bacterium B5. J. Environ. Sci. 2007, 19, 1336–1340. [Google Scholar] [CrossRef]
- Ren, J.; Ma, Q.W.; Huang, H.H. Oxidative degradation of microcystin-LR by combination of UV/H2O2. Fresen Environ. Bull. 2010, 12, 3037–3044. [Google Scholar]
- Hu, H.J.; Wei, Y.X. Freshwater Algae of China—Systems, Taxonomy and Ecology; Science Press: Beijing, China, 2006; pp. 67–68. [Google Scholar]
- Wu, G.; Xi, Y.; Zhao, Y.J. The latest progress in the research of algicidal bacteria. Environ. Sci. Res. 2002, 15, 43–46. [Google Scholar]
- Kim, J.D.; Kim, J.Y.; Park, J.K.; Lee, C.G. Selective Control of the Prorocentrum minimum Harmful Algal Blooms by a Novel Algal-Lytic Bacterium Pseudoalteromonas haloplanktis AFMB-008041. Mar. Biotechnol. 2009, 11, 463–472. [Google Scholar] [CrossRef]
- Kim, M.J.; Jeong, S.Y.; Lee, S.J. Isolation, identification, and algicidal activity of marine bacteria against Cochlodinium polykrikoides. J. Appl. Phycol. 2008, 20, 1069–1078. [Google Scholar] [CrossRef]
- Lawton, L.A.; Welgamage, A.; Manage, P.M.; Edwards, C. Novel bacterial strains for the removal of microcystins from drinking water. Water Sci. Technol. 2011, 63, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Lemes, G.A.F.; Kersanach, R.; Pinto, L.D.; Dellagostin, O.A.; Yunes, J.S.; Matthiensen, A. Biodegradation of microcystins by aquatic Burkholderia sp from a South Brazilian coastal lagoon. Ecotoxicol. Environ. Saf. 2008, 69, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Lewis, W.M.; Wurtsbaugh, W.A.; Paerl, H.W. Rationale for Control of Anthropogenic Nitrogen and Phosphorus to Reduce Eutrophication of Inland Waters. Environ. Sci. Technol. 2011, 45, 10300–10305. [Google Scholar] [CrossRef]
- Zeng, G.M.; Wang, P.; Huang, T. Extraction of Chlorophyll-a from Eutrophic Water by Repeated Freezing and Thawing-Extraction Method. Asian J. Chem. 2014, 26, 2289–2292. [Google Scholar] [CrossRef]
- Xie, J.; Hu, W.R.; Pei, H.Y. Detection of amount and activity of living algae in fresh water by dehydrogenase activity (DHA). Environ. Monit. Assess. 2008, 146, 473–478. [Google Scholar] [CrossRef]
- Ma, H.; Cui, F.Y.; Fan, Z.Q.; Liu, Z.Q.; Zhao, Z.W. Efficient control of Microcystis blooms by promoting biological filter-feeding in raw water. Ecol. Eng. 2012, 47, 71–75. [Google Scholar] [CrossRef]
- Manage, P.M.; Edwards, C.; Singh, B.K.; Lawton, L.A. Isolation and Identification of Novel Microcystin-Degrading Bacteria. Appl. Environ. Microbiol. 2009, 75, 6924–6928. [Google Scholar] [CrossRef] [Green Version]
- Matthijs, H.C.P.; Visser, P.M.; Reeze, B.; Meeuse, J.; Slot, P.C.; Wijn, G.; Talens, R.; Huisman, J. Selective suppression of harmful cyanobacteria in an entire lake with hydrogen peroxide. Water Res. 2012, 46, 1460–1472. [Google Scholar] [CrossRef]
- Paerl, H.W.; Xu, H.; McCarthy, M.J.; Zhu, G.W.; Qin, B.Q.; Li, Y.P.; Gardner, W.S. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N & P) management strategy. Water Res. 2011, 45, 1973–1983. [Google Scholar]
- Park, J.H.; Yoshinaga, I.; Nishikawa, T.; Imai, I. Algicidal bacteria in particle-associated form and in free-living form during a diatom bloom in the Seto Inland Sea, Japan. Aquat. Microb. Ecol. 2010, 60, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Park, S.C.; Lee, J.K.; Kim, S.W.; Park, Y. Selective Algicidal Action of Peptides against Harmful Algal Bloom Species. PLoS ONE 2011, 6, e26733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, H.F.; Chen, W.; Sun, L.W.; Jin, Y.X.; Liu, W.P.; Fu, Z.W. Inhibitory effects of paraquat on photosynthesis and the response to oxidative stress in Chlorella vulgaris. Ecotoxicology 2009, 18, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.H.; Fan, M.H. The Current State of Water Quality and Technology Development for Water Pollution Control in China. Crit. Rev. Environ. Sci. Technol. 2010, 40, 519–560. [Google Scholar] [CrossRef]
- Imamura, N.; Motoike, I.; Shimada, N. An efficient screening approach for anti-Microcystis compounds based on knowledge of aquatic microbial ecosystem. J. Antibiot. 2001, 54, 582–587. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.O.; Kato, J.; Takiguchi, N. Involvement of an extracellular protease in algicidal activity of the marine fungi Pseudoalteromonas sp strain A28. Appl. Environ. Microbiol. 2000, 66, 4334–4339. [Google Scholar] [CrossRef] [Green Version]
- Banin, E.; Khare, S.K.; Naider, F. Proline-rich peptide from the coral pathogen Vibrio shiloi that inhibits photosynthesis of zooxanthellae. Appl. Environ. Microbiol. 2001, 67, 1536–1541. [Google Scholar] [CrossRef] [Green Version]
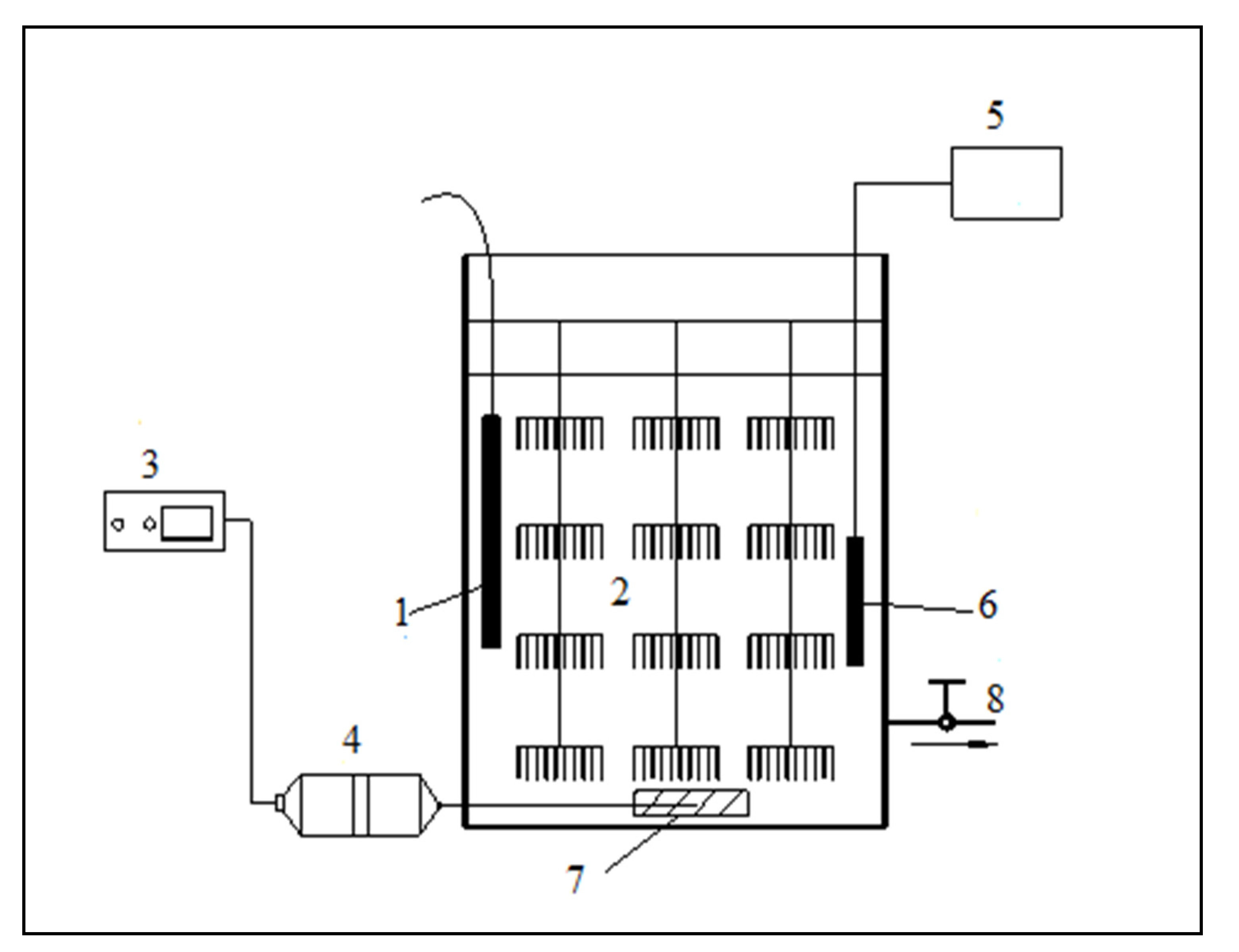
Figure 1.
The experimental equipment of immobilizing white rot fungi: (1) heating units; (2) immobilization carriers; (3) timer; (4) oxygenator; (5) DO detector; (6) DO probe; (7) sand head; (8) drainage and sampling port.
Figure 1.
The experimental equipment of immobilizing white rot fungi: (1) heating units; (2) immobilization carriers; (3) timer; (4) oxygenator; (5) DO detector; (6) DO probe; (7) sand head; (8) drainage and sampling port.

Figure 2.
Effect of three addition methods with Phanerochaete chrysosporium on Chl-a.

Figure 3.
Effect of three addition methods with Phanerochaete chrysosporium on DHA.

Figure 4.
Effect of three addition methods with Phanerochaete chrysosporium on the protein content.

Figure 5.
Effect of three addition methods with Phanerochaete chrysosporium on the MDA content.

Figure 6.
Effect of three addition methods with Phanerochaete chrysosporium on Turpin FACHB-507.

Figure 7.
FTIR on three addition methods with Phanerochaete chrysosporium on Turpin FACHB-507.


Figure 8.
The algae lysis mechanism of white rot fungi.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The growth medium used for Turpin FACHB-507 was SE (pH 7.5).
| Ingredient | Content (g/L) | Soil Nutrient Solution, Content (g/L) | Ingredient |
|---|---|---|---|
| NaNO3 | 0.025 | H3BO3 | 0.286 |
| NaCl | 0.0025 | MnCl2·4H2O | 0.181 |
| K2HPO4·3H2O | 0.0075 | ZnSO4·7H2O | 0.0022 |
| MgSO4·7H2O | 0.0075 | CuSO4·5H2O | 0.0079 |
| CaCl2·2H2O | 0.0025 | Na2MoO4·2H2O | 0.0039 |
| KH2PO4 | 0.0175 | CoCl2·6H2O | 0.0020 |
| FeCl3·6H2O | 0.0005 | ||
| EDTA-Na | 0.0002 | ||
| Soil extract solution | 40 mL−L |
Table 2.
The algicidal efficiency of three addition methods of white rot fungi on Turpin FACHB-507.
| Inlet Water (ug/L) | 145.58 (Low) | 151.67 (Medium) | 161.64 (High) |
|---|---|---|---|
| 1#Out of the water (ug/L) | 21.82 | 23.48 | 26.38 |
| 2#Out of the water (ug/L) | 71.15 | 73.34 | 78.56 |
| 3#Out of the water (ug/L) | 102.38 | 109.58 | 119.26 |
| 4#Comparison of water output (ug/L) | 149.53 | 154.67 | 165.79 |
| 1#Algae lysis rate (%) | 85.01 | 84.52 | 83.68 |
| 2#Algae lysis rate (%) | 51.13 | 51.65 | 51.40 |
| 3#Algae lysis rate (%) | 29.67 | 27.75 | 26.22 |
| 4#Algae lysis rate (%) | - | - | - |
Table 3.
The DHA variation in three addition methods with Phanerochaete chrysosporium.
| Inlet Water (ug TPF/mL.h) | 0.83 | 0.87 | 0.89 |
|---|---|---|---|
| 1#Out of the water (ug TPF/mL.h) | 0.34 | 0.36 | 0.37 |
| 2#Out of the water (ug TPF/mL.h) | 0.47 | 0.50 | 0.49 |
| 3#Out of the water (ug TPF/mL.h) | 0.61 | 0.64 | 0.66 |
| 4#Comparison of water output (ug TPF/mL.h) | 0.86 | 0.90 | 0.95 |
| 1#Removal rate (%) | 59.04 | 58.62 | 58.43 |
| 2#Removal rate (%) | 43.37 | 42.53 | 44.94 |
| 3#Removal rate (%) | 26.51 | 26.44 | 25.84 |
| 4#Removal rate (%) | - | - | - |
Table 4.
The protein content variation in three addition methods with Phanerochaete chrysosporium.
| Inlet Water (ug/mL) | 10.76 | 10.84 | 10.87 |
|---|---|---|---|
| 1#Out of the water (ug/mL) | 3.73 | 3.71 | 3.74 |
| 2#Out of the water (ug/mL) | 4.63 | 4.67 | 4.69 |
| 3#Out of the water (ug/mL) | 8.17 | 8.21 | 8.25 |
| 4#Comparison of water output (ug/mL) | 10.81 | 10.89 | 10.94 |
| 1#Removal rate (%) | 65.33 | 65.77 | 65.59 |
| 2#Removal rate (%) | 56.97 | 56.92 | 56.85 |
| 3#Removal rate (%) | 24.07 | 24.26 | 24.10 |
| 4#Removal rate (%) | - | - | - |
Table 5.
The MDA content variation in three addition methods with Phanerochaete chrysosporium.
| Inlet Water (umol/L) | 0.031 | 0.031 | 0.030 |
|---|---|---|---|
| 1#Out of the water (umol/L) | 0.092 | 0.093 | 0.093 |
| 2#Out of the water (umol/L) | 0.067 | 0.065 | 0.067 |
| 3#Out of the water (umol/L) | 0.054 | 0.051 | 0.053 |
| 4#Out of the water (umol/L) | 0.031 | 0.031 | 0.030 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, J.; Zeng, G.; Wang, F.; Huang, X.; Li, Y.; Liang, D.; Zhang, M.; Sun, D. Study on the Algae Lysis Method of White Rot Fungi Algae Control System. Water 2022, 14, 903. https://doi.org/10.3390/w14060903
AMA Style
Wang J, Zeng G, Wang F, Huang X, Li Y, Liang D, Zhang M, Sun D. Study on the Algae Lysis Method of White Rot Fungi Algae Control System. Water. 2022; 14(6):903. https://doi.org/10.3390/w14060903
Chicago/Turabian StyleWang, Jun, Guoming Zeng, Fei Wang, Xin Huang, Yan Li, Dong Liang, Maolan Zhang, and Da Sun. 2022. "Study on the Algae Lysis Method of White Rot Fungi Algae Control System" Water 14, no. 6: 903. https://doi.org/10.3390/w14060903
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.