
Physiological and Biochemical Responses of Kandelia obovata to Upwelling Stress

1
State Key Laboratory of Tropical Oceanography, South China Sea Institute of Oceanology, Chinese Academy of Sciences, Guangzhou 510301, China
2
Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou), Guangzhou 511458, China
3
Innovation Academy of South China Sea Ecology and Environmental Engineering, Guangzhou 510301, China
4
College of Earth and Planetary Science, University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
Water 2022, 14(6), 899; https://doi.org/10.3390/w14060899
Submission received: 21 January 2022
/
Revised: 9 March 2022
/
Accepted: 11 March 2022
/
Published: 13 March 2022
(This article belongs to the Special Issue Origin and Dynamic of Micropollutants in Contaminated River, Estuary, and Surrounding Coastal Water Systems)
Abstract
:Mangroves growing in intertidal areas are faced with various stresses caused by coastal human activities and oceanic and atmospheric sources. Although the study of the physiological and biochemical characteristics of mangroves has been developing over the past four decades, the effect of upwelling on mangroves in plants stress resistance has seldom been investigated. Here, changes in the physiological and biochemical characteristics of the leaves of Kandelia obovata seedlings in response to upwelling were investigated (air temperature: 25 °C; water temperature: control 25 °C, 13 °C, and 5 °C; salinity: 10‰). The results revealed that upwelling treatment caused an increase in chlorophyll content but a decrease in photosynthetic fluorescence parameters. Hydrogen peroxide (H2O2) production and malondialdehyde activity (MDA) increased with the decrease in upwelling temperature. The proline content increased under upwelling stress, whereas the soluble sugar content decreased. Further, the activities of antioxidant enzymes, such as superoxide dismutase activity (SOD) and peroxidase activity (POD), showed an increasing trend during the treatment, while catalase activity (CAT) decreased. It was evidenced that upwelling stress triggered the physiological and biochemical responses of Kandelia obovata seedlings. This effect became more intense as the upwelling temperature decreased, and all these indicators showed different responses to upwelling stress. Through synthesizing more energy and regulating enzyme activity and osmotic pressure, the leaves of K. obovata formed a resistance mechanism to short-term upwelling.
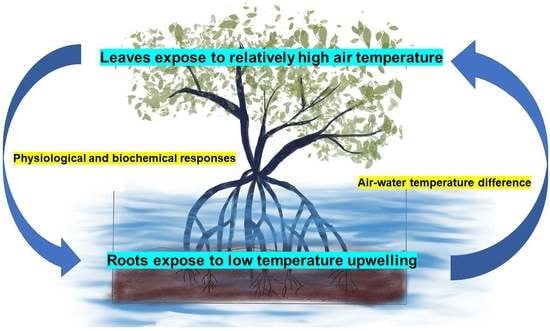
1. Introduction
Mangroves located at the intertidal zones along tropical and subtropical latitudes are considered to be one of the most valuable and productive coastal ecosystems [1,2]. As unique marine ecosystems, mangrove ecosystems have the four distinct characteristics of high productivity, high return rate, high decomposition rate, and high resistance to extreme weather events and anthropogenic activities [3]. They also provide a variety of significant services, including acting as food sources and shelter for many terrestrial and marine species [4,5], protecting coastal areas from storms and floods [6], purifying the environment [7,8], and many other economic and ecological functions [9,10].
Even though mangroves are increasingly valued, forests are rapidly declining and 40% of the tropical mangroves have ecologically degraded over the last century [11]. Plenty of studies have investigated mangrove extinction due to anthropogenic activities such as aquaculture and urban development [12,13] and environmental changes such as extreme weather events, sea level rise affected by climate change, and plastic and microplastic pollution [14,15,16,17]. This loss causes a serious ecological problem and mangroves need urgent conservation to deal with further increases in climate change, which is expected to have a dramatic effect on coastal areas [18,19].
Mangrove estuaries are seawater mixing zones between freshwater and seawater. As a result, mangroves are subjected to stressful situations such as temperature change, salinity, water mixing, and human activities that cause hydrogen, eutrophication, etc., [20,21,22]. The photosynthesis and antioxidant functions of mangrove plants can be inhibited by low temperature and can even cause death [23]. Excessive salinity can result in an osmotic imbalance in mangrove organisms [24]. Inundation can affect the photosynthesis rate and the reactive oxygen production of mangrove plants [25]. Upwelling is an oceanic process in which deep, cold, saline, and nutrient-rich water is transported to the surface, and it plays an important role in coastal ecosystems [26,27,28]. Mangrove plants distributed at low tide level are frequently exposed to upwelling. The difference between an upwelling and non-upwelling area was examined in the tropical bay of Panama. First, although the air temperature is almost the same, the seawater temperature in the upwelling (25.61 °C) and non-upwelling (28.80 °C) areas was different. Second, the mangrove’s mean daily minimum mud in the upwelling (24.2 ± 0.07 °C) area was lower than that in the non-upwelling (25.6 ± 0.06 °C) area. These small but distinct differences caused a reduction in mangrove egg masses and capsules [29]. The upwelling phenomenon has also been reported in the eastern coast of Hainan Island, Leizhou Peninsula, and the Pearl River Estuary where mangroves are largely distributed [30,31]. In particular, the sea surface temperature in the east of Guangdong was as low as 22 °C during an upwelling event [32]. The studies of upwelling in these areas mainly focused on nutrient enrichment and phytoplankton and zooplankton changes [27,33,34]. There is still a lack of knowledge on the relationship between mangrove growth, distribution, and resistance with an air–sea temperature difference caused by upwelling. Hence, the research into the adaptability of mangrove plants to upwelling will provide a critical theoretical basis for mangrove protection.
Kandelia obovata is not only a dominant mangrove species along the south-eastern coast of China but also an important species for mangrove afforestation [35]. As a pioneer species, K. obovata is mainly distributed at low tide and middle tide zones, where it is frequently subjected to upwelling [36,37]. K. obovata can survive in areas with salinity of up to 27.58‰ [38]. Simultaneously, as a mangrove species successfully introduced to northernmost China, K. obovata can survive in unusually cold regions [39]. In mangrove growth and distribution, temperature is the main constraint. Upwelling has the dual characteristics of low temperature and high salinity. To determine the effect of upwelling on K. obovata seedlings, our study investigated the photosynthetic parameters, oxidative stress markers, and antioxidant system of their leaves in different upwelling temperature. Changes in physiological and biochemical characteristics demonstrated the special response mechanism of K. obovata to upwelling.
2. Materials and Methods
2.1. Seedling Collection and Treatment Conditions
Seedlings of well-developed K. obovata with no pests or diseases and similar length were collected from the nursery of the Qi’ao Island Mangrove Nature Reserve in Guangdong. They were transferred to sandy soil and grown using 1/2 Hoagland’s solution for one month to help the plants adapt to the new environment.
Following one month of acclimation in Hoagland nutrient solution, K. obovata seedlings were divided into three groups; each group contained ten seedlings. The second pair of fresh leaves was taken as the measurement material for each index, and three biological replicates were guaranteed. To simulate the upwelling, these three experimental groups were subject to three different levels of water temperature (25 °C (control), 13 °C, 5 °C) and the same air temperature (25 °C) for 12 h. In addition, the water salinity was maintained at 10‰.
2.2. Determination Method
2.2.1. Determination of Chlorophyll Content and Fluorescence Parameters
Chlorophyll a (Chl a) and chlorophyll b (Chl b) content were detected by spectrophotometry after 95% ethanol extraction [40]. Chlorophyll fluorescence parameters were measured using a fluorometer mini-PAM-Ⅱ (Walz, Germany) [41,42]. The following parameters were measured: ratio of variable and maximal fluorescence (Fv/Fm), effective quantum yield of photosystem Ⅱ (Y(Ⅱ)), electron transport rate (ETR), photochemical quenching coefficient (qP), and non-photochemical quenching coefficient (qN).
2.2.2. Determination of Soluble Sugar and Proline
The soluble protein content was measured by the method of Bradford using BSA as a standard [43]. The soluble sugar content was measured using anthrone colorimetry [44]. The concentration of proline was determined according to the Ninhydrin colorimetry by the assay kit (for plant) purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China) [45].
2.2.3. Extraction and Assay of H2O2, MDA, SOD, POD, and CAT
Fresh leaf samples (roughly 1.0 g) were ground using an ice-cold mortar and pestle with 0.05 mol/L Tris hydrochloride buffer (PH 7.8) followed by centrifugation at 3500 rpm and 4 °C for 20 min [46]. The clear supernatant was collected for the measurement of hydrogen peroxide (H2O2), malondialdehyde activity (MDA), superoxide dismutase activity (SOD), peroxidase activity (POD), and catalase activity (CAT) following the instruments of the assay kit (for plant) purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China).
The concentration of H2O2 was measured by monitoring the absorbance of the titanium–peroxide complex at 405 nm [47]. Lipid peroxidation was determined as the amount of MDA produced in the thiobarbituric acid (TBA) reaction [48]. The SOD activity was measured at 560 nm by the nitro-blue-tetrazolium (NBT) photoreduction method [49]. The POD activity was assayed at 470 nm using the guaiacol method [22]. The CAT activity was measured based on the hydrolysis reaction of H2O2 with CAT [49].
2.3. Statistical Analyses
Experimental data were presented as the mean ± SD. All data were verified for normality and homogeneity of variances with the Shapiro Wilk and Levene’s test, respectively. A one-way analysis of variance (ANOVA) and Tukey’s post hoc test were carried out to determine the significance of differences in the various leaf response indexes. p < 0.05 was considered a significant difference. The statistical analysis was performed in SPSS (version 18). Principal component analysis (PCA) was performed using Origin, and the variables were standardized to balance their impact on the weighting in the analysis.
3. Results
3.1. Changes in Photosynthetic Pigment and Fluorescence Parameters
The effect of decreasing temperature on the upwelling stress on chlorophyll contents and fluorescence parameters in K. obovata seedlings is shown in Table 1. A significant difference was only found in qN (p < 0.05). Photosynthetic pigments, Y(Ⅱ), ETR, qP, and Fv/Fm values displayed no significant differences among the groups.
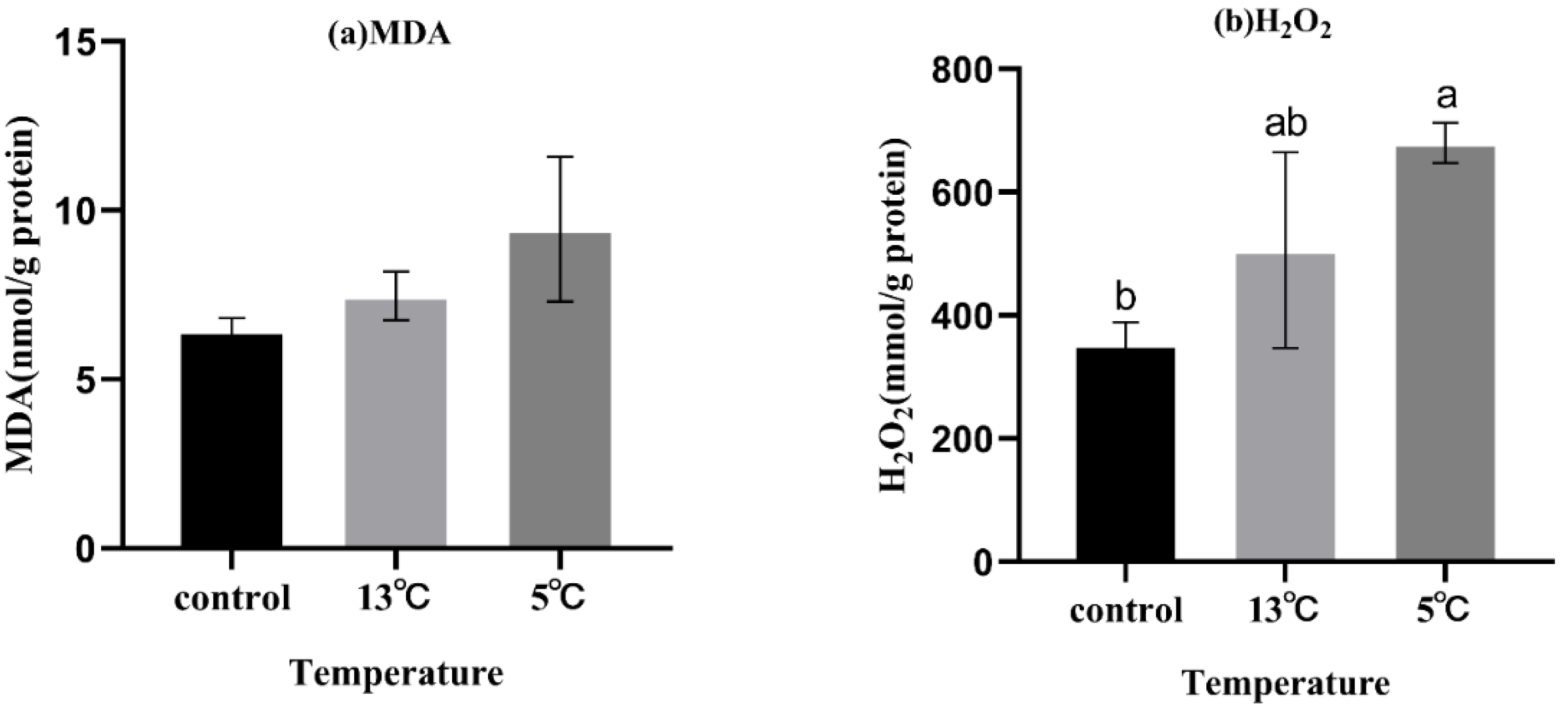
3.2. Changes in H2O2 Production and MDA Content
To evaluate the oxidative stress in K. obovata seedlings under upwelling stress, the MDA and H2O2 contents were measured (Figure 1). The content of H2O2 increased significantly under 5 °C upwelling treatment (p < 0.05). Although with no statistical difference, the content of MDA showed a similar trend to H2O2 after the upwelling treatment.
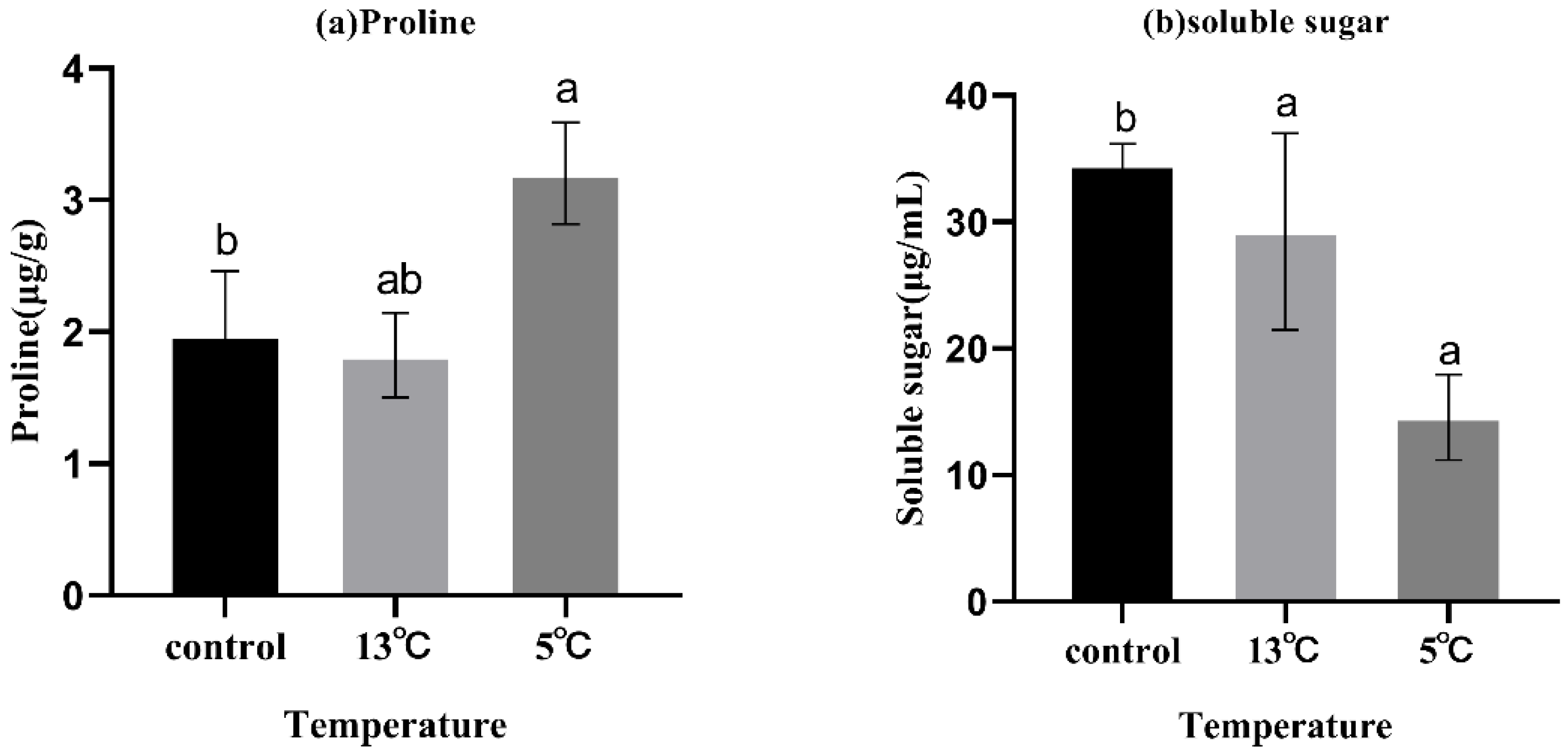
3.3. Changes in Proline and Soluble Sugar Content
The changes in proline and soluble sugar contents in K. obovata seedlings under upwelling stress are shown in Figure 2. Proline content was significantly higher in the 5 °C treatment than in the control (p < 0.05). The content of soluble sugar decreased significantly with the upwelling treatment (p < 0.05), and there was no significant difference between the 13 °C and 5 °C treatments.
3.4. Changes in SOD, POD, and CAT Activity
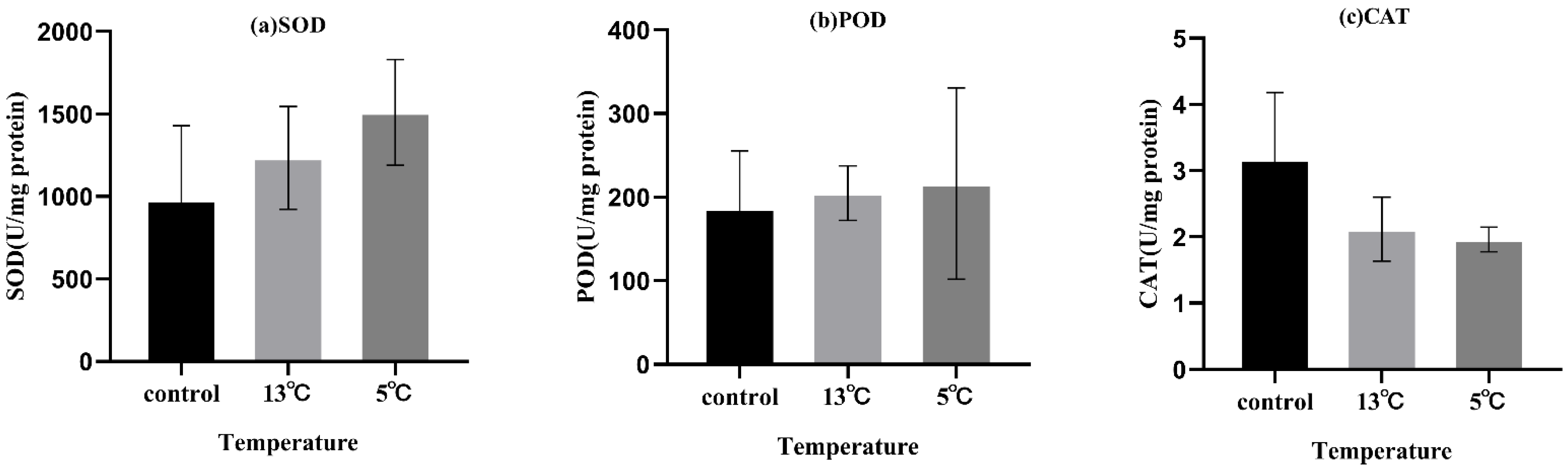
The anti-oxidant enzymes activity of SOD, POD, and CAT are shown in Figure 3. No significant difference was detected in any of the analyzed parameters.
3.5. Principal Component Analysis (PCA)
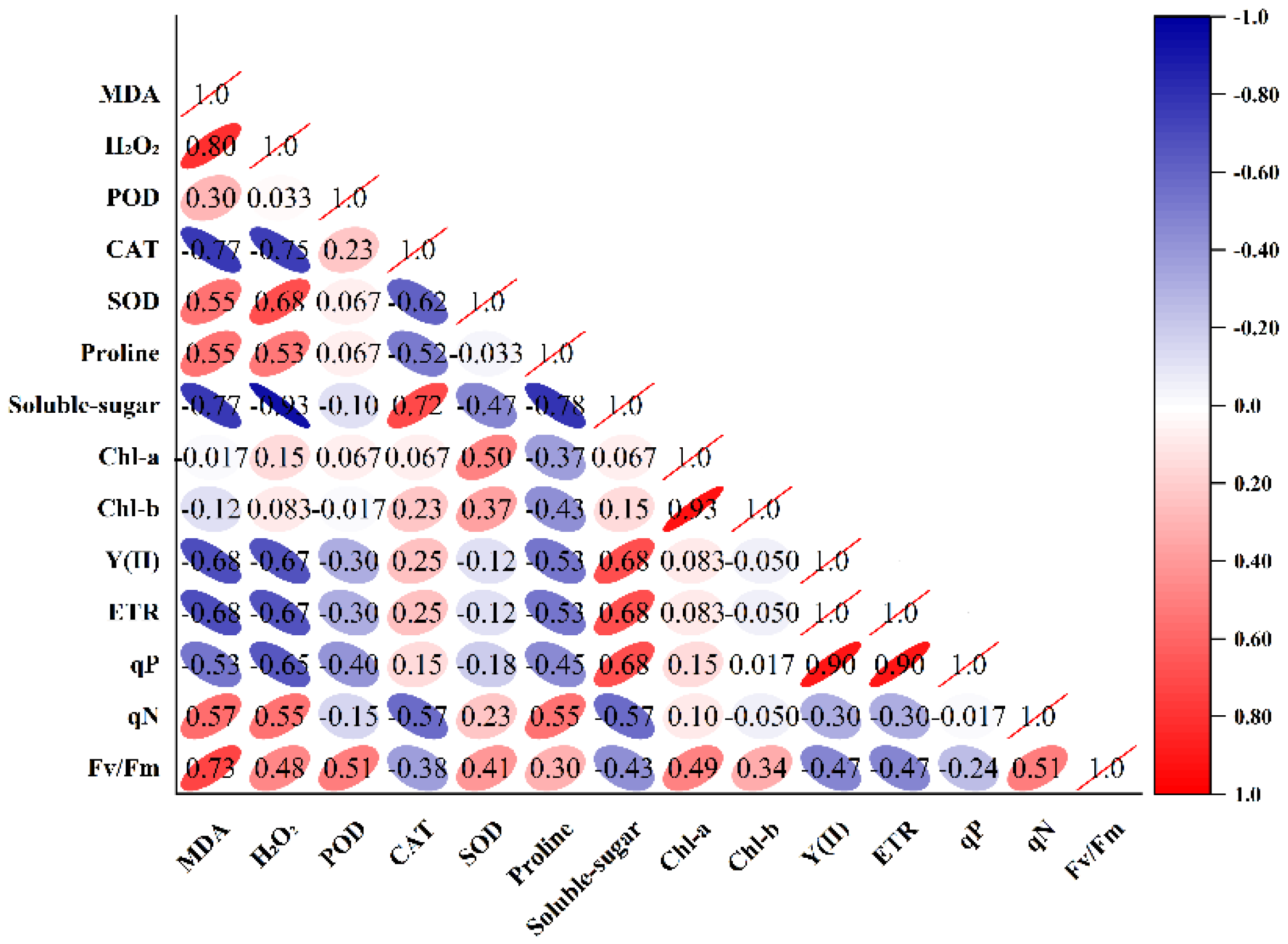
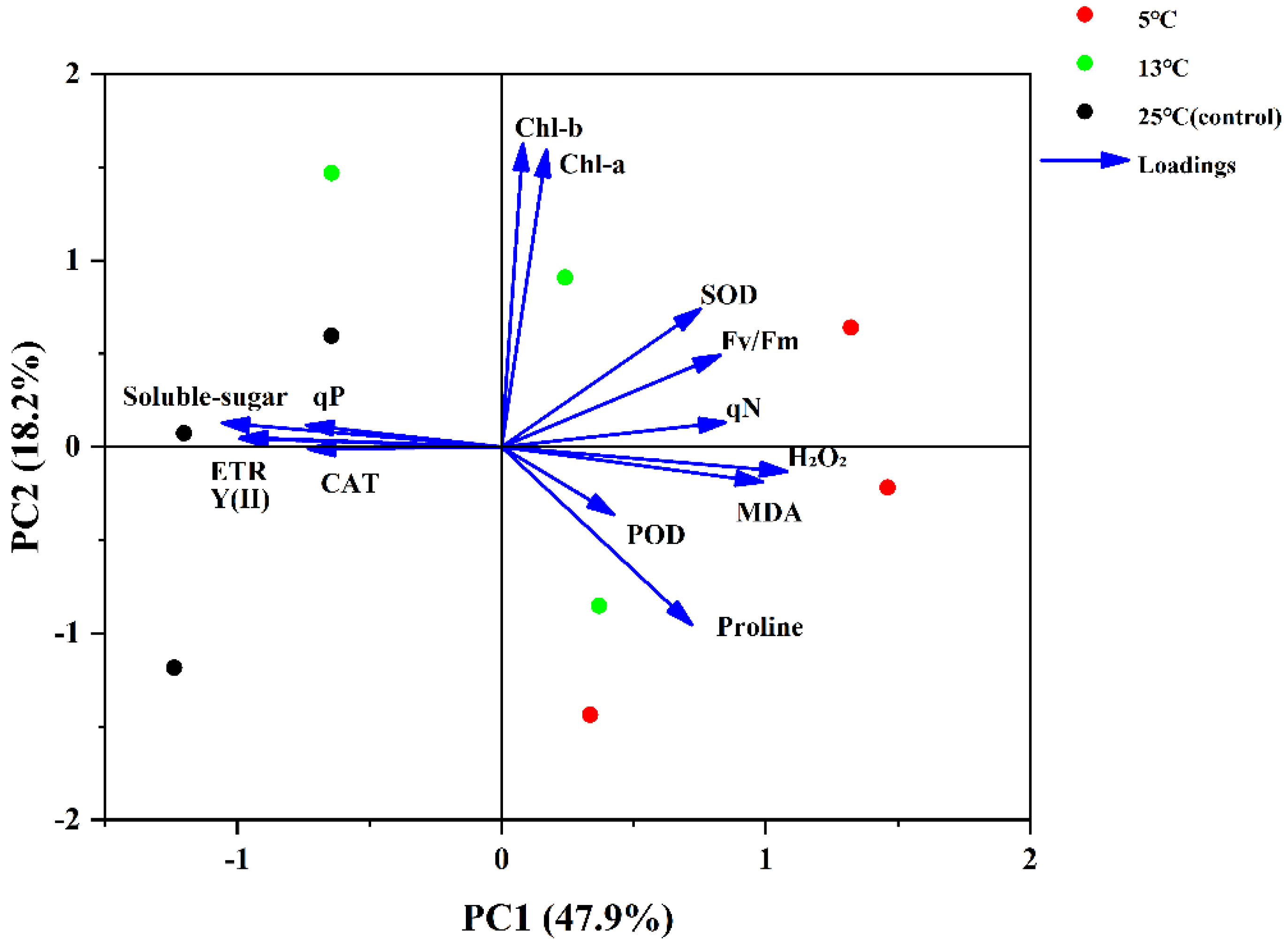
A two-dimensional data matrix, where each row corresponded to a sample at a different upwelling temperature and each column corresponded to a response variable, was employed for principal component analysis (PCA). The results of the correlation coefficient (mostly > 0.3) confirmed the adequacy of the dataset for PCA (Figure 4). The PCA generated eight principal components, and the first two principal components with eigenvalues >1 accounted for 66.01% of total variation (Table 2). PC1 explained 47.86% of the total variance. It was most highly correlated with MDA, H2O2, SOD, soluble sugar, Y(Ⅱ), and ETR. PC1 could be described as the oxidative damage factor, which reflected the accumulation and scavenge system of oxidative stress markers. PC2 described 18.15% of the variance, mainly influenced by the chlorophyll contents (Figure 5). PC2 could be described as the photosynthetic efficiency factor, which showed the strength of the photosynthetic capacity.
4. Discussion
4.1. Response of the Photosynthetic System of K. obovata Seedlings to Upwelling
Chlorophyll is a critical biomolecule in photosynthesis, absorbing light and transforming light energy [50]. Previous studies have shown that the chlorophyll content of K. obovata was decreased under abiotic stress, such as low temperature, high salinity, and flooding [51,52,53]. However, in our study, despite the absence of significant differences for Chl a and Chl b content, their contents were increased under treatment (Table 1). As the leaves were exposed to the air temperature (25 °C), and the roots were exposed to low-temperature upwelling directly, this phenomenon might be a type of response strategy to the air–water temperature difference: increasing chlorophyll content to produce more carbon-based defensive chemicals, thus alleviating root stress [54].
Chlorophyll fluorescence analysis technology is often used in plant photosynthesis mechanism and stress resistance physiology [55]. The value of qN significantly increased under treatment. This is similar to a study where the qN value increased in cucumber and sweet pepper at 4 °C temperature exposure [56]. Our result showed that upwelling treatment induced qN and enhanced the photoprotective mechanism. The Fv/Fm values of all the seedlings obtained in this study were close to 0.8, showing that impairment in plant health status did not occur [57]. The values of Y(Ⅱ), ETR, and qP values showed a decrease compared to the control but no significant change. These results demonstrated that the original light energy capture efficiency in the PS Ⅱ reaction center decreased slightly but was still maintained at a regular level [42]. Combining the chlorophyll content changes with chlorophyll fluorescence analysis, the photosynthetic system of K. obovata seedlings was not seriously damaged, and it could be speculated that K. obovata seedlings can adapt to short-term upwelling stress through different energy allocation strategies for roots and leaves.
4.2. Membrane Damage and Oxidative Stress of K. obovata Seedlings under Upwelling
Mangrove subjected to abiotic stress can experience oxidative damage [47]. Changes in the content of MDA and H2O2 in plants are important indicators of oxidative stress. H2O2 is one of the most significant reactive oxygen species (ROS), and its overproduction promotes lipid oxidation and increases MDA content [58,59]. In the present study, MDA and H2O2 content increased under treatment (Figure 1). In addition, the content of H2O2 significantly increased in the 5 °C treatment (p < 0.05). Similar to the research in strawberry seedlings, MDA and H2O2 increased at low temperatures [60]. The results showed that upwelling could induce H2O2 accumulation and, thus, increase MDA content, resulting in cell membrane damage. This indicates that K. obovata seedlings were somewhat damaged by upwelling stress, and the injury increased as the upwelling temperature decreased.
4.3. Response of Antioxidant Mechanisms of K. obovata Seedlings to Upwelling
In order to scavenge excess ROS and alleviate abiotic stress, plants generate antioxidant mechanisms to protect themselves including enzymatic and non-enzymatic antioxidant response systems [61]. In the enzymatic system, SOD converts free O2– radicals to H2O2 and O2, POD and CAT scavenge the accumulated H2O2 to nontoxic levels or form water and O2 [62,63]. In non-enzymatic antioxidant systems, ascorbate, proline, and soluble sugar can reduce the damaging effect of ROS [64].
In the present study, SOD and POD activity showed a slight increase though no significant difference. Interestingly, the opposite change was observed in CAT activity. The CAT activity decreased under upwelling stress, which was consistent with figleaf gourd under low temperature [65]. This phenomenon might be explained by the fact that K. obovata was a chilling-adaptive plant and had a certain adaptability to low temperature upwelling. In addition, the enzymatic systems of POD and CAT have a shifting peak regulation to reduce ROS accumulation [66]. POD mainly functioned in the early stage of stress, while the 12 h upwelling treatment did not reach the response time of CAT. Therefore, POD and CAT showed a different trend in our study. At the same time, the contents of H2O2 and MDA remained at a rather high level. All these findings indicate that the antioxidant enzyme activities could not maintain the balance between ROS production and elimination.
In the non-enzymatic antioxidant systems, proline plays an important role in scavenging ROS, maintaining osmotic pressure balance, and regulating complex metabolic processes [67]. Soluble sugar, as an important metabolite in plants, also helps to increase osmotic pressure, provides energy and substrate, and induces other physiological and biochemical processes [68]. Proline accumulation in plant tissues under abiotic stress conditions leads to osmotic regulation [69], and this is consistent with our results. However, the soluble sugar content significantly decreased under treatment in our study (p < 0.05). This result was different to alfalfa seedlings [70] and broccoli seedlings [71], when exposed to low temperature. The decrease in soluble sugar content might be because the K. obovata seedlings did not respond in time under the short-term stress, so they needed to consume the stored soluble sugar to generate energy to adapt to upwelling stress, which occurred in bamboo and sweet persimmon [72,73]. Another possible reason could be that the roots were directly exposed to upwelling to cause stress; thus, the leaves consumed the stored soluble sugar to enhance the ability to maintain homeostasis. It could be concluded that K. obovata seedlings alleviate upwelling stress by stimulating enzyme activities, producing osmotic adjustment substances, and generating more energy.
4.4. Relationship between Upwelling Treatment and K. obovata Seedling Response
As the stress response mechanism of plants is sophisticated, a multivariate analysis method must be used to evaluate the comprehensive stress response index [74]. In our study, PCA was implemented to further explore the relationship between upwelling treatment and the K. obovata seedlings’ response mechanism. PC1 was mainly correlated with H2O2 and MDA (positively) as well as soluble sugar (negatively). The results indicated that under upwelling stress, excessive H2O2 and MDA were accumulated in the leaves of K. obovata, and soluble sugar was consumed to resist the stress. Oxidative stress markers (MDA and H2O2) were more sensitive to low temperature, which was unanimous with the results of one-way ANOVA (Figure 1). PC2 was principally determined by chlorophyll content (positively), demonstrating that upwelling treatment enhanced photosynthesis to synthesize more energy to resist stress. In the PCA biplot (Figure 5), the samples of the group treated with 5 °C are located on the right of the figure, whereas the control group is located on the left side of the figure. This separation suggests that the upwelling temperature significantly affected the response mechanism of K. obovata. With the decrease in upwelling temperature, the response of K. obovata seedlings was more obvious. Comparatively, chlorophyll content, H2O2, MDA, SOD, POD, Fv/Fm, and qN increased more significantly at low temperatures. The increase in soluble sugar, ETR, Y(Ⅱ), and CAT were more striking at high temperatures. These results might not only be due to the temperature of upwelling but also to the water–air difference especially for the inconsistent trend for soluble sugar and chlorophyll content compared with former studies [75,76]. In conclusion, PCA revealed the effect of different upwelling temperatures on K. obovata seedlings. A lower upwelling temperature caused more intense response on K. obovata seedlings. The oxidative damage factor and photochemical efficiency factor could be used as the main evaluation indexes of the upwelling resistance of K. obovata. These indicators are also in the stress response systems of other mangrove plants, such as Sonneratia apetala, Bruguiera gymnorhiza, and Kandelia candel (L.) Druce [47,77,78].
5. Conclusions
In this study, chlorophyll content, chlorophyll fluorescence parameters (except qN), H2O2 production, MDA content, SOD activity, POD activity, and proline content increased, while CAT activity, qN value, and soluble sugar content decreased in the seedling leaves of K. obovata under short-term upwelling. The lower the upwelling temperature was, the more obvious the changes were.
The results showed that the leaves of K. obovata can enhance photosynthesis by increasing the chlorophyll content and consume stored soluble sugar to produce more organic substances to alleviate the stress of the air–water temperature difference caused by upwelling. When encountering upwelling, H2O2 and MDA accumulation was observed in K. obovata and caused oxidative stress, which would be alleviated by increased antioxidant enzymes (SOD and POD) and osmotic adjustment substances (proline). The PCA results supported the notion that the chlorophyll content, oxidative stress markers, osmotic adjustment substance, and antioxidant enzymes had different responses to low temperature upwelling. As for the limitation of the experimental conditions, our experiments only revealed the stress response of mangroves to short-term upwelling (12 h). Through improving experimental facilities, we will further study the response of mangroves to long-term upwelling stress in order to better explore the resistance mechanisms of mangroves to upwelling.
Author Contributions
Conceptualization, M.W., Y.W., J.D. and X.L.; methodology, M.W. and X.L.; software, X.L.; writing—original draft preparation, X.L.; writing—review and editing, M.W.; data curation, X.L.; funding acquisition, M.W. and Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (31971480), Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou) (No. GML2019ZD0303) and Innovation Academy of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences (ISEE2019ZR02 and ISEE2018ZD02).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tatongjai, S.; Kraichak, E.; Kermanee, P. Comparative anatomy and salt management of Sonneratia caseolaris (L.) Engl. (Lythraceae) grown in saltwater and freshwater. PeerJ 2021, 9, e10962. [Google Scholar] [CrossRef] [PubMed]
- Himes-Cornell, A.; Grose, S.O.; Pendleton, L. Mangrove ecosystem service values and methodological approaches to valuation: Where do we stand? Front. Mar. Sci. 2018, 5, 376. [Google Scholar] [CrossRef]
- Wang, Y.-S.; Gu, J.-D. Ecological responses, adaptation and mechanisms of mangrove wetland ecosystem to global climate change and anthropogenic activities. Int. Biodeterior. Biodegradat. 2021, 162, 105248. [Google Scholar] [CrossRef]
- Delfan, N.; Shojaei, M.G.; Naderloo, R. Patterns of structural and functional diversity of macrofaunal communities in a subtropical mangrove ecosystem. Estuarine Coast. Shelf Sci. 2021, 252, 107288. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Blaber, S.J.M.; Bouillon, S.; Green, P.; Haywood, M.; Kirton, L.G.; Meynecke, J.-O.; Pawlik, J.; Penrose, H.M.; Sasekumar, A.; et al. The habitat function of mangroves for terrestrial and marine fauna: A review. Aquat. Bot. 2008, 89, 155–185. [Google Scholar] [CrossRef] [Green Version]
- Kathiresan, K.; Rajendran, N. Coastal mangrove forests mitigated tsunami. Estuarine Coast. Shelf Sci. 2005, 65, 601–606. [Google Scholar] [CrossRef]
- Song, H.; Wang, Y.-S.; Sun, C.-C.; Wang, Y.-T.; Peng, Y.-L.; Cheng, H. Effects of pyrene on antioxidant systems and lipid peroxidation level in mangrove plants, Bruguiera gymnorrhiza. Ecotoxicology 2012, 21, 1625–1632. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Sun, X.; Xu, Y.; Zhang, Q.; Li, X. Accumulation and tolerance of mangroves to heavy metals: A review. Curr. Pollut. Rep. 2017, 3, 302–317. [Google Scholar] [CrossRef]
- Anneboina, L.R.; Kumar, K.K. Economic analysis of mangrove and marine fishery linkages in India. Ecosyst. Serv. 2017, 24, 114–123. [Google Scholar] [CrossRef]
- Walters, B.B.; Rönnbäck, P.; Kovacs, J.M.; Crona, B.; Hussain, S.A.; Badola, R.; Primavera, J.H.; Barbier, E.; Dahdouh-Guebas, F. Ethnobiology, socio-economics and management of mangrove forests: A review. Aquat. Bot. 2008, 89, 220–236. [Google Scholar] [CrossRef] [Green Version]
- Padhy, S.R.; Bhattacharyya, P.; Dash, P.K.; Reddy, C.S.; Chakraborty, A.; Pathak, H. Seasonal fluctuation in three mode of greenhouse gases emission in relation to soil labile carbon pools in degraded mangrove, Sundarban, India. Sci. Total Environ. 2020, 705, 135909. [Google Scholar] [CrossRef] [PubMed]
- Pérez, A.; Machado, W.; Sanders, C.J. Anthropogenic and environmental influences on nutrient accumulation in mangrove sediments. Mar. Pollut. Bull. 2021, 165, 112174. [Google Scholar] [CrossRef]
- Pendleton, L.; Donato, D.C.; Murray, B.C.; Crooks, S.; Jenkins, W.A.; Sifleet, S.; Craft, C.B.; Fourqurean, J.; Kauffman, J.B.; Marbà, N.; et al. Estimating Global “Blue Carbon” Emissions from Conversion and Degradation of Vegetated Coastal Ecosystems. PLoS ONE 2012, 7, e43542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodroffe, C.; Rogers, K.; McKee, K.; Lovelock, C.; Mendelssohn, I.; Saintilan, N. Mangrove Sedimentation and Response to Relative Sea-Level Rise. Annu. Rev. Mar. Sci. 2016, 8, 243–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.; Han, J.; Cheung, S.G.; Chong, R.K.Y.; Lo, C.-M.; Lee, F.W.-F.; Xu, S.J.-L.; Yang, Y.; Tam, N.F.-Y.; Zhou, H.-C. How mangrove plants affect microplastic distribution in sediments of coastal wetlands: Case study in Shenzhen Bay, South China. Sci. Total Environ. 2020, 767, 144695. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.D. Marine debris: A proximate threat to marine sustainability in Bootless Bay, Papua New Guinea. Mar. Pollut. Bull. 2012, 64, 1880–1883. [Google Scholar] [CrossRef] [PubMed]
- Leoville, A.; Lagarde, R.; Grondin, H.; Faivre, L.; Rasoanirina, E.; Teichert, N. Influence of environmental conditions on the distribution of burrows of the mud crab, Scylla serrata, in a fringing mangrove ecosystem. Reg. Stud. Mar. Sci. 2021, 43, 101684. [Google Scholar] [CrossRef]
- Yang, Y.; Li, J.; Yang, S.; Li, X.; Fang, L.; Zhong, C.; Duke, N.C.; Zhou, R.; Shi, S. Effects of Pleistocene sea-level fluctuations on mangrove population dynamics: A lesson from Sonneratia alba. BMC Evol. Biol. 2017, 17, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, Q.; Wang, Z.; Tao, J.; Kimura, M.K.; Liu, H.; Hogetsu, T.; Lian, C. Ocean currents drove genetic structure of seven dominant mangrove species along the coastlines of Southern China. Front. Genet. 2021, 12, 268. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Singh, M.; Wang, J.; Xiao, L.; Guan, D. Effects of marine pollution, climate, and tidal range on biomass and sediment organic carbon in Chinese mangrove forests. Catena 2021, 202, 105270. [Google Scholar] [CrossRef]
- Chen, L.; Wang, W.; Li, Q.Q.; Zhang, Y.; Yang, S.; Osland, M.J.; Huang, J.; Peng, C. Mangrove species’ responses to winter air temperature extremes in China. Ecosphere 2017, 8, e01865. [Google Scholar] [CrossRef]
- Barnuevo, A.; Asaeda, T. Integrating the ecophysiology and biochemical stress indicators into the paradigm of mangrove ecology and a rehabilitation blueprint. PLoS ONE 2018, 13, e0202227. [Google Scholar] [CrossRef] [Green Version]
- Du, H.Y.; Zhao, J.; Shi, Y.F.; Che, Y.J. The succession of potential vegetation in China and its sensitivity under climate change. Chin. J. Ecol. 2018, 37, 1459–1466. [Google Scholar] [CrossRef]
- Cheng, H.; Inyang, A.; Li, C.-D.; Fei, J.; Zhou, Y.-W.; Wang, Y.-S. Salt tolerance and exclusion in the mangrove plant Avicennia marina in relation to root apoplastic barriers. Ecotoxicology 2020, 29, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, Z.; Shen, Z.-J.; Luo, M.-R.; Liu, Y.-L.; Wei, M.-Y.; Wang, W.-H.; Qin, Y.-Y.; Gao, C.-H.; Li, K.-K.; et al. Physiological and proteomic responses of mangrove plant Avicennia marina seedlings to simulated periodical inundation. Plant Soil 2020, 450, 231–254. [Google Scholar] [CrossRef]
- Lu, X.; Huang, C.; Chen, F.; Zhang, S.; Lao, Q.; Chen, C.; Wu, J.; Jin, G.; Zhu, Q. Carbon and nitrogen isotopic compositions of particulate organic matter in the upwelling zone off the east coast of Hainan Island, China. Mar. Pollut. Bull. 2021, 167, 112349. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xie, N.; Bai, M.; Li, J.; Wang, G. Composition change and decreased diversity of microbial eukaryotes in the coastal upwelling waters of South China Sea. Sci. Total Environ. 2021, 795, 148892. [Google Scholar] [CrossRef] [PubMed]
- Krek, A.; Krek, E.; Danchenkov, A.; Krechik, V.; Kapustina, M. The role of upwellings in the coastal ecosystem of the Southeastern Baltic Sea. Reg. Stud. Mar. Sci. 2021, 44, 101707. [Google Scholar] [CrossRef]
- Collin, R.; Ochoa, I. Influence of seasonal environmental variation on the reproduction of four tropical marine gastropods. Mar. Ecol. Prog. Ser. 2016, 555, 125–139. [Google Scholar] [CrossRef]
- Risheng Wu, L.L. Summarization of study on upwelling system in the South China Sea. J. Oceanogr. Taiwan Strait 2003, 22, 8. [Google Scholar]
- Jing, Z.-Y.; Qi, Y.-Q.; Hua, Z.-L.; Zhang, H. Numerical study on the summer upwelling system in the northern continental shelf of the South China Sea. Cont. Shelf Res. 2009, 29, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Huang, Z.; Hu, J. Using TPI to map spatial and temporal variations of significant coastal upwelling in the Northern South China Sea. Remote Sens. 2021, 13, 1065. [Google Scholar] [CrossRef]
- Jiang, R.; Wang, Y.-S. Modeling the ecosystem response to summer coastal upwelling in the northern South China Sea. Oceanologia 2018, 60, 32–51. [Google Scholar] [CrossRef]
- Chen, C.-C.; Shiah, F.-K.; Gong, G.-C.; Chen, T.-Y. Impact of upwelling on phytoplankton blooms and hypoxia along the Chinese coast in the East China Sea. Mar. Pollut. Bull. 2021, 167, 112288. [Google Scholar] [CrossRef]
- Pan, D.; Wang, L.; Tan, F.; Lu, S.; Lv, X.; Zaynab, M.; Cheng, C.-L.; Abubakar, Y.S.; Chen, S.; Chen, W. Phosphoproteomics unveils stable energy supply as key to flooding tolerance in Kandelia candel. J. Proteom. 2018, 176, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lin, P. Mangrove Ecosystem in China; Science Press: Beijing, China, 1997. [Google Scholar]
- Lang, T.; Wei, P.; Chen, X.; Fu, Y.; Tam, N.; Hu, Z.; Chen, Z.; Li, F.; Zhou, H. Microcosm study on allelopathic effects of leaf litter leachates and purified condensed tannins from Kandelia obovata on germination and growth of Aegiceras corniculatum. Forests 2021, 12, 1000. [Google Scholar] [CrossRef]
- Liu, Y.-L.; Shen, Z.-J.; Simon, M.; Li, H.; Ma, D.-N.; Zhu, X.-Y.; Zheng, H.-L. Comparative proteomic analysis reveals the regulatory effects of h2s on salt tolerance of mangrove plant Kandelia obovata. Int. J. Mol. Sci. 2019, 21, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.-L.; Wang, Y.-S.; Fei, J.; Sun, C.-C. Isolation and expression analysis of two novel C-repeat binding factor (CBF) genes involved in plant growth and abiotic stress response in mangrove Kandelia obovata. Ecotoxicology 2020, 29, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Jian-Jun Hao, Z.-L.K.; Yang, Y. Experimental Rechnique of Plant Physiology; Chemical Industry Press: Beijing, China, 2006. [Google Scholar]
- Gan, T.; Zhao, N.; Yin, G.; Chen, M.; Wang, X.; Liu, J.; Liu, W. Optimal chlorophyll fluorescence parameter selection for rapid and sensitive detection of lead toxicity to marine microalgae Nitzschia closterium based on chlorophyll fluorescence technology. J. Photochem. Photobiol. B Biol. 2019, 197, 111551. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, G.-q.; Chai, J.; Wang, M.; Jia, X.; Guo, Z.; Sun, L.; Nie, X. Effect of barley yellow dwarf virus infection on photosynthesis and chlorophyll fluorescence parameters of oat. ACTA Agrestia Sin. 2020, 28, 923–931. [Google Scholar] [CrossRef]
- Tao, L.; Ren, H.; Ren, J. Assessment of cultured media for desert moss crust by physiological responses. J. Basic Microbiol. 2021, 61, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Guizhu, C. Physiological adaptability of three mangrove species to salt stress. Acta Ecol. Sin. 2007, 27, 2208–2214. [Google Scholar] [CrossRef]
- Afefe, A.A.; Khedr, A.-H.A.; Abbas, M.S.; Soliman, A.S. Responses and Tolerance Mechanisms of Mangrove Trees to the Ambient Salinity along the Egyptian Red Sea Coast. Limnol. Rev. 2021, 21, 3–13. [Google Scholar] [CrossRef]
- Wang, X.; Wang, D.; Zhang, T.; Zhang, Q.; Zhao, J. iTRAQ-Based Quantitative Proteomic Analysis Reveals Toxicity Mechanisms in Chlamys farreri Exposed to Okadaic Acid. Front. Mar. Sci. 2021, 8, 792050. [Google Scholar] [CrossRef]
- Ru, Q.M.; Xiao, Q.; Lin, P.; Pei, Z.M.; Zheng, H.L. Short- and long-term effects of NaCl on physiological and biochemical characteristics in leaves of a true mangrove, Kandelia candel. Russ. J. Plant Physiol. 2009, 56, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Lu, H.; Wang, Y.; Hong, H.; Wang, Q.; Liu, J.; Yan, C. Uptake, biotransformation and physiological response of TBBPA in mangrove plants after hydroponics exposure. Mar. Pollut. Bull. 2020, 151, 110832. [Google Scholar] [CrossRef]
- Li, H.-X.; Xiao, Y.; Cao, L.-L.; Yan, X.; Li, C.; Shi, H.-Y.; Wang, J.-W.; Ye, Y.-H. Cerebroside C Increases Tolerance to Chilling Injury and Alters Lipid Composition in Wheat Roots. PLoS ONE 2013, 8, e73380. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Yu, K.; He, T.; Li, F.; Zhang, D.; Liu, J. The low temperature induced physiological responses of Avena nuda L.; a cold-tolerant plant species. Sci. World J. 2013, 2013, 658793. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; An, B.; Cao, D.; Xu, R.; Wang, S.; Zhang, Z.; Liu, X.; Sun, X. Improving photosynthetic capacity, alleviating photosynthetic inhibition and oxidative stress under low temperature stress with exogenous hydrogen sulfide in blueberry seedlings. Front. Plant Sci. 2020, 11, 108. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Xue, C.; Chen, H.; He, C.; Wang, Q. Low-Temperature Adaptation of the Snow Alga Chlamydomonas nivalis Is Associated With the Photosynthetic System Regulatory Process. Front. Microbiol. 2020, 11, 1233. [Google Scholar] [CrossRef]
- Wunderlich, A.; Pinheiro, M.A.A. Mangrove habitat partitioning by Ucides cordatus (Ucididae): Effects of the degree of tidal flooding and tree-species composition during its life cycle. Helgol. Mar. Res. 2012, 67, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Agathokleous, E.; Feng, Z.; Peñuelas, J. Chlorophyll hormesis: Are chlorophylls major components of stress biology in higher plants? Sci. Total Environ. 2020, 726, 138637. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-H.; Lin, K.-H.; Jiang, J.-Y.; Wang, C.-W.; Chen, C.-I.; Huang, M.-Y.; Weng, J.-H. Comparisons between yellow and green leaves of sweet potato cultivars in chlorophyll fluorescence during various temperature regimes under high light intensities. Sci. Hortic. 2021, 288, 110335. [Google Scholar] [CrossRef]
- Li, X.-G.; Meng, Q.-W.; Jiang, G.-Q.; Zou, Q. the susceptibility of cucumber and sweet pepper to chilling under low irradiance is related to energy dissipation and water-water cycle. Photosynthetica 2003, 41, 259–265. [Google Scholar] [CrossRef]
- Popov, V.N.; Antipina, O.V.; Selivanov, A.A.; Rakhmankulova, Z.F.; Deryabin, A. Functional activity of the photosynthetic apparatus in tobacco and arabidopsis plants exposed to chilling temperatures. Russ. J. Plant Physiol. 2019, 66, 102–109. [Google Scholar] [CrossRef]
- Farzana, S.; Chen, J.; Pan, Y.; Wong, Y.-S.; Tam, N.F.Y. Antioxidative response of Kandelia obovata, a true mangrove species, to polybrominated diphenyl ethers (BDE-99 and BDE-209) during germination and early growth. Mar. Pollut. Bull. 2016, 124, 1063–1070. [Google Scholar] [CrossRef]
- Kamran, R.V.; Toorchi, M.; Moghadam, M.; Mohammadi, H. The effect of cold stress on H2O2 and MDA contents in barely genotypes. J. Biodivers. Enviromental Sci. 2015, 7, 66–75. [Google Scholar]
- Xu, C.; Wang, M.; Yang, Z.; Zheng, Q. Low temperature and low irradiation induced irreversible damage of strawberry seedlings. Photosynthetica 2020, 58, 156–164. [Google Scholar] [CrossRef]
- Sun, W.H.; Duan, M.; Li, F.; Shu, D.F.; Yang, S.; Meng, Q.W. Overexpression of tomato tAPX gene in tobacco improves tolerance to high or low temperature stress. Biol. Plant. 2010, 54, 614–620. [Google Scholar] [CrossRef]
- Janků, M.; Luhová, L.; Petřivalský, M. On the origin and fate of reactive oxygen species in plant cell compartments. Antioxidants 2019, 8, 105. [Google Scholar] [CrossRef] [Green Version]
- Castro, B.; Citterico, M.; Kimura, S.; Stevens, D.M.; Wrzaczek, M.; Coaker, G. Stress-induced reactive oxygen species compartmentalization, perception and signalling. Nat. Plants 2021, 7, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Amari, T.; Abdelly, C. Biochemical responses of Digitaria commutata and Cenchrus ciliaris to water stress: Antioxidative reactions, proline and soluble sugars accumulation. Bioagro 2021, 33, 171–180. [Google Scholar] [CrossRef]
- Zhang, Y.-P.; Jia, F.-F.; Zhang, X.-M.; Qiao, Y.-X.; Shi, K.; Zhou, Y.-H.; Yu, J.-Q. Temperature effects on the reactive oxygen species formation and antioxidant defence in roots of two cucurbit species with contrasting root zone temperature optima. Acta Physiol. Plant. 2011, 34, 713–720. [Google Scholar] [CrossRef]
- Liang, F.; Pan, Y.J.; Deng, X.; Wu, Y.S.; Liang, Z.R.; Zhao, S.H.; Tian, X.H. Responses of Barringtonia racemosa to Tidal Flooding. Fujian J. Agric. Sci. 2020, 35, 1346–1356. [Google Scholar] [CrossRef]
- de Oliveira, M.M.T.; Lu, S.; Zurgil, U.; Raveh, E.; Tel-Zur, N. Grafting in Hylocereus (Cactaceae) as a tool for strengthening tolerance to high temperature stress. Plant Physiol. Biochem. 2021, 160, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Zhou, J.; Di, B.; Liu, Y.; Zhang, G.; Yang, X. Using electrical impedance tomography for rapid determination of starch and soluble sugar contents in Rosa hybrida. Sci. Rep. 2021, 11, 2871. [Google Scholar] [CrossRef] [PubMed]
- Mohammadrezakhani, S.; Rezanejad, F.; Hajilou, J. Effect of putrescine and proline on proflies of GABA, antioxidant activities in leaves of three Citrus species in response to low temperature stress. J. Plant Biochem. Biotechnol. 2021, 30, 545–553. [Google Scholar] [CrossRef]
- Bao, G.; Tang, W.; An, Q.; Liu, Y.; Tian, J.; Zhao, N.; Zhu, S. Physiological effects of the combined stresses of freezing-thawing, acid precipitation and deicing salt on alfalfa seedlings. BMC Plant Biol. 2020, 20, 104. [Google Scholar] [CrossRef]
- Kalisz, A.; Gil, J.; Kunicki, E.; Sękara, A.; Sałata, A.; Caruso, G. Different temperature regimes influenced the quality of broccoli seedlings, which caused a change in the chemical composition of mature heads. Agronomy 2021, 11, 1806. [Google Scholar] [CrossRef]
- Xu, C. Changes of three osmotic regulatory metabolites contents in leaves of bamboo under low temperature stress. J. Henan Agric. Sci. 2011, 40, 127–130. [Google Scholar] [CrossRef]
- Ok, J.; Kim, S.H.; Ma, K.B.; Kim, D.; Jeong, H.S.; Shin, H. Seaonal fluctuation of freezing tolerance and soluble sugar content in three sweet persimmon cultivars. Hortic. Sci. Technol. 2020, 39, 305–313. [Google Scholar] [CrossRef]
- Das, L.; Patel, R.; Salvi, H.; Kamboj, R. Assessment of natural regeneration of mangrove with reference to edaphic factors and water in Southern Gulf of Kachchh, Gujarat, India. Heliyon 2019, 5, e02250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, M.; Zhang, Q.; Zheng, X.; Zhai, J.; Peng, C. Comparison of leaves and stems of Paederia scandens (Lour.) Merr. in tolerance to low temperature. Photosynthetica 2020, 58, 846–852. [Google Scholar] [CrossRef]
- Zhang, W.; Xie, J.Z. Effects of Low Temperature Stress on Resistance Indices of Sympodial Bamboo Seedlings. Adv. Mater. Res. 2013, 726–731, 4241–4247. [Google Scholar] [CrossRef]
- Shen, Z.-J.; Qin, Y.-Y.; Luo, M.-R.; Li, Z.; Ma, D.-N.; Wang, W.-H.; Zheng, H.-L. Proteome analysis reveals a systematic response of cold-acclimated seedlings of an exotic mangrove plant Sonneratia apetala to chilling stress. J. Proteom. 2021, 248, 104349. [Google Scholar] [CrossRef]
- Miyama, M.; Tada, Y. Transcriptional and physiological study of the response of Burma mangrove (Bruguiera gymnorhiza) to salt and osmotic stress. Plant Mol. Biol. 2008, 68, 119–129. [Google Scholar] [CrossRef]
Figure 1.
Effect of upwelling on MDA and H2O2 content in the leaves of K.obovata. (a) MDA, malondialdehyde activity; (b) H2O2, hydrogen peroxide. Values are presented as mean ± SD, and bars with different letters indicate significant differences (p < 0.05).
Figure 1.
Effect of upwelling on MDA and H2O2 content in the leaves of K.obovata. (a) MDA, malondialdehyde activity; (b) H2O2, hydrogen peroxide. Values are presented as mean ± SD, and bars with different letters indicate significant differences (p < 0.05).

Figure 2.
Effect of upwelling on the proline and soluble sugar content in the leaves of K. obovata. (a) Proline; (b) soluble sugar. Values are presented as mean ± SD, and bars with different letters indicate significant differences (p < 0.05).
Figure 2.
Effect of upwelling on the proline and soluble sugar content in the leaves of K. obovata. (a) Proline; (b) soluble sugar. Values are presented as mean ± SD, and bars with different letters indicate significant differences (p < 0.05).

Figure 3.
Effect of upwelling on SOD, POD, and CAT activity in the leaves of K. obovata. (a) SOD, superoxide dismutase; (b) POD, peroxidase; and (c) CAT, catalase. Values are presented as mean ± SD, and no significant differences were detected in these parameters (p < 0.05).
Figure 3.
Effect of upwelling on SOD, POD, and CAT activity in the leaves of K. obovata. (a) SOD, superoxide dismutase; (b) POD, peroxidase; and (c) CAT, catalase. Values are presented as mean ± SD, and no significant differences were detected in these parameters (p < 0.05).

Figure 4.
Matrix of correlation for response indexes of K. obovata seedlings under upwelling. MDA, malondialdehyde activity; H2O2, hydrogen peroxide; POD, peroxidase; CAT, catalase; SOD, superoxide dismutase; Chl a, chlorophyll a; Chl b, chlorophyll b; Fv/Fm, ratio of variable and maximal fluorescence; Y(Ⅱ), effective quantum yield of photosystem Ⅱ; ETR, electron transport rate; qP, photochemical quenching coefficient; qN, non-photochemical quenching coefficient.
Figure 4.
Matrix of correlation for response indexes of K. obovata seedlings under upwelling. MDA, malondialdehyde activity; H2O2, hydrogen peroxide; POD, peroxidase; CAT, catalase; SOD, superoxide dismutase; Chl a, chlorophyll a; Chl b, chlorophyll b; Fv/Fm, ratio of variable and maximal fluorescence; Y(Ⅱ), effective quantum yield of photosystem Ⅱ; ETR, electron transport rate; qP, photochemical quenching coefficient; qN, non-photochemical quenching coefficient.

Figure 5.
Principal component analysis biplot for PC1 and PC2. MDA, malondialdehyde activity; H2O2, hydrogen peroxide; POD, peroxidase; CAT, catalase; SOD, superoxide dismutase; Chl a, chlorophyll a; Chl b, chlorophyll b; Fv/Fm, ratio of variable and maximal fluorescence; Y(Ⅱ), effective quantum yield of photosystem Ⅱ; ETR, electron transport rate; qP, photochemical quenching coefficient; qN, non-photochemical quenching coefficient.
Figure 5.
Principal component analysis biplot for PC1 and PC2. MDA, malondialdehyde activity; H2O2, hydrogen peroxide; POD, peroxidase; CAT, catalase; SOD, superoxide dismutase; Chl a, chlorophyll a; Chl b, chlorophyll b; Fv/Fm, ratio of variable and maximal fluorescence; Y(Ⅱ), effective quantum yield of photosystem Ⅱ; ETR, electron transport rate; qP, photochemical quenching coefficient; qN, non-photochemical quenching coefficient.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effect of upwelling on chlorophyll contents and fluorescence parameters in the leaves of K. obovata.
Table 1.
Effect of upwelling on chlorophyll contents and fluorescence parameters in the leaves of K. obovata.
| Control | Treated | |||
|---|---|---|---|---|
| 25 °C | 13 °C | 5 °C | p-Value | |
| Photosynthetic pigments, mg/L | ||||
| Chlorophyll a | 7.458 ± 1.045 | 8.358 ± 1.769 | 7.611 ± 1.817 | 0.767 |
| Chlorophyll b | 2.902 ± 0.613 | 3.179 ± 0.584 | 2.856 ± 0.545 | 0.772 |
| Photosynthetic fluorescence parameters | ||||
| Y(Ⅱ) | 0.387 ± 0.047 | 0.348 ± 0.041 | 0.324 ± 0.021 | 0.197 |
| ETR | 70.233 ± 8.451 | 63 ± 7.375 | 58.633 ± 3.88 | 0.193 |
| qP | 0.542 ± 0.061 | 0.542 ± 0.066 | 0.504 ± 0.054 | 0.692 |
| qN | 0.459 ± 0.02 b | 0.659 ± 0.017 a | 0.678 ± 0.041 a | 0.000 |
| Fv/Fm | 0.833 ± 0.003 | 0.842 ± 0.003 | 0.845 ± 0.011 | 0.120 |
Notes: Values are presented as mean ± SD, and bars with different letters indicate significant differences (p < 0.05). Chl a, chlorophyll a; Chl b, chlorophyll b; Fv/Fm, ratio of variable and maximal fluorescence; Y(Ⅱ), effective quantum yield of photosystem Ⅱ; ETR, electron transport rate; qP, photochemical quenching coefficient; qN, non-photochemical quenching coefficient.
Table 2.
Principal component analysis eigenvalues for response indexes of K. obovata seedlings under upwelling.
Table 2.
Principal component analysis eigenvalues for response indexes of K. obovata seedlings under upwelling.
| PC1 | PC2 | PC3 | |
|---|---|---|---|
| Eigenvalue | 6.700 | 2.541 | 1.875 |
| Proportion % | 47.86 | 18.15 | 13.39 |
| Cumulative % | 47.86 | 66.01 | 79.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, X.; Wang, Y.; Dong, J.; Wu, M. Physiological and Biochemical Responses of Kandelia obovata to Upwelling Stress. Water 2022, 14, 899. https://doi.org/10.3390/w14060899
AMA Style
Li X, Wang Y, Dong J, Wu M. Physiological and Biochemical Responses of Kandelia obovata to Upwelling Stress. Water. 2022; 14(6):899. https://doi.org/10.3390/w14060899
Chicago/Turabian StyleLi, Xiaomei, Youshao Wang, Junde Dong, and Meilin Wu. 2022. "Physiological and Biochemical Responses of Kandelia obovata to Upwelling Stress" Water 14, no. 6: 899. https://doi.org/10.3390/w14060899
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.