
Seasonal Dynamics of Algal Net Primary Production in Response to Phosphorus Input in a Mesotrophic Subtropical Plateau Lake, Southwestern China

Abstract
:1. Introduction
2. Materials and Methods
2.1. EFDC Model Description
2.1.1. Phytoplankton Kinetic
2.1.2. Nutrient Limitation
2.1.3. Temperature Limitation
2.1.4. Basal Metabolism and Predation
2.1.5. Calculation of NPP
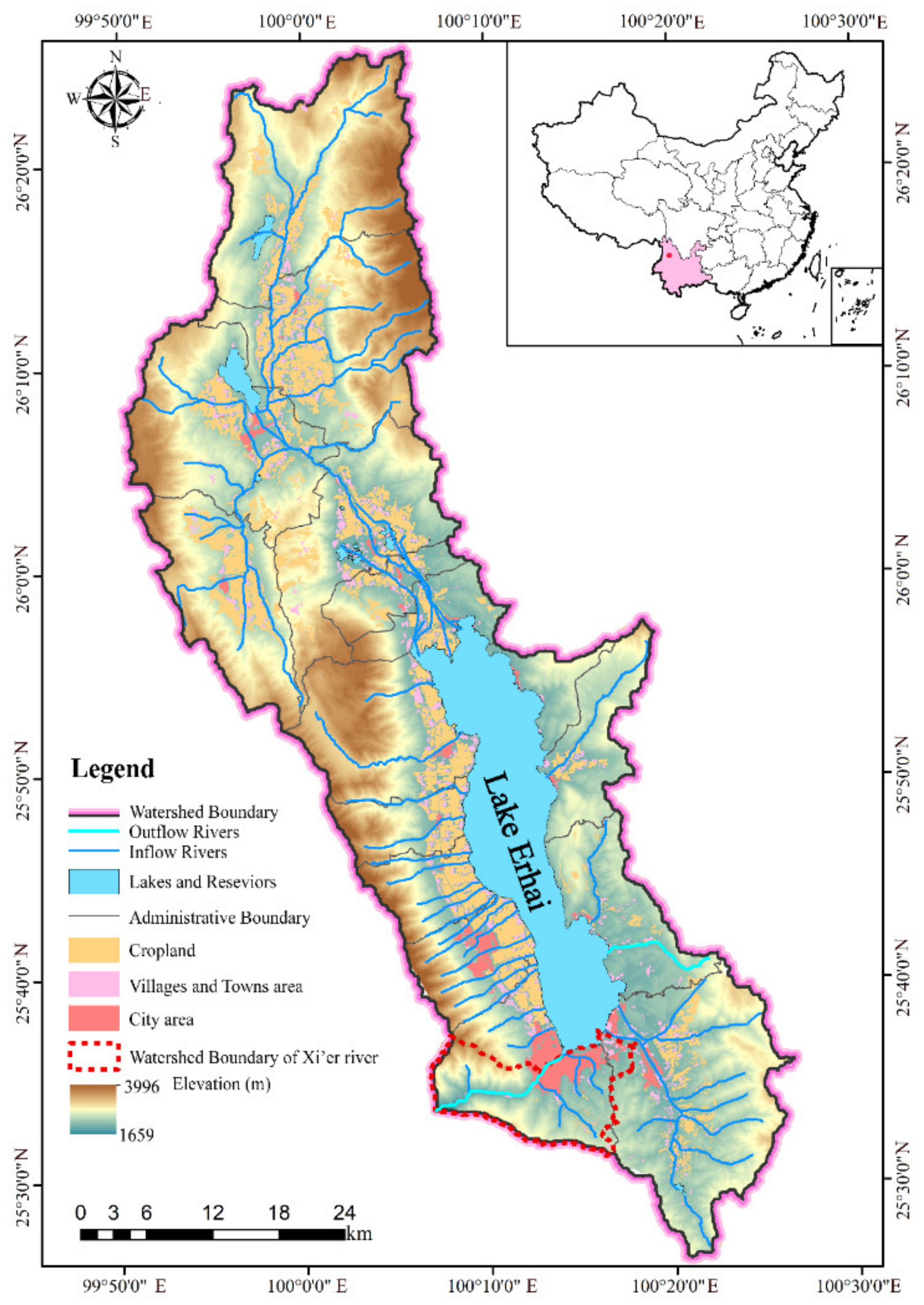
2.2. Study Site
2.3. Sample Collection and Analysis
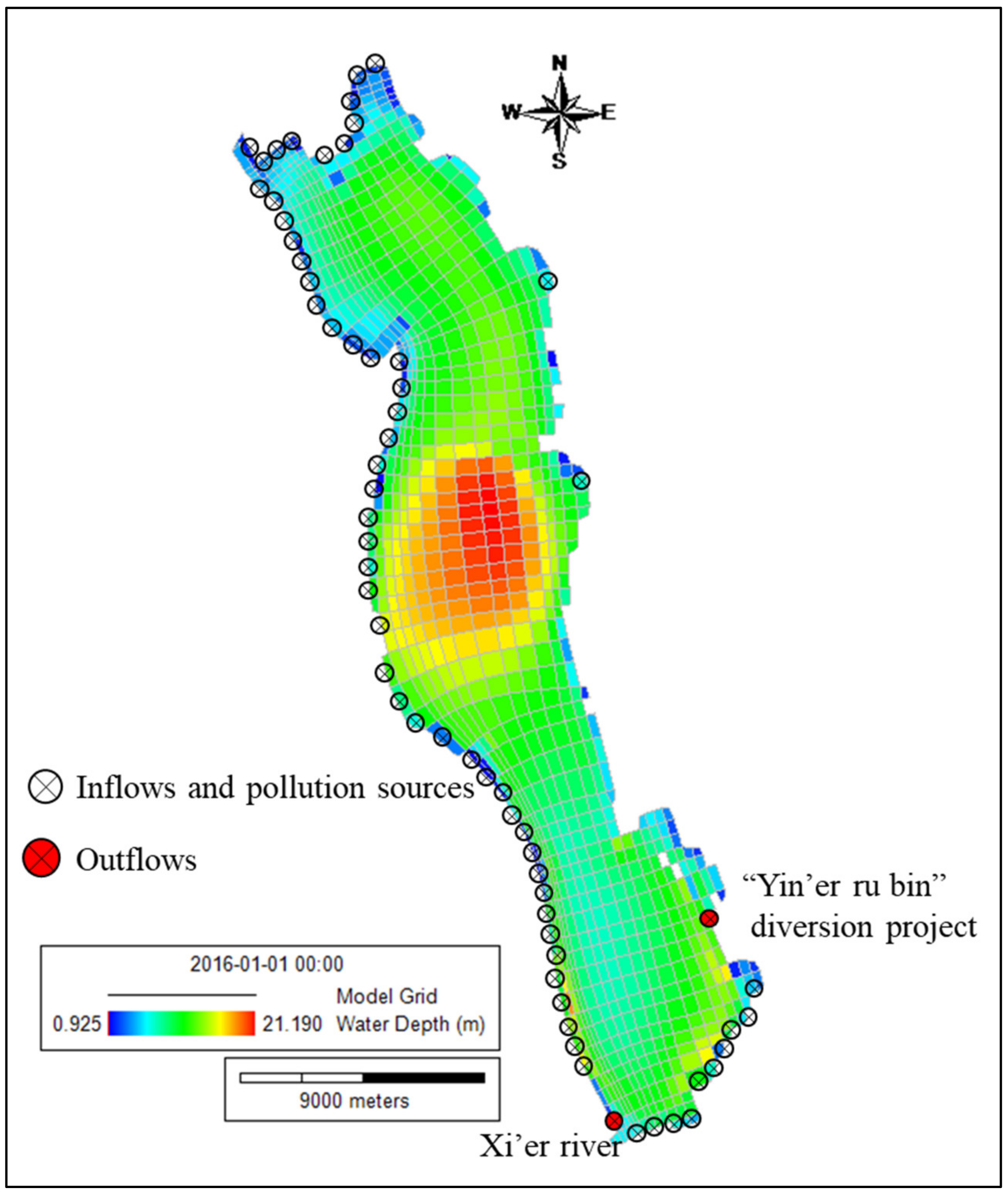
2.4. Model Setup
2.4.1. Grid Generation
2.4.2. Boundary Condition and Nutrient Input
2.4.3. Model Calibration and Validation
2.5. Sensitivity Analysis
2.6. Statistical Analysis
3. Results and Discussion
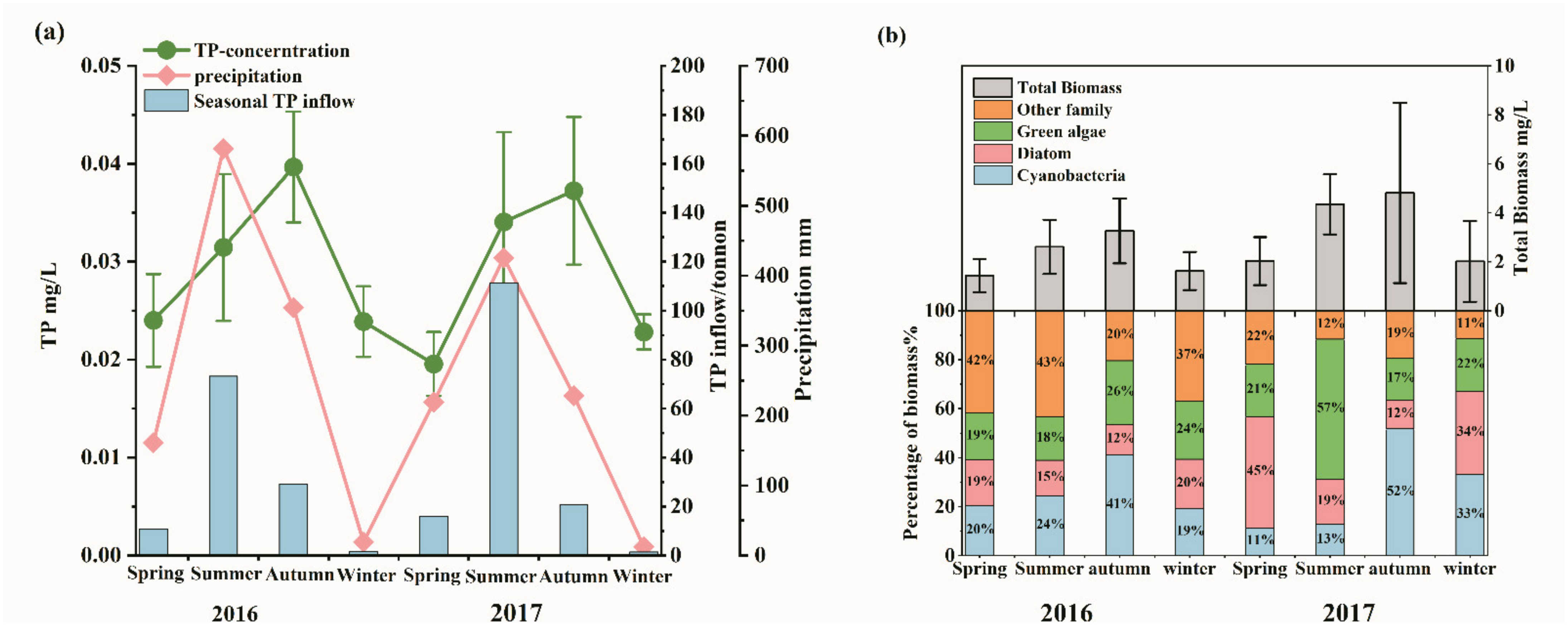
3.1. Seasonal Variation of Phosphorus Load and Algal Biomass
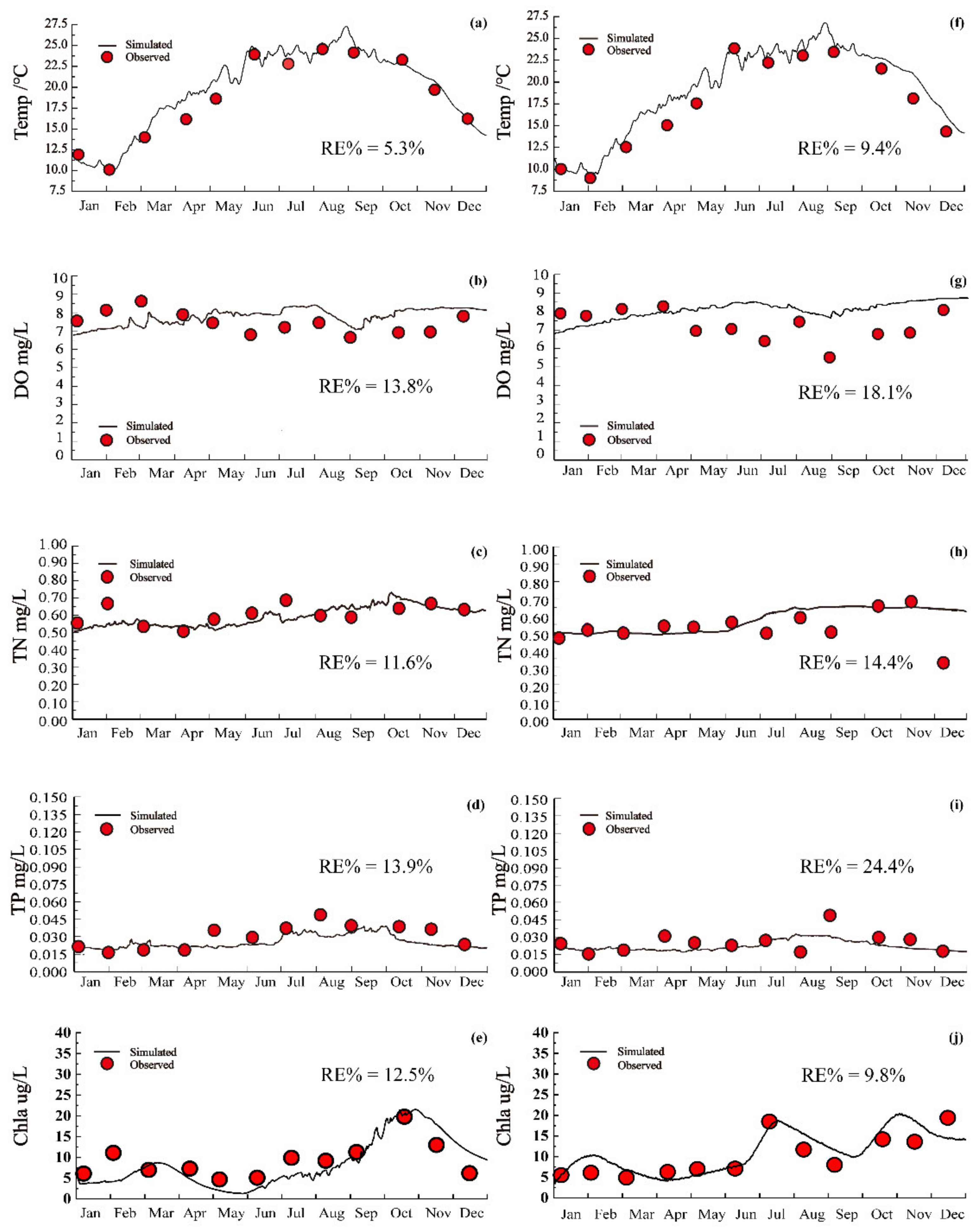
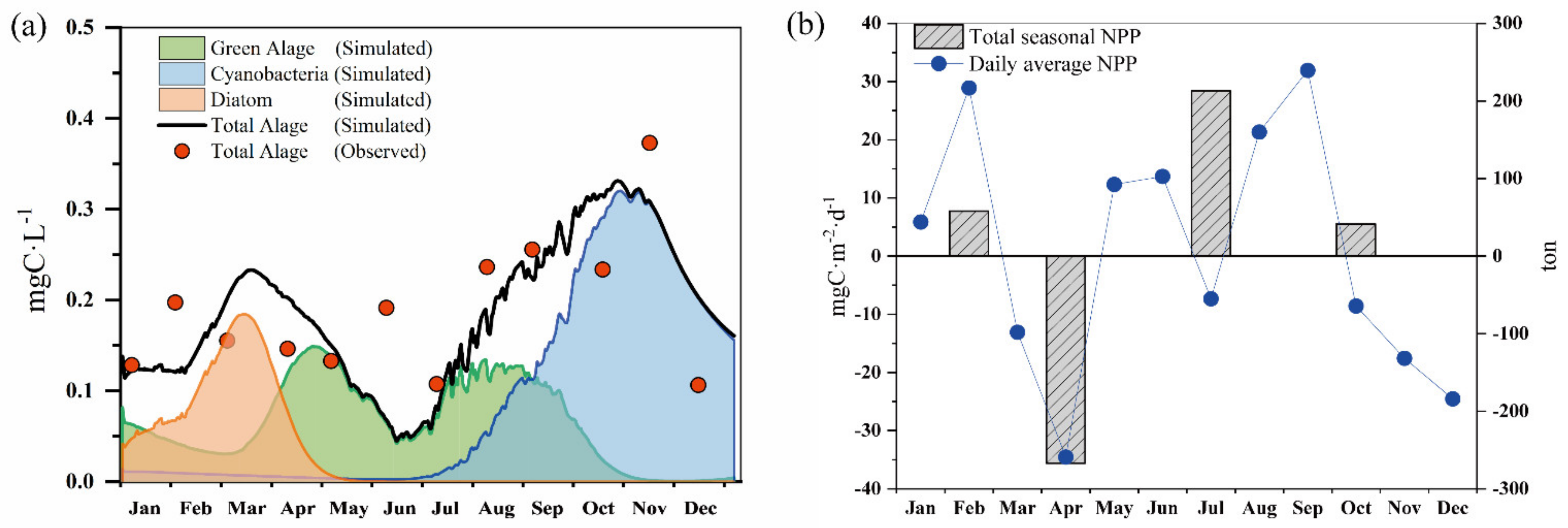
3.2. Model Results with Calibration and Validation
3.3. The Seasonal Dynamic of Phytoplankton Biomass and Net Primary Production
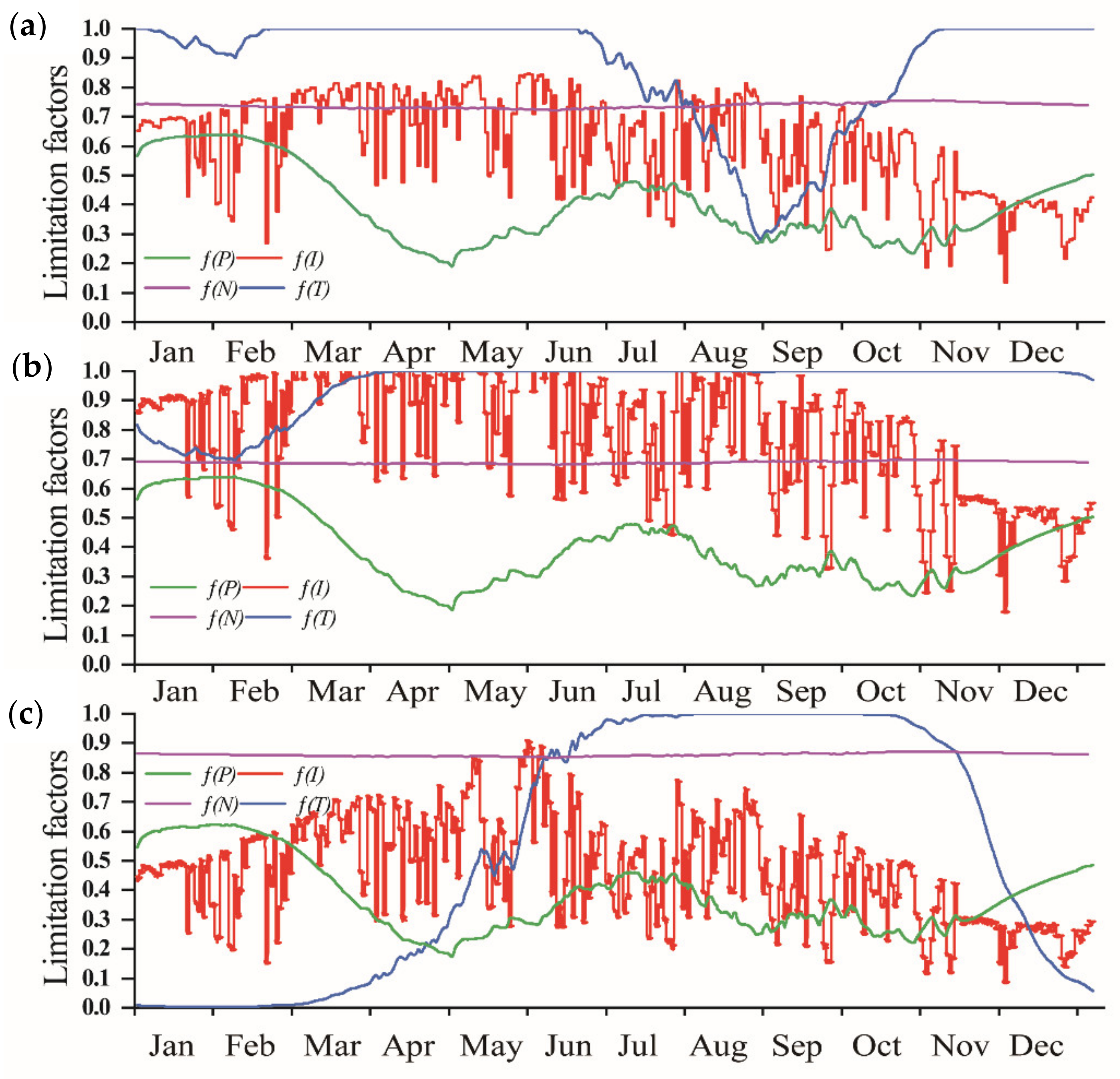
3.4. Limiting Factors of Algal Growth in Erhai
3.5. Model Sensitivity
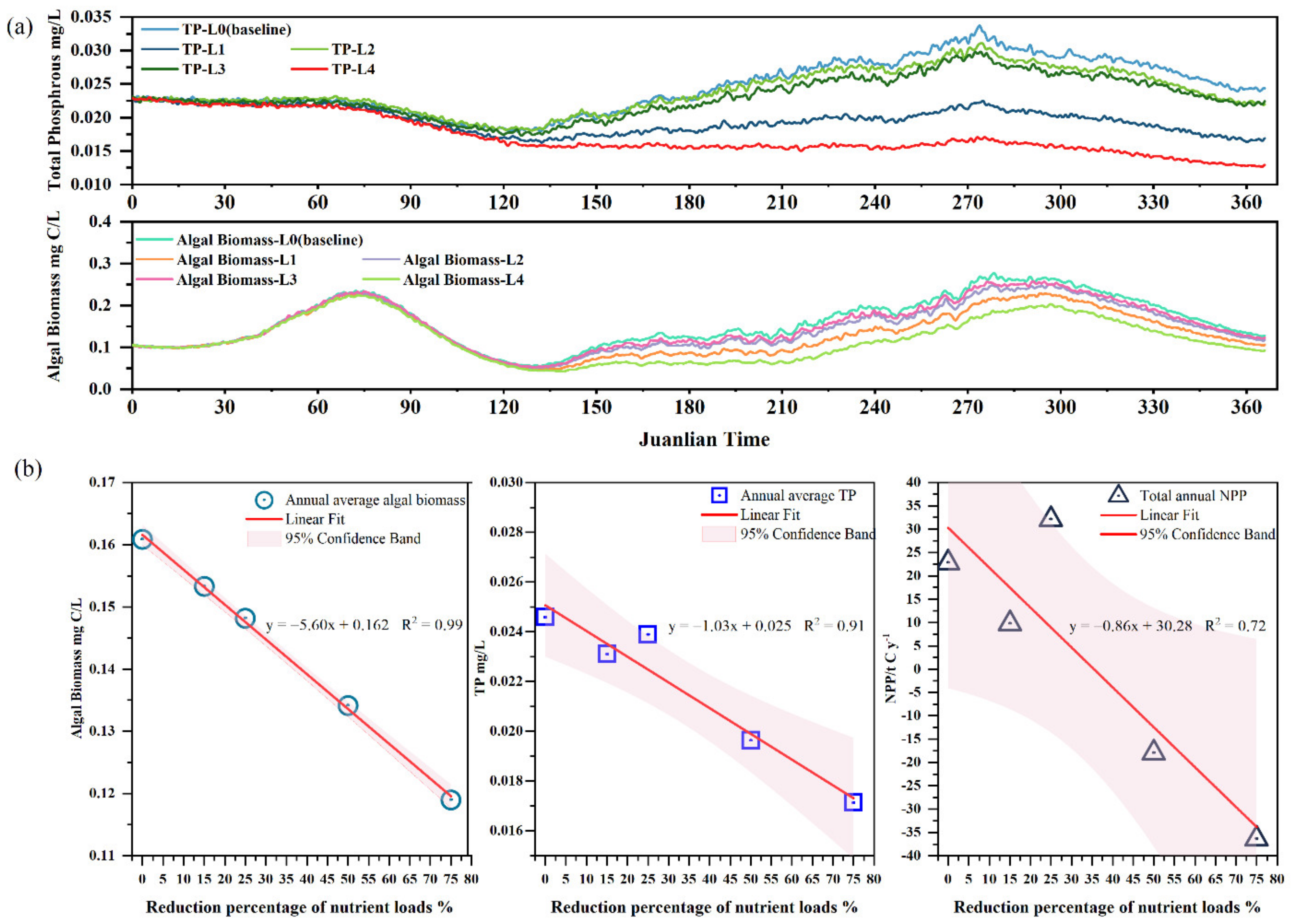
3.6. Response of NPP to the Phosphorous Load Reduction
4. Conclusions
- A 3D hydrodynamic and water quality model was developed for Lake Erhai, China. In the field investigation, the water surface elevation, temperature, nutrients and algal biomass concentration in the lake were accurately simulated by the numerical model, revealing a reasonable picture of the lake’s hydrodynamics and eutrophication process. The model is suitable to calculate the seasonal NPP in the lake.
- According to the modeling results, the average daily NPP ranged from −3.45 and 3.19 mg C/m2/d. The seasonal fluctuation in total NPP indicates that Lake Erhai is net heterotrophic in spring and net autotrophic in summer, autumn, and winter. The difference in seasonal NPP can be attributed to the combined impact of phosphorus supply and temperature limitation of the phytoplankton community within the year.
- The results of loading reduction scenarios indicate that phosphorus loading reduction directly affects the water quality of Lake Erhai, and the efficiency of improvement in TP concentration and NPP is proportionate to the degree of load decrease.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Unit | Description | Calibrated Value | Range |
|---|---|---|---|---|
| Pc | 1/day | Maximum growth Rate for Cyanobacteria | 1.35 | 0.2–9.0 |
| Pd | 1/day | Maximum growth Rate for Diatoms | 0.95 | 0.2–9.0 |
| Pg | 1/day | Maximum growth Rate for Greens | 1.7 | 0.2–9.0 |
| BMc | 1/day | Basal Metabolism Rate for Cyanobacteria | 0.013 | 0.01–0.92 |
| BMd | 1/day | Basal Metabolism Rate for Diatoms | 0.15 | 0.01–0.92 |
| BMg | 1/day | Basal Metabolism Rate for Greens | 0.13 | 0.01–0.92 |
| PRc | 1/day | Predation Rate on Cyanobacteria | 0.01 | 0.01–0.06 |
| PRd | 1/day | Predation Rate on Diatoms | 0.155 | 0.03–0.3 |
| PRg | 1/day | Predation Rate on Greens | 0.14 | 0.03–0.3 |
| Keb | 1/m | Background Light Extinction Coefficient | 0.38 | 0.25–0.45 |
| Sc | m/day | Settling velocity for Cyanobacteria | 0.02 | 0.01–0.3 |
| Sd | m/day | Settling velocity for Diatoms | 0.12 | 0.01–0.3 |
| Sg | m/day | Settling velocity for Greens | 0.1 | 0.01–0.3 |
| SRP | m/day | Settling velocity for refractory particulate organic matter (RPOM) | 0.05 | 0.02–9.0 |
| SLP | m/day | Settling velocity for liable particulate organic matter (LPOM) | 0.01 | 0.02–9.0 |
| CChlc | mg C/ug Chl | C:chlorophyll ratio for Cyanobacteria | 0.02 | 0.01–0.05 |
| CChld | mg C/ug Chl | C:chlorophyll ratio for Algae:Diatoms | 0.033 | 0.01–0.05 |
| CChlg | mg C/ug Chl | C:chlorophyll ratio for Algae:Greens | 0.033 | 0.01–0.05 |
| CPprm1 | Constant1 used in Determining Algae C:P Ratio (gC/gP) | 45 | 30–65 | |
| Keb | 1/m | Background Light Extinction Coefficient | 0.38 | 0.15–0.45 |
| KHNc | mg/L | Nitrogen Half-Saturation for Cyanobacteria | 0.045 | 0.01–0.25 |
| KHNd | mg/L | Nitrogen Half-Saturation for Algae:Diatoms | 0.05 | 0.01–0.25 |
| KHNg | mg/L | Nitrogen Half-Saturation for Algae:Greens | 0.1 | 0.01–0.25 |
| KHPc | mg/L | Phosphorus Half-Saturation for Cyanobacteria | 0.0033 | 0.001–0.05 |
| KHPd | mg/L | Phosphorus Half-Saturation for Algae:Diatoms | 0.0043 | 0.001–0.05 |
| KHPg | mg/L | Phosphorus Half-Saturation for Algae:Greens | 0.0035 | 0.001–0.05 |
| TMc1 | °C | Lower Optimal Temperature for Growth, Cyanobacteria | 23 | 20–25 |
| TMc2 | °C | Upper Optimal Temperature for Growth, Cyanobacteria | 30 | 25–30 |
| TMd1 | °C | Lower Optimal Temperature for Growth, Diatoms | 10 | 10–15 |
| TMd2 | °C | Upper Optimal Temperature for Growth, Diatoms | 22 | 10–15 |
| TMg1 | °C | Lower Optimal Temperature for Growth, Greens | 20 | 22–25 |
| TMg2 | °C | Upper Optimal Temperature for Growth, Greens | 25 | 22–26 |
| TMp1 | °C | Lower Optimal Temperature for Predation, Diatoms | 15 | 15–25 |
| TMp2 | °C | Upper Optimal Temperature for Predation, Diatoms | 20 | 15–26 |
| KTG1c | Suboptimal Temperature Effect Coeff for Growth, Cyanobacteria | 0.2 | 0.001–0.01 | |
| KTG2c | Superoptimal Temperature Effect Coeff for Growth, Cyanobacteria | 0.03 | 0.001–0.01 | |
| KTG1d | Suboptimal Temperature Effect Coeff for Growth, Diatoms | 0.069 | ||
| KTG2d | Superoptimal Temperature Effect Coeff for Growth, Diatoms | 0.1 | ||
| KTG1g | Suboptimal Temperature Effect Coeff for Growth, Greens | 0.1 | ||
| KTG2g | Superoptimal Temperature Effect Coeff for Growth, Greens | 0.01 | ||
| FNRc | Fraction of Metabolized Nitrogen Produced as RPON, Cyanobacteria | 0.15 | ||
| FNRd | Fraction of Metabolized Nitrogen Produced as RPON, Diatoms | 0.15 | ||
| FNRg | Fraction of Metabolized Nitrogen Produced as RPON, Greens | 0.15 | ||
| FNLc | Fraction of Metabolized Nitrogen Produced as LPON, Cyanobacteria | 0.25 | ||
| FNLd | Fraction of Metabolized Nitrogen Produced as LPON, Diatoms | 0.25 | ||
| FNLg | Fraction of Metabolized Nitrogen Produced as LPON, Greens | 0.25 | ||
| FNDc | Fraction of Metabolized Nitrogen Produced as DON, Cyanobacteria | 0.5 | ||
| FNDd | Fraction of Metabolized Nitrogen Produced as DON, Diatoms | 0.5 | ||
| FNDg | Fraction of Metabolized Nitrogen Produced as DON, Greens | 0.5 | ||
| FNIc | Fraction of Metabolized Nitrogen Produced as DIN, Cyanobacteria | 0.1 | ||
| FNId | Fraction of Metabolized Nitrogen Produced as DIN, Diatoms | 0.1 | ||
| FNIg | Fraction of Metabolized Nitrogen Produced as DIN, Greens | 0.1 | ||
| FPRc | Fraction of Metabolized Phosphorus Produced as RPOP, Cyanobacteria | 0 | ||
| FPRd | Fraction of Metabolized Phosphorus Produced as RPOP, Diatoms | 0 | ||
| FPRg | Fraction of Metabolized Phosphorus Produced as RPOP, Greens | 0 | ||
| FPLc | Fraction of Metabolized Phosphorus Produced as LPOP, Cyanobacteria | 0 | ||
| FPLd | Fraction of Metabolized Phosphorus Produced as LPOP, Diatoms | 0 | ||
| FPLg | Fraction of Metabolized Phosphorus Produced as LPOP, Greens | 0 | ||
| FPDc | Fraction of Metabolized Phosphorus Produced as DOP, Cyanobacteria | 0.95 | ||
| FPDd | Fraction of Metabolized Phosphorus Produced as DOP, Diatoms | 0.9 | ||
| FPDg | Fraction of Metabolized Phosphorus Produced as DOP, Greens | 0.9 | ||
| FPIc | Fraction of Metabolized Phosphorus Produced as P4T, Cyanobacteria | 0.05 | ||
| FPId | Fraction of Metabolized Phosphorus Produced as P4T, Diatoms | 0.1 | ||
| FPIg | Fraction of Metabolized Phosphorus Produced as P4T, Greens | 0.1 | ||
| KRP | 1/day | Minimum Hydrolysis Rate of RPOP | 0.005 | 0.001–0.01 |
| KLP | 1/day | Minimum Hydrolysis Rate of LPOP | 0.005 | 0.01–0.1 |
| KDP | 1/day | Minimum Mineralization Rate of DOP | 0.008 | 0.01–0.3 |
| KRPALG | m3(gc)/d | Constant relating Hydrolysis Rate of RPOP to Algae: | 0.005 | |
| KLPALG | m3(gc)/d | Constant relating Hydrolysis Rate of LPOP to Algae: | 0.01 | |
| KDPALG | m3(gc)/d | Constant relating Mineralization Rate of DOP to Algae: | 0.01 | |
| KRN | 1/day | Minimum Hydrolysis Rate of RPON | 0.005 | 0.001–0.01 |
| KLN | 1/day | Minimum Hydrolysis Rate of LPON | 0.0075 | 0.01–0.1 |
| KDN | 1/day | Minimum Mineralization Rate of DON | 0.01 | 0.01–0.08 |
| KRNALG | m3(gc)−1d−1 | Constant relating Hydrolysis Rate of RPON to Algae | 0.005 | 0.0001–0.01 |
| KLNALG | m3(gc)−1d−1 | Constant relating Hydrolysis Rate of LPON to Algae | 0.005 | 0.0001–0.02 |
| KDNALG | m3(gc)−1d−1 | Constant relating Mineralization Rate of DON to Algae | 0.01 | 0.001–0.01 |
| KHORDO | gO2/m3 | oxygen Half-Sat Constant for Algal Respiration | 1 | 0.5–2 |
| KHDNN | gN/m3 | half-sat. constant for denitrification | 0.001 | 0.05–0.2 |
| KHCOD | mg/L O2 | oxygen half-saturation constant for COD decay | 0.7667 | 1–1.5 |
References
- Pomati, F.; Matthews, B.; Seehausen, O.; Ibelings, B.W. Eutrophication and Climate Warming Alter Spatial (Depth) Co-Occurrence Patterns of Lake Phytoplankton Assemblages. Hydrobiologia 2017, 787, 375–385. [Google Scholar] [CrossRef]
- Wells, M.L.; Trainer, V.L.; Smayda, T.J.; Karlson, B.S.O.; Trick, C.G.; Kudela, R.M.; Ishikawa, A.; Bernard, S.; Wulff, A.; Anderson, D.M.; et al. Harmful Algal Blooms and Climate Change: Learning from the Past and Present to Forecast the Future. Harmful Algae 2015, 49, 68–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Moal, M.; Gascuel-Odoux, C.; Ménesguen, A.; Souchon, Y.; Étrillard, C.; Levain, A.; Moatar, F.; Pannard, A.; Souchu, P.; Lefebvre, A.; et al. Eutrophication: A New Wine in an Old Bottle? Sci. Total Environ. 2019, 651, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Goyette, J.O.; Bennett, E.M.; Maranger, R. Low Buffering Capacity and Slow Recovery of Anthropogenic Phosphorus Pollution in Watersheds. Nat. Geosci. 2018, 11, 921–925. [Google Scholar] [CrossRef]
- Schurz, C.; Hollosi, B.; Matulla, C.; Pressl, A.; Ertl, T.; Schulz, K.; Mehdi, B. A Comprehensive Sensitivity and Uncertainty Analysis for Discharge and Nitrate-Nitrogen Loads Involving Multiple Discrete Model Inputs under Future Changing Conditions. Hydrol. Earth Syst. Sci. 2019, 23, 1211–1244. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Zhang, W.; Wang, X.; Couture, R.M.; Larssen, T.; Zhao, Y.; Li, J.; Liang, H.; Liu, X.; Bu, X.; et al. Decline in Chinese Lake Phosphorus Concentration Accompanied by Shift in Sources since 2006. Nat. Geosci. 2017, 10, 507–511. [Google Scholar] [CrossRef]
- Gao, Y.; Jia, J.; Lu, Y.; Yang, T.; Lyu, S.; Shi, K.; Zhou, F.; Yu, G. Determining Dominating Control Mechanisms of Inland Water Carbon Cycling Processes and Associated Gross Primary Productivity on Regional and Global Scales. Earth Sci. Rev. 2021, 213, 103497. [Google Scholar] [CrossRef]
- López-Sandoval, D.C.; Duarte, C.M.; Agustí, S. Nutrient and Temperature Constraints on Primary Production and Net Phytoplankton Growth in a Tropical Ecosystem. Limnol. Oceanogr. 2021, 66, 2923–2935. [Google Scholar] [CrossRef]
- Trochine, C.; Guerrieri, M.E.; Liboriussen, L.; Lauridsen, T.L.; Jeppesen, E. Effects of Nutrient Loading, Temperature Regime and Grazing Pressure on Nutrient Limitation of Periphyton in Experimental Ponds. Freshw. Biol. 2014, 59, 905–917. [Google Scholar] [CrossRef]
- Val, J.; Pino, R.; Navarro, E.; Chinarro, D. Addressing the Local Aspects of Global Change Impacts on Stream Metabolism Using Frequency Analysis Tools. Sci. Total Environ. 2016, 569–570, 798–814. [Google Scholar] [CrossRef]
- Genzoli, L.; Hall, R.O., Jr. Shifts in Klamath River Metabolism Following a Reservoir Cyanobacterial Bloom. Freshw. Sci. 2016, 35, 795–809. [Google Scholar] [CrossRef]
- Kuehl, L.C.; Troelstrup, N.H., Jr. Relationships between Net Primary Reproduction, Water Transparency, Chlorophyll a, and Total Phosphorous in Oak Lake, Brookings County, South Dakota. Proc. S. D. Acad. Sci. 2013, 92, 67–78. [Google Scholar]
- Kemp, W.M.; Smith, E.M.; Marvin-DiPasquale, M.; Boynton, W.R. Organic Carbon Balance and Net Ecosystem Metabolism in Chesapeake Bay. Mar. Ecol. Prog. Ser. 1997, 150, 229–248. [Google Scholar] [CrossRef]
- Gazeau, F.; Borges, A.V.; Barrón, C.; Duarte, C.M.; Iversen, N.; Middelburg, J.J.; Delille, B.; Pizay, M.D.; Frankignoulle, M.; Gattuso, J.P. Net Ecosystem Metabolism in a Micro-Tidal Estuary (Randers Fjord, Denmark): Evaluation of Methods. Mar. Ecol. Prog. Ser. 2005, 301, 23–41. [Google Scholar] [CrossRef]
- Staehr, P.A.; Bade, D.; van de Bogert, M.C.; Koch, G.R.; Williamson, C.; Hanson, P.; Cole, J.J.; Kratz, T. Lake Metabolism and the Diel Oxygen Technique: State of the Science. Limnol. Oceanogr. Methods 2010, 8, 628–644. [Google Scholar] [CrossRef]
- Gelda, R.K.; Effler, S.W. Metabolic Rate Estimates for a Eutrophic Lake from Diel Dissolved Oxygen Signals. Hydrobiologia 2002, 485, 51–66. [Google Scholar] [CrossRef]
- Caffrey, J.M. Production, Respiration and Net Ecosystem Metabolism U.S. Estuaries. Environ. Monit. Assess. 2003, 81, 207–219. [Google Scholar] [CrossRef]
- Quay, P.D.; Wilbur, D.; Richey, J.E.; Devol, A.H.; Benner, R.; Forsberg, B.R. The 18O:16O of Dissolved Oxygen in Rivers and Lakes in the Amazon Basin: Determining the Ratio of Respiration to Photosynthesis Rates in Freshwaters. Limnol. Oceanogr. 1995, 40, 718–729. [Google Scholar] [CrossRef]
- Tobias, C.R.; Böhlke, J.K.; Harvey, J.W. The Oxygen-18 Isotope Approach for Measuring Aquatic Metabolism in High-Productivity Waters. Limnol. Oceanogr. 2007, 52, 1439–1453. [Google Scholar] [CrossRef]
- Hamilton, D.P.; Schladow, S.G. Prediction of Water Quality in Lakes and Reservoirs. Part I—Model Description. Ecol. Model. 1997, 96, 91–110. [Google Scholar] [CrossRef]
- Schladow, S.G.; Hamilton, D.P. Prediction of Water Quality in Lakes and Reservoirs: Part II—Model Calibration, Sensitivity Analysis and Application. Ecol. Model. 1997, 96, 111–123. [Google Scholar] [CrossRef]
- Riley, M.J.; Stefan, H.G. Development of the Minnesota Lake Water Quality Management Model “Minlake”. Lake Reserv. Manag. 1988, 4, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Cloern, J.E. The Relative Importance of Light and Nutrient Limitation of Phytoplankton Growth: A Simple Index of Coastal Ecosystem Sensitivity to Nutrient Enrichment. Aquat. Ecol. 1999, 33, 3–15. [Google Scholar] [CrossRef]
- Mortazavi, B.; Iverson, R.L.; Landing, W.M.; Lewis, F.G.; Huang, W. Control of Phytoplankton Production and Biomass in a River-Dominated Estuary: Apalachicola Bay, Florida, USA. Mar. Ecol. Prog. Ser. 2000, 198, 19–31. [Google Scholar] [CrossRef]
- Muylaert, K.; Tackx, M.; Vyverman, W. Phytoplankton Growth Rates in the Freshwater Tidal Reaches of the Schelde Estuary (Belgium) Estimated Using a Simple Light-Limited Primary Production Model. Hydrobiologia 2005, 540, 127–140. [Google Scholar] [CrossRef]
- Pinckney, J.L.; Paerl, H.W.; Harrington, M.B. Responses of the Phytoplankton Community Growth Rate to Nutrient Pulses in Variable Estuarine Environments. J. Phycol. 1999, 35, 1455–1463. [Google Scholar] [CrossRef]
- Strock, J.P.; Menden-Deuer, S. Temperature Acclimation Alters Phytoplankton Growth and Production Rates. Limnol. Oceanogr. 2021, 66, 740–752. [Google Scholar] [CrossRef]
- Klausmeier, C.A.; Litchman, E.; Levin, S.A. Phytoplankton Growth and Stoichiometry under Multiple Nutrient Limitation. Limnol. Oceanogr. 2004, 49, 1463–1470. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, I.C.; Bordalo, A.A.; Duarte, P.M. Influence of River Discharge Patterns on the Hydrodynamics and Potential Contaminant Dispersion in the Douro Estuary (Portugal). Water Res. 2010, 44, 3133–3146. [Google Scholar] [CrossRef]
- Azevedo, I.C.; Bordalo, A.A.; Duarte, P. Influence of Freshwater Inflow Variability on the Douro Estuary Primary Productivity: A Modelling Study. Ecol. Model. 2014, 272, 1–15. [Google Scholar] [CrossRef]
- Yan, J.; Liu, J.; You, X.; Shi, X.; Zhang, L. Simulating the Gross Primary Production and Ecosystem Respiration of Estuarine Ecosystem in North China with AQUATOX. Ecol. Model. 2018, 373, 1–12. [Google Scholar] [CrossRef]
- Hamrick, J.M. A Three-Dimensional Environmental Fluid Dynamics Computer Code: Theoretical and Computational Aspects; Virginia Institute of Marine Science: Gloucester Point, VA, USA, 1992. [Google Scholar]
- Qin, Q.; Shen, J. The Contribution of Local and Transport Processes to Phytoplankton Biomass Variability over Different Timescales in the Upper James River, Virginia. Estuar. Coast. Shelf Sci. 2017, 196, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Camacho, R.A.; Martin, J.L.; Watson, B.; Paul, M.J.; Zheng, L.; Stribling, J.B. Modeling the Factors Controlling Phytoplankton in the St. Louis Bay Estuary, Mississippi and Evaluating Estuarine Responses to Nutrient Load Modifications. J. Environ. Eng. 2015, 141, 04014067. [Google Scholar] [CrossRef]
- Rong, Z.; Huan, W.; Dezhao, Y.; Cheng, Z.; Hong, S.; Jun, C. Dynamic Changes of Microcystins and Phytoplankton during the Cyanobacterial Bloom in Lake Erhai in 2013. J. Lake Sci. 2015, 27, 378–384. [Google Scholar] [CrossRef]
- Turner, M.A.; Sigurdson, L.J.; Findlay, D.L.; Howell, E.T.; Robinson, G.G.C.; Brewster, J.F. Growth Characteristics of Bloom-Forming Filamentous Green Algae in the Littoral Zone of an Experimentally Acidified Lake. Can. J. Fish. Aquat. Sci. 1995, 52, 2251–2263. [Google Scholar] [CrossRef]
- Xiao, M.; Willis, A.; Burford, M.A. Differences in Cyanobacterial Strain Responses to Light and Temperature Reflect Species Plasticity. Harmful Algae 2017, 62, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Lu, X.; Chen, Y. The Effects of Temperature and Nutrient Ratios on Microcystis Blooms in Lake Taihu, China: An 11-Year Investigation. Harmful Algae 2011, 10, 337–343. [Google Scholar] [CrossRef]
- Cerco, C.F.; Cole, T.M. Three-Dimensional Eutrophication Model of Chesapeake Bay. Volume 1: Main Report; Final Report; FAO: Rome, Italy, 2006. [Google Scholar]
- Park, K.; Kuo, A.Y.; Shen, J.; Hamrick, J.M. A Three-Dimensional Hydrodyamic-Eutrophication Model (HEM-3D): Description of Water Quality and Sediment Process Submodels (EFDC Water Quality Model); William & Mary: Williamsburg, VA, USA, 1995; p. 327. [Google Scholar]
- Odum, H.T. Primary Production in Flowing Waters. Limnol. Oceanogr. 1956, 1, 102–117. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, L.; Ni, L.; Zhao, H.; Jiao, L.; Yang, S.; Guo, L.; Shen, J. Ecological Degeneration of the Erhai Lake and Prevention Measures. Environ. Earth Sci. 2015, 74, 3839–3847. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, K.; Wang, S.; Wang, S.; Li, Y.; Li, Q.; Meng, Z. Characteristics of Dissolved Organic Nitrogen in Overlying Water of Typical Lakes of Yunnan Plateau. China. Ecol. Indic. 2017, 84, 727–737. [Google Scholar] [CrossRef]
- American Public Health Association (APHA). Standard Methods for the Examination of Water and Wastewater, 21st ed.; APHA: Washington, DC, USA, 2005. [Google Scholar]
- Hasle, G.R.; Sournia, A. Phytoplankton Manual, Monographs on Oceanographic Methodology. In Concentrating Phytoplankton. Settling. The Inverted Microscope Method; Unesco: Paris, France, 1978. [Google Scholar]
- Hu, H. The Freshwater Algae of China: Systematics, Taxonomy and Ecology; Science Press: Beijing, China, 2006; ISBN 7030166337. [Google Scholar]
- Sun, J.; Liu, D. Study on Phytoplankton Biomass I. Phytoplankton Measurement Biomass from Cell Volume or Plasma Volume. Acta Oceanol. Sin. 1999, 21, 75–85. [Google Scholar]
- Reynolds, C.S. The Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006; ISBN 1139454897. [Google Scholar]
- Spears, B.M.; Carvalho, L.; Perkins, R.; Kirika, A.; Paterson, D.M. Sediment Phosphorus Cycling in a Large Shallow Lake: Spatio-Temporal Variation in Phosphorus Pools and Release. Hydrobiologia 2007, 584, 37–48. [Google Scholar] [CrossRef]
- Zou, R.; Wu, Z.; Zhao, L.; Elser, J.J.; Yu, Y.; Chen, Y.; Liu, Y. Seasonal Algal Blooms Support Sediment Release of Phosphorus via Positive Feedback in a Eutrophic Lake: Insights from a Nutrient Flux Tracking Modeling. Ecol. Model. 2020, 416, 108881. [Google Scholar] [CrossRef]
- Fan, P.; Wang, Y.; Wang, W.; Chai, B.; Lu, X.; Zhao, J.; Fan, P.; Wang, W.; Lu, X. Release Characteristics of Nitrogen and Phosphorus from Sediments Formed under Different Supplemental Water Sources in Xi’an Moat, China. Environ. Sci. Pollut. Res. 2019, 26, 10746–10755. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhang, L.; Wang, S.; Jiao, L. Features and Influencing Factors of Nitrogen and Phosphorus Diffusive Fluxes at the Sediment-Water Interface of Erhai Lake. Environ. Sci. Pollut. Res. 2018, 25, 1933–1942. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Lin, L.; Huang, Z.; Liu, M.; Dong, L.; Chen, J.; Crittenden, J. Distribution Characteristics and Pollution Risk Evaluation of the Nitrogen and Phosphorus Species in the Sediments of Lake Erhai, Southwest China. Environ. Sci. Pollut. Res. 2019, 26, 22295–22304. [Google Scholar] [CrossRef]
- Ni, Z.; Wang, S. Historical Accumulation and Environmental Risk of Nitrogen and Phosphorus in Sediments of Erhai Lake, Southwest China. Ecol. Eng. 2015, 79, 42–53. [Google Scholar] [CrossRef]
- Chaturvedi, S.; Rajasekar, E. Application of a Probabilistic LHS-PAWN Approach to Assess Building Cooling Energy Demand Uncertainties. Build. Simul. 2022, 15, 373–387. [Google Scholar] [CrossRef]
- Pianosi, F.; Wagener, T. A Simple and Efficient Method for Global Sensitivity Analysis Based On Cumulative Distribution Functions. Environ. Model. Softw. 2015, 67, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pianosi, F.; Wagener, T. Distribution-Based Sensitivity Analysis from a Generic Input-Output Sample. Environ. Model. Softw. 2018, 108, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Muleta, M.K.; Nicklow, J.W. Sensitivity and Uncertainty Analysis Coupled with Automatic Calibration for a Distributed Watershed Model. J. Hydrol. 2005, 306, 127–145. [Google Scholar] [CrossRef] [Green Version]
- Campolongo, F.; Cariboni, J.; Saltelli, A. An Effective Screening Design for Sensitivity Analysis of Large Models. Environ. Model. Softw. 2007, 22, 1509–1518. [Google Scholar] [CrossRef]
- Tang, T.J.; Yang, S.; Peng, Y.; Yin, K.; Zou, R. Eutrophication Control Decision Making Using EFDC Model for Shenzhen Reservoir, China. Water Resour. 2017, 44, 308–314. [Google Scholar] [CrossRef]
- Wu, G.; Xu, Z. Prediction of Algal Blooming Using EFDC Model: Case Study in the Daoxiang Lake. Ecol. Model. 2011, 222, 1245–1252. [Google Scholar] [CrossRef]
- Kim, J.; Lee, T.; Seo, D. Algal Bloom Prediction of the Lower Han River, Korea Using the EFDC Hydrodynamic and Water Quality Model. Ecol. Model. 2017, 366, 27–36. [Google Scholar] [CrossRef]
- Yi, X.; Zou, R.; Guo, H. Global Sensitivity Analysis of a Three-Dimensional Nutrients-Algae Dynamic Model for a Large Shallow Lake. Ecol. Model. 2016, 327, 74–84. [Google Scholar] [CrossRef]
- Arhonditsis, G.B.; Brett, M.T. Eutrophication Model for Lake Washington (USA): Part I. Model Description and Sensitivity Analysis. Ecol. Model. 2005, 187, 140–178. [Google Scholar] [CrossRef]
- Jiang, L.; Li, Y.; Zhao, X.; Tillotson, M.R.; Wang, W.; Zhang, S.; Sarpong, L.; Asmaa, Q.; Pan, B. Parameter Uncertainty and Sensitivity Analysis of Water Quality Model in Lake Taihu, China. Ecol. Model. 2018, 375, 1–12. [Google Scholar] [CrossRef]
- Cao, J.; Hou, Z.; Li, Z.; Chu, Z.; Yang, P.; Zheng, B. Succession of Phytoplankton Functional Groups and Their Driving Factors in a Subtropical Plateau Lake. Sci. Total Environ. 2018, 631–632, 1127–1137. [Google Scholar] [CrossRef]
- Yu, G.; Jiang, Y.; Song, G.; Tan, W.; Zhu, M.; Li, R. Variation of Microcystis and Microcystins Coupling Nitrogen and Phosphorus Nutrients in Lake Erhai, a Drinking-Water Source in Southwest Plateau, China. Environ. Sci. Pollut. Res. 2014, 21, 9887–9898. [Google Scholar] [CrossRef]
- Nelson, N.G.; Muñoz-Carpena, R.; Phlips, E. Parameter Uncertainty Drives Important Incongruities between Simulated Chlorophyll-a and Phytoplankton Functional Group Dynamics in a Mechanistic Management Model. Environ. Model. Softw. 2020, 129, 104708. [Google Scholar] [CrossRef]
- Sadeghian, A.; Chapra, S.C.; Hudson, J.; Wheater, H.; Lindenschmidt, K.E. Improving In-Lake Water Quality Modeling Using Variable Chlorophyll a/Algal Biomass Ratios. Environ. Model. Softw. 2018, 101, 73–85. [Google Scholar] [CrossRef]
- Scavia, D.; David Allan, J.; Arend, K.K.; Bartell, S.; Beletsky, D.; Bosch, N.S.; Brandt, S.B.; Briland, R.D.; Daloğlu, I.; DePinto, J.V.; et al. Assessing and Addressing the Re-Eutrophication of Lake Erie: Central Basin Hypoxia. J. Great Lakes Res. 2014, 40, 226–246. [Google Scholar] [CrossRef]
- Bertani, I.; Obenour, D.R.; Steger, C.E.; Stow, C.A.; Gronewold, A.D.; Scavia, D. Probabilistically Assessing the Role of Nutrient Loading in Harmful Algal Bloom Formation in Western Lake Erie. J. Great Lakes Res. 2016, 42, 1184–1192. [Google Scholar] [CrossRef] [Green Version]
- Rucinski, D.K.; DePinto, J.V.; Beletsky, D.; Scavia, D. Modeling Hypoxia in the Central Basin of Lake Erie under Potential Phosphorus Load Reduction Scenarios. J. Great Lakes Res. 2016, 42, 1206–1211. [Google Scholar] [CrossRef]
- Ni, Z.; Wang, S.; Cai, J.; Li, H.; Jenkins, A.; Maberly, S.C.; May, L. The Potential Role of Sediment Organic Phosphorus in Algal Growth in a Low Nutrient Lake. Environ. Pollut. 2019, 255, 113235. [Google Scholar] [CrossRef]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime Shifts, Resilience, and Biodiversity in Ecosystem Management. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar] [CrossRef] [Green Version]
- Scheffer, M.; Carpenter, S.R.; Lenton, T.M.; Bascompte, J.; Brock, W.; Dakos, V.; van de Koppel, J.; van de Leemput, I.A.; Levin, S.A.; van Nes, E.H.; et al. Anticipating Critical Transitions. Science 2012, 338, 344–348. [Google Scholar] [CrossRef] [Green Version]
- Spears, B.M.; Futter, M.N.; Jeppesen, E.; Huser, B.J.; Ives, S.; Davidson, T.A.; Adrian, R.; Angeler, D.G.; Burthe, S.J.; Carvalho, L.; et al. Ecological Resilience in Lakes and the Conjunction Fallacy. Nat. Ecol. Evol. 2017, 1, 1616–1624. [Google Scholar] [CrossRef]
- Mellor, G.L.; Yamada, T. Development of a turbulence closure model for geophysical fluid problems. Rev. Geophys. 1982, 20, 851. [Google Scholar] [CrossRef] [Green Version]
- Galperin, B.; Kantha, L.H.; Hassid, S.; Rosati, A. A quasi-equilibrium turbulent energy model for geophysical flows. J. Atmos. Sci. 1988, 45, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Qi, H.; Lu, J.; Chen, X.; Sauvage, S.; Sanchez-Pérez, J.M. Water Age Prediction and Its Potential Impacts on Water Quality Using a Hydrodynamic Model for Poyang Lake, China. Environ. Sci. Pollut. Res. 2016, 23, 13327–13341. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hua, Z.; Wang, L. Sensitivity Analysis of the Chaohu Lake Eutrophication Model with a New Index Based on the Morris Method. Water Sci. Technol. Water Supply 2018, 18, 1375–1387. [Google Scholar] [CrossRef]
- Jiang, Q.; Su, H.; Liu, Y.; Zou, R.; Ye, R.; Guo, H. Parameter Uncertainty-Based Pattern Identification and Optimization for Robust Decision Making on Watershed Load Reduction. J. Hydrol. 2017, 547, 708–717. [Google Scholar] [CrossRef]
- Tetra Tech. The Environmental Fluid Dynamics Code Theory and Computation. Volume 3: Water Quality Module; Tetra Tech: Fairfax, VA, USA, 2007; Volume 3, 60p. [Google Scholar]
- Jørgensen, S.E.; Friis, M.B.; Henriksen, J. Handbook of Environmental Data and Ecological Parameters; Oxford (UK) Pergamon Press for International Society for Ecological Modelling: Oxford, UK, 1979. [Google Scholar]







| Parameters | Units | Description | Min | Max |
|---|---|---|---|---|
| PMc | 1/day | Maximum growth rate for cyanobacteria | 1.5 | 2 |
| PMd | 1/day | Maximum growth rate for diatom | 1.0 | 2 |
| PMg | 1/day | Maximum growth rate for green algae | 1.5 | 2.5 |
| BMRd | 1/day | Basal metabolism rate for diatom | 0.15 | 0.3 |
| BMRg | 1/day | Basal metabolism rate for green algae | 0.06 | 0.12 |
| PRRd | 1/day | Predation rate for diatoms | 0.01 | 0.1 |
| PRRg | 1/day | Predation rate for green algae | 0.01 | 0.1 |
| TMc1 | °C | Lower optimal temperature for growth of cyanobacteria | 20 | 25 |
| TMc2 | °C | Upper optimal temperature for growth of cyanobacteria | 26 | 30 |
| KTG1c | / | Suboptimal temperature effect coefficient for growth of cyanobacteria | 0.001 | 0.01 |
| KHNc | mg/L | Nitrogen half-saturation for cyanobacteria | 0.1 | 0.35 |
| KHPc | mg/L | Phosphorus half-saturation for cyanobacteria | 0.0015 | 0.0025 |
| KHPg | mg/L | Phosphorus half-saturation for phytoplankton:greens | 0.002 | 0.004 |
| WSd | 1/day | Settling velocity for diatoms | 0.3 | 0.5 |
| WSrp | 1/day | Settling velocity for Refractory particulate organic matter (RPOM) | 0.0 | 0.15 |
| CPprm1 | g c/g P | Minimum algae carbon to phosphorus ratio | 40 | 60 |
| rNitM | 1/day | Maximum nitrification rate | 0.15 | 0.3 |
| KHNitN | gN/m3 | NH4 half-saturation constant for nitrification | 0.3 | 0.8 |
| KHNitDO | gO2/m3 | Oxygen half-saturation constant for nitrification | 0.5 | 1 |
| KHORDO | gO2/m3 | Oxygen half-saturation constant for algal respiration | 1.5 | 2.5 |
| KDC | 1/day | Minimum dissolution rate of dissolved organic carbon (DOC) | 0.0015 | 0.0025 |
| Parameters Variables | Rank of Sensitivity | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |
| Total Biomass | KHPg | PRRg | KHPc | PMc | WSrp | PMg |
| KS value | 0.274 | 0.241 | 0.212 | 0.211 | 0.196 | 0.188 |
| Biomass (cyanobacteria) | PRRg | KHPg | KHPg | KHPc | PMc | BMRg |
| KS value | 0.292 | 0.290 | 0.234 | 0.212 | 0.211 | 0.196 |
| Biomass (diatom) | BMRd | PRRd | PMd | KHNc | KHPc | PRRg |
| KS value | 0.342 | 0.236 | 0.230 | 0.200 | 0.193 | 0.192 |
| Biomass (green) | PRRg | BMRd | KHPc | BMRg | KHNc | PMd |
| KS value | 0.275 | 0.220 | 0.219 | 0.214 | 0.204 | 0.200 |
| Loading Reduction Ratio | Reduction in TP t/a | |
|---|---|---|
| L0 | / | 0 |
| L1 | 15% reduction in TP | 22.35 |
| L2 | 25% reduction in TP | 37.25 |
| L3 | 50% reduction in TP | 74.5 |
| L4 | 75% reduction and TP | 111.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Zhang, J.; Hou, Z.; Tian, Z.; Chu, Z.; Wang, S. Seasonal Dynamics of Algal Net Primary Production in Response to Phosphorus Input in a Mesotrophic Subtropical Plateau Lake, Southwestern China. Water 2022, 14, 835. https://doi.org/10.3390/w14050835
Wu Y, Zhang J, Hou Z, Tian Z, Chu Z, Wang S. Seasonal Dynamics of Algal Net Primary Production in Response to Phosphorus Input in a Mesotrophic Subtropical Plateau Lake, Southwestern China. Water. 2022; 14(5):835. https://doi.org/10.3390/w14050835
Chicago/Turabian StyleWu, Yue, Jinpeng Zhang, Zeying Hou, Zebin Tian, Zhaosheng Chu, and Shengrui Wang. 2022. "Seasonal Dynamics of Algal Net Primary Production in Response to Phosphorus Input in a Mesotrophic Subtropical Plateau Lake, Southwestern China" Water 14, no. 5: 835. https://doi.org/10.3390/w14050835