
Development of Quantitative Real-Time PCR for Detecting Environmental DNA Derived from Marine Macrophytes and Its Application to a Field Survey in Hiroshima Bay, Japan

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. Quantitation of Extracted DNA and Plasmid DNA
2.3. DNA Sequences of Internal Transcribed Spacer (ITS) and Maturasek (matK) Genes of Various Macrophyte DNA
2.4. Design of Primers and DLPs for Detecting Various Macrophyte DNA
2.5. Plasmid Development for Construction of the Standard Curve
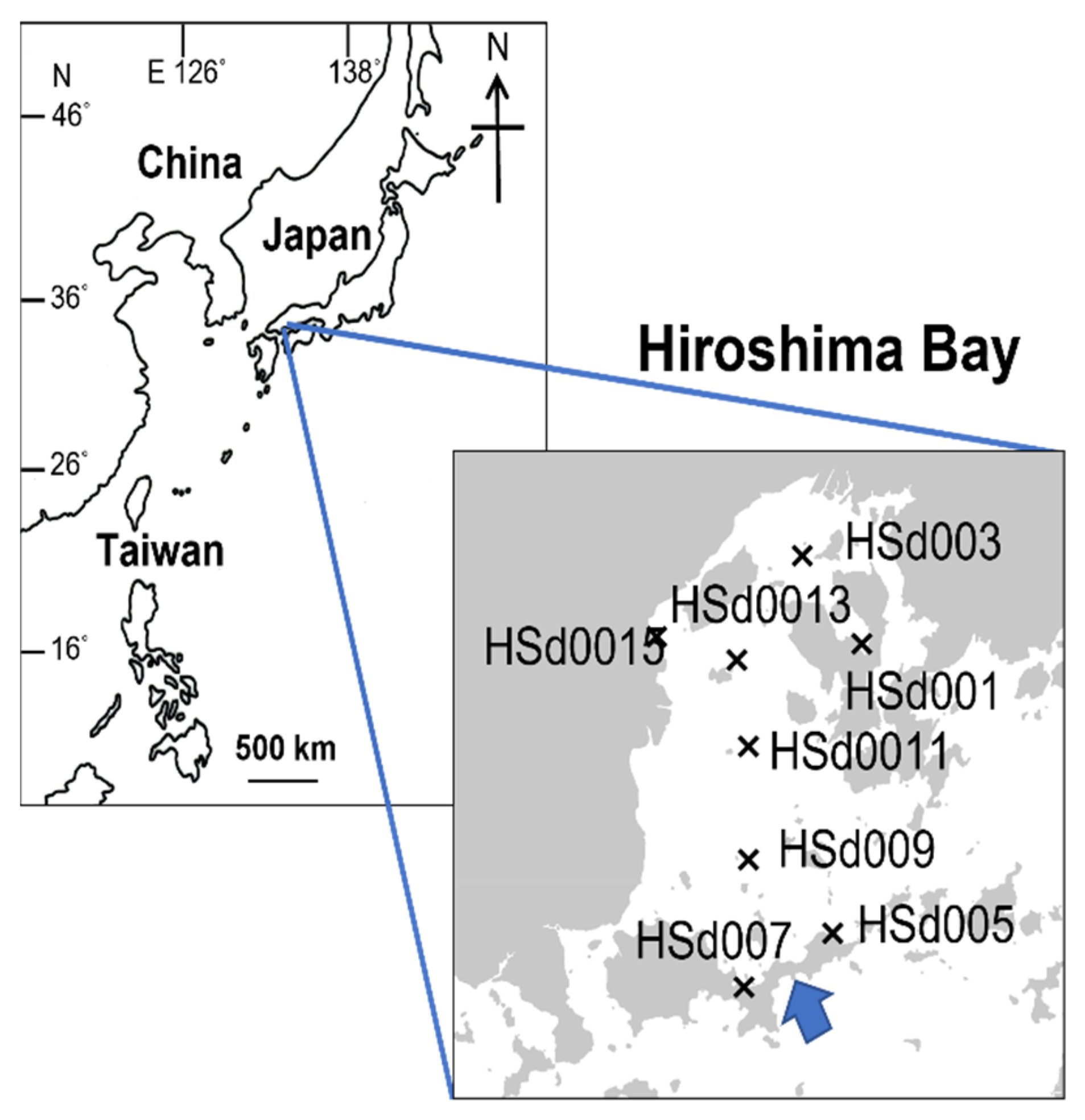
2.6. Application of the Newly Developed qPCR System to Field Sediment Samples
2.7. Calculation of Limit of Detection (LOD) and Limit of Quantification (LOQ) for eDNA
2.8. Analysis of Physical and Chemical Properties of the Sediment Samples Collected from Hiroshima Bay
3. Results
3.1. Design of Primers and DLPs for Detecting DNA of Various Macrophyte Taxa
3.2. Detection of eDNA Derived from Mangroves
3.3. Detection of eDNA Derived from Seagrass
3.4. Detection of eDNA Derived from Phaeophyceae, Rhodophyta and Chlorophyta
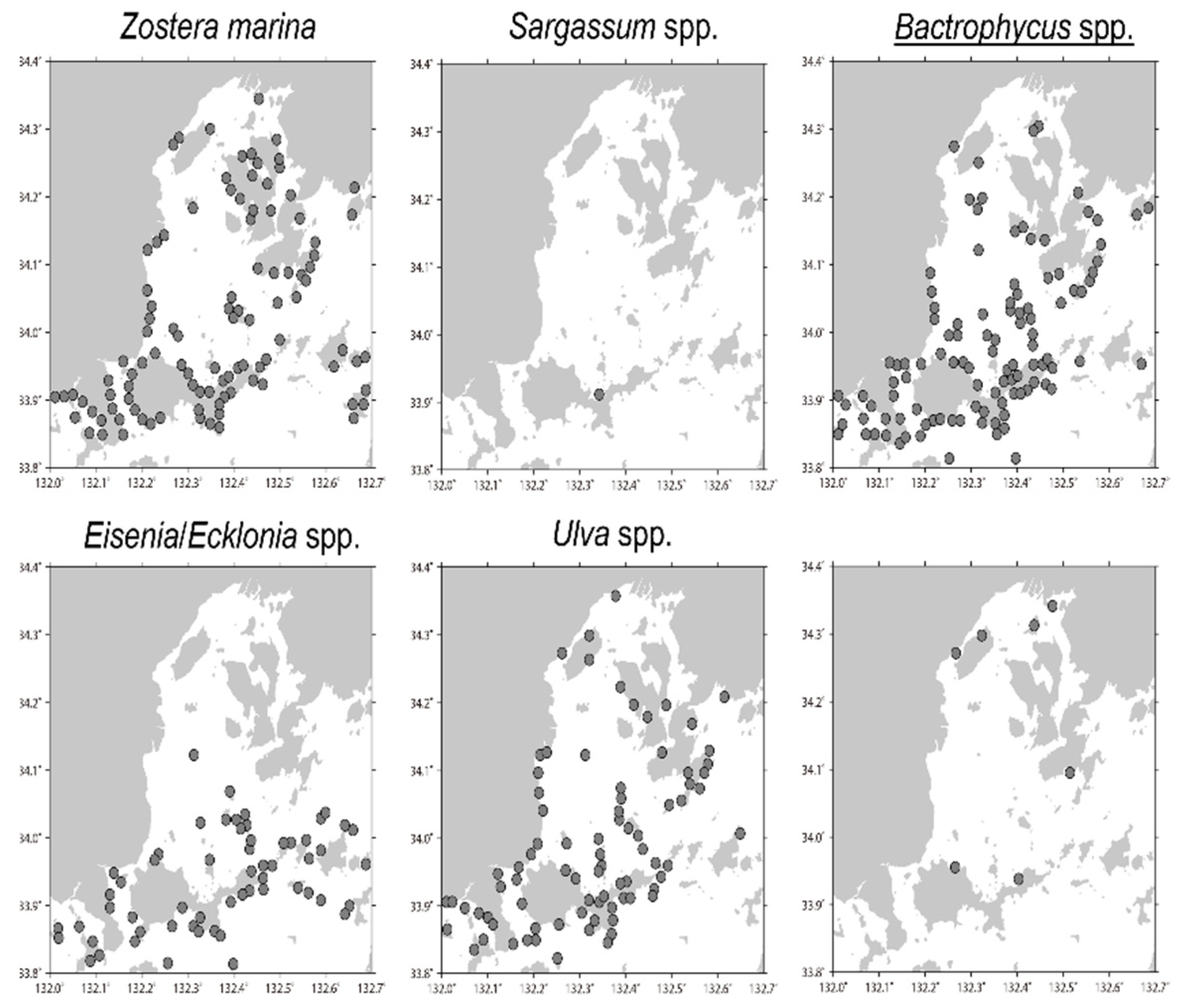
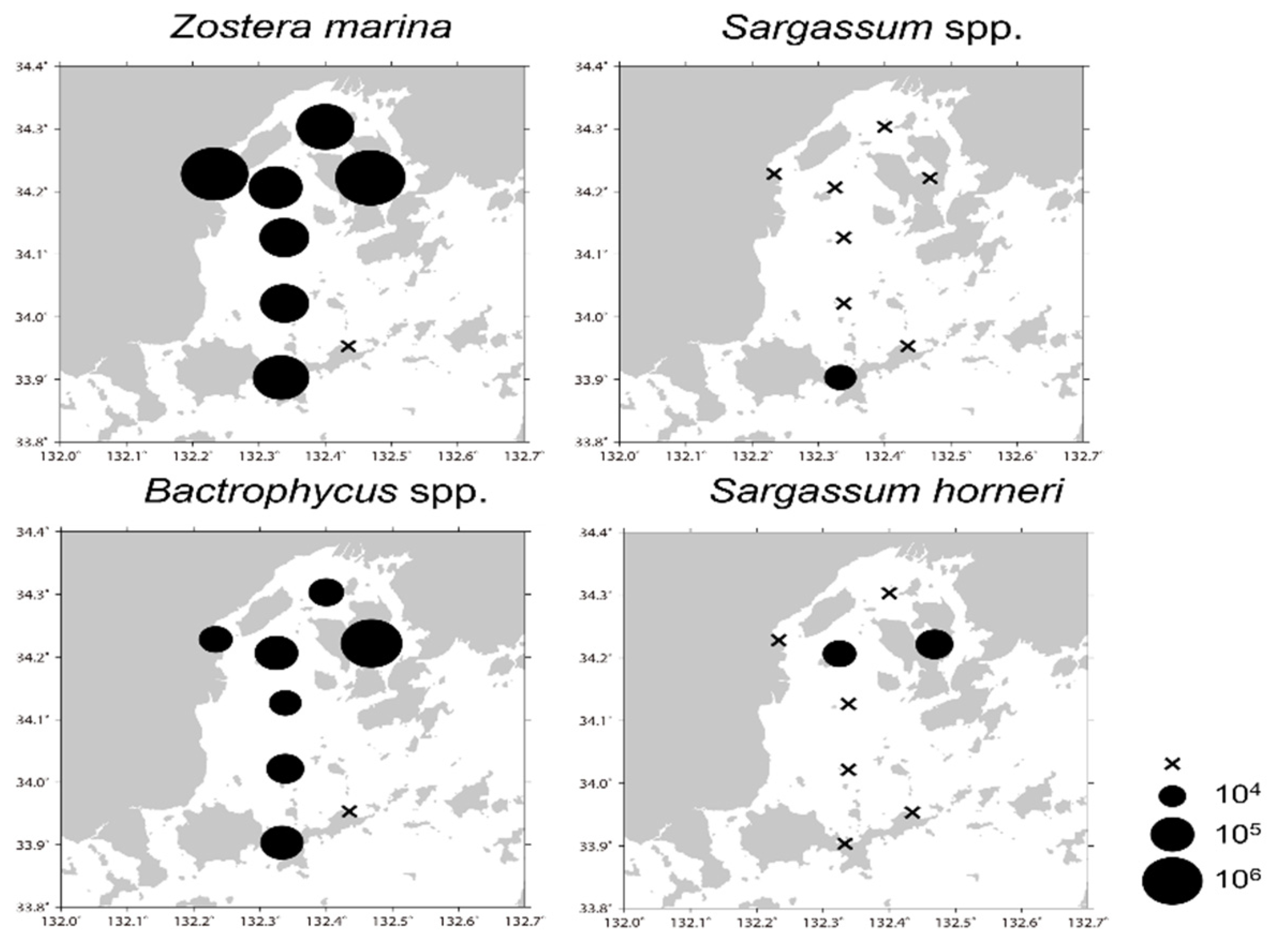
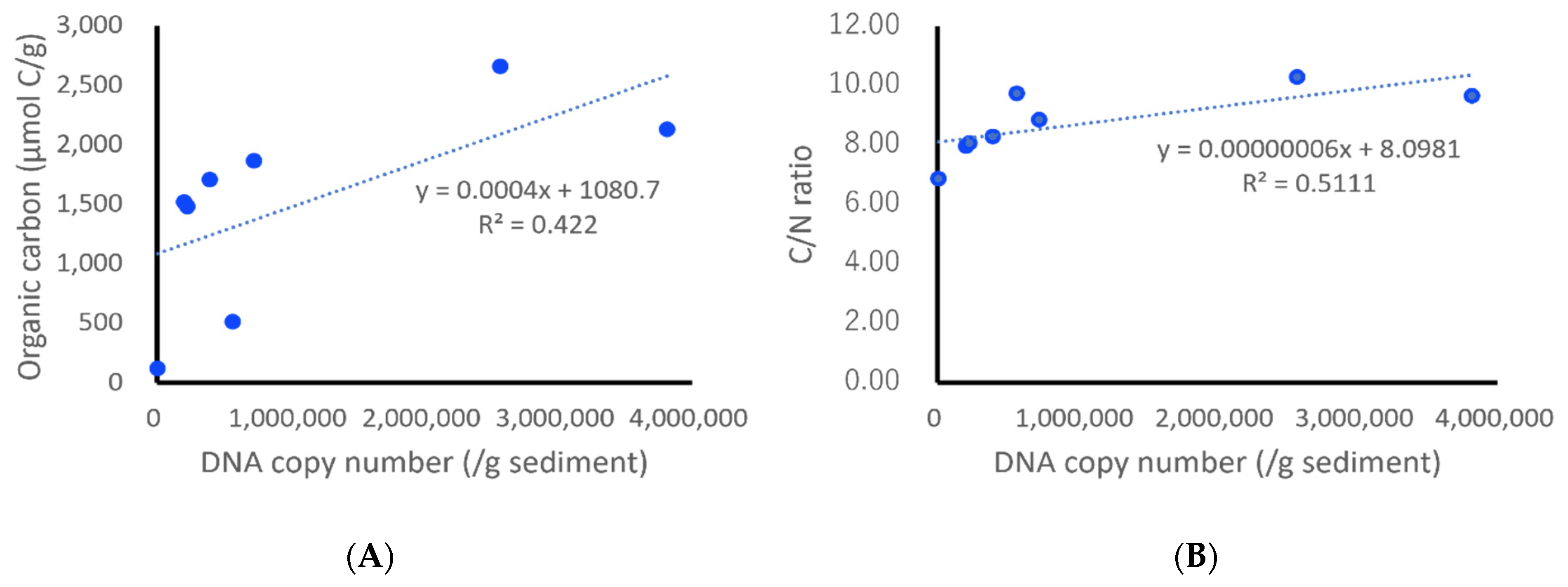
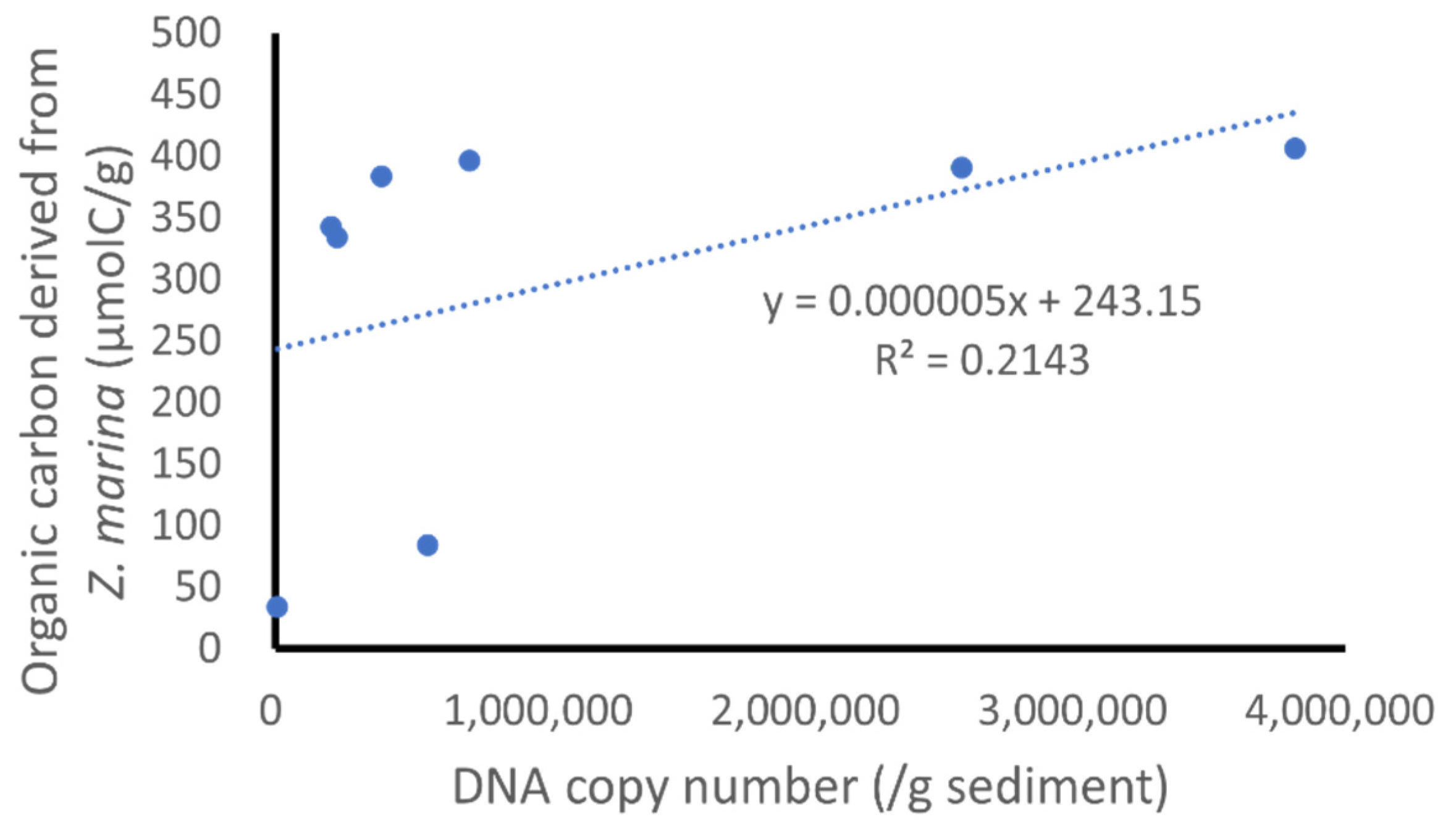
3.5. Application of the Designed qPCR System to Field Samples
4. Discussion
4.1. Recommended Reaction System
4.1.1. Points to Note in the Design of Detection Systems for qPCR
4.1.2. Which DNA Polymerase and Real-Time PCR Instruments to Use?
4.2. Detection of eDNA Derived from Mangroves
4.3. Detection of eDNA Derived from Seagrasses
4.4. Detection of eDNA Derived from Phaeophyceae, Rhodophyta and Chlorophyta
4.5. Application of the Designed qPCR System to Field Samples
4.6. Future Research Focus and Concepts
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nellemann, C.; Corcoran, E.; Duarte, C.M.; Valdés, L.; De Young, C.; Fonseca, L.; Grimsditch, G. (Eds.) Blue Carbon: The Role of Healthy Oceans in Binding Carbon. A Rapid Response Assessment; United Nations Environment Programme: Nairobi, Kenya, 2009. [Google Scholar]
- Hori, M.; Kuwae, T. Blue Carbon -CO2 Uptake and Carbon Storage in Shallow Coastal Ecocystems and Their Utilization; Chijin-Kan: Tokyo, Japan, 2017; pp. 1–254. (In Japanese) [Google Scholar]
- Hori, M.; Bayne, C.J.; Kuwae, T. Blue carbon: Characteristics of the Ocean’s sequestration and storage ability of carbon Dioxide. In Blue Carbon in Shallow Coastal Ecosystems; Kuwae, T., Hori, M., Eds.; Springer: Singapore, 2018; pp. 1–32. [Google Scholar]
- Bertram, C.; Quaas, M.; Reusch, T.B.H.; Vafeidis, A.T.; Wolff, C.; Rickels, W. The blue carbon wealth of nations. Nat. Clim. Chang. 2021, 11, 704–709. [Google Scholar] [CrossRef]
- Krause-Jensen, D.; Lavery, P.; Serrano, O.; Marba, N.; Masque, P.; Duarte, C.M. Sequestration of macroalgal carbon: The elephant in the blue carbon room. Biol. Lett. 2018, 14, 20180236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Jiang, Z.; Wu, Y.; Deng, Y.; Chen, Q.; Zhao, C.; Cui, L.; Huang, X. Macroalgae bloom decay decreases the sediment organic carbon sequestration potential in tropical seagrass meadows of the South China Sea. Mar. Pollut. Bull. 2019, 138, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Moura Queiros, A.; Stephens, N.; Widdicombe, S.; Tait, K.; McCoy, S.J.; Ingels, J.; Ruhi, S.; Airs, R.; Beesley, A.; Carnovale, G.; et al. Connected macroalgal-sediment systems: Blue carbon and food webs in the deep coastal ocean. Ecol. Monogr. 2019, 89, e01366. [Google Scholar]
- Bartolo, A.G.; Zammit, G.; Peters, A.F.; Küpper, F.C. The current state of DNA barcoding of macroalgae in the Mediterranean Sea: Presently lacking but urgently required. Bot. Mar. 2020, 63, 253–272. [Google Scholar] [CrossRef]
- Besterman, A.F.; McGlathery, K.J.; Reidenbach, M.A.; Wiberg, P.L.; Pace, M.L. Predicting benthic macroalgal abundance in shallow lagoons from geomorphology and hydrologic flow patterns. Limnol. Oceanogr. 2021, 66, 123–140. [Google Scholar] [CrossRef]
- Hilmi, N.; Chami, R.; Sutherland, M.D.; Hall-Spencer, J.M.; Lebleu, L.; Benitez, M.B.; Levin, L.A. The role of blue carbon in climate change mitigation and carbon stock conservation. Front. Clim. 2021, 3, 710546. [Google Scholar] [CrossRef]
- Rees, H.C.; Maddison, B.C.; Middleditch, J.; Patmore, J.R.M.; Gough, K.C. The detection of aquatic animal species using environmental DNA—A review of eDNA as a survey tool in ecology. J. Appl. Ecol. 2014, 51, 1450–1459. [Google Scholar] [CrossRef]
- Pedersen, M.W.; Overballe-Peterson, S.; Ermini, L.; Sarkissian, C.D.; Haile, J.; Hellstrom, M.; Spens, J.; Thomsen, P.F.; Bohmann, K.; Cappellini, E.; et al. Ancient and modern environmental DNA. Phil. Trans. R. Soc. B 2015, 370, 20130383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevathan-Tackett, S.M.; Jeffries, T.C.; Macreadie, P.L.; Manojlovic, B.; Ralph, P. Long-term decomposition captures key steps in microbial breakdown of seagrass litter. Sci. Total Environ. 2020, 705, 135806. [Google Scholar] [CrossRef] [PubMed]
- Jeunen, G.-J.; Knapp, M.; Spencer, H.G.; Taylor, H.R.; Lamare, M.D.; Stat, M.; Bunce, M.; Gemmell, N.J. Species-level biodiversity assessment using marine environmental DNA metabarcoding requires protocol optimization and standardization. Ecol. Evol. 2019, 9, 1323–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, D.C.; Bunce, M.; Cannell, B.L.; Oliver, R.; Houston, J.; White, N.E.; Barrero, R.A.; Bellgard, M.I.; Haile, J. DNA-based faecal dietary analysis: A comparison of qPCR and high throughput sequencing approaches. PLoS ONE 2011, 10, e25776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, L.R.; Handley, L.L.; Hahn, C.; Boonham, N.; Rees, H.C.; Gough, K.C.; Lewis, E.; Adams, I.P.; Brotherton, P.; Phillips, S.; et al. Needle in a haystack? A comparison of eDNA metabarcoding and targeted qPCR for detection of the great crested newt (Triturus cristatus). Ecol. Evol. 2018, 8, 6330–6341. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, S.; Chaves, C.; Velo-Anton, G.; Beja, P.; Egeter, B. Species detection from aquatic eDNA: Assessing the importance of capture methods. Environ. DNA 2021, 3, 435–448. [Google Scholar] [CrossRef]
- Reef, R.; Atwood, T.B.; Samper-Villarreal, J.; Adame, M.F.; Sampayo, E.M.; Lovelock, C.E. Using eDNA to determine the source of organic carbon in seagrass meadows. Limnol. Oceanogr. 2017, 62, 1254–1265. [Google Scholar] [CrossRef]
- Ortega, A.; Geraldi, N.R.; Duarte, C.M. Environmetal DNA identifies marine macrophyte contributions to blue cabon sediments. Limnol. Oceanogr. 2020, 9999, 1–11. [Google Scholar]
- Ortega, A.; Geraldi, N.R.; Diaz-Rua, R.; Orberg, S.B.; Wesselmann, M.; Krause-Jensen, D.; Duarte, C.M. A DNA mini-barcoding for marine macrophytes. Mol. Ecol. Resour. 2020, 20, 920–935. [Google Scholar] [CrossRef] [PubMed]
- Ushio, M.; Murakami, H.; Masuda, R.; Sado, T.; Miya, M.; Sakurai, S.; Yamanaka, H.; Minamoto, T.; Kondoh, M. Quantitative monitoring of multispecies fish environmental DNA using high-throughput sequencing. Metabarcoding Metagenom. 2018, 2, 1–15. [Google Scholar] [CrossRef]
- Sato, M.; Inoue, N.; Nambu, R.; Furuichi, N.; Imaizumi, T.; Ushio, M. Quantitative assessment of multiple fish species around artificial reefs combining environmental DNA metabarcoding and acoustic survey. Sci. Rep. 2021, 11, 19477. [Google Scholar] [CrossRef]
- Holland, P.M.; Abramson, R.D.; Watson, R.; Gelfand, D.H. Detection of specific polymerase chain reaction product by utilizing the 5′ –3′ exonuclease activity of Thermus aquaticus DNA polymerase. Proc. Natl. Acad. Sci. USA 1991, 88, 7276–7280. [Google Scholar] [CrossRef] [Green Version]
- Kevadiya, B.D.; Machhi, J.; Herskovitz, J.; Oleynikov, M.D.; Blomberg, W.R.; Bajwa, N.; Soni, D.; Das, S.; Hasan, M.; Patel, M.; et al. Diagnostics for SARS-CoV-2 infections. Nat. Mater. 2021, 20, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Vitásková1, E.; Černíková1, L.; Křivda1, V.; Jiřincová, H.; Sedlák, K.; Horníčková, J.; Havlíčková, M. Evaluation of TaqMan qPCR system integrating two identically labelled hydrolysis probes in single assay. Sci. Rep. 2017, 7, 41392. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, N.; Hamaguchi, M. Multiplex reverse transcription quantitative PCR detection of a single-stranded RNA virus HcRNAV infecting the bloom-forming dinoflagellate Heterocapsa circularisquama. Limnol. Oceanogr. Methods 2016, 14, 370–380. [Google Scholar] [CrossRef] [Green Version]
- Hamaguchi, M.; Shimabukuro, H.; Hori, M.; Yoshida, G.; Terada, T.; Miyajima, T. Quantitative real-time polymerase chain reaction (PCR) and droplet digital PCR duplex assays for detecting Zostera marina DNA in coastal sediments. Limnol. Oceanogr. Methods 2018, 16, 253–264. [Google Scholar] [CrossRef] [Green Version]
- Task Force on National Greenhouse Gas Inventories: Wetlands. Available online: https://www.ipcc-nggip.iges.or.jp/public/wetlands/index.html (accessed on 1 February 2022).
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffi, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative Real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymaekers, M.; Smets, R.; Maes, B.; Cartuyvels, R. Chechlist for optimization and validation of real-time PCR assays. J. Clin. Lab. Anal. 2009, 23, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Abliz, P.; Fukushima, K.; Takizawa, K.; Nieda, N.; Miyaji, M.; Nishimura, K. Rapid identification of the genus Fonsecaea by PCR with specific oligonucleotide primers. J. Clin. Microbiol. 2003, 41, 873–876. [Google Scholar] [CrossRef] [Green Version]
- Lucas, C.; Thangaradjou, T.; Papenbrock, J. Development of a DNA barcoding system for seagrasses: Successful but not simple. PLoS ONE 2012, 7, e29987. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Xue, J.-H.; Zhou, S.-L. New universal matK primers for DNA barcoding angiosperms. J. Syst. Evol. 2011, 49, 176–181. [Google Scholar] [CrossRef]
- Stiger, V.; Horiguchi, T.; Yoshida, T.; Coleman, A.W.; Matsuda, M. Phylogenetic relationships within the genus Sargassum (Fucales, Phaeophyceae), inferred from ITS-2 nrDNA, with an emphasis on the taxonomic subdivision of the genus. Phycol. Res. 2003, 51, 1–10. [Google Scholar] [CrossRef]
- Yotsukura, N.; Shimizu, T.; Katayama, T.; Druehl, L.D. Mitochondrial DNA sequence variation of four Saccharina species (Laminariales, Phaeophyceae) growing in Japan. J. Appl. Phycol. 2010, 22, 243–251. [Google Scholar] [CrossRef]
- Rana, S.; Valentin, K.; Riehl, J.; Balanfune, A.; Reynes, L.; Thibaut, T.; Bartsch, I.; Eichinger, L.; Glockner, G. Analysis of organellar genomes in brown algae reveals an independent introduction of similar foreign sequences into the mitochondrial genome. Genomics 2021, 113, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xiao, S.; Yu, Z. Mitochondrial DNA and morphological identification of Crassostrea zhanjiangensis sp. nov. (Bivalvia: Ostreidae): A new species in Zhanjiang, China. Aquat. Living Resour. 2013, 26, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Orr, W.J.; Wildes, S.; Kai, Y.; Raring, T.; Nakabo, T.; Katugin, O.; Guyon, J. Systematics of North Pacific sand lances of the genus Ammodytes based on molecular and morphological evidence, with the description of a new species from Japan. Fish. Bull. 2015, 113, 129–156. [Google Scholar] [CrossRef]
- Broeders, S.; Huber, I.; Grohmann, I.; Berben, G.; Taverniers, I.; Mazzara, M.; Roosens, N.; Morisset, D. Guidelines for validation of qualitative real-time PCR methods. Trends Food Sci. Technol. 2014, 37, 115–126. [Google Scholar] [CrossRef]
- Ma, H.; Bell, K.N.; Loker, R.N. qPCR and qRT-PCR analysis: Regulatory points to consider when conducting biodistribution and vector shedding studies. Mol. Ther. Methods Clin. Develop. 2020, 20, 152–168. [Google Scholar] [CrossRef] [PubMed]
- Schrader, C.; Schelke, A.; Ellerbroek, L.; Johne, R. PCR inhibitors- occurrence, properties, and removal. Appl. Microbiol. 2012, 113, 1014–1026. [Google Scholar] [CrossRef]
- Sidstedt, M.; Radstrom, P.; Hedman, J. PCR inhibition in qPCR, dPCR and MPS-mechanisms and solutions. Anal. Bioanal. Cem. 2020, 412, 2009–2023. [Google Scholar] [CrossRef] [Green Version]
- The European Union Reference Laboratories: Guidance Document on the Estimation of LOD and LOQ for Measurements in the Field of Contaminants in Feed and Food. Available online: https://ec.europa.eu/jrc/sites/default/files/lod_loq_guidance_document_food_contaminants_2016.pdf (accessed on 1 February 2022).
- Miyajima, T.; Hori, M.; Hamaguchi, M.; Shimabukuro, H.; Adachi, H.; Yamano, H.; Nakaoka, M. Geographic variability in organic carbon stock and accumulation rate in sediments of East and Southeast Asian seagrass meadows. Glob. Biogeochem. Cycles 2015, 29, 397–415. [Google Scholar] [CrossRef]
- Miyajima, T.; Hori, M.; Hamaguchi, M.; Shimabukuro, H.; Yoshida, G. Geophysical constraints for organic carbon sequestration capacity of Zostera marina seagrass meadows and surrounding habitats. Limnol. Oceanogr. 2017, 62, 954–972. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. A Language and Environment Statistic Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.rproject.Org (accessed on 3 March 2018).
- Primer3Plus. Available online: https://primer3plus.com/cgi-bin/dev/primer3plus.cgi (accessed on 1 February 2022).
- DNA Software. Available online: https://www.dnasoftware.com/multiplex-pcr-designs-in-hours-not-months/?gclid=EAIaIQobChMIyaGwsfv_9AIV0VBgCh2W0wB0EAAYASAAEgJMEfD_BwE (accessed on 1 February 2022).
- Nakasuga, T.; Oyama, H.; Haruki, M. Studies on the mangrove community. 1. The distribution of the mangrove community in Japan. Jpn. J. Ecol. 1974, 24, 237–246, (In Japanese with English Abstract). [Google Scholar]
- Lo, E.Y.Y.; Duke, N.C.; Sun, M. Phylogeographic pattern of Rhizophora (Rhizophoraceae) reveals the importance of both vicariance and long-distance oceanic dispersal to modern mangrove distribution. BMC Evol. Biol. 2014, 14, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, T. Carbon sequestration in Mangroves. In Blue Carbon in Shallow Coastal Ecosystems; Kuwae, T., Hori, M., Eds.; Springer: Singapore, 2018; pp. 73–100. [Google Scholar]
- Miyajima, T.; Hamaguchi, M.; Nakamura, T.; Katayama, H.; Hori, M. Export and dispersal of coastal macrophyte-derived organic matter in deep, offshore sediment around Tokara and Yaeyama Islands, southwest Japan: Evaluation using quantative DNA probing techniques. Bull. Geol. Surv. Jpn. 2021, submitted.
- Seagrassbed 2020. Available online: http://www.biodic.go.jp/moni1000/findings/newsflash/pdf/amamoba_2020.pdf (accessed on 1 February 2022).
- Stokes, H.; Mortimer, J.A.; Hays, G.C.; Unsworth, R.K.F.; Laloe, J.-O.; Esteban, N. Green turtle diet is dominated by seagreass in the Western Indian Ocean except amongst gravid females. Mar. Biol. 2019, 166, 135. [Google Scholar] [CrossRef] [Green Version]
- Kato, Y.; Aioi, K.; Omori, Y.; Takahata, N.; Satta, Y. Phylogenetic analyses of Zostera species based on rbcL and matK nucleotide sequences: Implications for the origin and diversification of aeagrasses in Japanese waters. Genes Genet. Syst. 2003, 78, 329–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yotaukura, N. A review on nomenclature of laminariacean kelp growing in cold water areas of Japan. Jpn. J. Phycol. 2007, 55, 167–172. [Google Scholar]
- Wang, X.; Shao, Z.; Fu, W.; Yao, J.; Hu, Q.; Duan, D. Chloroplast genome of one brown seaweed, Saccharina japonica (Laminariales, Pheophyta): Its structural features and phylogenetic analyses with other photosynthetic plastids. Mar. Genom. 2013, 10, 1–9. [Google Scholar] [CrossRef]
- Ye, N.; Zhang, W.; Miao, M.; Fan, X.; Zheng, Y.; Xu, D.; Wang, J.; Zhou, L.; Wang, D.; Gao, Y.; et al. Saccharina genomes provide novel insight into kelp biology. Nar. Commun. 2015, 6, 6986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothman, M.D.; Mattio, L.; Wernberg, T.; Anderson, R.J.; Uwai, S.; Mohring, M.B.; Bolton, J.J. A molecular investigation of the genus Ecklonia (Phaeophyceae, Laminariales) with special focus on the southern hemisphere. J. Phycol. 2015, 51, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Kawai, H.; Akita, S.; Hashimoto, K.; Hanyuda, T. A multigene molecular phylogeny of Eisenia reveals evidence for a new species, Eisenia nipponica (Laminariales), from Japan. Eur. J. Phycol. 2020, 55, 234–241. [Google Scholar] [CrossRef]
- Ichihara, K.; Arai, S.; Uchimura, M.; Fay, E.J.; Ebata, H.; Hiraoka, M.; Shimada, S. New species of freshwater Ulva, Ulva limnetica (Ulvales, Ulvophyceas) from the Ryukyu Islands, Japan. Phycol. Res. 2009, 57, 94–103. [Google Scholar] [CrossRef]
- Masakiyo, Y.; Shimada, S. Species diversity the genus Ulva (Ulvophyceae, Chlorophyta) in Japanese waters, with special reference to Ulva tepida Masakiyo et S. Shimada sp. nov. Bull. Natl. Mus. Nat. Sci. Ser. B 2014, 40, 1–13. [Google Scholar]
- Ichihara, K.; Suzuki, R.; Yamazaki, T.; Ota, S.; Mogi, Y.; Kagami, Y.; Kuwano, K.; Kawano, S. Ulva partita sp. nov., a novel Enteromorpha-like Ulva species from Japanese coastal areas. Cytologia 2015, 80, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Hanyuda, T.; Heesch, S.; Nelson, W.; Sutherland, J.; Arai, S.; Boo, S.M.; Kawai, H. Genetic diversity and biogeography of native and introduced populations of Ulva pertusa (Ulvales, Chlorophyta). Phycol. Res. 2016, 64, 102–109. [Google Scholar] [CrossRef]
- Yang, L.; Deng, Y.-Y.; Xu, G.-P.; Russell, S.; Lu, Q.-Q.; Bradie, J. Redefining Pyropia (Bangiales, Rhodophyta): Four new genera, Resurrection of Porphyrella and Description of Calidia Psedolobata sp. nov. from China. J. Phycol. 2020, 56, 862–879. [Google Scholar] [CrossRef] [PubMed]
- Fort, A.; HcHale, M.; Cascella, K.; Potin, P.; Usadel, B.; Guiry, M.D.; Sulpice, R. Foliose Ulva species show considerable inter-specific genetic diversity, low intra-specific genetic variation, and the rare occurrence of interspecific hybrids in the wild. J. Phycol. 2021, 57, 219–233. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, Y.; Du, M.; Lin, F.; Jiang, W.; Li, W.; Li, F.; Lv, X.; Fang, J.; Jiang, Z. Dissolved organic carbon from cultured kelp Saccharina japonica: Production, bioavailability, and bacterial degradation rates. Aquacult. Environ. Inetract. 2021, 13, 101–110. [Google Scholar] [CrossRef]
- Yoshida, G.; Uchimura, M.; Hiraoka, M.; Terawaki, T.; Arai, S.; Iseki, K. Accumulation of Ulva spp. (Chlorophyta) and other seaweed thalli on the shallow sea bottom of Hiroshima Bay (A preliminary survey). Biosph. Sci. 2010, 49, 31–38, (In Japanese with English Abstract). [Google Scholar]
- Miyajima, T.; Hamaguchi, M. Carbon sequestration in sediment as an ecosystem function of seagrass meadows. In Blue Carbon in Shallow Coastal Ecosystems; Kuwae, T., Hori, M., Eds.; Springer: Singapore, 2018; pp. 33–71. [Google Scholar]
- Miyajima, T.; Hamaguchi, M.; Hori, M. Evaluation of the baseline carbon sequestration rates of Indo-Pacific temperate and tropical seagrass meadow sediments. Ecol. Res. 2021, 37, 9–20. [Google Scholar] [CrossRef]
- Yoshida, G.; Hori, M.; Shimabukuro, H.; Hamaoka, H.; Yatsuya, K.; Watanabe, K.; Nakaoka, M. Carbon sequestration by seagrass and macroalgae in Japan: Estimates and future Needs. In Blue Carbon in Shallow Coastal Ecosystems; Kuwae, T., Hori, M., Eds.; Springer: Singapore, 2018; pp. 101–128. [Google Scholar]
- Kokubu, Y.; Komatsu, T. Fate of floating seaweeds: A case in waters off southeast Hokkaido island, Japan. GekkanKiyou 2015, 47, 265–270. (In Japanese) [Google Scholar]
- Shimabukuro, H.; Yoshida, G.; Miura, S.; Terawaki, T. Change of vegetation seaweed beds along the stone-built seawall mound in Hiroshima Bay, Hatsukaichi City, Hiroshima Prefecture, Japan. Jpn. J. Phycol. 2015, 63, 1–9. [Google Scholar]
- Wei, N.; Nakajima, F.; Tobino, T. Variation of environment DNA in sediment at different temporal scales in nearshore area of Tokyo Bay. J. Water Environ. Tech. 2019, 3, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Doi, H.; Uchii, K.; Takahara, T.; Matsuhashi, S.; Yamanaka, H.; Minamoto, T. Use of droplet digital PCR for estimation of fish abundance and biomass in environmental DNA surveys. PLoS ONE 2015, 10, e0122763. [Google Scholar] [CrossRef] [Green Version]
- Guardiola, M.; Jesus-Uriz, M.; Taberlet, P.; Coissac, E.; Wangensteen, O.S.; Turon, X. Deep-Sea, deep-sequencing: Metabarcoding extracellular DNA from sediments of marine canyons. PLoS ONE 2015, 10, e0139633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, P.F.; Willerslev, E. Environmental DNA—An emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 2015, 183, 4–18. [Google Scholar] [CrossRef]
- Hamaguchi, M.; Shimabukuro, H.; Usuki, H.; Hori, M. Occurrences of the indo-west pacific rock oyster Saccostrea cucullata in mainland Japan. Mar. Biodivers. Rec. 2014, 7, e84. [Google Scholar] [CrossRef]
- Hamaguchi, M.; Manabe, M.; Kajihara, N.; Shimabukuro, H.; Yamada, Y.; Nishi, E. DNA barcoding of flat oyster species reveals the presence of Ostrea stemtina Payraudeau, 1826 (Bivalvia: Ostreidae) in Japan. Mar. Biodivers. Rec. 2017, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Reich, S.; DiMartinob, E.; Toddb, J.A.; Wesselingh, F.P.; Renema, W. Indirect paleo-seagrass indicators (IPSIs): A review. Earth-Sci. Rev. 2015, 143, 161–186. [Google Scholar] [CrossRef]
- Geraldi, N.R.; Ortega, A.; Serrano, O.; Macreadie, P.I.; Lovelock, C.E.; Krause-Jensen, D.; Kennedy, H.; Lavery, P.S.; Pace, M.L.; Kaal, J.; et al. Fingerprinting blue carbon: Rationale and tools to determine the source of organic carbon in marine depositional environments. Front. Mar. Sci. 2019, 6, 263. [Google Scholar] [CrossRef] [Green Version]
- Miya, M.; Sato, Y.; Fukunaga, T.; Sado, T.; Poulsen, J.Y.; Sato, K.; Minamoto, T.; Yamamoto, S.; Yamanaka, H.; Araki, H.; et al. MiFish, a set of universal PCR primers for metabarcoding environmental DNA from fishes: Detection of more than 230 subtropical marine species. R. Soc. Open Sci. 2015, 2, 150088. [Google Scholar] [CrossRef] [Green Version]
- Kress, W.J.; Erickson, D.L. A two-locus global DNA barcode for land plants: The coding rbcL gene complements the non-coding trnH-psbA spacer region. PLoS ONE 2007, 6, e508. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Huang, J.; Xu, Y. The cellulose synthase superfamily in fully sequenced plants and algae. BMC. Plant Biol. 2009, 9, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, Y.; Ohi-Toma, T.; Murata, J.; Tanaka, N. Hybridization and polyploidy of an aquatic plant, Ruppia (Ruppiaceae), inferred from plastid and nuclear DNA phylogenies. Am. J. Bot. 2010, 97, 1156–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; De La Torre, A.R.; Sterck, L.; Canovas, F.M.; Avila, C.; Merino, I.; Carvera, M.T.; Ingvarsson, P.K.; Van de Peer, Y. Single-copy genes as molecular markers for phylogenomic studies in seed plants. Genome Biol. Evol. 2017, 9, 1130–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Japanese Food Guide. Available online: https://www.japanesefoodguide.com/edible-japanese-seaweed-types (accessed on 1 February 2022).
- Sato-Umi Net. Available online: https://www.env.go.jp/water/heisa/satoumi/en/02_e.html (accessed on 1 February 2022).
- Yamamuro, M.; Hiratsuka, J.-I.; Ishitobi, Y.; Hosokawa, S.; Nakamura, Y. Ecosystem shift resulting from loss of eelgrass and other submersed aquatic vegetation in two estuarine lagoons, Lake Nakaumi and Lake Shinji, Japan. J. Oceanog. 2006, 62, 551–558. [Google Scholar] [CrossRef]
- Murata, M.; Nakazoe, J.-I. Production and use of marine algae in Japan. Jpn. Agric. Res. Q. (JARQ) 2001, 35, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Sondak, C.F.A.; Ang, P.O., Jr.; Beardall, J.; Bellgrove, A.; Boo, S.M.; Gerung, G.S.; Hepburn, C.D.; Hong, D.D.; Hu, Z.; Kawai, H.; et al. Carbon dioxide mitigation potential of seaweed aquaculture beds (SABs). J. Appl. Phycol. 2017, 29, 2363–2373. [Google Scholar] [CrossRef]
- Brakel, J.; Sibonga, R.C.; Dumilag, R.V.; Montalescot, V.; Campbell, I.; Cottier-Cook, E.J.; Le Ward, V.; Liu, T.; Msuya, F.E.; Brodie, J.; et al. Exploring, harnessing and conserving marine genetic resources towards a sustainable seaweed aquaculture. Plants People Planet 2021, 3, 337–349. [Google Scholar] [CrossRef]
- Nobutoki, M.; Yoshihara, S.; Kuwae, T. Carbon offset utilizing coastal waters: Yokohama blue carbon project. In Blue Carbon in Shallow Coastal Ecosystems; Kuwae, T., Hori, M., Eds.; Springer: Singapore, 2018; pp. 321–346. [Google Scholar]
- Halpern, B.S.; Longo, C.; Hardy, D.; McLeod, K.L.; Samhouri, J.F.; Katona, S.K.; Kleisner, K.; Lester, S.E.; O’Leary, J.; Ranelletti, M.; et al. An index to assess the health and benefits of the global ocean. Nature 2012, 488, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Akhand, A.; Watanabe, K.; Chanda, A.; Tokoro, T.; Chakraborty, K.; Moki, H.; Tanaya, T.; Ghosh, J.; Kuwae, T. Lateral carbon fluxes and CO2 evasion from a subtropical mangrove-seagrass-coral continuum. Sci. Total Environ. 2021, 752, 142190. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Species | Target Gene | Abbreviations for DLPs and PCR Primers | Nucleotide Sequences 5′–3′ | Reaction Temperature (°C) | Amplicon Size (bp) |
|---|---|---|---|---|---|
| Rhizophora mucronata | ITS | YaeITS-P | Cy5-GCGCTGCGACTCCACGATGAGTACC-BHQ3 | 64.0 | 116 |
| YaeITS-F | TCGCGCCAAGGAAATCAAAGA | ||||
| YaeITS-R | ATATCCGTTGCCGAGAGTCG | ||||
| MatK | YehiruMatK-P | FAM-TCATCAGAAGAGGCGTACACTTTGAAGCCA-BHQ1 | 64.0 | 169 | |
| YehiruMatK-F | GTCTTTGCTAATGATTTTCCGTCT | ||||
| YehiruMatK-R | ACCACACATAAAAATGACATTGACC | ||||
| Bruguiera gymnorrhiza | ITS | OhiITS-P | HEX-GGCCGCTGCGTCTCCACCATCA-BHQ1 | 64.0 | 114 |
| OhiITS-F | GCGCCAAGGAAATCACAGG | ||||
| OhiITS-R | GATATCCGTTGCCGAGAGTCG | ||||
| MatK | OhiMatK-P | HEX-AAGAACCCGCGCTTCTTCCGTCAAAAA-BHQ1 | 64.0 | 142 | |
| OhiMatK-F | CTTGTAAACACAAAAGTCCTGTCC | ||||
| OhiMatK-R | CCTCGATATAACGTTTGTGAAATAGAAG | ||||
| Sonneratia alba | ITS | MayaITS-P | FAM-ACCTCGCCTTCTCCAGTGTGACAATGATGA-BHQ1 | 64.0 | 101 |
| MayaITS-F | CGGCACTGGATGCTTCCC | ||||
| MayaITS-R | GTGCGAGAGCCAAGATATCCA | ||||
| MatK | MayaMatK-P | Cy5-CCCGTTAGTAAGTCGACCCGGACCGAT-BHQ3 | 64.0 | 124 | |
| MayaMatK-F | TTCCCGTTATTCCTTGATTGGATGC | ||||
| MayaMatK-R | CGCACAAATCGGTCGATAATATCA |
| Target Species | Target Gene | Abbreviations for DLPs and PCR Primers | Nucleotide Sequences 5′–3′ | Reaction Temperature (°C) | Amplicon Size (bp) |
|---|---|---|---|---|---|
| Enhalus acoroides | ITS | Us-ITS-P | HEX-ACCGTTCGTTTCCACGTCACTCGCTC-BHQ1 | 64.0 | 99 |
| Us-ITS-F | ACCTGCGGAAGGATCATTGTC | ||||
| Us-ITS-R | AGGGTTGTTGTTCGGTGGTC | ||||
| MatK | UmishyoB-P | FAM-ACCCGGATACATCACAAAATTGAGCTTTCG-BHQ1 | 64.0 | 98 | |
| UmishyoB-F | AGTCCCAGTTATTCCTCTCATTGA | ||||
| UmishyoB-R | CGGATAAATCCGTCCAAATCCC | ||||
| Cymodocea rotundata | ITS | Ba-ITS-P | FAM-CGTTCGTTACGTTGGCTACCACTCTCCCT-BHQ1 | 64.0 | 91 |
| Ba-ITS-F | TGCCCATCTCGGAGTTCGTG | ||||
| Ba-ITS-R | GTGGACTCGCCTGACCTGG | ||||
| MatK | BeniA-P | Cy5-CCAAACTGGCTTACTAATGGGATACCCGGA-BHQ3 | 64.0 | 120 | |
| BeniA-F | CCCTCTCATTGGATCTTTATCTAAAGA | ||||
| BeniA-R | CATATACGACCAAACCGATCAATAATA | ||||
| Thalassia hemprichii | ITS | Rs-ITS-P | Cy5-CATTCACCACACGTCGGGATGCACC-BHQ3 | 64.0 | 108 |
| Rs-ITS-F | GCCGTCCCTGTGTTCCCTA | ||||
| Rs-ITS-R | TCACGACGCACGAAGCAC | ||||
| MatK | RyukyuS-P | HEX-AACCCGTCCAAATCGCCTTACTAACAGGA-BHQ1 | 64.0 | 122 | |
| RyukyuS-F | TTCCTCTCATTGGATCCTTGTCA | ||||
| RyukyuS-R | ACATATCCGACCAAATCGATCAATC | ||||
| Halophila ovalis Iriomote-type | ITS | Hal-ITS-P | Cy5-ACGTGAAGCAACGAGTGCGATCCACC-BHQ3 | 64.0 | 117 |
| Hal-ITS-F | TCTCGACGATGTCCTGCCTC | ||||
| Hal-ITS-R | GGGGATACTCAGCTTGGGGA | ||||
| Cymodocea serrulata | ITS | Ra-ITS-P | FAM-CACTTGAGCACACCGCTTCCACTCCAC-BHQ1 | 64.0 | 93 |
| Ra-ITS-F | GGCCTTCCGGTTCTCTCTCT | ||||
| Ra-ITS-R | CATCGCATCGCATCCATCGA | ||||
| Halodule uninervis | ITS | Umj-ITS-P | HEX-TTGCCATCCACCATCCATGCTGCGTC-BHQ1 | 64.0 | 98 |
| Umj-ITS-F | GGCTGAAGTACGTTGGGCTC | ||||
| Umj-ITS-R | GGCGATCCAAGGGAAGCATC |
| Target Species | Target Gene | Abbreviations for DLPs and PCR Primers | Nucleotide Sequences 5′–3′ | Reaction Temperature (°C) | Amplicon Size (bp) |
|---|---|---|---|---|---|
| Zostera marina | ITS | AmaITS-P3 | FAM-ACCCACCATGCCATGTACCGAACATGC-BHQ1 | 64.0 | 122 |
| AmaITS-F3 | TGTAAAGAATCAGAGAATGACCTTC | ||||
| AmaITS-R3 | CAGTTTCAGAATGGTAAACATTCTAA | ||||
| Zostera caulescens | ITS | TachiITS-P3 | HEX-CGTTTGCCTTGGCAACAATTGTGCCGTG-BHQ1 | 64.0 | 129 |
| TachiITS-F31 | CTTAAAGGATGCACAATCAAGT | ||||
| TachiITS-R3 | ATCGAATGAATACGTTTCACCA | ||||
| Phyllospadix iwatensis | ITS | Sugamo-P | Cy5-AGCCTGCGTGTCGTGCCGTGTAGCGCAGTGTAG-BHQ3 | 62.0 | 103 |
| Sugamo-F | GTGGATTGTTGCAGACGGTTTGTC | ||||
| Sugamo-R | ACAACGACAGATGGCGCACTAAG |
| Target Species | Target Gene | Abbreviations for DLPs and PCR Primers | Nucleotide Sequences 5′–3′ | Reaction Temperature (°C) | Amplicon Size (bp) |
|---|---|---|---|---|---|
| Undaria pinnatifida | ITS | WakameITS-P | HEX-TCGCCCAACATCGCGTAAC-BHQ1 | 56.0 | 83 |
| WakameITS-F | GCGTCGGTTTTGTAAA | ||||
| WakameITS-R | TCGAGGGAATTAACCC | ||||
| Saccharina japonica | ITS | MakonbuITS-P | FAM-CGAGGCGCCCCTCGCCCAACTTCGCA-BHQ1 | 68.0 | 116 |
| MakonbuITS-F | TATAAATTGTCTGTGAGGCCGCTTCGT | ||||
| MakonbuITS-R | TGATTCGAGGGCCTTTTCACAGGCT | ||||
| Saccharina longissima | ITS | NagaKoCo1-P | HEX-GCTGGGACAGGTTGGACGGTGTACCCACCT-GHQ1 | 68.0 | 163 |
| NagaKoCo1-F | CCTCTTTAATTTTGCTTCTAGCGTCTTCATT | ||||
| NagaKoCo1-R | CCTAAAATAGAAGCAGCACCCGAGAG | ||||
| Ecklonia kurome | ITS | KuromeITS-P | FAM-CGCCTCCCTCGGGTTTTAATTA-BHQ1 | 65.0 | 101 |
| KuromeITS-F | CGTTTGTAACCTCATCTTT | ||||
| KuromeITS-R | GCCCAACTTCGCATAA | ||||
| Sargassum horneri | ITS | AkamokuITS2-P | HEX-AGCCTCTAGCAACGCTCCAA-BHQ1 | 58.0 | 106 |
| AkamokuITS2-F | TCGCTATATGCAGGTTTA | ||||
| AkamokuITS2-R | GACTGCCTACCGTCAA | ||||
| Sargassum muticum | ITS | TamahaITS2-P | Cy5-TGTCATCAGCGCCGCAAAG-BHQ3 | 58.0 | 122 |
| TamahaITS2-F | GGTGGGTATTTTTGTACC | ||||
| TamahaITS2-R | GGAAGACACGGGTTAA |
| Target Species Complex | Target Gene | Abbreviations for DLPs and PCR Primers | Nucleotide Sequences 5′–3′ | Reaction Temperature (°C) | Amplicon Size (bp) |
|---|---|---|---|---|---|
| Sargassum spp. | ITS | SubSar-DLP | FAM-GGTGGACTCAGGGGACGAGCAGG-BHQ1 | 62.0 | 151 |
| SubSar-F | GKGTTCGATCTCGATCTCAAG | ||||
| SubSar-R | CAAAGACAATAGAAGCCTGGACAAT | ||||
| Bactrophycus spp. | ITS | TemSar-DLP | HEX-CGACCCGTCGTACAACGGATCCTC-BHQ1 | 62.0 | 102 |
| TemSar-F | TGTGCGGGTGAGTTTGAAG | ||||
| TemSar-R | CAAACTCACCCGCGYACAT | ||||
| Ulva spp. | ITS | Ulva-ITS-P | FAM-CGGATATCTTGGCTCTCGCAACGATGAAGAACGC-BHQ1 | 65.0 | 159 |
| Ulva-ITS-F | CTGAAGCAGCTTCGYAMGGGGACAC | ||||
| Ulva-ITS-R | AATGTGCGTTCAAGATTCGATGACTC | ||||
| Neopyropia spp. | ITS | Susabi-ITS-P | HEX-TGGGCGTTGCCCTCTGGAACGTGCT-BHQ1 | 65.0 | 105 |
| Susabi-ITS-F2 | TCTGACGTAGAGACAGGTGCCGTC | ||||
| Susabi-ITS-R2 | CCGTCAAGCACAATCTGCCTCTTTTGA | ||||
| Ecklonia/Eisenia spp. | ITS | EckEiseITS-P | FAM-TTCTCGGGGTATAAACGCTCGCCTCCCTCGG-BHQ1 | 66.0 | 119 |
| EckEiseITS-F | GTCTGAGACGTCGCCGTTTGTAACCTCA | ||||
| EckEiseITS-R | CCCCTCGCCCAACTTCGCATAACAAA | ||||
| Saccharina spp. | ITS | SacchaITS-P | HEX-GTCGCGGCGGCGGACTTTGAGTGTTCCG-BHQ1 | 66.0 | 150 |
| SacchaITS-F | ACTCGCCCCTCTTCTCTCCTGTCTCA | ||||
| SacchaITS-R | GAAGCGAGCGCCGTCAACAACTCTG |
| Group | Target Species or Species Complex | Gene | Sloop | Efficiency | R2 | DNA Polymerase |
|---|---|---|---|---|---|---|
| Mangrove | Rhizophora mucronata | ITS | −3.60 | 0.89 | 0.99 | B |
| MatK | −3.41 | 0.96 | 0.99 | A,B | ||
| Bruguiera gymnorrhiza | ITS | −3.64 | 0.88 | 0.99 | B | |
| MatK | −3.06 | 1.12 | 0.99 | A,B | ||
| Sonneratia alba | ITS | −3.34 | 0.99 | 0.99 | A,B | |
| MatK | −3.33 | 1.00 | 0.99 | A,B | ||
| Seagrass (subtropical zone) | Enhalus acoroides | ITS | −3.29 | 1.02 | 0.99 | A,B |
| MatK | −3.32 | 1.00 | 0.99 | A,B | ||
| Cymodocea rotundata | ITS | −3.42 | 0.96 | 0.99 | A,B | |
| MatK | −3.45 | 0.95 | 1.00 | A,B | ||
| Thalassia hemprichii | ITS | −3.28 | 1.02 | 0.99 | A,B | |
| MatK | −3.24 | 1.04 | 0.99 | A,B | ||
| Halophila ovalis (Iriomote-type) | ITS | −3.44 | 0.95 | 0.99 | A,B | |
| Cymodocea serrulata | ITS | −3.34 | 0.99 | 0.99 | A,B | |
| Halodule uninervis | ITS | −3.39 | 0.97 | 1.00 | A,B | |
| Seagrass (temperate and subarctic zone) | Zostera marina | ITS | −3.40 | 0.97 | 0.99 | A,B |
| Zostera caulescens | ITS | −3.24 | 1.04 | 0.99 | A,B | |
| Phyllospadix iwatensis | ITS | −3.31 | 1.01 | 1.00 | A,B | |
| Phaeophyceae (single species) | Undaria pinnatifida | ITS | −3.51 | 0.93 | 0.99 | B |
| Saccharina japonica | ITS | −3.42 | 0.96 | 1.00 | B | |
| Saccharina longissima | COI | −3.34 | 0.99 | 0.99 | A,B | |
| Ecklonia kurome | ITS | −3.42 | 0.96 | 1.00 | A,B | |
| Sargassum horneri | ITS | −3.49 | 0.94 | 0.99 | A,B | |
| Phaeophyceae, Rhodophyta and Chlorophyta (Species complex) | Sargassum spp. | ITS | −3.47 | 0.94 | 0.99 | A,B |
| Bactrophycus spp. | ITS | −3.40 | 0.97 | 0.99 | A,B | |
| Ulva spp. | ITS | −3.40 | 0.97 | 0.99 | A,B | |
| Neopyropia spp. | ITS | −3.40 | 0.97 | 1.00 | A,B | |
| Ecklonia/Eisenia spp. | ITS | −3.37 | 0.98 | 0.99 | A,B | |
| Saccharina spp. | ITS | −3.21 | 1.05 | 0.99 | A,B | |
| Internal standard | Crassostrea zhanjiangensis | COI | −3.46 | 0.95 | 0.99 | A,B |
| Ammodytes hexapterus | CYTB | −3.48 | 0.97 | 0.99 | A,B | |
| DNA polymerase | A:SsoFast Probes Supermix with ROX, B:iQ Multiplex Powermix | |||||
| Sampling Site | Sampling Position | Total Nitrogen (µmol N/g) | Organic Coarbon (µmol C/g) | C/N Ratio | Nitrogen Stable Isotope Ratio (d15N) | Carbon Stable Isotope Ratio (d13C) | Specific Surface Area (m2/g) | Total Pore Volume (cm3/g) | Average Pore Diameter (nm) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Longitude | Latitude | |||||||||
| HSd-001 | 34-13.3 | 132-28.1 | 220 | 2,130 | 9.66 | 7.75 | −21.29 | 26.02 | 0.119 | 18.30 |
| HSd-003 | 34-18.2 | 132-24.0 | 210 | 1,863 | 8.87 | 8.07 | −21.04 | 33.99 | 0.146 | 17.16 |
| HSd-005 | 33-57.2 | 132-26.1 | 17 | 117 | 6.87 | 9.11 | −19.95 | 3.60 | 0.015 | 16.45 |
| HSd-007 | 33-54.2 | 132-20.0 | 52 | 512 | 9.76 | 7.21 | −21.74 | 5.64 | 0.028 | 19.84 |
| HSd-009 | 34-01.3 | 132-20.3 | 190 | 1,517 | 7.98 | 7.85 | −20.86 | 32.09 | 0.123 | 15.39 |
| HSd-011 | 34-07.6 | 132-20.3 | 183 | 1,479 | 8.07 | 7.84 | −20.82 | 31.71 | 0.113 | 14.24 |
| HSd-013 | 34-12.4 | 132-19.5 | 206 | 1,705 | 8.30 | 7.93 | −20.85 | 35.03 | 0.110 | 12.58 |
| HSd-015 | 34-13.7 | 132-14.0 | 258 | 2,658 | 10.30 | 6.19 | −22.46 | 31.84 | 0.106 | 13.29 |
| Sampling Site | Organic Carbon Derived from Z. marina (µmol C/g) | |||
|---|---|---|---|---|
| 4-Source Model | SD* | 3-Source Model | SD* | |
| HSd-001 | 253 | 185 | 407 | 211 |
| HSd-003 | 235 | 168 | 397 | 175 |
| HSd-005 | 19 | 12 | 34 | 10 |
| HSd-007 | 56 | 41 | 84 | 50 |
| HSd-009 | 200 | 140 | 343 | 141 |
| HSd-011 | 195 | 136 | 334 | 139 |
| HSd-013 | 225 | 157 | 384 | 162 |
| HSd-015 | 255 | 189 | 391 | 231 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamaguchi, M.; Miyajima, T.; Shimabukuro, H.; Hori, M. Development of Quantitative Real-Time PCR for Detecting Environmental DNA Derived from Marine Macrophytes and Its Application to a Field Survey in Hiroshima Bay, Japan. Water 2022, 14, 827. https://doi.org/10.3390/w14050827
Hamaguchi M, Miyajima T, Shimabukuro H, Hori M. Development of Quantitative Real-Time PCR for Detecting Environmental DNA Derived from Marine Macrophytes and Its Application to a Field Survey in Hiroshima Bay, Japan. Water. 2022; 14(5):827. https://doi.org/10.3390/w14050827
Chicago/Turabian StyleHamaguchi, Masami, Toshihiro Miyajima, Hiromori Shimabukuro, and Masakazu Hori. 2022. "Development of Quantitative Real-Time PCR for Detecting Environmental DNA Derived from Marine Macrophytes and Its Application to a Field Survey in Hiroshima Bay, Japan" Water 14, no. 5: 827. https://doi.org/10.3390/w14050827