
Role of Ecohydrographical Barriers on the Spatio-Temporal Distribution of Chaetognath Community in the Gulf of Aqaba during Summer

 ,
,
Abstract
:1. Introduction
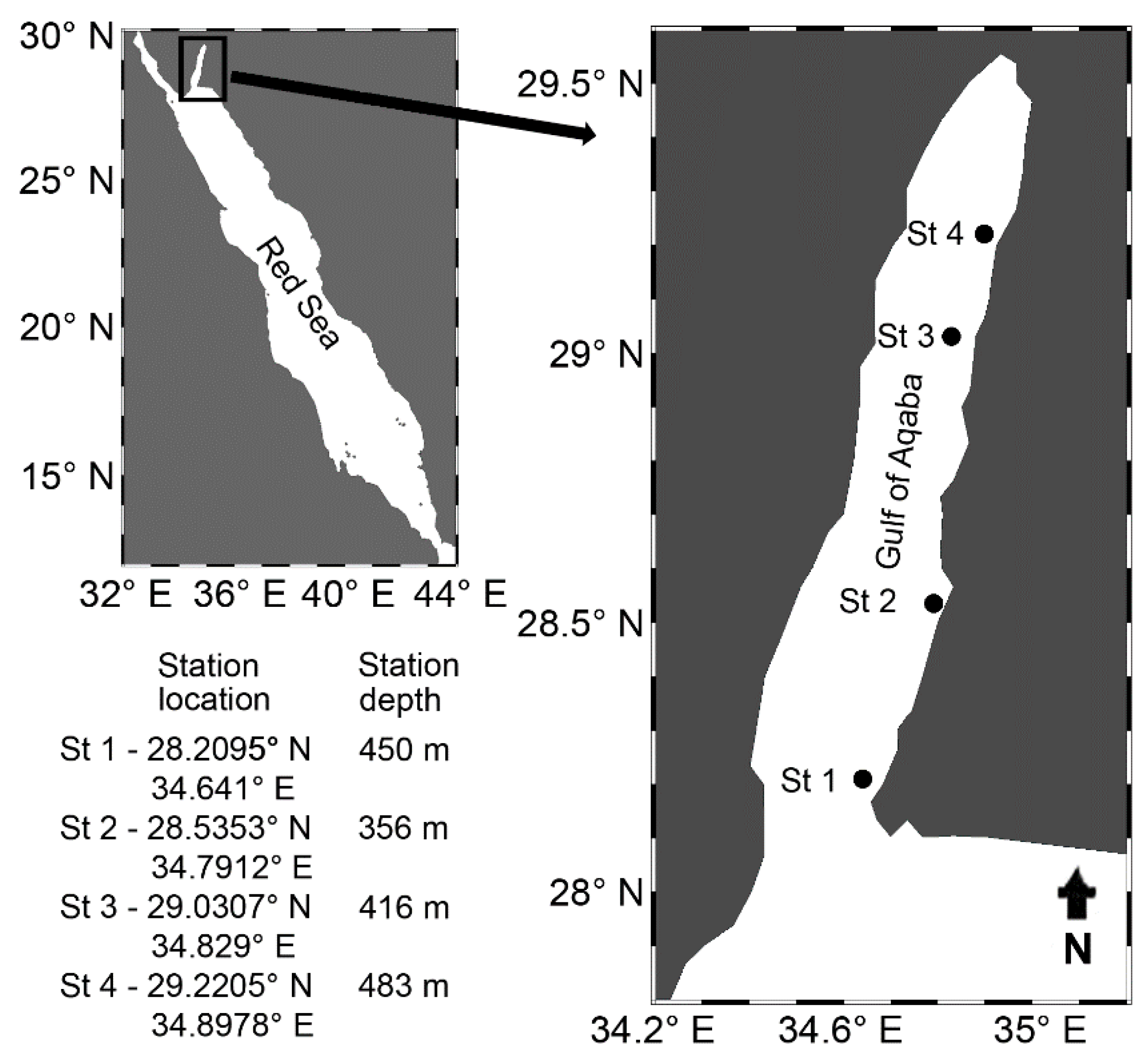
2. Materials and Methods
2.1. Collections
2.2. Data Analyses
3. Results
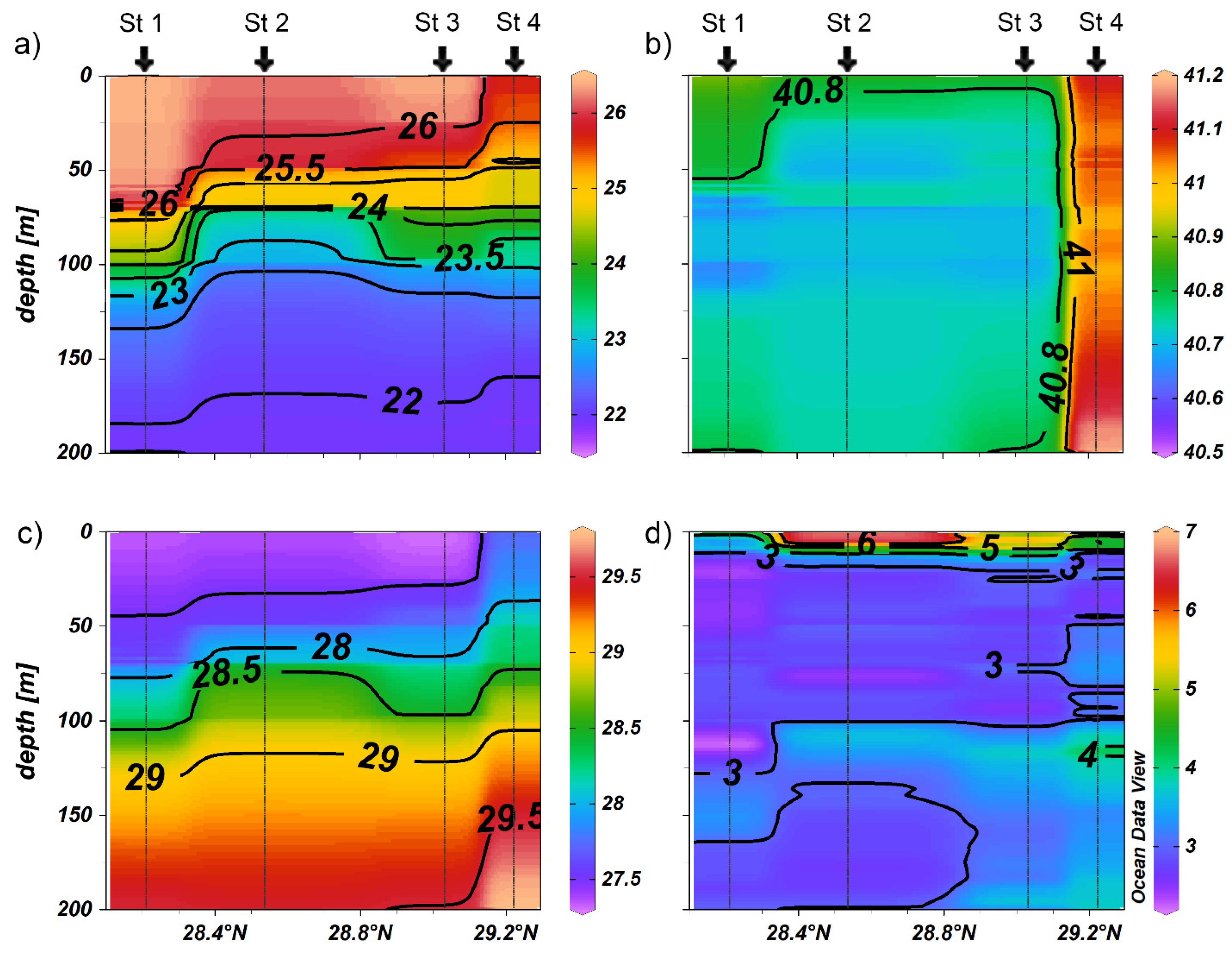
3.1. Physico-Chemical Environment
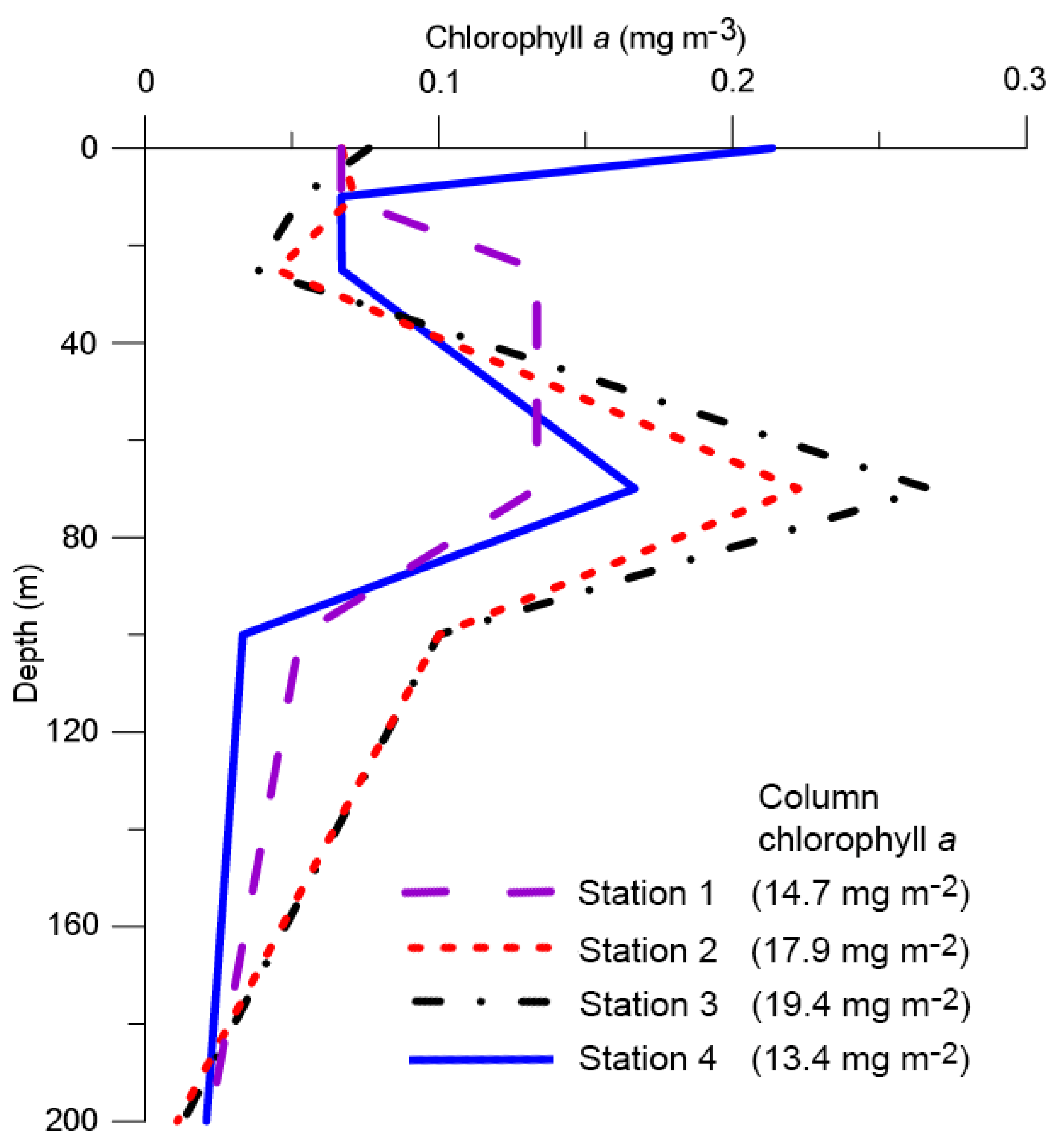
3.2. Phytoplankton Biomass (Chlorophyll a)
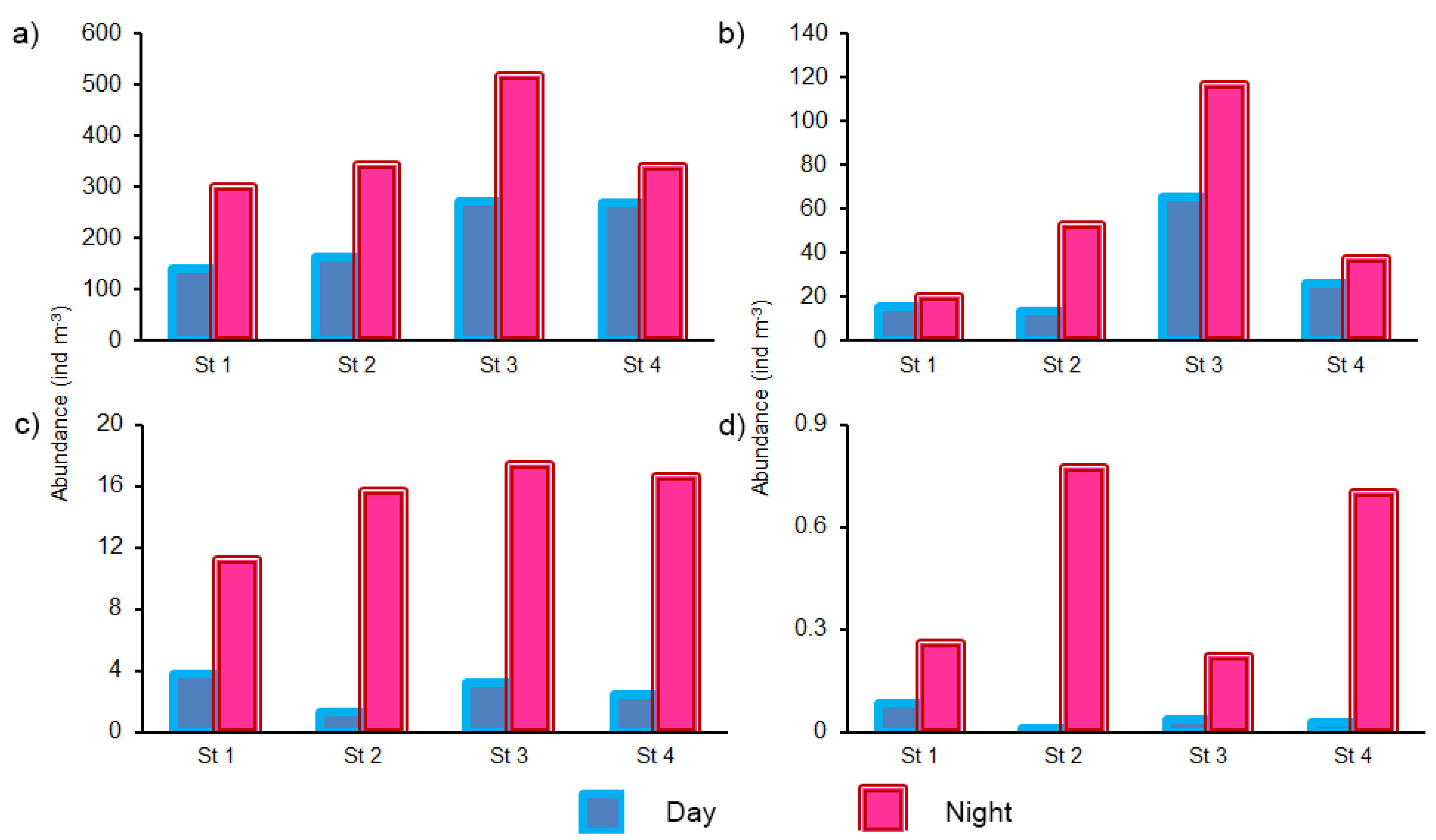
3.3. Abundance of Copepod
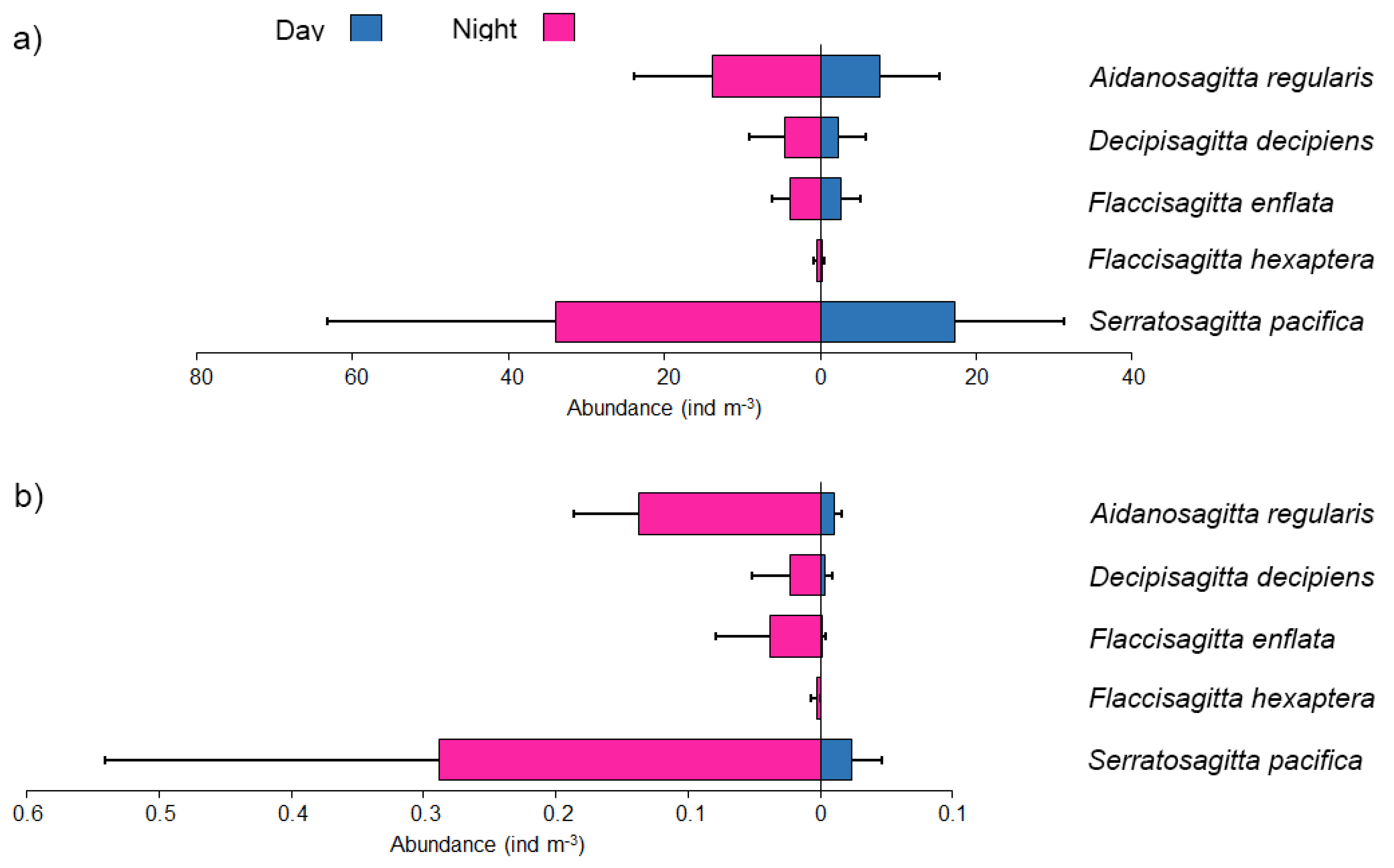
3.4. Composition and Abundance of Chaetognath
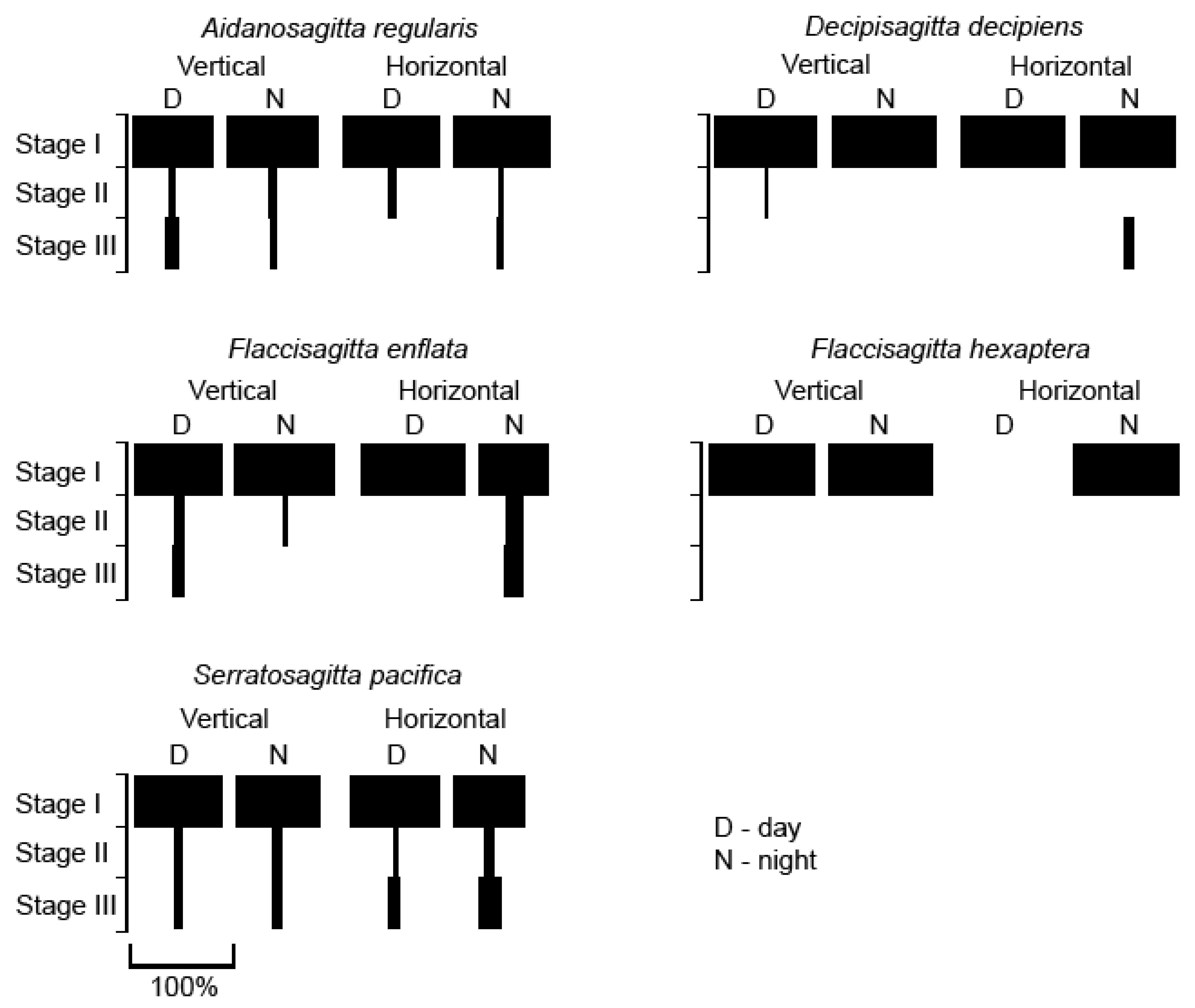
3.5. Maturity Stage-Wise Distribution
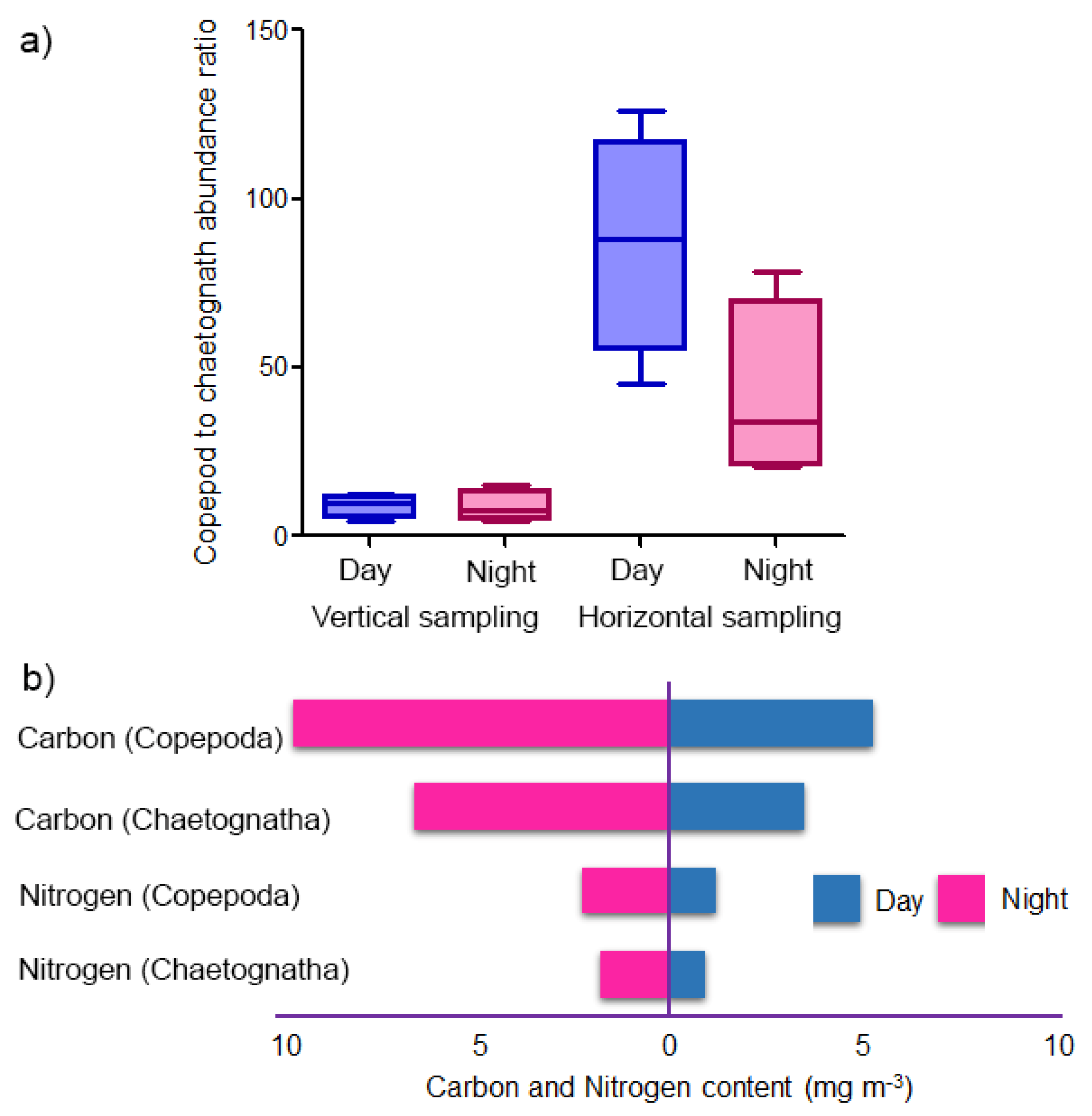
3.6. Copepod–Chaetognath Interrelation
3.7. Species Evenness
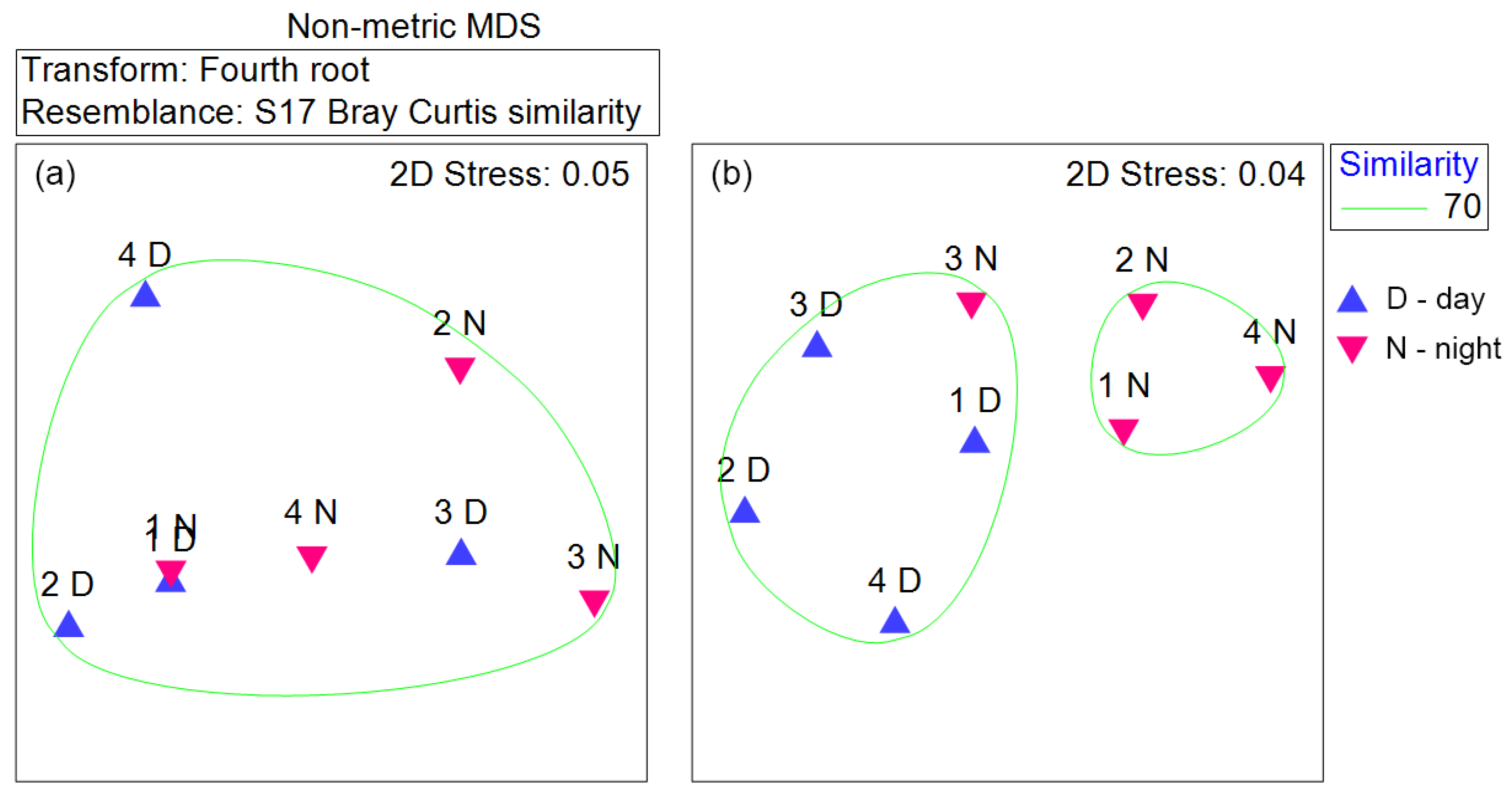
3.8. n-MDS Analysis
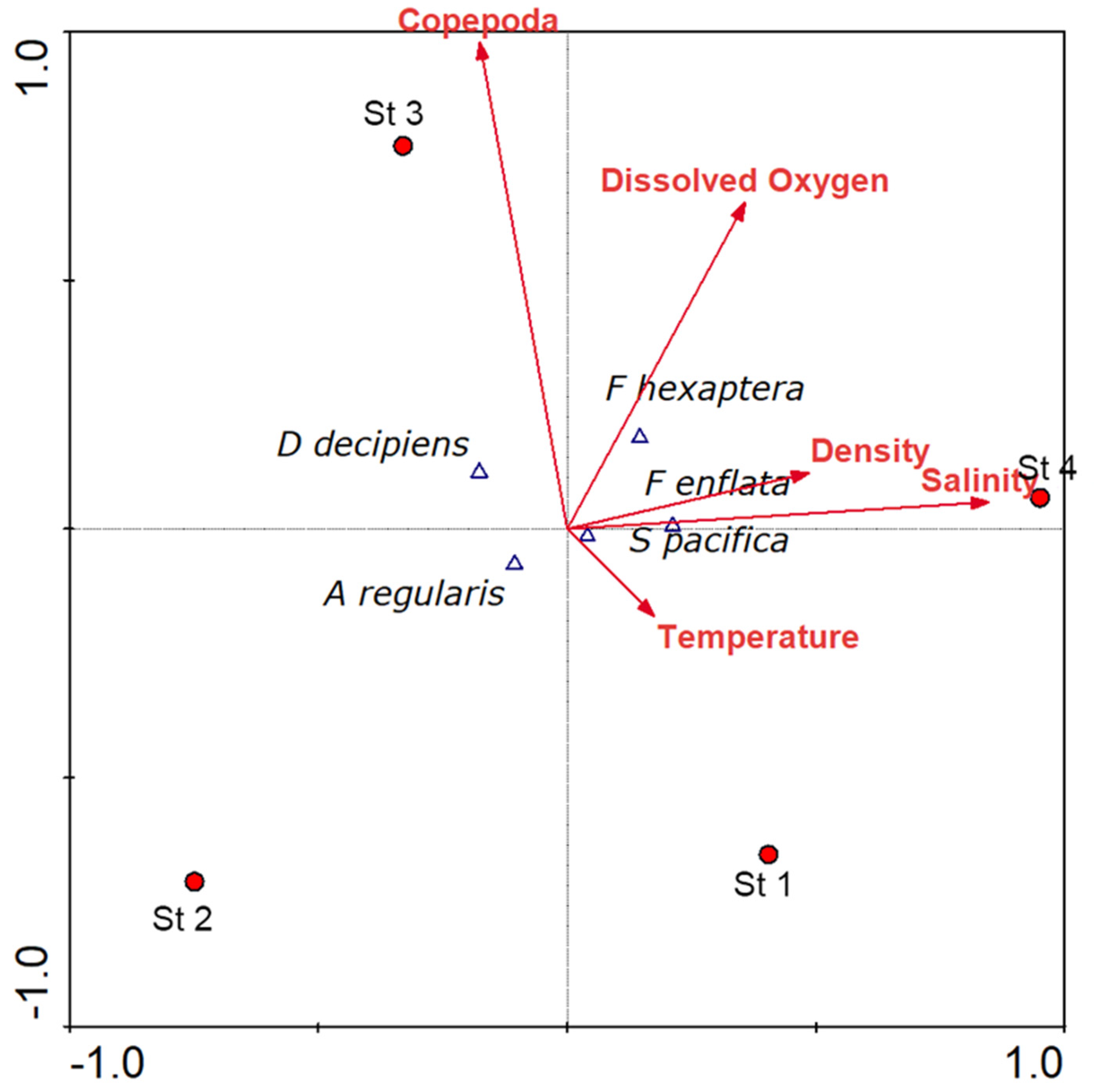
3.9. Canonical Correspondence Analysis (CCA)
4. Discussion
4.1. Role of Hydrography
4.2. Trophic Relationship
4.3. Distinct Nature of Chaetognath Community in the GoA and Their Ecology
4.4. Diel Variability
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cornils, A.; Schnack-Schiel, S.B.; Hagen, W.; Dowidar, M.; Stambler, N.; Plähn, O.; Richter, C. Spatial and temporal distribution of mesozooplankton in the Gulf of Aqaba and the northern Red Sea in February/March 1999. J. Plankton Res. 2005, 27, 505–518. [Google Scholar] [CrossRef]
- Terazaki, M. The role of carnivorous zooplankton, particularly chaetognaths in ocean flux. In Biogeochemical Processes and Ocean Flux in the Western Pacific; Sakai, H., Nozaki, Y., Eds.; TERRAPUB: Tokyo, Japan, 1995; pp. 319–330. [Google Scholar]
- Pakhomov, E.A.; Perissinotto, R.; Froneman, P.W. Predation impact of carnivorous macrozooplankton and micronekton in the Atlantic sector of the Southern Ocean. J. Mar. Syst. 1999, 19, 47–64. [Google Scholar] [CrossRef]
- Kehayias, G. Quantitative aspects of feeding of chaetognaths in the eastern Mediterranean pelagic waters. J. Mar. Biol. Ass. UK 2003, 83, 559–569. [Google Scholar] [CrossRef]
- Casanova, J.P. Similarity of plankton distribution patterns in two nearly land-locked seas: The Mediterranean and the Red Sea. UNESCO Tech. Pap. Mar. Sci. 1986, 49, 42–46. [Google Scholar]
- Al-Aidaroos, A.M.; Karati, K.K.; El-Sherbiny, M.M.; Devassy, R.P.; Kürten, B. Latitudinal environmental gradients and diel variability influence abundance and community structure of Chaetognatha in Red Sea coral reefs. Syst. Biodivers. 2017, 15, 35–48. [Google Scholar] [CrossRef]
- Bieri, R. The distribution of the planktonic chaetognatha in the Pacific and their relationship to the water masses. Limnol. Oceanogr. 1959, 4, 1–28. [Google Scholar] [CrossRef]
- Ulloa, R.; Palma, S.; Silva, N. Bathymetric distribution of chaetognaths and their association with water masses off the coast of Valparaıso, Chile. Deep Sea Res. Part I 2000, 47, 2009–2027. [Google Scholar] [CrossRef]
- Froneman, P.W.; Pakhomov, E.A. Trophic importance of the chaetognaths Eukrohnia hamata and Sagitta gazellae in the pelagic system of the Prince Edward Islands (Southern Ocean). Polar Biol. 1998, 19, 242–249. [Google Scholar] [CrossRef]
- Gibbons, M.J. Diel feeding and vertical migration of Sagitta serratodentata Krohn tasmanica Thomson (Chaetognatha) in the southern Benguela. J. Plankton Res. 1992, 14, 249–259. [Google Scholar] [CrossRef]
- Besiktepe, S.; Unsal, M. Population structure, vertical distribution and diel migration of Sagitta setosa (Chaetognatha) in the south-western part of the Black Sea. J. Plankton Res. 2000, 22, 669–683. [Google Scholar] [CrossRef] [Green Version]
- Conway, D.V.P.; Williams, R. Seasonal population structure, vertical distribution and migration of the chaetognath Sagitta elegans in the Celtic Sea. Mar. Biol. 1986, 93, 377–387. [Google Scholar] [CrossRef]
- Pan, J.; Cheng, F.; Yu, F. The diel vertical migration of zooplankton in the hypoxia area observed by video plankton recorder. Indian J. Geo-Mar. Sci. 2018, 47, 1353–1363. [Google Scholar]
- Paldor, N.; Anati, D.A. Seasonal variations of temperature and salinity in the Gulf of Eilat (Aqaba). Deep Sea Res. 1979, 26, 661–672. [Google Scholar] [CrossRef]
- Assaf, G.; Kessler, J. Climate and energy in the Gulf of Aqaba (Elat). Mon. Weather Rev. 1974, 104, 381–385. [Google Scholar] [CrossRef] [Green Version]
- Silverman, J.; Gildor, H. The residence time of an active versus a passive tracer in the Gulf of Aqaba: A box model approach. J. Mar. Syst. 2008, 71, 159–170. [Google Scholar] [CrossRef]
- Klinker, J.; Reiss, Z.; Kropach, C.; Levanon, I.; Harpaz, H.; Shapiro, Y. Nutrients and biomass distribution in the Gulf of Aqaba (Elat) Red Sea. Mar. Biol. 1978, 45, 53–64. [Google Scholar] [CrossRef]
- Manasrah, R.; Lass, H.U.; Fennel, W. Circulation in the Gulf of Aqaba (Red Sea) during winter—Spring. J. Oceanogr. 2006, 62, 219–225. [Google Scholar] [CrossRef]
- Wankel, S.D.; Chen, Y.; Kendall, C.; Post, A.F.; Paytan, A. Sources of aerosol nitrate to the Gulf of Aqaba: Evidence from δ15N and δ18O of nitrate and trace metal chemistry. Mar. Chem. 2010, 120, 90–99. [Google Scholar] [CrossRef]
- Carlson, D.F.; Fredj, E.; Gildor, H.; Biton, E.; Steinbuck, J.V.; Monismith, S.G.; Genin, A. Observations of tidal currents in the Northern Gulf of Eilat/Aqaba (Red Sea). J. Mar. Syst. 2012, 102–104, 14–28. [Google Scholar] [CrossRef]
- Carlson, D.F.; Fredj, E.; Gildor, H. The annual cycle of vertical mixing and restratification in the Northern Gulf of Eilat/Aqaba (Red Sea) based on high temporal and vertical resolution observations. Deep-Sea Res. Part I 2014, 84, 1–17. [Google Scholar] [CrossRef]
- Lindell, D.; Post, A.F. Ultraphytoplankton succession is triggered by deep winter mixing in the Gulf of Aqaba (Eilat), Red Sea. Limnol. Oceanogr. 1995, 40, 1130–1141. [Google Scholar] [CrossRef]
- Labiosa, R.G.; Arrigo, K.R.; Genin, A.; Monismith, S.G.; van Dijken, G. The interplay between upwelling and deep convective mixing in determining the seasonal phytoplankton dynamics in the Gulf of Aqaba: Evidence from SeaWiFS and MODIS. Limnol. Oceanogr. 2003, 48, 2355–2368. [Google Scholar] [CrossRef] [Green Version]
- Al-Najjar, T.; Badran, M.I.; Richter, C.; Meyerhoefer, M.; Sommer, U. Seasonal dynamics of phytoplankton in the Gulf of Aqaba, Red Sea. Hydrobiologia 2007, 579, 69–83. [Google Scholar] [CrossRef]
- Mackey, K.R.; Labiosa, R.G.; Calhoun, M.; Street, J.H.; Post, A.F.; Paytan, A. Phosphorus availability, phytoplankton community dynamics, and taxon-specific phosphorus status in the Gulf of Aqaba, Red Sea. Limnol. Oceanogr. 2007, 52, 873–885. [Google Scholar] [CrossRef] [Green Version]
- Echelman, T.; Fishelson, L. Surface zooplankton dynamics and community structure in the Gulf of Aqaba (Eilat), Red Sea. Mar. Biol. 1990, 107, 179–190. [Google Scholar] [CrossRef]
- Khalil, M.T.; El-Rakman, N.S.A. Abundance and diversity of surface zooplankton in the Gulf of Aqaba, Red Sea, Egypt. J. Plankton Res. 1997, 19, 927–936. [Google Scholar] [CrossRef]
- Farstey, V.; Lazar, B.; Genin, A. Expansion and homogeneity of the vertical distribution of zooplankton in a very deep mixed layer. Mar. Ecol. Prog. Ser. 2002, 238, 91–100. [Google Scholar] [CrossRef]
- El-Sherbiny, M.M.; Reny, P.D.; Muller, E.M.; Al- Sofyani, A.A.; Al-Aidaroos, A.M. Biodiversity and abundance of plankton communities along the coastal waters of Gulf of Aqaba, Saudi Arabia. Pak. J. Zool. 2019, 51, 1823–1836. [Google Scholar] [CrossRef]
- Almeida Prado-Por, M.S. The diversity and dynamics of Calanoida (Copepoda). Oceanol. Acta 1983, 6, 139–145. [Google Scholar]
- Böttger-Schnack, R.; Hagen, W.; Schnack-Schiel, S.B. The microcopepod fauna in the Gulf of Aqaba, northern Red Sea: Species diversity and distribution of Oncaeidae (Poecilostomatoida). J. Plankton Res. 2001, 23, 1029–1035. [Google Scholar] [CrossRef] [Green Version]
- Cornils, A.; Niehoff, B.; Richter, C.; Al-Najjar, T.; Schnack-Schiel, S.B. Seasonal abundance and reproduction of clausocalanid copepods in the northern Gulf of Aqaba (Red Sea). J. Plankton Res. 2006, 29, 57–70. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Biological and Chemical Methods for Seawater Analysis; Pergamon: Oxford, UK, 1984. [Google Scholar]
- Dyson, N.; Jitts, H.R.; Scott, B.D. Techniques for measuring oceanic primary production using radioactive carbon. In Division of Fisheries and Oceanography; CSIRO: Melbourne, Australia, 1965. [Google Scholar]
- Sameoto, D.; Wiebe, P.; Runge, J.; Postel, L.; Dunn, J.; Miller, C.; Coombs, S. Collecting zooplankton. In ICES Zooplankton Methodology Manual; Harris, R., Wiebe, P., Lenz, J., Skjoldal, H.R., Huntley, M.E., Eds.; Academic Press: London, UK, 2000; pp. 55–81. [Google Scholar]
- Tokioka, T. The taxonomical outline of Chaetognatha. Publ. Seto Mar. Biol. Lab. 1965, 12, 335–357. [Google Scholar] [CrossRef]
- Michel, H.B. Chaetognatha of the Carribean sea and adjacent areas. NOAA Technical Rep. NMFS 1984, 15, 1–33. [Google Scholar]
- Mclelland, J.A. An illustrated key to the chaetognatha of the northern Gulf of Mexico with notes on their distribution. Gulf Res. Rep. 1989, 8, 145–172. [Google Scholar] [CrossRef]
- McLaren, I.A. Population and production ecology of zooplankton in Ogac Lake, a landlocked fiord on Baffin Island. J. Fish. Res. Board Can. 1969, 26, 1485–1559. [Google Scholar] [CrossRef]
- Zo, Z. Breeding and growth of the Chaetognath Sagitta elegans in Bedford basin. Limnol. Oceanogr. 1973, 18, 750–756. [Google Scholar] [CrossRef]
- Kürten, B.; Al-Aidaroos, A.M.; Kürten, S.; El-Sherbiny, M.M.; Devassy, R.P.; Struck, U.; Zarokanellos, N.; Jones, B.H.; Hansen, T.; Bruss, G.; et al. Carbon and nitrogen stable isotope ratios of pelagic zooplankton elucidate ecohydrographic features in the oligotrophic Red Sea. Prog. Oceanogr. 2016, 140, 69–90. [Google Scholar] [CrossRef] [Green Version]
- Karati, K.K.; Al-Aidaroos, A.M.; Devassy, R.P.; El-Sherbiny, M.M.; Jones, B.H.; Sommer, U.; Kürten, B. Ecohydrographic control on the community structure and vertical distribution of pelagic Chaetognatha in the Red Sea. Mar. Biol. 2019, 166, 30. [Google Scholar] [CrossRef] [Green Version]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E Ltd.: Plymouth, UK, 2006. [Google Scholar]
- ter Braak, C.J.; Verdonschot, P.F. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 255–289. [Google Scholar] [CrossRef]
- Reiss, Z.; Hottinger, L. The Gulf of Aqaba: Ecological Micropaleontology; Springer Science and Business Media: New York, NY, USA, 2012. [Google Scholar]
- Genin, A.; Lazar, B.; Brenner, S. Vertical mixing and coral death in the Red Sea following the eruption of Mount Pinatubo. Nature 1995, 377, 507–510. [Google Scholar] [CrossRef]
- Berman, T.; Paldor, N.; Brenner, S. Simulation of wind-driven circulation in the Gulf of Elat (Aqaba). J. Mar. Sys. 2000, 26, 349–365. [Google Scholar] [CrossRef]
- Yahel, G.; Post, A.F.; Fabricius, K.; Marie, K.D.; Vaulot, D.; Genin, A. Phytoplankton distribution and grazing near coral reefs. Limnol. Oceanogr. 1998, 43, 551–563. [Google Scholar] [CrossRef]
- Timonin, A.G. The structure of plankton communities of the Indian Ocean. Mar. Biol. 1971, 9, 281–289. [Google Scholar] [CrossRef]
- Terazaki, M. Life history strategy of the chaetognath Sagitta elegans in the World Oceans. Coast. Mar. Sci. 2004, 29, 1–12. [Google Scholar]
- Yoon, H.; Ko, A.R.; Kang, J.H.; Choi, J.K.; Ju, S.J. Diet of chaetognatha Sagitta crassa and S. nagae in the yellow sea inferred from Gut content and fatty acid analyses. Ocean Polar Res. 2016, 38, 5–46. [Google Scholar] [CrossRef] [Green Version]
- Kusum, K.K.; Vineetha, G.; Raveendran, T.V.; Muraleedharan, K.R.; Nair, M.; Achuthankutty, C.T. Impact of oxygen-depleted water on the vertical distribution of Chaetognatha in the northeastern Arabian Sea. Deep Sea Res. Part I 2011, 58, 1163–1174. [Google Scholar] [CrossRef]
- Nair, V.R. Bathymetric distribution of Chaetognatha in the Indian Ocean. Indian J. Mar. Sci. 1978, 7, 276–282. [Google Scholar]
- Kusum, K.K.; Vineetha, G.; Raveendran, T.V.; Nair, V.R.; Muraleedharan, K.R.; Achuthankutty, C.T.; Joseph, T. Chaetognath community and their responses to varying environmental factors in the northern Indian Ocean. J. Plankton Res. 2014, 36, 1146–1152. [Google Scholar] [CrossRef] [Green Version]
- Nair, V.R.; Kusum, K.K.; Gireesh, R.; Nair, M. The distribution of the Chaetognatha population and its interaction with environmental characteristics in the Bay of Bengal and the Arabian Sea. Mar. Biol. Res. 2015, 11, 269–282. [Google Scholar] [CrossRef]
- Buchanan, P.J.; Beckley, L.E. Chaetognaths of the Leeuwin Current system: Oceanographic conditions drive epi-pelagic zoogeography in the south-east Indian Ocean. Hydrobiologia 2016, 763, 81–96. [Google Scholar] [CrossRef] [Green Version]
- Maillard, C.; Soliman, G. Hydrography of the Red Sea and exchange with Indian Ocean in summer. Oceanol. Acta 1986, 9, 249–269. [Google Scholar]
- Furnestin, M.L.; Codaccioni, J.C. Chaetognathes du Nord-Ouest de l’Océan Indien (golfe d’Aden, Mer d’Arabie, golfe d’Oman, golfe Persique). Cah. ORSTOM Sér Océanogr. 1968, 6, 143–171. [Google Scholar]
- Haghi, M.; Savari, A.; Madiseh, S.D.; Zakeri, M. Abundance of pelagic chaetognaths in northwestern Persian Gulf. Plankton Benthos Res. 2010, 5, 44–48. [Google Scholar] [CrossRef] [Green Version]
- Klinker, J.; Reiss, Z.; Kropach, C.; Levanon, I.; Harpaz, H.; Halicz, E.; Assaf, G. Observations on the circulation pattern in the Gulf of Aqaba (Elat), Red Sea. Israel J. Earth-Sci. 1976, 25, 85–103. [Google Scholar]
- Kusum, K.K.; Vineetha, G.; Raveendran, T.V.; Muraleedharan, K.R.; Biju, A.; Achuthankutty, C.T. Influence of upwelling on distribution of chaetognath (zooplankton) in the oxygen deficient zone of the eastern Arabian Sea. Cont. Shelf Res. 2014, 78, 16–28. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, Z.L. Estimating optimal salinity and temperature of chaetognaths. J. Mar. Biol. Assoc. UK 2012, 92, 1399–1407. [Google Scholar] [CrossRef]
- Schmidt, H.E. The vertical distribution and diurnal migration of some zooplankton in the Bay of Eilat (Red Sea). Helgoländer Wiss. Meeresunters. 1973, 24, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Hays, G.C.; Harris, R.P.; Head, R.N. Diel changes in the near-surface biomass of zooplankton and the carbon content of vertical migrants. Deep-Sea Res. Part II 2001, 48, 1063–1068. [Google Scholar] [CrossRef]
- Vineetha, G.; Jyothibabu, R.; Madhu, N.V.; Sooria, P.M.; Shivaprasad, A.; Reny, P.D.; Deepak, M.P. Tidal influence on the diel vertical migration pattern of zooplankton in a tropical monsoonal estuary. Wetlands 2015, 35, 597–610. [Google Scholar] [CrossRef]
- Lie, A.A.; Tse, P.; Wong, C.K. Diel vertical migration and feeding of three species of chaetognaths (Flaccisagitta enflata. Aidanosagitta delicata and Aidanosagitta neglecta) in two shallow, subtropical bays in Hong Kong. J. Plankton Res. 2012, 34, 670–684. [Google Scholar]
- Hays, G.C. A review of the adaptive significance and ecosystem consequences of zooplankton diel vertical migrations. Hydrobiologia 2003, 503, 163–170. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chaetognaths | St 1 Vertical | St 2 Vertical | St 3 Vertical | St 4 Vertical | St 1 Surface | St 2 Surface | St 3 Surface | St 4 Surface |
|---|---|---|---|---|---|---|---|---|
| Aidanosagitta regularis | 5.1 ± 2 | 14 ± 12.3 | 20.7 ± 2.4 | 3.2 ± 1.1 | 0.051 ± 0.054 | 0.071 ± 0.094 | 0.112 ± 0.133 | 0.061 ± 0.076 |
| Decipisagitta decipiens | 0.6 ± 0.1 | 2.9 ± 3 | 9.1 ± 2.3 | 1.2 ± 1.1 | 0.009 ± 0.003 | 0.006 ± 0.007 | 0.005 ± 0.002 | 0.033 ± 0.047 |
| Flaccisagitta enflata | 2 ± 0.3 | 1.3 ± 1 | 3.8 ± 2.9 | 6 ± 0.2 | 0.003 ± 0.0004 | 0.032 ± 0.045 | 0.001 ± 0.001 | 0.043 ± 0.055 |
| Flaccisagitta hexaptera | 0.4 ± 0.2 | 0.04 ± 0.1 | 0.8 ± 0.3 | 0.3 ± 0.5 | 0.002 ± 0.002 | – | – | 0.004 ± 0.006 |
| Serratosagitta pacifica | 9.7 ± 1.5 | 14.8 ± 11.9 | 56.8 ± 28.2 | 21.1 ± 5.3 | 0.106 ± 0.069 | 0.283 ± 0.394 | 0.012 ± 0.006 | 0.222 ± 0.292 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karati, K.K.; Vineetha, G.; Devassy, R.P.; Al-Aidaroos, A.M.; El-Sherbiny, M.M. Role of Ecohydrographical Barriers on the Spatio-Temporal Distribution of Chaetognath Community in the Gulf of Aqaba during Summer. Water 2022, 14, 822. https://doi.org/10.3390/w14050822
Karati KK, Vineetha G, Devassy RP, Al-Aidaroos AM, El-Sherbiny MM. Role of Ecohydrographical Barriers on the Spatio-Temporal Distribution of Chaetognath Community in the Gulf of Aqaba during Summer. Water. 2022; 14(5):822. https://doi.org/10.3390/w14050822
Chicago/Turabian StyleKarati, Kusum Komal, Gopinath Vineetha, Reny P. Devassy, Ali M. Al-Aidaroos, and Mohsen M. El-Sherbiny. 2022. "Role of Ecohydrographical Barriers on the Spatio-Temporal Distribution of Chaetognath Community in the Gulf of Aqaba during Summer" Water 14, no. 5: 822. https://doi.org/10.3390/w14050822