
Multi-Soil-Layering Technology: A New Approach to Remove Microcystis aeruginosa and Microcystins from Water

 ,
,  ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Microcystins in Water: Treatment Methods
2.1. Ozonation/Oxidation
2.2. Filtration and Complexation
Microcystin Immobilization onto Soil Particles
3. Fate of Immobilized Microcystins: Biological Activity
4. Some Eco-Technologies for the Elimination of Cyanobacterial Pollution
5. Multi-Soil-Layering (MSL) Technology
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguilera, A.; Haakonsson, S.; Martin, M.V.; Salerno, G.L.; Echenique, R.O. Bloom-forming cyanobacteria and cyanotoxins in Argentina: A growing health and environmental concern. Limnologica 2018, 69, 103–114. [Google Scholar] [CrossRef]
- Wood, R. Acute animal and human poisonings from cyanotoxin exposure—A review of the literature. Environ. Int. 2016, 91, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Francis, G. Poisonous Australian lake. Nature 1878, 18, 11–12. [Google Scholar] [CrossRef] [Green Version]
- Stewart, I.; Seawright, A.A.; Shaw, G.R. Cyanobacterial Poisoning in Livestock, Wild Mammals and Birds—An overview. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs. Advances in Experimental Medicine and Biology; Hudnell, H.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 619, pp. 613–637. [Google Scholar] [CrossRef]
- Thomas, A.D.; Saker, M.L.; Norton, J.H.; Olsen, R.D. Cyanobacterium Cylindrospermopsis raciborskii as a probable cause of death in cattle in northern Queensland. Aust. Vet. J. 1998, 76, 592–594. [Google Scholar] [CrossRef] [Green Version]
- Falconer, I. Algal Toxins and Human Health. In The Handbook of Environmental Chemistry; Hrubec, J., Ed.; Springer: Berlin/Heidelberg, Germany, 1998; pp. 53–82. [Google Scholar]
- Carmichael, W.W.; Azevedo, S.M.; An, J.S.; Molica, R.J.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human fatalities from cyanobacteria: Chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 2001, 109, 663–668. [Google Scholar] [CrossRef]
- Chen, J.; Xie, P.; Li, L.; Xu, J. First identification of the hepatotoxic microcystins in the serum of a chronically exposed human population together with indication of hepatocellular damage. Toxicol. Sci. 2009, 108, 81–89. [Google Scholar] [CrossRef]
- Kubickova, B.; Babica, P.; Hilscherová, K.; ŠIndlerová, L. Effects of cyanobacterial toxins on the human gastrointestinal tract and the mucosal innate immune system. Environ. Sci. Eur. 2019, 31, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Bouaïcha, N.; Miles, C.; Beach, D.; Labidi, Z.; Djabri, A.; Benayache, N.; Nguyen-Quang, T. Structural Diversity, Characterization and Toxicology of Microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef] [Green Version]
- ElKhalloufi, F.; Oufdou, K.; Lahrouni, M.; Faghire, M.; Peix, A.; Ramírez-Bahena, M.H.; Vasconcelos, V.; Oudra, B. Physiological and antioxidant responses of Medicago sativa-rhizobia symbiosis to cyanobacterial toxins (Microcystins) exposure. Toxicon 2013, 76, 167–177. [Google Scholar] [CrossRef]
- Via-Ordorika, L.; Fastner, J.; Kurmayer, R.; Hisbergues, M.; Dittmann, E.; Komarek, J.; Erhard, M.; Chorus, I. Distribution of Microcystin-Producing and Non-Microcystin-Producing Microcystis sp. in European Freshwater Bodies: Detection of Microcystins and Microcystin Genes in Individual Colonies. Syst. Appl. Microbiol. 2004, 27, 592–602. [Google Scholar] [CrossRef] [Green Version]
- Brient, L.; ben Gamra, N.; Periot, M.; Roumagnac, M.; Zeller, P.; Bormans, M.; Méjean, A.; Ploux, O.; Biegala, I.C. Rapid Characterization of Microcystin-Producing Cyanobacteria in Freshwater Lakes by TSA-FISH (Tyramid Signal Amplification-Fluorescent in Situ Hybridization). Front. Environ. Sci. 2017, 5, 43. [Google Scholar] [CrossRef] [Green Version]
- Overlingė, D.; Toruńska-Sitarz, A.; Kataržytė, M.; Pilkaitytė, R.; Gyraitė, G.; Mazur-Marzec, H. Characterization and Diversity of Microcystins Produced by Cyanobacteria from the Curonian Lagoon (SE Baltic Sea). Toxins 2021, 13, 838. [Google Scholar] [CrossRef] [PubMed]
- Fortin, N.; Aranda-Rodriguez, R.; Jing, H.; Pick, F.; Bird, D.; Greer, C.W. Detection of Microcystin-Producing Cyanobacteria in Missisquoi Bay, Quebec, Canada, Using Quantitative PCR. Appl. Environ. Microbiol. 2010, 76, 5105–5112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oudra, B.; Dadi-El Andaloussi, M.; Vasconcelos, V.M. Identification and quantification of microcystins from a Nostoc muscorum bloom occurring in Oukaïmeden River (High-Atlas mountains of Marrakech, Morocco). Environ. Monit. Assess. 2008, 149, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management, 2nd ed.; CRC Press: London, UK, 2021. [Google Scholar] [CrossRef]
- Stewart, I.; Eaglesham, G.K.; McGregor, G.B.; Chong, R.; Seawright, A.A.; Wickramasinghe, W.A.; Sadler, R.; Hunt, L.; Graham, G. First Report of a Toxic Nodularia spumigena (Nostocales/ Cyanobacteria) Bloom in Sub-Tropical Australia. II. Bioaccumulation of Nodularin in Isolated Populations of Mullet (Mugilidae). Int. J. Environ. Res. Public Health 2012, 9, 2396–2411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, Z.; Bakr, A.; Campos, A.; Vasconcelos, V.; Nasr SA, M. Growth inhibition and microcystin accumulation in bush bean (Phaseolus vulgaris L.) plant irrigated with water containing toxic Chrooccocus minutus. Agric. Water Manag. 2022, 261, 107381. [Google Scholar] [CrossRef]
- Mohamed, Z.A. Breakthrough of Oscillatoria limnetica and microcystin toxins into drinking water treatment plants—Examples from the Nile River, Egypt. Water SA 2016, 42, 161. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Guillén, R.; Prieto, A.I.; Moreno, I.; Vasconcelos, V.M.; Moyano, R.; Blanco, A.; Cameán Fernandez, A.M. Cyanobacterium producing cylindrospermopsin cause histopathological changes at environmentally relevant concentrations in subchronically exposed tilapia (Oreochromis niloticus). Environ. Toxicol. 2013, 30, 261–277. [Google Scholar] [CrossRef]
- Nguyen TT, L.; Hoang, T.H.; Nguyen, T.K.; Duong, T.T. The occurrence of toxic cyanobacterium Cylindrospermopsis raciborskii and its toxin cylindrospermopsin in the Huong River, Thua Thien Hue province, Vietnam. Environ. Monit. Assess. 2017, 189, 490. [Google Scholar] [CrossRef]
- Osswald, J.; Rellán, S.; Gago-Martinez, A.; Vasconcelos, V. Production of anatoxin-a by cyanobacterial strains isolated from Portuguese fresh water systems. Ecotoxicology 2009, 18, 1110–1115. [Google Scholar] [CrossRef]
- Casali, S.P.; Dos Santos, A.C.A.; de Falco, P.B.; Calijuri, M.D.C. Influence of environmental variables on saxitoxin yields by Cylindrospermopsis raciborskii in a mesotrophic subtropical reservoir. J. Water Health 2017, 15, 509–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harr, K.E.; Szabo, N.J.; Cichra, M.; Phlips, E.J. Debromoaplysiatoxin in Lyngbya-dominated mats on manatees (Trichechus manatus latirostris) in the Florida King’s Bay ecosystem. Toxicon 2008, 52, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Walls, J.T.; Wyatt, K.H.; Doll, J.C.; Rubenstein, E.M.; Rober, A.R. Hot and toxic: Temperature regulates microcystin release from cyanobacteria. Sci. Total Environ. 2018, 610, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Figueroa-Sanchez, M.A.; Nandini, S.; Sarma, S.S.S. Zooplankton community structure in the presence of low levels of cyanotoxins: A case study in a high altitude tropical reservoir (Valle de Bravo, Mexico). J. Limnol. 2014, 73, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Duan, Z.; Tan, X.; Parajuli, K.; Upadhyay, S.; Zhang, D.; Shu, X.; Liu, Q. Colony formation in two Microcystis morphotypes: Effects of temperature and nutrient availability. Harmful Algae 2018, 72, 14–24. [Google Scholar] [CrossRef]
- Rollwagen-bollens, G.; Lee, T.; Rose, V.; Bollens, S.M. Beyond Eutrophication: Vancouver Lake, WA, USA as a Model System for Assessing Multiple, Interacting Biotic and Abiotic Drivers of Harmful Cyanobacterial Blooms. Water 2018, 10, 757. [Google Scholar] [CrossRef] [Green Version]
- Corbel, S.; Mougin, C.; Martin-laurent, F.; Crouzet, O.; Bru, D.; Nélieu, S.; Bouaïcha, N. Evaluation of phytotoxicity and ecotoxicity potentials of a cyanobacterial extract containing microcystins under realistic environmental concentrations and in a soil-plant system. Chemosphere 2015, 128, 332–340. [Google Scholar] [CrossRef]
- Jankowiak, J.; Hattenrath-Lehmann, T.; Kramer, B.J.; Ladds, M.; Gobler, C.J. Deciphering the effects of nitrogen, phosphorus, and temperature on cyanobacterial bloom intensification, diversity, and toxicity in western Lake Erie. Limnol. Oceanogr. 2019, 64, 1347–1370. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.W.; Harke, M.J.; Marcoval, M.A.; Goleski, J.; Orano-dawson, C.; Berry, D.L.; Gobler, C.J. Effects of nitrogenous compounds and phosphorus on the growth of toxic and non-toxic strains of Microcystis during cyanobacterial blooms. Aquat Microb. Ecol. 2010, 61, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, S.R. Eutrophication of aquatic ecosystems: Bistability and soil phosphorus. Proc. Natl. Acad. Sci. USA 2005, 102, 10002–10005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Wu, X.; Hao, H.; He, Z. Mechanisms and assessment of water eutrophication. J. Zhejiang Univ. Sci. B 2008, 9, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Chaffin, J.D.; Bridgeman, T.B. Organic and inorganic nitrogen utilization by nitrogen-stressed cyanobacteria during bloom conditions. J. Appl. 2014, 26, 299–309. [Google Scholar] [CrossRef]
- Lu, J.; Zhu, B.; Struewing, I.; Xu, N.; Duan, S. Nitrogen-phosphorus- associated metabolic activities during the development of a cyanobacterial bloom revealed by metatranscriptomics. Sci. Rep. 2019, 9, 2480. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Li, L.; Gan, N.; Zheng, L.; Ma, H.; Shan, K.; Liu, J.; Xiao, B.; Song, L. Seasonal dynamics of water bloom-forming Microcystis morphospecies and the associated extracellular microcystin concentrations in large, shallow, eutrophic Dianchi Lake. J. Environ. Sci. 2014, 26, 1921–1929. [Google Scholar] [CrossRef]
- Lee, S.; Jiang, X.; Manubolu, M.; Riedl, K.; Ludsin, S.A.; Martin, J.F.; Lee, J. Fresh produce and their soils accumulate cyanotoxins from irrigation water: Implications for public health and food security. Food Res. Int. 2017, 102, 234–245. [Google Scholar] [CrossRef]
- Bukaveckas, P.A.; Franklin, R.; Tassone, S.; Trache, B.; Egerton, T. Cyanobacteria and cyanotoxins at the river-estuarine transition. Harmful Algae 2018, 76, 11–21. [Google Scholar] [CrossRef]
- Kibuye, F.A.; Zamyadi, A.; Wert, E.C. A critical review on operation and performance of source water control strategies for cyanobacterial blooms: Part I-chemical control methods. Harmful Algae 2021, 109, 102099. [Google Scholar] [CrossRef]
- Eke, J.; Wagh, P.; Escobar, I.C. Separation and Purification Technology Ozonation, biofiltration, and the role of membrane surface charge and hydrophobicity in removal and destruction of algal toxins at basic pH values. Sep. Purif. Technol. 2018, 194, 56–63. [Google Scholar] [CrossRef]
- Herrera, N.; Florez, M.T.; Velasquez, J.P.; Echeverri, F. Effect of phenyl-acyl compounds on the growth, morphology, and toxin production of Microcystisaeruginosa Kützing. Water 2019, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Zohdi, E.; Abbaspour, M. Harmful algal blooms (red tide): A review of causes, impacts, and approaches to monitoring and prediction. Int. J. Environ. Sci. Technol. 2019, 16, 1789–1806. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndlela, L.L.; Oberholster, P.J.; Van Wyk, J.H.; Cheng, P.H. A laboratory-based exposure of Microcystis and Oscillatoria cyanobacterial isolates to heterotrophic bacteria. Toxicon 2019, 165, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Águila-Ramírez, R.N.; Arenas-González, A.; Hernández-Guerrero, C.J.; González-Acosta, B.; Borges-Souza, J.M.; Véron, B.; Pope, J.; Hellio, C. Antimicrobial and antifouling activities achieved by extracts of seaweeds from Gulf of California, Mexico. Hidrobiologica 2012, 22, 8–15. [Google Scholar]
- Sedmak, B.; Carmeli, S.; Eleršek, T. “Non-Toxic” Cyclic Peptides Induce Lysis of Cyanobacteria—An Effective Cell Population Density Control Mechanism in Cyanobacterial Blooms. Microb. Ecol. 2019, 56, 201–209. [Google Scholar] [CrossRef]
- Van Wichelen, J.; Vanormelingen, P.; Codd, G.A.; Vyverman, W. The common bloom-forming cyanobacterium Microcystis is prone to a wide array of microbial antagonists. Harmful Algae 2016, 55, 97–111. [Google Scholar] [CrossRef]
- Su, X.; Steinman, A.D.; Tang, X.; Xue, Q.; Zhao, Y.; Xie, L. Response of bacterial communities to cyanobacterial harmful algal blooms in Lake Taihu, China. Harmful Algae 2017, 68, 168–177. [Google Scholar] [CrossRef]
- Kumar, P.; Hegde, K.; Brar, S.K.; Kermanshahi-pour, A.; Roy-lachapelle, A.; Galvez-cloutier, R. Co-culturing of Native Bacteria from Drinking Water Treatment Plant with Known—Biorefining and Remediation Laboratory, Department of Process Engineering and Applied. Chem. Eng. J. 2019, 383, 123090. [Google Scholar] [CrossRef]
- Zhang, S.J.; Du, X.P.; Zhu, J.M.; Meng, C.X.; Zhou, J.; Zuo, P. The complete genome sequence of the algicidal bacterium Bacillus subtilis strain JA and the use of quorum sensing to evaluate its antialgal ability. Biotechnol. Rep. 2020, 25, e00421. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zuo, J.; Song, L.; Gan, N. Microcystin-degrading bacteria affect mcyD expression and microcystin synthesis in Microcystis spp. J. Environ. Sci. 2015, 41, 195–201. [Google Scholar] [CrossRef]
- Trainic, M.; Koren, I.; Sharoni, S.; Frada, M.; Segev, L.; Rudich, Y.; Vardi, A. Infection Dynamics of a Bloom-Forming Alga and Its Virus Determine Airborne Coccolith Emission from Seawater. Iscience 2018, 6, 327–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, A. Cyanophages: Starving the Host to Recruit Resources. Curr. Biol. 2016, 26, R511–R513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawa, N.; Tatsuke, T.; Ogawa, A.; Hirokawa, Y.; Osanai, T.; Hanai, T. Modification of carbon metabolism in Synechococcus elongatus PCC 7942 by cyanophage-derived sigma factors for bioproduction improvement. J. Biosci. Bioeng. 2018, 127, 256–264. [Google Scholar] [CrossRef] [PubMed]
- El Amrani Zerrifi, S.; Mugani, R.; Redouane, E.M.; El Khalloufi, F.; Campos, A.; Vasconcelos, V.; Oudra, B. Harmful Cyanobacterial Blooms (HCBs): Innovative green bioremediation process based on anti-cyanobacteria bioactive natural products. Arch. Microbiol. 2021, 203, 31–44. [Google Scholar] [CrossRef]
- El Amrani Zerrifi, S.; Redouane, E.M.; Mugani, R.; Ribeiro, I.; de Fátima Carvalho, M.; Campos, A.; Barakate, M.; Vasconcelos, V.; Oudra, B.; El Khalloufi, F. Moroccan actinobacteria with promising activity against toxic cyanobacteria Microcystis aeruginosa. Environ. Sci. Pollut. Res. 2021, 28, 235–245. [Google Scholar] [CrossRef]
- Xie, P.; Liu, J. Practical Success of Biomanipulation using Filter-Feeding Fish to Control Cyanobacteria Blooms: A Synthesis of Decades of Research and Application in a Subtropical Hypereutrophic Lake. Sci. World J. 2001, 1, 337–356. [Google Scholar] [CrossRef] [Green Version]
- Aba, R.P.; Mugani, R.; Hejjaj, A.; Brugerolle de Fraissinette, N.; Oudra, B.; Ouazzani, N.; Campos, A.; Vasconcelos, V.; Carvalho, P.N.; Mandi, L. First Report on Cyanotoxin (MC-LR) Removal from Surface Water by Multi-Soil-Layering (MSL) Eco-Technology: Preliminary Results. Water 2021, 13, 1403. [Google Scholar] [CrossRef]
- WHO. Guidelines for Drinking-Water Quality; WORLD Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Von Gunten, U. Ozonation of drinking water: Part I. Oxidation kinetics and product formation. Water Res. 2003, 37, 1443–1467. [Google Scholar] [CrossRef]
- Chang, J.; Chen, Z.; Wang, Z.; Shen, J.; Chen, Q. Ozonation degradation of microcystin-LR in aqueous solution: Intermediates, byproducts, and pathways. Water Res. 2014, 63, 52–61. [Google Scholar] [CrossRef]
- Lu, S.Y.; Wang, N.Y.; Wang, C. Oxidation and biotoxicity assessment of microcystin-LR using different AOPs based on UV, O3, and H2O2. Front. Environ. Sci. Eng. 2018, 12. [Google Scholar] [CrossRef]
- Von Gunten, U. Ozonation of drinking water: Part II. Disinfection and by-product formation in presence of bromide, iodide, or chlorine. Water Res. 2003, 37, 1469–1487. [Google Scholar] [CrossRef]
- Sharma, V.K.; Triantis, T.M.; Antoniou, M.G.; He, X.; Pelaez, M.; Han, C.; Song, W.; O’Shea, K.E.; Armah, A.; Kaloudis, T.; et al. Destruction of microcystins by conventional and advanced oxidation processes: A review. Sep. Purif. Technol. 2012, 91, 3–17. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, G.; Chen, Y.; Jia, N.; Li, R. Four decades of progress in cylindrospermopsin research: The ins and outs of a potent cyanotoxin. J. Hazard. Mater. 2021, 406, 124653. [Google Scholar] [CrossRef] [PubMed]
- Pelaez, M.; Nolan, N.T.; Pillai, S.C.; Seery, M.K.; Falaras, P.; Kontos, A.G.; Dunlop, P.S.; Hamilton, J.W.; Byrne, J.; O’Shea, K.; et al. A review on the visible light active titanium dioxide photocatalysts for environmental applications. Appl. Catal. B Environ. 2012, 125, 331–349. [Google Scholar] [CrossRef] [Green Version]
- Merel, S.; Clément, M.; Thomas, O. State of the art on cyanotoxins in water and their behaviour towards chlorine. Toxicon 2010, 55, 677–691. [Google Scholar] [CrossRef]
- Staehelld, J.; Jurg, H.; Cited, L.; Staehelld, J.; Jurg, H. Decomposition of Ozone in Water in the Presence of Organic Solutes Acting as Promoters and Inhibitors of Radical Chain Reactions. Environ. Sci. Technol. 1985, 19, 1206–1213. [Google Scholar] [CrossRef]
- Ershov, B.G.; Morozov, P.A. The Kinetics of Ozone Decomposition in Water, the Influence of pH and Temperature. Chem. Kinet. Catal. 2009, 83, 1295–1299. [Google Scholar] [CrossRef]
- Yershov, B.G.; Morozov, P.A.; Gordeev, A.V.; Seliverstov, A.F. Kinetic Regularities of Ozone Decomposition in Water. Phys. Chem. Water Treat. Processes 2009, 31, 381–388. [Google Scholar] [CrossRef]
- Chang, J.; Chen, Z.; Wang, Z.; Kang, J.; Chen, Q.; Yuan, L.; Shen, J. Oxidation of microcystin-LR in water by ozone combined with UV radiation: The removal and degradation pathway. Chem. Eng. J. 2015, 276, 97–105. [Google Scholar] [CrossRef]
- Onstad, G.D.; Strauch, S.; Meriluoto, J.; Codd, G.A.; Von Gunten, U. Selective oxidation of key functional groups in cyanotoxins during drinking water ozonation. Environ. Sci. Technol. 2007, 41, 4397–4404. [Google Scholar] [CrossRef]
- Hitzfeld, B.C.; Höger, S.J.; Dietrich, D.R.; Hoger, S.J.; Dietrich, D.R. Cyanobacterial Toxins: Removal during Drinking Water Treatment, and Human Risk Assessment Cyanobacteria. Environ. Health Perspect. 2000, 108, 113–122. [Google Scholar] [PubMed] [Green Version]
- Xie, G.; Hu, X.; Du, Y.; Jin, Q.; Liu, Y.; Tang, C.; Hu, X.; Li, G.; Chen, Z.; Zhou, D.; et al. Light-driven breakdown of microcystin-LR in water: A critical review. Chem. Eng. J. 2021, 417, 129244. [Google Scholar] [CrossRef]
- Bai, M.; Zheng, Q.; Zheng, W.; Li, H.; Lin, S.; Huang, L. OH, Inactivation of Cyanobacterial Blooms and Degradation of Toxins in Drinking Water Treatment System. Water Res. 2019, 154, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Bai, M.; Yang, X.; Zhong, Z.; Gao, M.; Tian, Y. OH, pre-treatment of algae blooms and degradation of microcystin-LR in a drinking water system of 480 m3/day: Comparison with ClO2. Chem. Eng. J. 2019, 367, 189–197. [Google Scholar] [CrossRef]
- Papadimitriou, T.; Kormas, K.; Dionysiou, D.D. Using H2O2 treatments for the degradation of cyanobacteria and microcystins in a shallow hypertrophic reservoir. Environ. Sci. Pollut. Res. 2016, 23, 21523–21535. [Google Scholar] [CrossRef] [PubMed]
- He, X.; de la Cruz, A.A.; Hiskia, A.; Kaloudis, T.; O’Shea, K.; Dionysiou, D.D. Destruction of microcystins (cyanotoxins) by UV-254nm-based direct photolysis and advanced oxidation processes (AOPs): Influence of variable amino acids on the degradation kinetics and reaction mechanisms. Water Res. 2015, 74, 227–238. [Google Scholar] [CrossRef]
- Jasim, S.Y.; Saththasivam, J. Advanced oxidation processes to remove cyanotoxins in water. Desalination 2017, 406, 83–87. [Google Scholar] [CrossRef]
- Kumar, P.; Dayana, H.; Rubio, P.; Hegde, K.; Kaur, S.; Cledon, M.; Kermanshahi-pour, A.; Sauvé, S. Agro-industrial residues as unique support in a sand filter to enhance the bioactivity to remove microcystin-Leucine arginine and organics. Sci. Total Environ. 2019, 670, 971–981. [Google Scholar] [CrossRef]
- Miao, H.; Qin, F.; Tao, G.; Tao, W.; Ruan, W. Detoxification and degradation of microcystin-LR and -RR by ozonation. Chemosphere 2010, 79, 355–361. [Google Scholar] [CrossRef]
- Griffiths, D.J.; Saker, M.L. The Palm Island mystery disease 20 years on a review of research on the cyanotoxin cylindrospermopsin. Environ. Toxicol. 2003, 18, 78–93. [Google Scholar] [CrossRef]
- Serrà, A.; Philippe, L.; Perreault, F.; Garcia-Segura, S. Photocatalytic treatment of natural waters. Reality or hype ? The case of cyanotoxins remediation. Water Res. 2021, 188, 116543. [Google Scholar] [CrossRef] [PubMed]
- Azzam, E.I.; Jay-Gerin, J.P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Chen, A.; Peng, L.; Luo, S.; Zeng, Q.; Shao, J. Physiological characteristics and toxin production of Microcystis aeruginosa (Cyanobacterium) in response to DOM in anaerobic digestion effluent. Sci. Total Environ. 2019, 685, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Crittenden, J.C.; Trussell, R.R.; Hand, D.W.; Howe, K.J.; Tchobanoglous, G. Coagulation and Flocculation, MWH’s Water Treatment: Principles and Design, 3rd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 541–639. [Google Scholar]
- Gągała, I.; Mankiewicz-boczek, J. The Natural Degradation of Microcystins (Cyanobacterial Hepatotoxins) in Fresh Water—The Future of Modern Treatment Systems and Water Quality Improvement. Pol. J. Environ. Stud. 2012, 21, 1125–1139. [Google Scholar]
- Romero, L.G.; Mondardo, R.I.; Sens, M.L.; Grischek, T. Removal of cyanobacteria and cyanotoxins during lake bank filtration at Lagoa do Peri, Brazil. Clean Technol. Environ Policy 2014, 16, 1133–1143. [Google Scholar] [CrossRef]
- Şengül, A.B.; Ersan, G.; Tüfekçi, N. Removal of intra- and extracellular microcystin by submerged ultrafiltration (UF) membrane combined with coagulation/flocculation and powdered activated carbon (PAC) adsorption. J. Hazard. Mater. 2018, 343, 29–35. [Google Scholar] [CrossRef]
- Frišták, V.; Laughinghouse, H.D.; Bell, S.M. The Use of Biochar and Pyrolysed Materials to Improve Water Quality through Microcystin Sorption Separation. Water 2020, 12, 2871. [Google Scholar] [CrossRef]
- Chaffin, J.D.; Fox, E.L.; Nauman, C.A.; Slodysko, K.N. The ability of household pitcher-style water purifiers to remove microcystins depends on filtration rate and activated carbon source. Water Science and Technology: Water Supply 2019, 19, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Abbas, T.; Kajjumba, G.W.; Ejjada, M.; Masrura, S.U.; Marti, E.J.; Khan, E.; Jones-Lepp, T.L. Recent Advancements in the Removal of Cyanotoxins from Water Using Conventional and Modified Adsorbents—A Contemporary Review. Water 2020, 12, 2756. [Google Scholar] [CrossRef]
- Huang, W.; Cheng, B.; Cheng, Y. Adsorption of microcystin- LR by three types of activated carbon. J. Hazard. Mater. 2007, 141, 115–122. [Google Scholar] [CrossRef]
- Villars, K.; Huang, Y.; Lenhart, J.J. Drinking Water Using Granular Activated Carbon. Environ. Eng. Sci. 2020, 37, 585–595. [Google Scholar] [CrossRef]
- Pendleton, P.; Schumann, R.; Wong, S.H. Microcystin-LR adsorption by activated carbon. J. Colloid Interface Sci. 2001, 240, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Robert, L.; Burwell, J.R. Manualof symbols and terminology for physicochemical quantities and units—Appendix ii. Pure Appl. Chem. 1976, 46, 71–90. [Google Scholar]
- Zdravkov, B.D.; Čermák, J.J.; Šefara, M.; Janků, J.; Sefara, M.; Zdravkov, B.D.; Čermák, J.J.; Šefara, M.; Janků, J.; Sefara, M. Pore classification in the characterization of porous materials: A perspective. Cent. Eur. J. Chem. 2007, 5, 385–395. [Google Scholar] [CrossRef]
- Araújo, L.S.; Coutinho, A.D.; Alvarez-Mendez, M.O.; Moruzzi, R.B.; Calijuri, M.D.; Cunha, D.G. Caracterização e avaliação de fatores que determinam a remoção de microcistin-LR em carvão ativado granular produzido a partir de diferentes matérias-primas. Eng. Sanit. E Ambient. 2018, 23, 1131–1142. [Google Scholar] [CrossRef]
- Wu, X.; Xiao, B.; Li, R.; Wang, C.; Huang, J.; Wang, Z. Mechanisms and Factors Affecting Sorption of Microcystins onto Natural Sediments. Environ. Sci. Technol. 2011, 45, 2641–2647. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque Júnior, E.C.; Méndez, M.O.; Coutinho, A.D.; Franco, T.T. Removal of Cyanobacteria Toxins from Drinking Water by Adsorption on Activated Carbon Fibers. Mater. Res. 2008, 11, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Bajracharya, A.; Liu, Y.L.; Lenhart, J.J. The influence of natural organic matter on the adsorption of microcystin-LR by powdered activated carbon. Environ. Sci. Water Res. Technol. 2019, 5, 582. [Google Scholar] [CrossRef]
- Zhao, Y.X.; Gao, B.Y.; Shon, H.K.; Cao, B.C.; Kim, J.H. Coagulation characteristics of titanium (Ti) salt coagulant compared with aluminum (Al) and iron (Fe) salts. J. Hazard. Mater. 2011, 185, 1536–1542. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, Y.; Gao, B.; Zhao, Q. Enhanced algae removal by Ti-based coagulant: Comparison with conventional Al- and Fe-based coagulants. Environ. Sci. Pollut. Res. 2018, 25, 13147–13158. [Google Scholar] [CrossRef]
- Fuentes, M.; Olaetxea, M.; Baigorri, R.; Zamarreño, A.M.; Etienne, P.; Laîné, P.; Ourry, A.; Yvin, J.; Garcia-mina, J.M. Main binding sites involved in Fe (III) and Cu (II) complexation in humic-based structures. J. Geochem. Explor. J. 2013, 129, 14–17. [Google Scholar] [CrossRef]
- Klein, A.R.; Baldwin, D.S.; Silvester, E. Proton and iron-binding by the cyanobacterial toxin microcystin-LR. Environ. Sci. Technol. 2013, 47, 5178–5184. [Google Scholar] [CrossRef] [PubMed]
- Pochodylo, A.L.; Klein, A.R.; Aristilde, L. Metal-binding selectivity and coordination dynamics for cyanobacterial microcystins with Zn, Cu, Fe, Mg, and Ca. Environ. Chem. Lett. 2017, 15, 695–701. [Google Scholar] [CrossRef]
- Dai, G.; Gan, N.; Song, L.; Fang, S.; Peng, N. Fast adsorption of microcystin-LR by Fe (III)-modified powdered activated carbon. J. Oceanol. Limnol. 2018, 36, 1103–1111. [Google Scholar] [CrossRef]
- El Bouaidi, W.; Essalhi, S.; Douma, M.; Tazart, Z.; Ounas, A.; Enaime, G.; Yaacoubi, A. Evaluation of the potentiality of Vicia faba and Opuntia ficus indica as eco-friendly coagulants to mitigate Microcystis aeruginosa blooms. Desalination Water Treat. 2020, 195, 198–213. [Google Scholar] [CrossRef]
- Nakamura, T.; Adachi, Y.; Suzuki, M. Flotation and sedimentation of a single Microcystis floc collected from surface bloom. Water Res. 1993, 27, 979–983. [Google Scholar] [CrossRef]
- Zhu, W.; Li, M.; Luo, Y.; Dai, X.; Guo, L.; Xiao, M.; Huang, J.; Tan, X. Vertical distribution of Microcystis colony size in Lake Taihu: Its role in algal blooms. J. Great Lakes Res. 2014, 40, 949–955. [Google Scholar] [CrossRef] [Green Version]
- Piezer, K.; Li, L.; Jeon, Y.; Kadudula, A.; Seo, Y. The Application of Potassium Permanganate to Treat Cyanobacteria-Laden Water: A Review. Process Saf. Environ. Prot. 2021, 148, 400–414. [Google Scholar] [CrossRef]
- Xu, Z.; Woodhouse, J.N.; Te, S.H.; Yew-Hoong Gin, K.; He, Y.; Xu, C.; Chen, L.; Gin, K.Y.; He, Y. Seasonal variation in the bacterial community composition of a large estuarine reservoir and response to cyanobacterial proliferation. Chemosphere 2018, 202, 576–585. [Google Scholar] [CrossRef]
- Omori, K.; Sato, M.; Amano, Y.; Machida, M. Induction of colony formation of Microcystis aeruginosa by controlling extracellular polysaccharides and metal cation concentrations. J. Chem. Eng. Jpn. 2018, 51, 289–297. [Google Scholar] [CrossRef]
- Sakurai, S.; Omori, K.; Amano, Y.; Machida, M. Removal of Microcystis blooms using enhanced colony formation and buoyancy by controlling extracellular polysaccharides and cation concentrations. Int. J. Environ. Sci. Technol. 2019, 16, 4793–4802. [Google Scholar] [CrossRef]
- Hadjoudja, S.; Deluchat, V.; Baudu, M. Cell surface characterisation of Microcystis aeruginosa and Chlorella vulgaris. J. Colloid Interface Sci. 2010, 342, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Takaara, T.; Sano, D.; Masago, Y.; Omura, T. Surface-retained organic matter of Microcystis aeruginosa inhibiting coagulation with polyaluminum chloride in drinking water treatment. Water Res. 2010, 44, 3781–3786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amano, Y.; Hosoi, T.; Machida, M.; Imazeki, F. Effects of extracellular polymeric substances (EPS) and iron on colony formation of unicellular Microcystis aeruginosa. J. Jpn. Soc. Civ. Eng. 2013, 69, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Yu, G.; Jiang, H. Investigation on extracellular polymeric substances from mucilaginous cyanobacterial blooms in eutrophic freshwater lakes. Chemosphere 2013, 93, 75–81. [Google Scholar] [CrossRef]
- Pochodylo, A.L.; Aoki, T.G.; Aristilde, L. Adsorption mechanisms of microcystin variant conformations at water–mineral interfaces: A molecular modeling investigation. J. Colloid Interface Sci. 2016, 480, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Datta, T.; Amano, Y.; Omori, K. Influence of Extracellular Polysaccharides and Calcium Ion on Colony Formation of Unicellular Microcystis aeruginosa. Environ. Eng. Sci. 2017, 34, 149–157. [Google Scholar] [CrossRef]
- Sato, M.; Amano, Y.; Machida, M.; Imazeki, F. Colony formation of highly dispersed Microcystis aeruginosa by controlling extracellular polysaccharides and calcium ion concentrations in aquatic solution. Limnology 2017, 18, 111–119. [Google Scholar] [CrossRef]
- Bali, M.; Gueddari, M. Removal of phosphorus from secondary effluents using infiltration–percolation process. Appl. Water Sci. 2019, 9, 54. [Google Scholar] [CrossRef] [Green Version]
- Latrach, L.; Ouazzani, N.; Masunaga, T.; Hejjaj, A.; Bouhoum, K.; Mahi, M.; Mandi, L. Domestic wastewater disinfection by combined treatment using multi-soil-layering system and sand filters (MSL-SF): A laboratory pilot study. Ecol. Eng. 2016, 91, 294–301. [Google Scholar] [CrossRef]
- Corbel, S.; Bouaïcha, N.; Mougin, C. Dynamics of the toxic cyanobacterial microcystin-leucine-arginine peptide in agricultural soil. Environ. Chem. Lett. 2014, 12, 535–541. [Google Scholar] [CrossRef]
- Holst, T.; Jørgensen, N.O.; Jørgensen, C.; Johansen, A. Degradation of microcystin in sediments at oxic and anoxic, denitrifying conditions. Water Res. 2003, 37, 4748–4760. [Google Scholar] [CrossRef]
- Babica, P.; Kohoutek, J.; Bláha, L.; Adamovský, O.; Maršálek, B.; Per, O.L.P.A. Evaluation of extraction approaches linked to ELISA and HPLC for analyses of microcystin-LR, -RR, and -YR in freshwater sediments with different organic material contents. Anal. Bioanal. Chem. 2006, 385, 1545–1551. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Song, L.; Gan, N.; Li, L. Sorption, degradation, and mobility of microcystins in Chinese agriculture soils: Risk assessment for groundwater protection. Environ. Pollut. 2006, 144, 752–758. [Google Scholar] [CrossRef]
- Chen, W.; Li, L.; Gan, N.; Song, L. Optimization of an effective extraction procedure for the analysis of microcystins in soils and lake sediments. Environ. Pollut. 2006, 143, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, K.; Masui, H.; Uemura, H.; Mori, Y.; Harada, K. Analysis of microcystins in sediments using MMPB method. Toxicon 2021, 39, 687–692. [Google Scholar] [CrossRef]
- De Maagd, P.G.; Hendriks, A.J.; Seinen, W.; Sijm, D.T. pH-dependent hydrophobicity of the cyanobacteria toxin microcystin-LR. Water Res. 1999, 33, 677–680. [Google Scholar] [CrossRef]
- Miller, M.J.; Critchley, M.M.; Hutson, J.; Fallowfield, H.J. The adsorption of cyanobacterial hepatotoxins from water onto soil during batch experiments. Water Res. 2001, 35, 1461–1468. [Google Scholar] [CrossRef]
- Chénard, C.; Kolundžija, S.; Lauro, F.M.; Kolund, S.; Lauro, F.M. Complete genome sequence of the cyanophage S-PRM1 isolated from Singapore coastal waters. Mar. Genom. 2018, 43, 58–60. [Google Scholar] [CrossRef]
- Kang, S.; Xing, B. Adsorption of dicarboxylic acids by clay minerals as examined by in situ ATR-FTIR and ex situ DRIFT. Langmuir 2007, 23, 7024–7031. [Google Scholar] [CrossRef]
- Morris, R.J.; Williams, D.E.; Luu, H.A.; Holmes, C.F.; Andersen, R.J.; Calvert, S.E. The adsorption of microcystin-LR by natural clay particles. Toxicon 2000, 38, 303–308. [Google Scholar] [CrossRef]
- Liu, Y.L.; Walker, H.W.; Lenhart, J.J. Adsorption of microcystin-LR onto kaolinite, illite, and montmorillonite. Chemosphere 2019, 220, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Naidja, A.; Huang, P.M. Aspartic acid interaction with Ca-montmorillonite: Adsorption, desorption, and thermal stability. Appl. Clay Sci. 1994, 9, 265–281. [Google Scholar] [CrossRef]
- Thirumavalavan, M.; Hu, Y.; Lee, J. Effects of humic acid and suspended soils on adsorption and photo-degradation of microcystin-LR onto samples from Taiwan reservoirs and rivers. J. Hazard. Mater. 2012, 217, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Munusamy, T.; Hu, Y.; Lee, J. Adsorption and photodegradation of microcystin-LR onto sediments collected from reservoirs and rivers in Taiwan: A laboratory study to investigate the fate, transfer, and degradation of microcystin-LR. Environ. Sci. Pollut. Res. 2012, 19, 2390–2399. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Rachid, C.; Pachecoc, A.B.; Magalhães, V. Biotic and abiotic factors affect microcystin-LR concentrations in water/ sediment interface. Microbiol. Res. 2020, 236, 126452. [Google Scholar] [CrossRef] [PubMed]
- Corbel, S.; Mougin, C.; Bouaïcha, N. Cyanobacterial toxins: Modes of actions, fate in aquatic and soil ecosystems, phytotoxicity and bioaccumulation in agricultural crops. Chemosphere 2014, 96, 1–15. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Al Shehri, A.M. Microcystins in groundwater wells and their accumulation in vegetable plants irrigated with contaminated waters in Saudi Arabia. J. Hazard. Mater. 2009, 172, 310–315. [Google Scholar] [CrossRef]
- Cousins, I.T.; Bealing, D.J.; James, H.A.; Sutton, A.; Vol, W.R. Biodegradation of microcystin-LR by indigenous mixed bacterial populations. Water Res. 1996, 30, 481–485. [Google Scholar] [CrossRef]
- El Khalloufi, F.; Oufdou, K.; Bertrand, M.; Lahrouni, M.; Oudra, B.; Ortet, P.; Barakat, M.; Heulin, T.; Achouak, W. Microbiote shift in the Medicago sativa rhizosphere in response to cyanotoxins extract exposure. Sci. Total Environ. 2015, 539, 135–142. [Google Scholar] [CrossRef]
- Redouane, E.M.; El Amrani Zerrifi, S.; El Khalloufi, F.; Oufdou, K.; Oudra, B.; Lahrouni, M.; Campos, A.; Vasconcelos, V. Mode of action and faith of microcystins in the complex soil-plant ecosystems. Chemosphere 2019, 225, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Lemes GA, F.; Kist, L.W.; Bogo, M.R.; Yunes, J.S. Biodegradation of [D-Leu 1] microcystin-LR by a bacterium isolated from sediment of Patos Lagoon estuary, Brazil. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Song, L.; Peng, L.; Wan, N.; Zhang, X.; Gan, N. Reduction in microcystin concentrations in large and shallow lakes: Water and sediment-interface contributions. Water Res. 2007, 42, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Terin, U.C.; Sabogal-Paz, L.P. Microcystis aeruginosa and microcystin-LR removal by household slow sand filters operating in continuous and intermittent flows. Water Res. 2019, 150, 29–39. [Google Scholar] [CrossRef]
- Li, H.; Xing, P.; Chen, M.; Bian, Y.Y.; Wu, Q.L. Short-term bacterial community composition dynamics in response to accumulation and breakdown of Microcystis blooms. Water Res. 2011, 45, 1702–1710. [Google Scholar] [CrossRef]
- Lezcano, M.Á.; Velázquez, D.; Quesada, A.; El-shehawy, R. Diversity and temporal shifts of the bacterial community associated with a toxic cyanobacterial bloom: An interplay between microcystin producers and degraders. Water Res. 2017, 125, 52–61. [Google Scholar] [CrossRef]
- Kumar, P.; Hegde, K.; Kaur, S.; Cledon, M.; Brar, S.K.; Cledon, M.; Kermanshahi-pour, A. Potential of biological approaches for cyanotoxin removal from drinking water: A review. Ecotoxicol. Environ. Saf. 2019, 172, 488–503. [Google Scholar] [CrossRef]
- Kormas, K.A.; Lymperopoulou, D.S. Cyanobacterial Toxin Degrading Bacteria: Who Are They? BioMed Res. Int. 2013, 2013, 463894. [Google Scholar] [CrossRef] [Green Version]
- Bourne, D.G.; Blakeley, R.L.; Riddles, P.; Jones, G.J. Biodegradation of the cyanobacterial toxin microcystin LR in natural water and biologically active slow sand filters. Water Res. 2006, 40, 1294–1302. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Shi, G.; Mei, Z.; Wang, R.; Li, D. Discerning biodegradation and adsorption of microcystin-LR in a shallow semi-enclosed bay and bacterial community shifts in response to associated process. Ecotoxicol. Environ. Saf. 2016, 132, 123–131. [Google Scholar] [CrossRef]
- Tsao, S.; Wei, D.J.; Chang, Y.T.; Lee, J.F. Aerobic biodegradation of microcystin-LR by an indigenous bacterial mixed culture isolated in Taiwan. Int. Biodeterior. Biodegrad. 2017, 124, 101–108. [Google Scholar] [CrossRef]
- Wang, R.; Tai, Y.; Wan, X.; Ruan, W.; Man, Y.; Wang, J.; Yang, Y.; Yang, Y. Enhanced removal of Microcystis bloom and microcystin-LR using microcosm constructed wetlands with bioaugmentation of degrading bacteria. Chemosphere 2018, 210, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Eleuterio, L.; Batista, J.R. Biodegradation studies and sequencing of microcystin-LR degrading bacteria isolated from a drinking water biofilter and a fresh water lake. Toxicon 2010, 55, 1434–1442. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.R.; Wilhelm, S.W.; Boyer, G.L. The fate of microcystins in the environment and challenges for monitoring. Toxins 2014, 6, 3354–3387. [Google Scholar] [CrossRef] [Green Version]
- Gurbuz, F.; Codd, G.A. Microcystin Removal by a Naturally-Occurring Substance: Pumice. Bull. Environ. Contam. Toxicol. 2008, 81, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yang, X.; Yang, L.; Xiao, B.; Wu, X.; Wang, J.; Wan, H. An effective pathway for the removal of microcystin LR via anoxic biodegradation in lake sediments. Water Res. 2010, 44, 1884–1892. [Google Scholar] [CrossRef]
- Kumar, P.; Hegde, K.; Brar, S.K.; Cledon, M.; Kermanshahi-pour, A.; Roy-Lachapelle, A.; Galvez-Cloutier, R. Novel fluidized-bed biofilm reactor for concomitant removal of microcystin-LR and organics. Chem. Eng. J. 2019, 359, 99–111. [Google Scholar] [CrossRef]
- Jeon, Y.; Li, L.; Calvillo, J.; Ryu, H.; Santo Domingo, J.W.; Choi, O.; Brown, J.; Seo, Y. Impact of algal organic matter on the performance, cyanotoxin removal, and biofilms of biologically-active filtration systems. Water Res. 2020, 184, 116120. [Google Scholar] [CrossRef]
- Westrick, J.A.; Szlag, D.C.; Southwell, B.J.; Sinclair, J.A.; Szlag, D.C.; Southwell, B.J.; Sinclair, J. A Review of Cyanobacteria and Cyanotoxins Removal/Inactivation in Drinking Water Treatment. In Analytical and Bioanalytical Chemistry; Springer Science and Business: Berlin/Heidelberg, Germany, 2010; Volume 397, pp. 1705–1714. [Google Scholar] [CrossRef]
- Bavithra, G.; Azevedo, J.; Oliveira, F.; Morais, J.; Pinto, E.; Ferreira, I.M.; Vasconcelos, V.; Campos, A.; Almeida, C.M. Assessment of Constructed Wetlands’ Potential for the Removal of Cyanobacteria and Microcystins (MC-LR). Water 2020, 12, 10. [Google Scholar] [CrossRef] [Green Version]
- Kochi, L.Y.; Freitas, P.L.; Maranho, L.T.; Juneau, P.; Gomes, M.P. Aquatic macrophytes in constructed wetlands: A fight against water pollution. Sustainability 2020, 12, 9202. [Google Scholar] [CrossRef]
- Cheng, R.; Zhu, H.; Shutes, B.; Yan, B. Treatment of microcystin (MC-LR) and nutrients in eutrophic water by constructed wetlands: Performance and microbial community. Chemosphere 2021, 263, 128139. [Google Scholar] [CrossRef] [PubMed]
- Morón-López, J.; Nieto-Reyes, L.; Molina, S.; Lezcano, M.Á. Exploring microcystin-degrading bacteria thriving on recycled membranes during a cyanobacterial bloom. Sci. Total Environ. 2020, 736, 139672. [Google Scholar] [CrossRef]
- Morón-López, J.; Molina, S. Optimization of Recycled-Membrane Biofilm Reactor (R-MBfR) as a sustainable biological treatment for microcystins removal. Biochem. Eng. J. 2020, 153, 107422. [Google Scholar] [CrossRef]
- Wei, C.; Wu, W. Performance of single-pass and by-pass multi-step multi-soil-layering systems for low- (C/N) -ratio polluted river water treatment. Chemosphere 2018, 206, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Huang, G.; An, C.; Shen, J.J.; Zhang, P.; Chen, X.; Shen, J.J.; Yao, Y. Treatment of rural domestic wastewater using multi-soil-layering systems: Performance evaluation, factorial analysis, and numerical modeling. Sci. Total Environ. 2018, 644, 536–546. [Google Scholar] [CrossRef]
- Lamzouri, K.; Mahi, M.; Ouatar, S.; Bartali, E.; Masunaga, T.; Latrach, L.; Mandi, L. Application of Multi-soil-layering technique for wastewater treatment in Moroccan rural areas: Study of the operation process for an engineering design. J. Mater. Environ. Sci. 2017, 7, 579–585. [Google Scholar]
- Latrach, L.; Ouazzani, N.; Hejjaj, A.; Zouhir, F.; Mahi, M.; Masunaga, T.; Mandi, L. Optimization of hydraulic efficiency and wastewater treatment performances using a new design of vertical flow Multi-Soil-Layering (MSL) technology. Ecol. Eng. 2018, 117, 140–152. [Google Scholar] [CrossRef]
- Latrach, L.; Ouazzani, N.; Hejjaj, A.; Mahi, M.; Masunaga, T.; Mandi, L. Two-stage vertical flow multi-soil-layering (MSL) technology for efficient removal of coliforms and human pathogens from domestic wastewater in rural areas under arid climate. Int. J. Hyg. Environ. Health 2018, 221, 64–80. [Google Scholar] [CrossRef]
- Shen, J.; Huang, G.; An, C.; Song, P.; Xin, X.; Yao, Y.; Zheng, R. Biophysiological and factorial analyses in the treatment of rural domestic wastewater using multi-soil-layering systems. J. Environ. Manag. 2018, 226, 83–94. [Google Scholar] [CrossRef]
- Attanandana, T.; Saitthiti, B.; Thongpae, S.; Kritapirom, S. Multi-media-layering system for food service wastewater treatment. Ecol. Eng. 2000, 15, 133–138. [Google Scholar] [CrossRef]
- Hong, Y.; Huang, G.; An, C.; Song, P.; Xin, X.; Chen, X. Enhanced nitrogen removal in the treatment of rural domestic sewage using vertical- flow multi-soil-layering systems: Experimental and modeling insights. J. Environ. Manag. 2019, 240, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Latrach, L.; Masunaga, T.; Ouazzani, N.; Hejjaj, A.; Mandi, L. Removal of bacterial indicators and pathogens from domestic wastewater by the multi-soil-layering (MSL) system. Soil Sci. Plant Nutr. 2015, 61, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Morató, J.; Codony, F.; Sánchez, O.; Pérez, L.M.; García, J.; Mas, J. Key design factors affecting microbial community composition and pathogenic organism removal in horizontal subsurface flow constructed wetlands. Sci. Total Environ. 2014, 481, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Lezcano, M.Á.; Quesada, A.; El-Shehawy, R. Seasonal dynamics of microcystin-degrading bacteria and toxic cyanobacterial blooms: Interaction and influence of abiotic factors. Harmful Algae 2018, 71, 19–28. [Google Scholar] [CrossRef]



{kind=link}
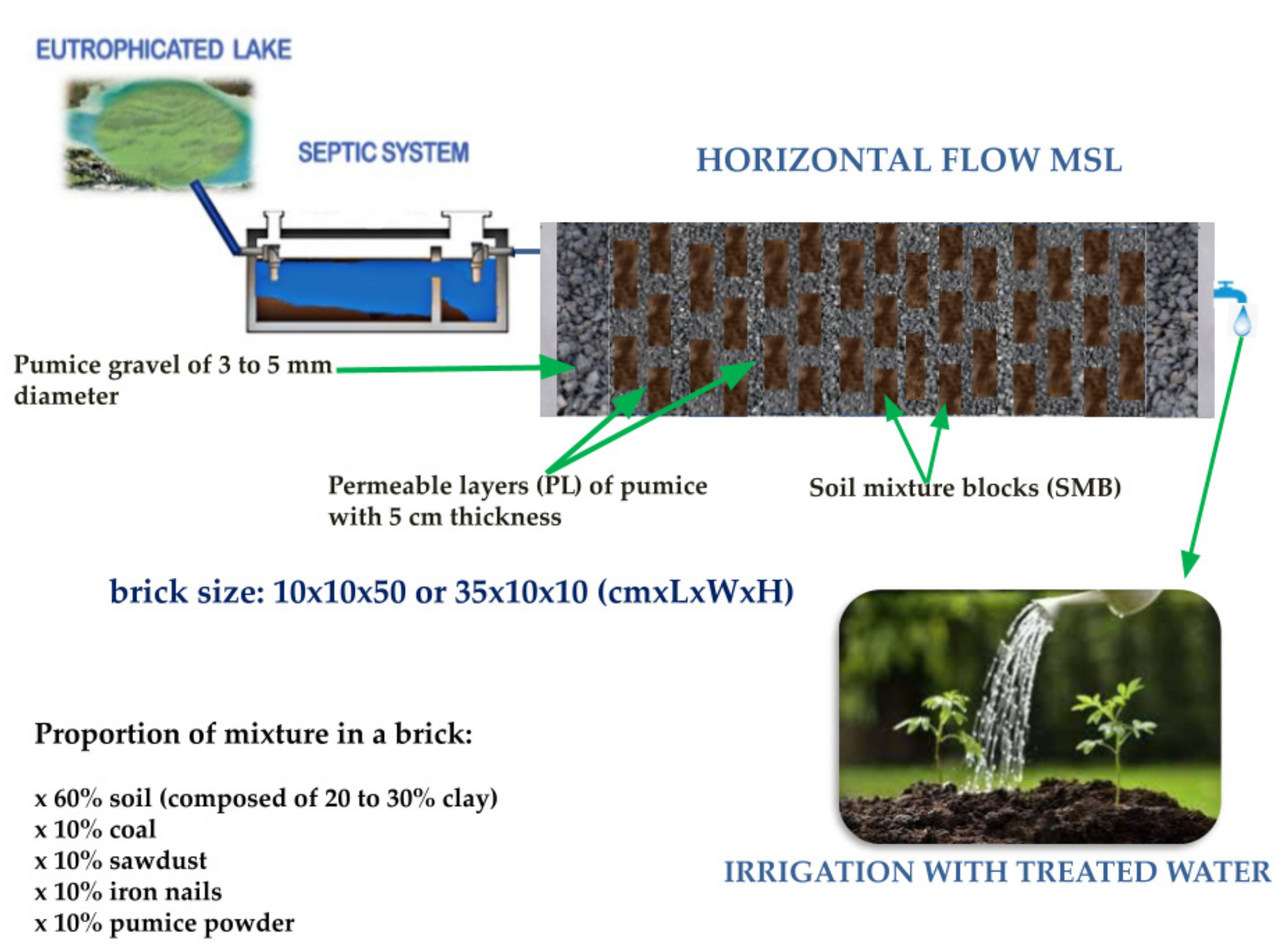{kind=link}
| Species | Produced Toxin | Toxin Family | Highest Amount of Toxin Quantified | Range of LD50 | References |
|---|---|---|---|---|---|
| Microcystis aeruginosa | Microcystin | Hepatotoxin | 11,500 μg MC-LR-eq g−1 DW | 50–1200 μgkg−1 mouse body weight | [10,11] |
| Microcystis botrys | Microcystins | Hepatotoxin | 90% of microcystins in analyzed colonies | - | [12] |
| Microcystis flos-aquae | Microcystins | Hepatotoxin | 50% of microcystins in analyzed colonies | - | [12] |
| Microcystis panniformis | Microcystins | Hepatotoxin | 53% of microcystins in analyzed colonies | - | [12] |
| Microcystis wesenbergii | Microcystins | Hepatotoxin | 0% of microcystins in analyzed colonies | - | [12] |
| Microcystis ichthyoblabe | Microcystins | Hepatotoxin | 20% of microcystins in analyzed colonies | - | [12] |
| Microcystis viridis | Microcystin | Hepatotoxin | 17% of microcystins in analyzed colonies | >1200 μgkg−1 | [12] |
| Planktothrix rubescens | Microcystin | Hepatotoxin | 1500 μg MC g−1 DW | [13] | |
| Planktothrix agardhii | Microcystin | Hepatotoxin | 4500 μg MC g−1 DW | - | [14] |
| Woronichinia naegeliana | Microcystin | Hepatotoxin | - | - | [14] |
| Anabaena spiroides | Microcystins | Hepatotoxin | - | - | [15] |
| Nostoc muscorum | Microcystin | Hepatotoxin | 229.4 μg MC g−1 DW | 15–125 mgkg−1 mouse body weight | [16] |
| Dolichospermum flosaquae | Microcystins | Hepatotoxin | - | 160–300 μgkg−1 mouse body weight | [14,17] |
| Nodularia spumigena | Nodularin | Hepatotoxin | 43.6 μg NOD/kg DW sea mullet livers | - | [18] |
| Chrooccocus minutus | Microcystins | Hepatotoxin | 132 MC μg L−1 | - | [19] |
| Oscillatoria limnetica | Microcystins | Hepatotoxin | 877 μg MC-LR-eq μg g−1 DW | - | [20] |
| - | [20] | ||||
| Aphanizomenon ovalisporum | cylindrospermopsin | Hepatotoxin | 8700 µg CYN g−1 DW | - | [21] |
| Cylindrospermopsis raciborskii | cylindrospermopsin | Hepatotoxin | 70.83 µg CYN g−1 DW | [22] | |
| Microcystis sp. | Anatoxin-a | Neurotoxin | 0.12μg ANTX-a g−1 DW | 31 μgkg−1 mouse body weight | [17,23] |
| Aphanizomenon flos-aquae | Anatoxin-a | Neurotoxin | 24.62 μg ANTX-a g−1 DW | - | [23] |
| Anabaena sp. | Anatoxin-a | Neurotoxin | 21.9 μg ANTX-a g−1 DW | - | [23] |
| Cylindrospermopsis raciborskii | saxitoxin | Neurotoxin | 0.20 μgL−1 STXs | 10 μgkg−1 mouse body weight | [17,24] |
| Lyngbya sp. | Debromoaplysiatoxin | Dermatotoxins | 6.31 μg DAT g−1 DW | - | [25] |
| Eco-Technology Name | Processing Basis: System Strength | Highest Initial MC Concentration Used (µg/L) | Highest MC Removal Rate Obtained (%) | Lowest MC Concentration Reached (µg/L) | Presence of Cyanobacterial Cells (cells/ml) * or Chl a (µg/L) ** | Highest Cyanobacterial Cell Abatement (log10 unit) a or (% Chl a Removal) b | References |
|---|---|---|---|---|---|---|---|
| Microbial bioaugmented constructed wetlands | Constructed wetlands material and biological activity | 16.07 | 90 | na | 179.3 | 90 | [157] |
| A household slow sand filter (C-HSSF) | HSSF material and biological activity | 5 | na | <1 | 1 × 105 * | 2.39 ± 0.34 a | [149] |
| A household slow sand filter (I-HSSF) | HSSF material and biological activity | 5 | na | <1 | 1 × 105 * | 2.01 ± 0.43 a | [149] |
| Constructed Wetlands | Constructed wetlands material and biological activity | 50 | 99.9 | na | 1 × 106 * | 94 | [165] |
| Multi-soil-layering system (MSL) | MSL material and biological activity | 10 | 99.35 | na | na | na | [60] |
| Repurposed Osmotic membrane | Discarded Osmotic membrane and bioaugmented MC degrading bacterial strain | 836 | 90 | <0.2 | na | na | [168] |
| Constructed wetlands | Constructed wetlands material and biological activity | 14.41 | 80 | <0.5 | na | na | [167] |
| Parameters | Recorded Removal Efficiency (%) | References Respectively to Recorded Efficiency % |
|---|---|---|
| SS | 93; 98.21; 95; 71 | [172]; [175]; [173]; [176] |
| BOD5 | 85; 93.66; 92.34; 90 | [172]; [175]; [173]; [176] |
| COD | 98.29; 70; 81; 98.53; 86.11; 81; 70 | [177]; [170]; [172]; [171]; [175]; [173]; [176] |
| NH4+-N | 76.60; 99; 88; 100; 92.81; 89; 76 | [177]; [170]; [172]; [171]; [175]; [173]; [176] |
| NTK | 82; 94 | [172]; [173] |
| TN | 69.86; 82; 83; 64; 83.67; 92; 91 | [170]; [172]; [171]; [175]; [173]; [177] |
| NO3−-N | 96.15; 96 | [177]; [172]; [173] |
| PO43−-P | 82; 98; 89 | [172]; [173]; [177] |
| TP | 100; 82; 84; 100; 86.20; 98.90 | [177]; [170]; [172]; [171]; [175]; [173] |
| Variables | Inactivation Ratio in Log10 Units | References |
|---|---|---|
| Total coliforms | 1.28; 2.2; 4.46; 2.3; 2.36 | [178]; [179]; [125]; [174]; [173] |
| Fecal coliforms | 1.21; 2.2; 4.47; 2.18; 2.38 | [178]; [179]; [125]; [173] |
| Fecal Streptococci | 1.16; 2.3; 4.13; 3.15; 2.21 | [178]; [179]; [125]; [173] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mugani, R.; Aba, R.P.; Hejjaj, A.; El Khalloufi, F.; Ouazzani, N.; Almeida, C.M.R.; Carvalho, P.N.; Vasconcelos, V.; Campos, A.; Mandi, L.; et al. Multi-Soil-Layering Technology: A New Approach to Remove Microcystis aeruginosa and Microcystins from Water. Water 2022, 14, 686. https://doi.org/10.3390/w14050686
Mugani R, Aba RP, Hejjaj A, El Khalloufi F, Ouazzani N, Almeida CMR, Carvalho PN, Vasconcelos V, Campos A, Mandi L, et al. Multi-Soil-Layering Technology: A New Approach to Remove Microcystis aeruginosa and Microcystins from Water. Water. 2022; 14(5):686. https://doi.org/10.3390/w14050686
Chicago/Turabian StyleMugani, Richard, Roseline Prisca Aba, Abdessamad Hejjaj, Fatima El Khalloufi, Naaila Ouazzani, C. Marisa R. Almeida, Pedro N. Carvalho, Vitor Vasconcelos, Alexandre Campos, Laila Mandi, and et al. 2022. "Multi-Soil-Layering Technology: A New Approach to Remove Microcystis aeruginosa and Microcystins from Water" Water 14, no. 5: 686. https://doi.org/10.3390/w14050686