
Review of the Distribution and Influence of Antibiotic Resistance Genes in Ballast Water
School of Environment and Architecture, University of Shanghai for Science and Technology, Shanghai 200093, China
*
Authors to whom correspondence should be addressed.
Water 2022, 14(21), 3501; https://doi.org/10.3390/w14213501
Submission received: 3 September 2022
/
Revised: 26 October 2022
/
Accepted: 27 October 2022
/
Published: 2 November 2022
(This article belongs to the Special Issue Soluble Microbial Products and Perfluorinated Compounds in Wastewater Treatment)
Abstract
:The misuse of antibiotics causes antibiotic resistance genes (ARGs) in bacteria to be gradually enriched by environmental selection, resulting in increased tolerance and resistance in bacteria to antibiotics. Ballast water is a mobile carrier for the global transfer of bacteria and genes, thus posing a certain risk of ARGs spreading into the global ocean. Therefore, it is important to investigate the current status of ARGs in ballast water, as well as control the abundance of ARGs. Herein, we attempt to comprehensively summarize the distribution and abundance of ARGs in ballast water from different sea areas and analyze the influencing factors (such as physical factors, chemical factors, temperature, pH, etc.) on the distribution of ARGs. Furthermore, we seek to review the changes in ARGs after differential disinfection technology treatment in ballast water (including chlorination, ultraviolet, ozone, and free radical technology), especially the enhancing effect of subinhibitory concentrations of disinfectants on ARGs transfer. Overall, we believe this review can serve as a guide for future researchers to establish a more reasonable standard of ballast water discharge that considers the pollution of ARGs and provide new insight into the risk of vertical and horizontal ARG transfer in ballast water after disinfection.
1. Introduction
Ballast water is the water that fills the bottom and side tanks of a ship to keep the hull balanced and accounts for approximately 40% of the cargo transportation volume [1]. According to the estimates of the International Maritime Organization (IMO), each year, up to 12 billion tons of ballast water are transshipped around the world, and every year more than 80,000 ships transport ballast water around the world [2]. The transfer and discharge of ballast water from ships is currently the main pathway leading to the invasion of alien organisms through offshore waters (about 29% of total biological invasions) [3], and it is estimated that approximately 3000 species are transferred through ship ballast water every day [4]. A large number of potentially pathogenic microorganisms have been detected in river estuaries, ports, and bays, posing potential harm to aquatic organisms and human health [5]. Therefore, ballast water is a mobile carrier for the global transfer of bacteria and genes, which has been attracting much attention in recent years [6].
Recently, various residual antibiotics found in the marine environment have become a focus of public concern [7,8,9]. The high concentrations of antibiotics in the marine environment are mainly derived from chemical production, breeding, aquaculture, and medical care [10]. Additionally, some kinds of antibiotics are resistant to degradation, resulting in their environmental accumulation in marine environments. Antibiotics at high concentrations can threaten the biodiversity and function of adjacent aquatic ecosystems and may cause bacteria to produce antibiotic resistance genes (ARGs) and threaten human health [11]. If antibiotics are allowed to remain in the marine environment, they will exert selective pressure on bacteria, inducing the enrichment of ARGs. ARGs can be horizontally transferred between different strains through mobile genetic elements (MGEs) [12,13], thereby enabling more microorganisms to acquire antibiotic resistance. Gene transfer from ARGs can allow antibiotic-resistant pathogenic bacteria to emerge. It also endangers people’s health and disturbs the ecosystem’s balance [14].
The aquatic ecosystem is a huge natural repository of ARGs. Recently, with frequent human activities, the increase in wastewater discharge, and the development of marine aquaculture activities, the abundance of ARGs in the marine environment has gradually increased [10,15]. Therefore, antibiotics from agriculture and human waste are often found in ballast water near the coast [16,17]. Because medical treatment and animal husbandry use a lot of antibiotics, medical wastewater and aquaculture wastewater are considered to be the main source of antibiotic-resistant genes [18,19]. Wastewater and domestic sewage containing antibiotics or resistance genes will be directly or indirectly discharged into rivers [6], lakes [20], and oceans. Therefore, the sea will become the final place for the influx of resistance genes [21]. There are three possible sources of ARGs in coastal waters. The first source is the direct input from land sources, and surface runoff carrying ARGs enters the coastal waters [22]. For example, effluent from sewage treatment plants [23,24,25]. The second source is the ARGs produced by the selective pressure of antibiotics or other poisons used in water aquaculture [26]. The third source is the inherent background value of those antibiotic resistance genes evolved by microorganisms in the marine environment to resist other populations that can secrete antibiotics [27].
Ballast operations are usually carried out in offshore waters [28]. Therefore, seawater containing various bacteria [29,30,31,32], ARGs [33,34], and antibiotic residues [6] can be pumped into the ship’s ballast tank. When antibiotic-resistant bacteria in ballast water are gradually enriched, the transfer of antibiotic-resistant genes between microbial communities occurs [28]. It has been proven that there are more ARGs in ship ballast water than in samples taken from the nearby ocean [5,35,36]. In addition, ARGs were found in many samples of ballast water from different sea areas, which shows that ballast water can spread antibiotic resistance [21]. Therefore, it is necessary to control the abundance of ARGs in ballast water.
When minimizing the hazards from ballast water discharge, ballast water must be discharged after treatment. At present, relevant studies have shown that adsorption, activated-sludge technology [37,38,39], and constructed wetlands [40] have been used to deal with ARGs pollution. These methods may cause secondary pollution problems. In these treatment methods, ARGs are mainly transferred rather than completely degraded. This paper summarized the common disinfection methods of antibiotic resistance genes and summarized their advantages and disadvantages. The common and traditional disinfection processes mainly include ultraviolet disinfection [41] and chlorination disinfection [42]. Free radical disinfection is currently a research hotspot, including the Fenton reaction [43], photocatalysis [44,45], and photoelectric catalysis [46], which show great advantages in removing ARGs.
In summary, the discharge of ballast water accelerates the spread and diffusion of ARGs in the marine environment, with each pollution source and carrier diffusing exponentially [47]. It is necessary to summarize the distribution and abundance of ARGs in ballast water. In addition, there are few reviews on the abundance changes of ARGs and the transfer risk of ARGs in ballast water after disinfection. Therefore, the main contents of this review mainly include (1) introducing the distribution and abundance of ARGs in ballast water from different sea areas and different sources; (2) investigating the abundance of resistance genes in ballast water after disinfection, as well as the vertical and horizontal transfer of ARGs under nonantibiotic stress mediated by different disinfection methods.
2. Methodology
2.1. Methods
In order to understand the distribution and impact of antibiotic resistance genes in ballast water, we reviewed the most relevant studies on the use of antibiotics, the generation of antibiotic-resistant bacteria in water, the biological invasion caused by transmission and ballast water transportation, and the transmission of antibiotic resistance genes. This paper combs the research on the distribution and transfer of antibiotic resistance genes in the past 25 years from several aspects, such as the issuing agency, the author’s cooperation network, keyword and research evolution, digging the frontier hot spots in this field, looking forward to identify the future research trends in this field, and provide a certain reference for the related research and time development to solve the harm of antibiotic resistance genes.
2.2. Identifification of Records
The scope of this paper is to review the distribution and impact of antibiotic resistance genes in ballast water, especially the papers that collect the distribution of antibiotic resistance genes in ballast water transportation, the impact of antibiotic resistance gene transfer on the environment, and the disinfection methods of antibiotic resistance genes. To ensure the quality of the comments, we collected peer-reviewed journal articles and research reports indexed by Science Direct and Web of Science databases. In collecting these references, we used any search term in the title, abstract, and keywords to identify the relevant research. Among them, the keywords “ballast water”, “antibiotic resistance gene”, “antibiotic-resistant bacteria”, “resistance gene transfer”, “resistance gene removal”, and “ballast water disinfection” were used to determine the reference library. The time range of the references in this review span 1998 to 2022.
2.3. Eligibility Criteria
The purpose of this review is to include searchable studies on the distribution of antibiotic resistance genes in ballast water and the disinfection of resistance genes during transportation. The main research that met the conditions for this review required the following criteria: (1) the distribution of resistance genes in ballast water was analyzed; (2) the effect of the disinfection technology on the inactivation of the resistant genes was analyzed; (3) the effects of the minimum inhibitory concentration on the horizontal transfer of resistant genes were analyzed.
3. Bacteria in Ship Ballast Water
At the Diplomatic Conference of the International Maritime Organization in 2004, the “International Convention on the Control and Management of Ships Ballast Water and Its Sediments” was passed. This is also called the “Ballast Water Convention”. The ballast water convention mainly has five chapters (chapters A–E). All ships subject to the ballast water convention have fully entered the stage of implementing ballast water management, which is the second regulation in Chapter D, namely, the D-2 standard [48] (Table 1). At present, people’s attention to ship ballast water discharge is focused on this standard.
Even though disinfection may remove the majority of the bacteria in ballast water, there are still many bacteria in the water that can spread rapidly to new maritime environments. According to reports, pathogenic germs and other types of bacteria have been discovered in ballast water, mostly in Chinese ports [35]. In addition, bacteria may also affect the sea area of the port with the ship’s ballast water after long-distance migration [49]. For example, Lv et al. [50] used 16 S rRNA technology to analyze sediment samples from the ballast tanks of nine ocean-going ships in Jiangyin, China, and detected 44 pathogenic bacteria from the sediments, among which Pseudomonas alkalogenes, Enterococcus Heidegger, Shigella, and Bacillus anthracis were the most abundant, accounting for 64.5% of the total pathogenic bacterial sequences. Yang et al. [51] found nine kinds of pathogenic bacteria in the ballast water of the Yangshan Port, including Pseudoalteromonas piscicida, Rhodococcus erythropolis, Vibrio sp., Bacillus aquimaris, Vibrio alginolyticus, etc. Altug et al. [6] investigated the ballast water from ships sailing to Turkey in the Marmara Sea and found 38 species, including 27 pathogens, which shows that ballast water could be harmful. Salleh et al. [52] investigated the ballast water from eight different sources, and 33 potential pathogens were detected from all of the ballast water samples. Among them, Pseudomonas spp., Tenacibaculum spp., Flavobacterium spp., Halomonas spp., and Acinetobacter junii are the main potential pathogens.
Furthermore, Ng et al. [53] analyzed the ballast water of six cabins in a Singapore port and found a variety of vibrios that showed antibiotic resistance. Rivera et al. [54] studied ballast water in Brazil and found the Vibrio cholerae O1-producing virus in the ballast water samples. Invasive Vibrio cholerae strains can be further transported to other regions by ships, which will bring harm to other regions. Dobbs et al. [55] also proved that Vibrio cholerae in ship ballast water gradually acquired resistance to antibiotics, which indicated that Vibrio cholerae isolates could transfer ARGs to other Vibrio cholerae strains by horizontal transfer. This phenomenon may be enhanced in the ballast tank [56]. Therefore, ballast water is not only a carrier of bacteria for global spreading, but it is also a source of genes.
The spread of bacteria, especially pathogenic bacteria carried by ballast water, not only threatens the local marine environment and aquatic fisheries but also threatens human life [55]. On the other hand, the transfer of drug-resistance genes to pathogenic bacteria will gradually enhance the threat of pathogenic bacteria [47]. In recent years, the existing microbial detection basis for ballast water have tended to be standardized; however, ARGs are an emerging pollutant that have been neglected in the detection of ballast water. Therefore, the discharge index of ballast water needs to be changed, and the management of ballast water discharge needs to be investigated [57].
4. Resistance Genes
4.1. Antibiotics and Antibiotic Resistance Genes
Antibiotics, which are important growth inhibitors to prevent bacterial diseases and stimulate growth in aquaculture, are often used as disease-prevention drugs and feed additives in aquaculture [10]. At present, the commonly used antibiotics with a high detection frequency and detection abundance in the environment mainly include the following categories (Table 2): tetracyclines, sulfonamides, quinolones, β-lactams, aminoglycosides, macrolides class, and amide alcohols (also known as chloramphenicol antibiotics) [9]. However, because of the abuse of antibiotics, bacteria suffer from sustained antibiotic stress, which induces the enrichment of ARGs in bacteria. Recently, antibiotic resistance has become a hot issue of global concern [12]. Antibiotic-resistant bacteria always show multiple antibiotic resistance (MARs), leading to the emergence of multiantibiotic-resistant bacteria and even “superbugs” that can tolerate most antibiotics [48]. Furthermore, ARGs will accumulate, diffuse, and be transferred into the environment.
4.2. ARGs in the Marine Environment
Recently, ARGs in the marine environment have mainly come from sewage treatment plants, aquaculture farms, and medical wastewater, and some also come from antibiotic production plants and animal husbandry [12,13,57]. ARGs, such as sulfonamides, tetracyclines, chloramphenicols, macrolides, and quinolones, have been found in the mariculture environment. They are usually found in mariculture and seawater sediments [59,60]. The detection abundances of sulfonamide (sul1 and sul2) and tetracycline (tetA and tetB) related to ARGs in cultured seawater are high, but there are large differences among different sampling sites [61,62]. Chen et al. [63] investigated the ARGs in 13 major mariculture farms in China and found that the absolute copy number and relative abundance of sulfonamide ARGs in cultured seawater and sediment were 4.3 and 2.3 times higher than those of tetracycline ARGs, respectively, of which the main ARG was sul2. To overcome the limitations of the existing ARG primers, Gao et al. [64] used Illumina high-throughput sequencing to analyze ARGs in the sediments of 10 aquaculture farms along the coast of China. The results showed that the abundance of bacA was the highest in all sediment samples, followed by mexF and mexB.
ARGs have also been detected in coastal waters [65,66,67,68,69,70], deep sea [71], estuary [72,73], mariculture [74,75,76,77,78,79], near shore area [80,81,82,83], polar environments [84,85,86], and other different marine environments. At present, resistant bacteria to tetracycline, sulfonamide, fluoroquinolone, third-generation cephalosporin, and chloramphenicol, as well as the related ARGs, have been widely detected in the marine environment [65,66,67,69,70]. Sulfonamide ARGs (sul1 and sul2) with high abundance were detected in the sediments of the Bohai Sea [65,66], the Yellow Sea [67], and the East China Sea [69] (Table 3), while macrolide and polypeptide ARGs were abundant in the sediments of the South China Sea [70]. Suzuki et al. [62] found that sul1 and sul2 are the main sulfa ARGs, while sul3 is transmitted by nonculturable bacteria in seawater. Moreover, some researchers proved that aquaculture will not only increase the abundance of ARGs in nearby waters but also increases the detection rate of multidrug-resistant bacteria in the environment far away from aquaculture areas [87,88,89]. Overall, ARGs are ubiquitous in the marine environment, and with the accumulation of antibiotics and the vertical and horizontal transfer of ARGs, bacteria in the marine environment will gradually acquire resistance to antibiotics.
5. ARGs in the Ballast Water
5.1. ARGs in the Ballast Water of Ships
In recent years, the proliferation of ARGs has gradually evolved into a serious global problem [5], and ARGs have been detected in sewage, rivers, inshore fishing grounds, marine sediments, and inshore seawater [56,60,61,62,63,64,90]. Seawater containing ARGs and human bacterial pathogens (HBPs) will be pumped into the ballast tank of the ship and then discharged to the port sea area of the destination [91]. This new pollutant can be transmitted to all parts of the world through ship ballast water, and the spread of ARGs will enhance the resistance of pathogenic bacteria to antibiotics.
Lv et al. [92] found that ARBs were detected in 25 out of 30 ship ballast water samples from different watersheds, with a detection rate of 83.3%; among these, penicillin-resistant bacteria had the highest detection rate, but their absolute abundance was relatively lower than that of hygromycin, chloramphenicol, tetracycline-like and vancomycin-resistant bacteria (Figure 1). According to the articles that were published, it seems that antibiotic stress may not be a particular technique to increase ARG production [92].
Penicillin was the first antibiotic discovered by human beings in the world, and it has been used for a long time. Therefore, the amount of penicillin-resistant bacteria in the global marine environment is relatively high [93]. However, other inexpensive and effective antibiotics, such as oxytetracycline, vancomycin, and tetracycline, have gradually developed and are now commonly used in aquaculture [94,95]. These antibiotics have a long half-life, slow metabolism, and nondegradable properties, which make them remain in the environment and continuously enrich ARGs. These persistent ARGs will aggravate the risk of ARG transmission in the environment after being discharged into the adjacent sea area. Schwartz et al. [96] concluded that the prevalence of vancomycin- and tetracycline-resistant bacteria is higher than that of penicillin-resistant bacteria.
The abundance of different ARGs in ballast water varies greatly. Lv et al. [92] found that the abundance of gentamicin-resistant bacteria was relatively lower than that of other ARBs in ballast water. Mohamed-Hatha et al. [97] and Souissi et al. [98] also proved that the abundance of gentamycin-resistant bacteria was low in marine environments. However, Lv et al. [99] conducted research on the ballast water of 28 ships in Chinese ports and proved that sul1 was the most abundant ARG with a concentration of 5.0 × 108 copies/mL. Niu et al. [66] and Stoll et al. [100] also found that sul1 had a high absolute abundance in surface water, and sulfonamide resistance genes dominated and persisted. Due to their widespread use, high discharge rate, high solubility, and persistence of sulfonamide antibiotics, sulfonamide-resistant bacteria can be gradually enriched in ballast water and persist in the aqueous environment for 5–10 years in the absence of selective pressure [73]. Quinolone antibiotics are a newer type of antibiotic, but bacteria that are resistant to them have been found in different types of seawater and sediments, which could be dangerous [101].
Many studies found that the ARG content in ballast water was higher than that in the samples collected in other environments [99,102]. Ballast water samples contained higher concentrations of ARGs than those in the Yellow Sea and Bohai Sea [61,103]. William et al. [47] found higher concentrations of tetM in ballast water than in seawater, suggesting the potential for ballast water to promote the global diffusion vector of ARGs. Furthermore, many HBPs have been detected in estuaries [73,104], ports [65,66], and coastal waters [59,60,61,62,63], which contain ARGs. The discharge of ballast water containing ARGs and HBPs into the seaport sea area of the destination will cause certain pollution risks to the water environment of the seaport area. The spread of ARGs will enhance the resistance of pathogenic bacteria to antibiotics, and HBPs will make humans sick [105,106]. Therefore, the prevention and control of pathogenic bacteria in ballast water have ushered in new challenges. The long-term existence and spread of ARGs in ballast water lead to the growth and spread of the drug resistance of pathogenic bacteria.
5.2. ARGs in Ballast Water Sediments
Any material that settles out of the ballast water tends to be deposited in the sediment at the bottom of the ballast tanks. Bottom sediments may contain a variety of marine organisms and microorganisms in their active and dormant stages, including pathogens. Prange et al. [107] found that the amount of sediment in the ballast tank of a merchant ship measured up to 200 tons. Living and nonliving organisms in ballast sediments need to be managed by appropriate methods; otherwise, they will pose risks. Hamer et al. [108] indicated that the thickness of the sediments in the double-bottomed ballast tanks of cargo ships increased to 30 cm after only two years of use. Most of the water left in the ships and sediments may contain non-native organisms. Gollasch et al. [109] recorded 990 species in ballast water sediment samples. Drake et al. [33] separated particle sludge that contained a variety of organisms in the ballast water sediment. Therefore, the ballast tank sediment provides a solid matrix rich in nutrients. The sediment in the ballast water tank will cause serious damage to the ecological environment of docks and port waters if it is not reasonably and effectively disposed of. More seriously, microorganisms that reside in sediments may exchange ARGs with one another. This has the potential to seriously affect the area surrounding the shipyard.
Lv et al. [28] studied ballast tank sediments and detected 10 ARGs in the sediments, among which the sulfonamide resistance genes (sul1 and sul2) had the highest absolute abundance, ranging from 8.36 × 107–1.61 × 109 copies/g to2.55 × 107–4.31 × 108 copies/g, respectively (Figure 2). Sulfonamide is the most commonly used antibiotic in marine aquaculture. Due to the particularities of aquaculture, the utilization rate of antibiotics is low, and the high concentration of antibiotics is eventually released into the surrounding marine water body or the sediments. Sulfonamide continues to spread and migrate, which, in turn, pollutes the marine environment [33] and affects coastal areas and marine sediments [110]. Ballast operations generally occur in offshore waters. Because of the relatively high abundance of sul1 and sul2 genes in offshore waters, sul1 and sul2 can be enriched in sediments. Lv et al. [28] found that the abundance of sulfonamide resistance genes was significantly higher than that of other ARGs, such as tetracycline resistance genes (tetM and tetQ). Chen et al. [69] and Lin et al. [111] found that tetracycline resistance genes were more easily removed than sulfonamide resistance genes. Gao et al. [112] investigated the pollution of ARGs in the aquaculture environments of Tianjin and proved that the abundance of sulfonamide resistance genes was much higher than others. There is a strong link between antibiotics and the ARGs they are linked to within the ballast water sediment environment. This means that antibiotic residues and ARGs may end up in the marine environment through the circulation of seawater, the suspension of sediment, and the movement of organisms. Because of this, it is important to constantly monitor the distribution and abundance of ARGs in ballast water, as well as develop appropriate methods to hinder the spread of ARGs via ballast water.
5.3. Influencing Factors of Resistance Genes
Recently, many studies have focused on the influence of a single type of antibiotic or environmental pressure on the distribution and spread of ARGs, aiming to reduce the ARGs and provide methods for the management of ARG contamination [33,109]. However, a variety of parameters, including physical factors and chemical factors, and specific components like temperature, pH, and alkalinity, all have an impact on the ARGs distributed in aquatic environments.
Some traditional pollutants, such as heavy metals, are selected to enrich antibiotic resistance coselection. It has been shown that the abundance of ARGs in the environment is related to the concentration of heavy metals [28]. Heavy metals can directly act on the microbial cell structure or indirectly affect bacteria by changing environmental conditions (such as pH, etc.), resulting in toxic effects on the bacteria. The long-term selective process of heavy metals can stimulate the self-defense mechanism of bacteria and gradually develop resistance. He et al. [113] found that both heavy metals and antibiotics can promote the transfer of resistance genes, and their combined effect is greater than their individual effects. Manaia et al. [25] believe that the influence of antibacterial residues or metals is the determinant of the fate of ARBs and ARGs in wastewater treatment. Yang et al. [20] summarized the distribution of antibiotics and ARGs in global lakes. It was found that chemical pollution, including antibiotics and heavy metals, affected the distribution of ARGs in lakes through selective pressure. Bacterial community is the main factor in forming drug resistance, followed by mobile genetic elements. Therefore, there was a certain correlation between heavy metal content and ARGs that influenced the abundance of ARGs.
Integrons (such as intI) often co-occur with antibiotic resistance genes due to the insertion of the antibiotic resistance gene. Lv et al. [28] found that intI was much more abundant than ARGs in shipyard sediments. The correlation between intI and ARGs was very strong. The high viability of intI enhances the propagation of ARGs through the horizontal gene transfer between microorganisms. Additionally, Zhang et al. [114] found that subinhibitory disinfectant doses may enhance the transfer of genes that cause bacteria to become resistant to antibiotics from one type of bacteria to another.
The content and distribution of antibiotic resistance genes in coastal waters are affected by human activities and environmental factors. Windil et al. [26] found that aquaculture changed the characteristics of ARGs and mobile genetic elements in Baltic Sea sediments. Lupo et al. [115] found that environmental pollution and water quality parameters affected the antibiotic resistance of bacteria in surface water. Rizzo et al. [23] and Bouki et al. [24] summarized the fate of ARB and ARGs in urban sewage treatment plants and believed that the effluent from urban sewage treatment plants was the main source of antibiotic release into the environment, causing ARGs and ARB to diffuse into the water environment. Antibiotic residues are the main driving force for the maintenance and transmission of ARB and its genes in wastewater. Moreover, it is also possible to change the formation and transmission rates under the action of antibiotics or heavy metals [59,116,117]. It is important to consider how the combined impacts of various antibiotics or combinations of antibiotics and environmental factors affect the characteristics of resistance gene contamination.
6. Changes in ARGs in Ballast Water after Disinfection
6.1. Ballast Water Disinfection
Now, ARGs have been found in a variety of water environments [118,119], but there is no special monitoring or control method for antibiotics and resistance genes. Many studies have focused on the removal of antibiotics and resistance genes in sewage treatment plants [120,121], while there are few studies on the treatment of antibiotics and resistance genes in ballast water. Therefore, it is necessary to emphasize the control of ARGs in ballast water. This chapter summarizes the research status of ARG-removal technology in aquatic environments in recent years and compares the advantages and disadvantages of each technology. Table 4 summarizes the removal effects of several treatment methods on ARGs and compares their advantages and disadvantages in detail.
6.2. Changes in ARGs in Ballast Water under Different Disinfection Technology
6.2.1. Chlorination Disinfection
Chlorination disinfection usually uses free chlorine and chlorine dioxide as disinfectants to eliminate pollutants, which is an important disinfection method for water treatment. Studies have shown that when the free chlorine concentration is 5 mg/L, 10 mg/L, and 15 mg/L, the pollutants, such as penicillin and tetracycline, in the urban sewage environment can be completely removed [131]. Chlorination may produce highly toxic disinfection byproducts. After chlorination disinfection, the toxicity of quinolones to photobacteria is enhanced [121]. In addition, a large amount of toxic disinfection byproducts can be generated after the chlorination disinfection of tetracyclines [132,133].
Free chlorine solution can act on target components such as DNA, RNA, and enzymes in the cytoplasm, causing the cleavage of resistance gene fragments [134]. Disinfection with higher doses of chlorine not only inactivates most donor bacteria but also damages the structure and function of the donor bacteria. Surviving donor bacteria have reduced extracellular secretions during the splicing process, making it difficult to transfer plasmids into the recipient bacteria, resulting in a significant reduction in the frequency of splice transfer [135]. However, it has also been proven that there was no significant removal of tetracycline resistance genes and erythromycin resistance genes after free chlorine disinfection (15 mg Cl2 min/L) [136]. In addition, some research indicated that a high concentration of free chlorine (300 mg Cl2 min/L) does not affect a decrease in resistance genes [42]. Moreover, some scholars claim that chlorination disinfection may promote the enrichment of resistance genes [137]. Liu et al. [138] found that the concentration of a-eARG increased 7.8 times after chlorine disinfection. It shows that the ARG released into the extracellular parts after chlorine disinfection tends to combine with the substances in the water to form a-eARG.
6.2.2. Ultraviolet Disinfection
Ultraviolet disinfection technology is widely used in water treatment because of its good effect, convenient use, and lack of chemical additions. Some studies indicate that ultraviolet (UV) irradiation can effectively remove antibiotics from aqueous solutions, especially antibiotics that are difficult to degrade [139]. UV can efficiently degrade lactams, fluoroquinolones, and tetracyclines [140].
Guo et al. [41] showed that, after UV treatment at an energy density of 5 mJ/cm2, the proportion of erythromycin-resistant bacteria in wastewater decreased, while the proportion of tetracycline-resistant bacteria increased. This result indicates that tetracyclin-resistant bacteria show higher tolerance to ultraviolet radiation and extrapolates that UV has a selectivity effect on ARBs [141,142]. In subsequent research, Guo et al. [143] found that UV disinfection (>10 mJ/cm2) can greatly inhibit horizontal gene transfer (HGT) and directly damage plasmid-containing ARGs, resulting in the death of the HGT donors and receptors. The above research shows that low-dose UV disinfection can effectively reduce ARGs in a water environment, but other researchers have reached the opposite conclusion. McKinney et al. [123] believed that high-dose UV disinfection could effectively remove ARGs. Through a series of experiments, they concluded that the UV dose required to reduce 3~4 log ARGs (mecA, vanA, tetA, and ampC) was 200~400 mJ/cm2 (Figure 3). However, the UV radiation dose in municipal sewage treatment plants is usually lower than 10 mJ/cm2 [143]. Therefore, in actual water treatment equipment, it is very difficult to achieve such a high UV dose. Also, ampicillin-resistant genes (ampC) are very resistant to UV disinfection, so this method cannot be applied to all ARGs [123].
6.2.3. Ozone Disinfection
Ozone is a strong oxidant that can react rapidly with organics in the form of direct oxidation. It can also indirectly oxidize most organic compounds with the hydroxyl radicals (OH) generated by water substrates [144]. In solutions of amoxicillin, doxycycline, ciprofloxacin, and sulfadiazine, the removal rate of the antibiotics accelerated with a continuous increase in the ozone concentration. Ozone (75 mg/L) can remove 95% of antibiotics, and its decomposition byproducts have no antibacterial activity or toxicity [145]. A total of eight antibiotics were completely removed when the ozone dose reached 125 mg O3/g for dissolved organic carbon (DOC) and the hydraulic retention was 40 min [146]. When the amount of ozone reached 657 mg/L and the water retention time was 120 min, 96% of the oxytetracycline was removed [146].
Compared with UV disinfection, ozone has a higher removal efficiency for antibiotics, yet it is easy to produce ozone byproducts that accumulate in water bodies. Ozone and ·OH oxidize and destroy bacterial cells. They then destroy the bacteria’s organelles, DNA, RNA, and other parts, directly [147]. Ozone disinfection kills a wide range of resistant bacteria, which causes the abundance of Escherichia coli, Staphylococcus, and Enterococcus to decrease sharply (0.73 mg O3/mg DOC, 20 min) [148]. Also, ozone treatment with 0.9 g O3/g DOC killed 60.2–98.9% of resistant bacteria like Enterococcus, Staphylococcus, Enterobacter, and Pseudomonas aeruginosa, with the abundance of ARGs, such as blaVIM, vanA, ampC, and ermB, decreasing by 18.7, 49.9, 69.8, and 99.3%, respectively [149]. Compared to UV disinfection and chlorination disinfection, ozone disinfection is better for eliminating resistant bacteria and resistant genes [123]. This may be because when a cell membrane is damaged, and proteins leak out, microbial gene fragments are exposed directly to ozone.
6.2.4. Free Radical Disinfection
Free radical disinfection techniques include photocatalysis and the Fenton reaction. Photocatalytic oxidation has a high removal rate of antibiotics and resistance genes in water and is a widely used method in free radical disinfection technology. Combining photocatalysis disinfection with the simultaneous dosing of UV irradiation (18.9 mJ/cm2) and H2O2 (40 mg/L) can completely degrade antibiotics such as ampicillin, erythromycin, and tetracycline [150]. Research on the elimination of antibiotics and their transformation products in wastewater suggested that the antibiotics in wastewater could be completely removed with a UV irradiation dose of 0.9 kJ/L for 90 min. Moreover, when UV was used in combination with H2O2/Fe, antibiotics could be completely removed within 30 min. In addition, research also shows that adding low doses of persulfate can get rid of antibiotics completely in 7–18 s [151].
Photocatalytic disinfection also effectively degrades resistance genes. Under UV/H2O2 treatment for 30 min, the abundance of resistance genes was reduced by 2.8–3.5 log [152]. Over the same treatment time, UV disinfection and chlorine disinfection could only reduce the abundance of resistance genes by 0.80–1.21 log and 1.65–2.28 log, respectively [122]. Although these results showed that UV/H2O2 treatment had a high removal rate for resistant bacteria and ARGs, the removal effect of free resistance genes in suspension was not obvious [153,154]. This could be due to a low oxidant dose or the inability of OH to react with other intracellular substances on DNA fragments [155,156]. When comparing OH to other disinfection methods, free radical technology is effective for antibiotic removal, resistant bacteria inactivation, and resistance gene degradation. However, because of the requirement for additional catalysts, free radical disinfection costs more than other methods, like UV and chlorination, because it needs more catalysts.
A single disinfection method cannot completely mineralize antibiotics or inactivate ARGs, and the continuous formation of disinfection byproducts may produce serious environmental toxicity. Some methods of disinfection, such as UV/H2O2 and UV Fenton, may be better for the removal of antibiotics and genes that make bacteria resistant to them. Zhang et al. [128] found that under optimal conditions, the UV/H2O2 method could reduce the abundance of resistance genes by 2.63–3.48 orders of magnitude. Hou et al. [157] found that among the treatment technologies, such as UV, ozone oxidation, Fenton oxidation, and Fenton/UV, Fenton oxidation and Fenton/UV are the most effective methods to remove the total bacteria and ARGs in water. Giannakis et al. [43] conducted a study on the elimination of antibiotic-resistant bacteria and ARGs by sunlight and the sunlight/Fenton method and found that both can effectively remove ARGs, and the removal rate of the sunlight/Fenton method was faster than that of the sunlight/Fenton method. Also, more attention should be paid to the byproducts of disinfection that are generated during the process. The ozone disinfection method can effectively remove antibiotics and resistance genes, but it can also easily produce disinfection byproducts. The UV photolysis of antibiotics may also generate more toxic intermediate products, thereby causing toxic effects on other organisms in the aquatic environment [158].
6.3. The Affect of Subinhibitory Concentrations of Disinfectants
The long-distance distribution of residual disinfectants through pipelines often decreases to subinhibitory levels (below minimum inhibitory concentrations [MICs]) or even becomes undetectable. Residual disinfectant in the retransmission pipeline is used to inhibit the regeneration of microorganisms in the pipeline. Therefore, ARGs and ARGs in treated ballast water should be exposed to relatively low concentrations of disinfectants. After a long voyage, the residual disinfectants in the ballast water may be at subinhibitory levels. The concentration of the disinfectant may also promote the transfer of antibiotic resistance. Xi et al. [159] found that ARGs were more abundant in tap and reclaimed water than in freshly-treated water. Anderssen et al. [160] indicated that subinhibitory levels of antibiotics could induce the transfer of ARGs, which may have an important impact on the spread of ARGs by participating in the reactive oxygen species (ROS) response system and SOS response pathway [114]. Guo et al. [143] investigated the effects of UV disinfection and chlorination on the transfer of ARGs and proved that UV disinfection and chlorination had a significant effect on their conjugate transfer (Figure 4). In contrast, the frequency of conjugation significantly increased with the subinhibitory dose (of up to 40 mg Cl2 min/L). Ye et al. [114] found that subinhibitory concentrations of disinfectants promote the conjugation transfer of ARGs within and between genera. Subinhibitory concentrations of chlorine, chloramine, and H2O2 promoted intragenus conjugation transfer by 3.4–6.4, 1.9–7.5, and 1.4–5.4 times, respectively (Figure 5) [114].
7. Conclusions
This paper summarizes the main types of ARGs in ballast water and ballast tank sediments and analyzes the factors that influence resistance genes. In addition, it analyzes the changes in ARGs after disinfection. The conclusions are shown as follows.
- Several resistance genes have been discovered in ballast water and sediments. This is generally due to the widespread use of antibiotics in aquaculture;
- The levels of ARGs in ballast water were higher than those in the samples collected from nearby marine environments. ARGs were found in ballast water samples from different sea areas. This result indicated that ballast water could promote the spread of ARGs, which should be further considered in the formulation of ballast water discharge standards;
- Disinfection treatment can enhance the removal of ARGs in ballast water. However, if the disinfectants are reduced to subinhibitory levels, a potential mechanism for the conjugate transfer of the ARGs will emerge.
ARG pollution is widespread in the water environment, and its horizontal gene transfer mode in the water environment is a hot topic of current research. However, there is little research on the transfer of ARGs in ballast water environments. Future research needs to further investigate the impact of ballast water discharge on the transmission of ARGs in local waters. Furthermore, some studies have shown that disinfectants with subinhibitory concentrations during water transportation can promote the conjugation and transfer of ARGs within and between genera. Therefore, it is necessary to determine the subinhibitory concentration of a disinfectant that causes the horizontal transfer of ARGs in ballast water. At present, most of the studies on ARG removal regarding disinfection technology report a single technology; there are few studies on ARG removal using more than two combined processes. The removal mechanism of ARGs by each disinfection process is different, and combination processes may produce synergistic effects.
Author Contributions
Writing—original draft preparation, J.G.; writing—review and editing, J.G. and S.; visualization, J.G. and Y.Z.; editing, B.J., C.W. and Y.Z.; supervision, Y.Z. and C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was founded by Natural Science Foundation of Shanghai, grant number No. 20ZR1438200, and the National Science Foundation of China, grant number No. 51778565.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lv, B.; Shi, J.; Li, T.; Ren, L.; Tian, W.; Lu, X.; Han, Y.; Cui, Y.; Jiang, T. Deciphering the characterization, ecological function and assembly processes of bacterial communities in ship ballast water and sediments. Sci. Total Environ. 2022, 816, 152721. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Q.; Xue, J.Z.; Xiao, N.Y.; Lv, B.Y.; Wu, H.X. Effects of holding time on the diversity and composition of potential pathogenic bacteria in ship ballast water. Mar. Environ. Res. 2020, 160, 104979. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Saebi, M.; Grey, E.K.; Corbett, J.J.; Chen, D.; Yang, D.; Wan, Z. Ballast water-mediated species spread risk dynamics and policy implications to reduce the invasion risk to the Mediterranean Sea. Mar. Pollut. Bull. 2022, 174, 113285. [Google Scholar] [CrossRef] [PubMed]
- Darling, J.A.; Martinson, J.; Gong, Y.G.; Oskum, S.; Pilgrim, E.; Lohan, K.M.P.; Carney, K.J.; Ruiz, G.M. Ballast Water Exchange and Invasion Risk Posed by Intracoastal Vessel Traffic: An Evaluation Using High Throughput Sequencing. Environ. Sci. Technol. 2018, 52, 9926–9936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gollasch, S.; David, M. Abiotic and biological differences in ballast water uptake and discharge samples. Mar. Pollut. Bull. 2021, 164, 112046. [Google Scholar] [CrossRef] [PubMed]
- Altug, G.; Gurun, S.; Cardak, M.; Ciftci, P.S.; Kalkan, S. The occurrence of pathogenic bacteria in some ships’ ballast water incoming from various marine regions to the Sea of Marmara, Turkey. Mar. Environ. Res. 2012, 81, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Werschkun, B.; Banerji, S.; Basurko, O.C.; David, M.; Fuhr, F.; Gollasch, S.; Grummt, T.; Haarich, M.; Jha, A.N.; Kacan, S.; et al. Emerging risks from ballast water treatment: The run-up to the International Ballast Water Management Convention. Chemosphere 2014, 112, 256–266. [Google Scholar] [CrossRef] [Green Version]
- Dong, P.Y.; Cui, Q.J.; Fang, T.T.; Huang, Y.; Wang, H. Occurrence of antibiotic resistance genes and bacterial pathogens in water and sediment in urban recreational water. J. Environ. Sci. 2019, 77, 65–74. [Google Scholar] [CrossRef]
- Cui, Q.J.; Huang, Y.; Wang, H.; Fang, T.T. Diversity and abundance of bacterial pathogens in urban rivers impacted by domestic sewage. Environ. Pollut. 2019, 249, 24–35. [Google Scholar] [CrossRef]
- Liu, X.; Steele, J.C.; Meng, X.Z. Usage, residue, and human health risk of antibiotics in Chinese aquaculture: A review. Environ. Pollut. 2017, 223, 161–169. [Google Scholar] [CrossRef]
- Hartmann, A.; Alder, A.C.; Koller, T.; Widmer, R.M. Identification of fluoroquinolone antibiotics as the main source of umuC genotoxicity in native hospital wastewater. Environ. Toxicol. Chem. 1998, 17, 377–382. [Google Scholar] [CrossRef]
- Jain, R.; Rivera, M.C.; Moore, J.E.; Lake, J.A. Horizontal gene transfer accelerates genome innovation and evolution. Mol. Biol. Evol. 2003, 20, 1598–1602. [Google Scholar] [CrossRef] [PubMed]
- Schluter, A.; Szczepanowski, R.; Puhler, A.; Top, E.M. Genomics of IncP-1 antibiotic resistance plasmids isolated from wastewater treatment plants provides evidence for a widely accessible drug resistance gene pool. FEMS Microbiol. Rev. 2007, 31, 449–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.K.; Ekka, R.; Mishra, M.; Mohapatra, H. Association study of multiple antibiotic resistance and virulence: A strategy to assess the extent of risk posed by bacterial population in aquatic environment. Environ. Monit. Assess. 2017, 189, 320. [Google Scholar] [CrossRef]
- Han, Y.; Wang, J.; Zhao, Z.L.; Chen, J.W.; Lu, H.; Liu, G.F. Fishmeal Application Induces Antibiotic Resistance Gene Propagation in Mariculture Sediment. Environ. Sci. Technol. 2017, 51, 10850–10860. [Google Scholar] [CrossRef]
- Ng, C.; Le, T.H.; Goh, S.G.; Liang, L.; Kim, Y.; Rose, J.B.; Yew-Hoong, K.G. A Comparison of Microbial Water Quality and Diversity for Ballast and Tropical Harbor Waters. PLoS ONE 2015, 10, e0154652. [Google Scholar] [CrossRef]
- Hoa, P.T.P.; Nonaka, L.; Viet, P.H.; Suzuki, S. Detection of the sul1, sul2, and sul3 genes in sulfonamide-resistant bacteria from wastewater and shrimp ponds of north Vietnam. Sci. Total Environ. 2008, 405, 377–384. [Google Scholar] [CrossRef]
- Ju, F.; Li, B.; Ma, L.; Wang, Y.; Huang, D.; Zhang, T. Antibiotic resistance genes and human bacterial pathogens: Co-occurrence, removal, and enrichment in municipal sewage sludge digesters. Water Res. 2016, 91, 1–10. [Google Scholar] [CrossRef]
- Beeton, M.L.; Chalker, V.J.; Jones, L.C.; Maxwell, N.C.; Spiller, O.B. Antibiotic Resistance among Clinical Ureaplasma Isolates Recovered from Neonates in England and Wales between 2007 and 2013. Antimicrob. Agents Chemother. 2016, 60, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Song, W.; Lin, H.; Wang, W.; Du, L.; Xing, W. Antibiotics and antibiotic resistance genes in global lakes: A review and meta-analysis. Environ. Int. 2018, 116, 60–73. [Google Scholar] [CrossRef]
- Hatosy, S.M.; Martiny, A.C. The Ocean as a Global Reservoir of Antibiotic Resistance Genes. Appl. Environ. Microbiol. 2015, 81, 7593–7599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felis, E.; Kalka, J.; Sochacki, A.; Kowalska, K.; Bajkacz, S.; Harnisz, M.; Korzeniewska, E. Antimicrobial pharmaceuticals in the aquatic environment—Occurrence and environmental implications. Eur. J. Pharmacol. 2020, 866, 172813. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef] [Green Version]
- Bouki, C.; Venieri, D.; Diamadopoulos, E. Detection and fate of antibiotic resistant bacteria in wastewater treatment plants: A review. Ecotoxicol. Environ. Saf. 2013, 91, 1–9. [Google Scholar] [CrossRef]
- Manaia, C.M.; Rocha, J.; Scaccia, N.; Marano, R.; Radu, E.; Biancullo, F.; Cerqueira, F.; Fortunato, G.; Iakovides, I.C.; Zammit, I.; et al. Antibiotic resistance in wastewater treatment plants: Tackling the black box. Environ. Int. 2018, 115, 312–324. [Google Scholar] [CrossRef] [PubMed]
- Muziasari, W.I.; Parnanen, K.; Johnson, T.A.; Lyra, C.; Karkman, A.; Stedtfeld, R.D.; Tamminen, M.; Tiedje, J.M.; Virta, M. Aquaculture changes the profile of antibiotic resistance and mobile genetic element associated genes in Baltic Sea sediments. FEMS Microbiol. Ecol. 2016, 92, fiw052. [Google Scholar] [CrossRef] [Green Version]
- Cordero, O.X.; Wildschutte, H.; Kirkup, B.; Proehl, S.; Ngo, L.; Hussain, F.; Le Roux, F.; Mincer, T.; Polz, M.F. Ecological Populations of Bacteria Act as Socially Cohesive Units of Antibiotic Production and Resistance. Science 2012, 337, 1228–1231. [Google Scholar] [CrossRef]
- Lv, B.; Cui, Y.; Tian, W.; Li, J.; Xie, B.; Yin, F. Abundances and profiles of antibiotic resistance genes as well as co-occurrences with human bacterial pathogens in ship ballast tank sediments from a shipyard in Jiangsu Province, China. Ecotoxicol. Environ. Saf. 2018, 157, 169–175. [Google Scholar] [CrossRef]
- Ruiz, G.M.; Fofonoff, P.W.; Carlton, J.T.; Wonham, M.J.; Hines, A.H. Invasion of coastal marine communities in North America: Apparent patterns, processes, and biases. Annu. Rev. Ecol. Syst. 2000, 31, 481–531. [Google Scholar] [CrossRef] [Green Version]
- Davidson, I.C.; Minton, M.S.; Carney, K.J.; Miller, A.W.; Ruiz, G.M. Pioneering patterns of ballast treatment in the emerging era of marine vector management. Mar. Policy. 2017, 78, 158–162. [Google Scholar] [CrossRef]
- Drillet, G. Protect aquaculture from ship pathogens. Nature 2016, 539, 31. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, D.M.; Smith, L.D.; Ruiz, G.M. The potential for intracoastal transfer of non-indigenous species in the ballast water of ships. Estuar. Coast. Shelf Sci. 1999, 48, 551–564. [Google Scholar] [CrossRef]
- Drake, L.A.; Doblin, M.A.; Dobbs, F.C. Potential microbial bioinvasions via ships’ ballast water, sediment, and biofilm. Mar. Pollut. Bull. 2007, 55, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Emami, K.; Askari, V.; Ullrich, M.; Mohinudeen, K.; Anil, A.C.; Khandeparker, L.; Burgess, J.G.; Mesbahi, E. Characterization of Bacteria in Ballast Water Using MALDI-TOF Mass Spectrometry. PLoS ONE 2012, 7, e38515. [Google Scholar] [CrossRef] [Green Version]
- Di Cesare, A.; Vignaroli, C.; Luna, G.M.; Pasquaroli, S.; Biavasco, F. Antibiotic-Resistant Enterococci in Seawater and Sediments from a Coastal Fish Farm. Microb. Drug Resist. 2012, 18, 502–509. [Google Scholar] [CrossRef]
- Akinbowale, O.L.; Peng, H.; Barton, M.D. Antimicrobial resistance in bacteria isolated from aquaculture sources in Australia. J. Appl. Microbiol. 2006, 100, 1103–1113. [Google Scholar] [CrossRef]
- Diehl, D.L.; LaPara, T.M. Effect of Temperature on the Fate of Genes Encoding Tetracycline Resistance and the Integrase of Class 1 Integrons within Anaerobic and Aerobic Digesters Treating Municipal Wastewater Solids. Environ. Sci. Technol. 2010, 44, 9128–9133. [Google Scholar] [CrossRef]
- Ghosh, S.; Ramsden, S.J.; LaPara, T.M. The role of anaerobic digestion in controlling the release of tetracycline resistance genes and class 1 integrons from municipal wastewater treatment plants. Appl. Microbiol. Biotechnol. 2009, 84, 791–796. [Google Scholar] [CrossRef]
- Zhang, T.; Yang, Y.; Pruden, A. Effect of temperature on removal of antibiotic resistance genes by anaerobic digestion of activated sludge revealed by metagenomic approach. Appl. Microbiol. Biotechnol. 2015, 99, 7771–7779. [Google Scholar] [CrossRef]
- Chen, J.; Wei, X.-D.; Liu, Y.-S.; Ying, G.-G.; Liu, S.-S.; He, L.-Y.; Su, H.-C.; Hu, L.-X.; Chen, F.-R.; Yang, Y.-Q. Removal of antibiotics and antibiotic resistance genes from domestic sewage by constructed wetlands: Optimization of wetland substrates and hydraulic loading. Sci. Total Environ. 2016, 565, 240–248. [Google Scholar] [CrossRef]
- Guo, M.-T.; Yuan, Q.-B.; Yang, J. Ultraviolet reduction of erythromycin and tetracycline resistant heterotrophic bacteria and their resistance genes in municipal wastewater. Chemosphere 2013, 93, 2864–2868. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.-B.; Guo, M.-T.; Yang, J. Fate of Antibiotic Resistant Bacteria and Genes during Wastewater Chlorination: Implication for Antibiotic Resistance Control. PLoS ONE 2015, 10, e0119403. [Google Scholar] [CrossRef] [PubMed]
- Giannakis, S.; Le, T.-T.M.; Entenza, J.M.; Pulgarin, C. Solar photo-Fenton disinfection of 11 antibiotic-resistant bacteria (ARB) and elimination of representative AR genes. Evidence that antibiotic resistance does not imply resistance to oxidative treatment. Water Res. 2018, 143, 334–345. [Google Scholar] [CrossRef]
- Ren, S.; Boo, C.; Guo, N.; Wang, S.; Elimelech, M.; Wang, Y. Photocatalytic Reactive Ultrafiltration Membrane for Removal of Antibiotic Resistant Bacteria and Antibiotic Resistance Genes from Wastewater Effluent. Environ. Sci. Technol. 2018, 52, 8666–8673. [Google Scholar] [CrossRef] [PubMed]
- Karaolia, P.; Michael-Kordatou, I.; Hapeshi, E.; Drosou, C.; Bertakis, Y.; Christofilos, D.; Armatas, G.S.; Sygellou, L.; Schwartz, T.; Xekoukoulotakis, N.P.; et al. Removal of antibiotics, antibiotic-resistant bacteria and their associated genes by graphene-based TiO2 composite photocatalysts under solar radiation in urban wastewaters. Appl. Catal. B 2018, 224, 810–824. [Google Scholar] [CrossRef]
- Jiang, Q.; Yin, H.; Li, G.; Liu, H.; An, T.; Wong, P.K.; Zhao, H. Elimination of antibiotic-resistance bacterium and its associated/dissociative blaTEM-1 and aac(3)-II antibiotic-resistance genes in aqueous system via photoelectrocatalytic process. Water Res. 2017, 125, 219–226. [Google Scholar] [CrossRef]
- Gerhard, W.A.; Gunsch, C.K. Higher normalized concentrations of tetracycline resistance found in ballast and harbor water compared to ocean water. Mar. Pollut. Bull. 2020, 151, 110796. [Google Scholar] [CrossRef]
- Sayinli, B.; Dong, Y.J.; Park, Y.; Bhatnagar, A.; Sillanpaa, M. Recent progress and challenges facing ballast water treatment-A review. Chemosphere 2022, 291, 132776. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. Transport of harmful marine microalgae via ship’s ballast water: Management and mitigation with special reference to the Arabian Gulf region. Aquat. Ecosyst. Health Manag. 2015, 18, 290–298. [Google Scholar] [CrossRef]
- Lv, B.Y.; Cui, Y.X.; Tian, W.; Feng, D.L. Composition and influencing factors of bacterial communities in ballast tank sediments: Implications for ballast water and sediment management. Mar. Environ. Res. 2017, 132, 14–22. [Google Scholar] [CrossRef]
- Yang, M.; Wang, Q.; Chen, J.; Wu, H. The occurrence of potential pathogenic bacteria on international ships’ ballast water at Yangshan Port, Shanghai, China. Mar. Pollut. Bull. 2022, 184, 114190. [Google Scholar] [CrossRef] [PubMed]
- Salleh, N.A.; Rosli, F.N.; Akbar, M.A.; Yusof, A.; Sahrani, F.K.; Razak, S.A.; Ahmad, A.; Usup, G.; Bunawan, H. Pathogenic hitchhiker diversity on international ships’ ballast water at West Malaysia port. Mar. Pollut. Bull. 2021, 172, 112850. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.; Goh, S.G.; Saeidi, N.; Gerhard, W.A.; Gunsch, C.K.; Gin, K.Y.H. Occurrence of Vibrio species, beta-lactam resistant Vibrio species, and indicator bacteria in ballast and port waters of a tropical harbor. Sci. Total Environ. 2018, 610, 651–656. [Google Scholar] [CrossRef]
- Rivera, I.N.G.; Souza, K.M.C.; Souza, C.P.; Lopes, R.M. Free-living and plankton-associated vibrios: Assessment in ballast water, harbor areas, and coastal ecosystems in Brazil. Front. Microbiol. 2013, 3, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobbs, F.C.; Goodrich, A.L.; Thomson, F.K.; Hynes, W. Pandemic Serotypes of Vibrio cholerae Isolated from Ships’ Ballast Tanks and Coastal Waters: Assessment of Antibiotic Resistance and Virulence Genes (tcpA and ctxA). Microb. Ecol. 2013, 65, 969–974. [Google Scholar] [CrossRef]
- Drake, L.A.; Meyer, A.E.; Forsberg, R.L.; Baier, R.E.; Doblin, M.A.; Heinemann, S.; Johnson, W.P.; Koch, M.; Rublee, P.A.; Dobbs, F.C. Potential invasion of microorganisms and pathogens via ‘interior hull fouling’: Biofilms inside ballast water tanks. Biol. Invasions 2005, 7, 969–982. [Google Scholar] [CrossRef]
- Faruque, S.M.; Albert, M.J.; Mekalanos, J.J. Epidemiology, genetics, and ecology of toxigenic Vibrio cholerae. Microbiol. Mol. Biol. Rev. 1998, 62, 1301–1314. [Google Scholar] [CrossRef] [Green Version]
- Kovalakova, P.; Cizmas, L.; McDonald, T.J.; Marsalek, B.; Feng, M.B.; Sharma, V.K. Occurrence and toxicity of antibiotics in the aquatic environment: A review. Chemosphere 2020, 251, 126351. [Google Scholar] [CrossRef]
- Zhao, Z.L.; Wang, J.; Han, Y.; Chen, J.W.; Liu, G.F.; Lu, H.; Yan, B.; Chen, S.S. Nutrients, heavy metals and microbial communities co-driven distribution of antibiotic resistance genes in adjacent environment of mariculture. Environ. Pollut. 2017, 220, 909–918. [Google Scholar] [CrossRef]
- Wang, J.H.; Lu, J.; Zhang, Y.X.; Wu, J.; Luo, Y.M.; Liu, H. Metagenomic analysis of antibiotic resistance genes in coastal industrial mariculture systems. Bioresour. Technol. 2018, 253, 235–243. [Google Scholar] [CrossRef]
- Suzuki, S.; Pruden, A.; Virta, M.; Zhang, T. Editorial: Antibiotic Resistance in Aquatic Systems. Front. Microbiol. 2017, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Nakanishi, S.; Tamminen, M.; Yokokawa, T.; Sato-Takabe, Y.; Ohta, K.; Chou, H.Y.; Muziasari, W.I.; Virta, M. Occurrence of sul and tet(M) genes in bacterial community in Japanese marine aquaculture environment throughout the year: Profile comparison with Taiwanese and Finnish aquaculture waters. Sci. Total Environ. 2019, 669, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Q.; Zheng, L.; Zhou, J.L.; Zhao, H. Persistence and risk of antibiotic residues and antibiotic resistance genes in major mariculture sites in Southeast China. Sci. Total Environ. 2017, 580, 1175–1184. [Google Scholar] [CrossRef]
- Gao, Q.X.; Li, Y.L.; Qi, Z.H.; Yue, Y.F.; Min, M.H.; Peng, S.M.; Shi, Z.H.; Gao, Y. Diverse and abundant antibiotic resistance genes from mariculture sites of China’s coastline. Sci. Total Environ. 2018, 630, 117–125. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Lu, J.; Wu, J.; Wang, J.H.; Lin, Y.C. Occurrence and distribution of antibiotic resistance genes in sediments in a semi-enclosed continental shelf sea. Sci. Total Environ. 2020, 720, 137712. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.G.; Zhang, K.; Zhang, Y. Occurrence and distribution of antibiotic resistance genes in the coastal area of the Bohai Bay, China. Mar. Pollut. Bull. 2016, 107, 245–250. [Google Scholar] [CrossRef]
- Na, G.S.; Zhang, W.R.; Zhou, S.Y.; Gao, H.; Lu, Z.H.; Wu, X.; Li, R.J.; Qiu, L.N.; Cai, Y.Q.; Yao, Z.W. Sulfonamide antibiotics in the Northern Yellow Sea are related to resistant bacteria: Implications for antibiotic resistance genes. Mar. Pollut. Bull. 2014, 84, 70–75. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, Y.; Wu, J.; Wang, J.; Zhang, C.; Lin, Y. Occurrence and spatial distribution of antibiotic resistance genes in the Bohai Sea and Yellow Sea areas, China. Environ. Pollut. 2019, 252, 450–460. [Google Scholar] [CrossRef]
- Chen, J.Y.; Su, Z.G.; Dai, T.J.; Huang, B.; Mu, Q.L.; Zhang, Y.M.; Wen, D.H. Occurrence and distribution of antibiotic resistance genes in the sediments of the East China Sea bays. J. Environ. Sci. 2019, 81, 156–167. [Google Scholar] [CrossRef]
- Chen, B.W.; Yang, Y.; Liang, X.M.; Yu, K.; Zhang, T.; Li, X.D. Metagenomic Profiles of Antibiotic Resistance Genes (ARGs) between Human Impacted Estuary and Deep Ocean Sediments. Environ. Sci. Technol. 2013, 47, 12753–12760. [Google Scholar] [CrossRef]
- Suzuki, S.; Ogo, M.; Miller, T.W.; Shimizu, A.; Takada, H.; Siringan, M.A.T. Who possesses drug resistance genes in the aquatic environment? Sulfarnethoxazole (SMX) resistance genes among the bacterial community in water environment of Metro-Manila, Philippines. Front. Microbiol. 2013, 4, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.G.; Zhao, Y.; Li, B.; Huang, C.L.; Zhang, S.Y.; Yu, S.; Chen, Y.S.; Zhang, T.; Gillings, M.R.; Su, J.Q. Continental-scale pollution of estuaries with antibiotic resistance genes. Nat. Microbiol. 2017, 2, 16270. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, K.; Wu, N.; Li, W.; Xu, W.; Zhang, Y.; Niu, Z. Estuarine sediments are key hotspots of intracellular and extracellular antibiotic resistance genes: A high-throughput analysis in Haihe Estuary in China. Environ. Int. 2020, 135, 105385. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Wu, X.M.; Yan, Q.P.; Ma, Y.; Huang, L.X.; Qin, Y.X.; Xu, X.J. Incidence of antimicrobial-resistance genes and integrons in antibiotic-resistant bacteria isolated from eels and aquaculture ponds. Dis. Aquat. Org. 2016, 120, 115–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, H.C.; Hu, X.J.; Wang, L.L.; Xu, W.J.; Xu, Y.; Wen, G.L.; Li, Z.J.; Cao, Y.C. Contamination of antibiotic resistance genes (ARGs) in a typical marine aquaculture farm: Source tracking of ARGs in reared aquatic organisms. J. Environ. Sci. Health B 2020, 55, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Xu, Y.B.; Xu, J.X.; Ling, J.Y.; Chen, J.L.; Zhou, J.L.; Zheng, L.; Du, Q.P. Antibiotic resistance genes (ARGs) in duck and fish production ponds with integrated or non-integrated mode. Chemosphere 2017, 168, 1107–1114. [Google Scholar] [CrossRef]
- Hong, B.; Ba, Y.; Niu, L.; Lou, F.; Zhang, Z.; Liu, H.; Pan, Y.; Zhao, Y. A Comprehensive Research on Antibiotic Resistance Genes in Microbiota of Aquatic Animals. Front. Microbiol. 2018, 9, 1617. [Google Scholar] [CrossRef]
- Thiang, E.L.; Lee, C.W.; Takada, H.; Seki, K.; Takei, A.; Suzuki, S.; Wang, A.; Bong, C.W. Antibiotic residues from aquaculture farms and their ecological risks in Southeast Asia: A case study from Malaysia. Ecosyst. Health Sust. 2021, 7, 1926337. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, Y.X.; Wu, J. Continental-scale spatio-temporal distribution of antibiotic resistance genes in coastal waters along coastline of China. Chemosphere 2020, 247, 125908. [Google Scholar] [CrossRef]
- Leonard, A.F.C.; Zhang, L.H.; Balfour, A.J.; Garside, R.; Hawkey, P.M.; Murray, A.K.; Ukoumunne, O.C.; Gaze, W.H. Exposure to and colonisation by antibiotic-resistant E. coli in UK coastal water users: Environmental surveillance, exposure assessment, and epidemiological study (Beach Bum Survey). Environ. Int. 2018, 114, 326–333. [Google Scholar] [CrossRef]
- Turgeon, P.; Michel, P.; Levallois, P.; Chevalier, P.; Daignault, D.; Crago, B.; Irwin, R.; McEwen, S.A.; Neumann, N.F.; Louie, M. Antimicrobial-resistant Escherichia coli in public beach waters in Quebec. Can. J. Occup. Ther. 2012, 23, E20–E25. [Google Scholar] [CrossRef] [Green Version]
- Kotlarska, E.; Luczkiewicz, A.; Pisowacka, M.; Burzynski, A. Antibiotic resistance and prevalence of class 1 and 2 integrons in Escherichia coli isolated from two wastewater treatment plants, and their receiving waters (Gulf of Gdansk, Baltic Sea, Poland). Environ. Sci. Pollut. Res. 2015, 22, 2018–2030. [Google Scholar] [CrossRef] [Green Version]
- Andrade, V.D.; Zampieri, B.D.; Ballesteros, E.R.; Pinto, A.B.; de Oliveira, A. Densities and antimicrobial resistance of Escherichia coli isolated from marine waters and beach sands. Environ. Monit. Assess. 2015, 187, 342. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.-Y.; Huang, F.-Y.; Zhao, Y.; Li, H.; Su, J.-Q. Increased levels of antibiotic resistance in urban stream of Jiulongjiang River, China. Appl. Microbiol. Biotechnol. 2015, 99, 5697–5707. [Google Scholar] [CrossRef]
- Tan, L.; Li, L.Y.; Ashbolt, N.; Wang, X.L.; Cui, Y.X.; Zhu, X.; Xu, Y.; Yang, Y.; Mao, D.Q.; Luo, Y. Arctic antibiotic resistance gene contamination, a result of anthropogenic activities and natural origin. Sci. Total Environ. 2018, 621, 1176–1184. [Google Scholar] [CrossRef]
- Cuadrat, R.R.C.; Sorokina, M.; Andrade, B.G.; Goris, T.; Davila, A.M.R. Global ocean resistome revealed: Exploring antibiotic resistance gene abundance and distribution in TARA Oceans samples. GigaScience 2020, 9, giaa046. [Google Scholar] [CrossRef]
- Eckert, E.M.; Di Cesare, A.; Stenzel, B.; Fontaneto, D.; Corno, G. Daphnia as a refuge for an antibiotic resistance gene in an experimental freshwater community. Sci. Total Environ. 2016, 571, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Ozaktas, T.; Taskin, B.; Gozen, A.G. High level multiple antibiotic resistance among fish surface associated bacterial populations in non-aquaculture freshwater environment. Water Res. 2012, 46, 6382–6390. [Google Scholar] [CrossRef]
- Jang, H.M.; Kim, Y.B.; Choi, S.; Lee, Y.; Shin, S.G.; Unno, T.; Kim, Y.M. Prevalence of antibiotic resistance genes from effluent of coastal aquaculture, South Korea. Environ. Pollut. 2018, 233, 1049–1057. [Google Scholar] [CrossRef]
- Campos, L.C.; Zahner, V.; Avelar, K.E.S.; Alves, R.M.; Pereira, D.S.G.; Brazil, J.M.V.; Freitas, F.S.; Salles, C.A.; Karaolis, D.K.R. Genetic diversity and antibiotic resistance of clinical and environmental Vibrio cholerae suggests that many serogroups are reservoirs of resistance. Epidemiol. Infect. 2004, 132, 985–992. [Google Scholar] [CrossRef]
- Liu, C.; Yoon, E.J.; Kim, D.; Shin, J.H.; Shin, J.H.; Shin, K.S.; Kim, Y.A.; Uh, Y.; Kim, H.S.; Kim, Y.R.; et al. Antimicrobial resistance in South Korea: A report from the Korean global antimicrobial resistance surveillance system (Kor-GLASS) for 2017. J. Infect. Chemother. 2019, 25, 845–859. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Jiang, T.; Wei, H.; Tian, W.; Han, Y.; Chen, L.; Zhang, D.; Cui, Y. Transfer of antibiotic-resistant bacteria via ballast water with a special focus on multiple antibiotic-resistant bacteria: A survey from an inland port in the Yangtze River. Mar. Pollut. Bull. 2021, 166, 112166. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, N.; Amal, M.N.A.; Saad, M.Z.; Yasin, I.S.M.; Zulkiply, N.A.; Mustafa, M.; Nasruddin, N.S. Virulence-associated genes and antibiotic resistance patterns of Vibrio spp. isolated from cultured marine fishes in Malaysia. BMC Vet. Res. 2019, 15, 176. [Google Scholar] [CrossRef] [PubMed]
- Guidi, L.R.; Santos, F.A.; Ribeiro, A.C.S.R.; Fernandes, C.; Silva, L.H.M.; Gloria, M.B.A. Quinolones and tetracyclines in aquaculture fish by a simple and rapid LC-MS/MS method. Food Chem. 2018, 245, 1232–1238. [Google Scholar] [CrossRef] [PubMed]
- Leal, J.F.; Henriques, I.S.; Correia, A.; Santos, E.B.H.; Esteves, V.I. Antibacterial activity of oxytetracycline photoproducts in marine aquaculture’s water. Environ. Pollut. 2017, 220, 644–649. [Google Scholar] [CrossRef]
- Schwartz, T.; Kohnen, W.; Jansen, B.; Obst, U. Detection of antibiotic-resistant bacteria and their resistance genes in wastewater, surface water, and drinking water biofilms. FEMS Microbiol. Ecol. 2003, 43, 325–335. [Google Scholar] [CrossRef]
- Hatha, A.A.M.; Neethu, C.S.; Nikhil, S.M.; Rahiman, K.M.M.; Krishnan, K.P.; Saramma, A.V. Relatively high antibiotic resistance among heterotrophic bacteria from arctic fjord sediments than water-Evidence towards better selection pressure in the fjord sediments. Polar Sci. 2015, 9, 382–388. [Google Scholar] [CrossRef]
- Souissi, M.; Laabidi, R.; Aissa, P.; Pringault, O.; Ben Said, O. Influence of Bizerte city wastewater treatment plant (WWTP) on abundance and antibioresistance of culturable heterotrophic and fecal indicator bacteria of Bizerte Lagoon (Tunisia). Ecotoxicol. Environ. Saf. 2018, 148, 201–210. [Google Scholar] [CrossRef]
- Lv, B.; Cui, Y.; Tian, W.; Wei, H.; Chen, Q.; Liu, B.; Zhang, D.; Xie, B. Vessel transport of antibiotic resistance genes across oceans and its implications for ballast water management. Chemosphere 2020, 253, 126697. [Google Scholar] [CrossRef]
- Stoll, C.; Sidhu, J.P.S.; Tiehm, A.; Toze, S. Prevalence of Clinically Relevant Antibiotic Resistance Genes in Surface Water Samples Collected from Germany and Australia. Environ. Sci. Technol. 2012, 46, 9716–9726. [Google Scholar] [CrossRef]
- Dong, P.; Wang, H.; Fang, T.; Wang, Y.; Ye, Q. Assessment of extracellular antibiotic resistance genes (eARGs) in typical environmental samples and the transforming ability of eARG. Environ. Int. 2019, 125, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.-p.; Yang, Y.; Lu, D.-p.; Niu, Z.-s.; Feng, J.-n.; Chen, Y.-r.; Tou, F.-y.; Garner, E.; Xu, J.; Liu, M.; et al. Biofilms as a sink for antibiotic resistance genes (ARGs) in the Yangtze Estuary. Water Res. 2018, 129, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Liang, X.; Huang, X.; Zhang, T.; Li, X. Differentiating anthropogenic impacts on ARGs in the Pearl River Estuary by using suitable gene indicators. Water Res. 2013, 47, 2811–2820. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.L.; Zhu, D.; Xu, Y.Y. Co-driving factors of tidal effect on the abundance and distribution of antibiotic resistance genes in the Yongjiang Estuary, China. Environ. Res. 2022, 213, 113649. [Google Scholar] [CrossRef]
- Neshani, A.; Zare, H.; Eidgahi, M.R.A.; Kakhki, R.K.; Safdari, H.; Khaledi, A.; Ghazvini, K. LL-37: Review of antimicrobial profile against sensitive and antibiotic-resistant human bacterial pathogens. Gene Rep. 2019, 17, 100519. [Google Scholar] [CrossRef]
- Zheng, J.; Gao, R.X.; Wei, Y.Y.; Chen, T.; Fan, J.Q.; Zhou, Z.C.; Makimilua, T.B.; Jiao, Y.N.; Chen, H. High-throughput profiling and analysis of antibiotic resistance genes in East Tiaoxi River, China. Environ. Pollut. 2017, 230, 648–654. [Google Scholar] [CrossRef]
- Prange, G.J.; Pereira, N.N. Ship Ballast Tank Sediment Reduction Methods. Nav. Eng. J. 2013, 125, 127–134. [Google Scholar]
- Casas-Monroy, O.; Roy, S.; Rochon, A. Dinoflagellate cysts in ballast sediments: Differences between Canada’s east coast, west coast and the Great Lakes. Aquat. Toxicol. 2013, 23, 254–276. [Google Scholar] [CrossRef]
- Gollasch, S.; Rosenthal, H.; Botnen, H.; Hamer, J.; Laing, I.; Leppakoski, E.; Macdonald, E.; Minchin, D.; Nauke, M.; Olenin, S.; et al. Fluctuations of zooplankton taxa in ballast water during short-term and long-term ocean-going voyages. Int. Rev. Hydrobiol. 2000, 85, 597–608. [Google Scholar] [CrossRef]
- Chen, B.; Liang, X.; Nie, X.; Huang, X.; Zou, S.; Li, X. The role of class I integrons in the dissemination of sulfonamide resistance genes in the Pearl River and Pearl River Estuary, South China. J. Hazard. Mater. 2015, 282, 61–67. [Google Scholar] [CrossRef]
- Lin, L.; Yuan, K.; Liang, X.; Chen, X.; Zhao, Z.; Yang, Y.; Zou, S.; Luan, T.; Chen, B. Occurrences and distribution of sulfonamide and tetracycline resistance genes in the Yangtze River Estuary and nearby coastal area. Mar. Pollut. Bull. 2015, 100, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Mao, D.; Luo, Y.; Wang, L.; Xu, B.; Xu, L. Occurrence of sulfonamide and tetracycline-resistant bacteria and resistance genes in aquaculture environment. Water Res. 2012, 46, 2355–2364. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Xu, Y.; Chen, J.; Ling, J.; Li, Y.; Huang, L.; Zhou, X.; Zheng, L.; Xie, G. Evolution of corresponding resistance genes in the water of fish tanks with multiple stresses of antibiotics and heavy metals. Water Res. 2017, 124, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gu, A.Z.; He, M.; Li, D.; Chen, J. Subinhibitory Concentrations of Disinfectants Promote the Horizontal Transfer of Multidrug Resistance Genes within and across Genera. Environ. Sci. Technol. 2017, 51, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Lupo, A.; Coyne, S.; Berendonk, T.U. Origin and evolution of antibiotic resistance: The common mechanisms of emergence and spread in water bodies. Front. Microbiol. 2012, 3, 18. [Google Scholar] [CrossRef] [Green Version]
- Vats, P.; Kaur, U.J.; Rishi, P. Heavy metal-induced selection and proliferation of antibiotic resistance: A review. J. Appl. Microbiol. 2022, 132, 4058–4076. [Google Scholar] [CrossRef]
- Lu, Z.; Na, G.; Gao, H.; Wang, L.; Bao, C.; Yao, Z. Fate of sulfonamide resistance genes in estuary environment and effect of anthropogenic activities. Sci. Total Environ. 2015, 527, 429–438. [Google Scholar] [CrossRef]
- Su, H.-C.; Liu, Y.-S.; Pan, C.-G.; Chen, J.; He, L.-Y.; Ying, G.-G. Persistence of antibiotic resistance genes and bacterial community changes in drinking water treatment system: From drinking water source to tap water. Sci. Total Environ. 2018, 616, 453–461. [Google Scholar] [CrossRef]
- Zheng, J.; Zhou, Z.; Wei, Y.; Chen, T.; Feng, W.; Chen, H. High-throughput profiling of seasonal variations of antibiotic resistance gene transport in a peri-urban river. Environ. Int. 2018, 114, 87–94. [Google Scholar] [CrossRef]
- Xu, Y.-B.; Hou, M.-Y.; Li, Y.-F.; Huang, L.; Ruan, J.-J.; Zheng, L.; Qiao, Q.-X.; Du, Q.-P. Distribution of tetracycline resistance genes and AmpC beta-lactamase genes in representative non-urban sewage plants and correlations with treatment processes and heavy metals. Chemosphere 2017, 170, 274–281. [Google Scholar] [CrossRef]
- Furukawa, T.; Hashimoto, R.; Mekata, T. Quantification of vancomycin-resistant enterococci and corresponding resistance genes in a sewage treatment plant. J. Environ. Sci. Health A 2015, 50, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhuang, Y.; Geng, J.; Ren, H.; Zhang, Y.; Ding, L.; Xu, K. Inactivation of antibiotic resistance genes in municipal wastewater effluent by chlorination and sequential UV/chlorination disinfection. Sci. Total Environ. 2015, 512, 125–132. [Google Scholar] [CrossRef]
- McKinney, C.W.; Pruden, A. Ultraviolet Disinfection of Antibiotic Resistant Bacteria and Their Antibiotic Resistance Genes in Water and Wastewater. Environ. Sci. Technol. 2012, 46, 13393–13400. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Hu, Y.; Jiang, L.; Yao, S.; Lin, K.; Zhou, Y.; Cui, C. Removal of antibiotic resistance genes and control of horizontal transfer risk by UV, chlorination and UV/chlorination treatments of drinking water. Chem. Eng. J. 2019, 358, 589–597. [Google Scholar] [CrossRef]
- Zhuang, Y.; Ren, H.; Geng, J.; Zhang, Y.; Zhang, Y.; Ding, L.; Xu, K. Inactivation of antibiotic resistance genes in municipal wastewater by chlorination, ultraviolet, and ozonation disinfection. Environ. Sci. Pollut. Res. 2015, 22, 7037–7044. [Google Scholar] [CrossRef] [PubMed]
- Asghari, F.B.; Dehghani, M.H.; Dehghanzadeh, R.; Farajzadeh, D.; Shanehbandi, D.; Mahvi, A.H.; Yaghmaeian, K.; Rajabi, A. Performance evaluation of ozonation for removal of antibiotic-resistant Escherichia coli and Pseudomonas aeruginosa and genes from hospital wastewater. Sci. Rep. 2021, 11, 24519. [Google Scholar] [CrossRef] [PubMed]
- Cengiz, M.; Uslu, M.O.; Balcioglu, I. Treatment of E. coli HB101 and the tetM gene by Fenton’s reagent and ozone in cow manure. J. Environ. Manag. 2010, 91, 2590–2593. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhuang, Y.; Geng, J.; Ren, H.; Xu, K.; Ding, L. Reduction of antibiotic resistance genes in municipal wastewater effluent by advanced oxidation processes. Sci. Total Environ. 2016, 550, 184–191. [Google Scholar] [CrossRef]
- Zhou, Z.; Shen, Z.; Cheng, Z.; Zhang, G.; Li, M.; Li, Y.; Zhan, S.; Crittenden, J.C. Mechanistic insights for efficient inactivation of antibiotic resistance genes: A synergistic interfacial adsorption and photocatalytic-oxidation process. Sci. Bull. 2020, 65, 2107–2119. [Google Scholar] [CrossRef]
- Cai, Q.; Hu, J. Effect of UVA/LED/TiO2 photocatalysis treated sulfamethoxazole and trimethoprim containing wastewater on antibiotic resistance development in sequencing batch reactors. Water Res. 2018, 140, 251–260. [Google Scholar] [CrossRef]
- Li, B.; Zhang, T. Different removal behaviours of multiple trace antibiotics in municipal wastewater chlorination. Water Res. 2013, 47, 2970–2982. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, T. pH significantly affects removal of trace antibiotics in chlorination of municipal wastewater. Water Res. 2012, 46, 3703–3713. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wei, D.; Du, Y. Acute toxicity evaluation for quinolone antibiotics and their chlorination disinfection processes. J. Environ. Sci. 2014, 26, 1837–1842. [Google Scholar] [CrossRef]
- Dodd, M.C. Potential impacts of disinfection processes on elimination and deactivation of antibiotic resistance genes during water and wastewater treatment. J. Environ. Monit. 2012, 14, 1754–1771. [Google Scholar] [CrossRef]
- Furukawa, T.; Jikumaru, A.; Ueno, T.; Sei, K. Inactivation Effect of Antibiotic-Resistant Gene Using Chlorine Disinfection. Water 2017, 9, 547. [Google Scholar] [CrossRef]
- Lin, W.; Zhang, M.; Zhang, S.; Yu, X. Can chlorination co-select antibiotic-resistance genes? Chemosphere 2016, 156, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Huang Jing, J.; Xi Jing, Y.; Hu Hong, Y.; Tang, F.; Pang Yu, C. Inactivation and Regrowth of Antibiotic-resistant Bacteria by PAA Disinfection in the Secondary Effluent of a Municipal Wastewater Treatment Plant. Biomed. Environ. Sci. 2013, 26, 865–868. [Google Scholar] [CrossRef]
- Liu, S.-S.; Qu, H.-M.; Yang, D.; Hu, H.; Liu, W.-L.; Qiu, Z.-G.; Hou, A.-M.; Guo, J.; Li, J.-W.; Shen, Z.-Q.; et al. Chlorine disinfection increases both intracellular and extracellular antibiotic resistance genes in a full-scale wastewater treatment plant. Water Res. 2018, 136, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Serna-Galvis, E.A.; Giraldo-Aguirre, A.L.; Silva-Agredo, J.; Florez-Acosta, O.A.; Torres-Palma, R.A. Removal of antibiotic cloxacillin by means of electrochemical oxidation, TiO2 photocatalysis, and photo-Fenton processes: Analysis of degradation pathways and effect of the water matrix on the elimination of antimicrobial activity. Environ. Sci. Pollut. Res. 2017, 24, 6339–6352. [Google Scholar] [CrossRef]
- Ding, Y.; Jiang, W.; Liang, B.; Han, J.; Cheng, H.; Haider, M.R.; Wang, H.; Liu, W.; Liu, S.; Wang, A. UV photolysis as an efficient pretreatment method for antibiotics decomposition and their antibacterial activity elimination. J. Hazard. Mater. 2020, 392, 122321. [Google Scholar] [CrossRef]
- Phattarapattamawong, S.; Chareewan, N.; Polprasert, C. Comparative removal of two antibiotic resistant bacteria and genes by the simultaneous use of chlorine and UV irradiation (UV/chlorine): Influence of free radicals on gene degradation. Sci. Total Environ. 2021, 755, 142696. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, B.A.; Vance, C.C.; Gentry, T.J.; Karthikeyan, R. Effects of chlorination and ultraviolet light on environmental tetracycline-resistant bacteria and tet(W) in water. J. Environ. Chem. Eng. 2017, 5, 777–784. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.T.; Yuan, Q.B.; Yang, J. Distinguishing Effects of Ultraviolet Exposure and Chlorination on the Horizontal Transfer of Antibiotic Resistance Genes in Municipal Wastewater. Environ. Sci. Technol. 2015, 49, 5771–5778. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Li, M.; Zhang, K.; Wang, N.; Wang, K.; Wang, H.; Meng, S.; Mu, R. Effect of ultrasound irradiation combined with ozone pretreatment on the anaerobic digestion for the biosludge exposed to trace-level levofloxacin: Degradation, microbial community and ARGs analysis. J. Environ. Manag. 2020, 262, 110356. [Google Scholar] [CrossRef]
- Alsager, O.A.; Alnajrani, M.N.; Abuelizz, H.A.; Aldaghmani, I.A. Removal of antibiotics from water and waste milk by ozonation: Kinetics, byproducts, and antimicrobial activity. Ecotoxicol. Environ. Saf. 2018, 158, 114–122. [Google Scholar] [CrossRef]
- Zheng, S.; Cui, C.; Liang, Q.; Xia, X.; Yang, F. Ozonation performance of WWTP secondary effluent of antibiotic manufacturing wastewater. Chemosphere 2010, 81, 1159–1163. [Google Scholar] [CrossRef]
- Chen, X.; Tang, R.; Wang, Y.; Yuan, S.; Wang, W.; Ali, I.M.; Hu, Z.-H. Effect of ultrasonic and ozone pretreatment on the fate of enteric indicator bacteria and antibiotic resistance genes, and anaerobic digestion of dairy wastewater. Bioresour. Technol. 2021, 320, 124356. [Google Scholar] [CrossRef]
- Lueddeke, F.; Hess, S.; Gallert, C.; Winter, J.; Guede, H.; Loeffler, H. Removal of total and antibiotic resistant bacteria in advanced wastewater treatment by ozonation in combination with different filtering techniques. Water Res. 2015, 69, 243–251. [Google Scholar] [CrossRef]
- Alexander, J.; Knopp, G.; Doetsch, A.; Wieland, A.; Schwartz, T. Ozone treatment of conditioned wastewater selects antibiotic resistance genes, opportunistic bacteria, and induce strong population shifts. Sci. Total Environ. 2016, 559, 103–112. [Google Scholar] [CrossRef]
- Pak, G.; Salcedo, D.E.; Lee, H.; Oh, J.; Maeng, S.K.; Song, K.G.; Hong, S.W.; Kim, H.-C.; Chandran, K.; Kim, S. Comparison of Antibiotic Resistance Removal Efficiencies Using Ozone Disinfection under Different pH and Suspended Solids and Humic Substance Concentrations. Environ. Sci. Technol. 2016, 50, 7590–7600. [Google Scholar] [CrossRef]
- Beretsou, V.G.; Michael-Kordatou, I.; Michael, C.; Santoro, D.; El-Halwagy, M.; Jager, T.; Besselink, H.; Schwartz, T.; Fatta-Kassinos, D. A chemical, microbiological and (eco)toxicological scheme to understand the efficiency of UV-C/H2O2 oxidation on antibiotic-related microcontaminants in treated urban wastewater. Sci. Total Environ. 2020, 744, 140835. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Chueca, J.; Varella della Giustina, S.; Rocha, J.; Fernandes, T.; Pablos, C.; Encinas, A.; Barcelo, D.; Rodriguez-Mozaz, S.; Manaia, C.M.; Marugan, J. Assessment of full-scale tertiary wastewater treatment by UV-C based-AOPs: Removal or persistence of antibiotics and antibiotic resistance genes? Sci. Total Environ. 2019, 652, 1051–1061. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Cao, S.; Jin, W.; Zhou, X.; Ding, W.; Tu, R.; Han, S.-F.; Wang, C.; Jiang, Q.; Huang, H.; et al. Inactivation of chlorine-resistant bacterial spores in drinking water using UV irradiation, UV/Hydrogen peroxide and UV/Peroxymonosulfate: Efficiency and mechanism. J. Clean. Prod. 2020, 243, 118666. [Google Scholar] [CrossRef]
- Ferro, G.; Guarino, F.; Castiglione, S.; Rizzo, L. Antibiotic resistance spread potential in urban wastewater effluents disinfected by UV/H2O2 process. Sci. Total Environ. 2016, 560, 29–35. [Google Scholar] [CrossRef]
- Guo, C.; Wang, K.; Hou, S.; Wan, L.; Lv, J.; Zhang, Y.; Qu, X.; Chen, S.; Xu, J. H2O2 and/or TiO2 photocatalysis under UV irradiation for the removal of antibiotic resistant bacteria and their antibiotic resistance genes. J. Hazard. Mater. 2017, 323, 710–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, Y.; Chung, H.J.; Di, D.Y.W.; Dodd, M.C.; Hur, H.-G.; Lee, Y. Inactivation efficiency of plasmid-encoded antibiotic resistance genes during water treatment with chlorine, UV, and UV/H2O2. Water Res. 2017, 123, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Chen, Z.; Gao, J.; Xie, Y.; Li, L.; Qin, S.; Wang, Q.; Mao, D.; Luo, Y. Simultaneous removal of antibiotics and antibiotic resistance genes from pharmaceutical wastewater using the combinations of up-flow anaerobic sludge bed, anoxic-oxic tank, and advanced oxidation technologies. Water Res. 2019, 159, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Su, C.; Zhou, J.W.; Xu, L.K.; Qian, Y.Y.; Chen, H. Effects and mechanisms of ultraviolet, chlorination, and ozone disinfection on antibiotic resistance genes in secondary effluents of municipal wastewater treatment plants. Chem. Eng. J. 2017, 317, 309–316. [Google Scholar] [CrossRef]
- Xi, C.; Zhang, Y.; Marrs, C.F.; Ye, W.; Simon, C.; Foxman, B.; Nriagu, J. Prevalence of Antibiotic Resistance in Drinking Water Treatment and Distribution Systems. Appl. Environ. Microbiol. 2009, 75, 5714–5718. [Google Scholar] [CrossRef] [Green Version]
- Andersson, D.I.; Hughes, D. Microbiological effects of sublethal levels of antibiotics. Nat. Rev. Microbiol. 2014, 12, 465–478. [Google Scholar] [CrossRef]
Figure 1.
Ten antibiotic-resistant bacteria (ARB) were found in ballast water [92].
Figure 1.
Ten antibiotic-resistant bacteria (ARB) were found in ballast water [92].

Figure 2.
Absolute concentrations of (a) sul1 and (b) sul2 in ballast tank sediments (BTS−1 to BTS−9 refer to ballast tank sediment samples) [28].
Figure 2.
Absolute concentrations of (a) sul1 and (b) sul2 in ballast tank sediments (BTS−1 to BTS−9 refer to ballast tank sediment samples) [28].

Figure 3.
UV (λ = 254 nm) disinfection curves for antibiotic resistance genes (ARGs). INC = Intracellular. EXC = extracellular [123].
Figure 3.
UV (λ = 254 nm) disinfection curves for antibiotic resistance genes (ARGs). INC = Intracellular. EXC = extracellular [123].

Figure 4.
Effect of (a) UV disinfection and (b) chlorination on the frequency of binding transfer [143].
Figure 4.
Effect of (a) UV disinfection and (b) chlorination on the frequency of binding transfer [143].

Figure 5.
Effect of (a) free chlorine, (b) chloramine and (c) H2O2 on intrageneric and transgeneric conjugation transfer [114].
Figure 5.
Effect of (a) free chlorine, (b) chloramine and (c) H2O2 on intrageneric and transgeneric conjugation transfer [114].


{kind=link}
{kind=link}
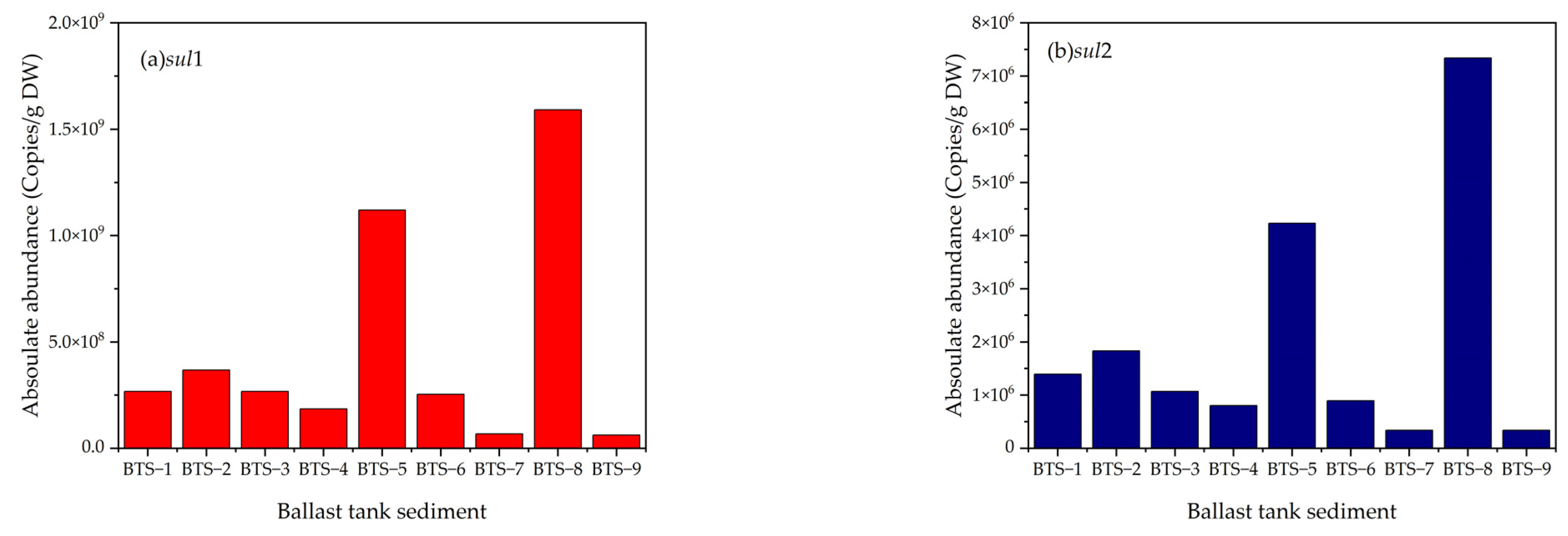{kind=link}
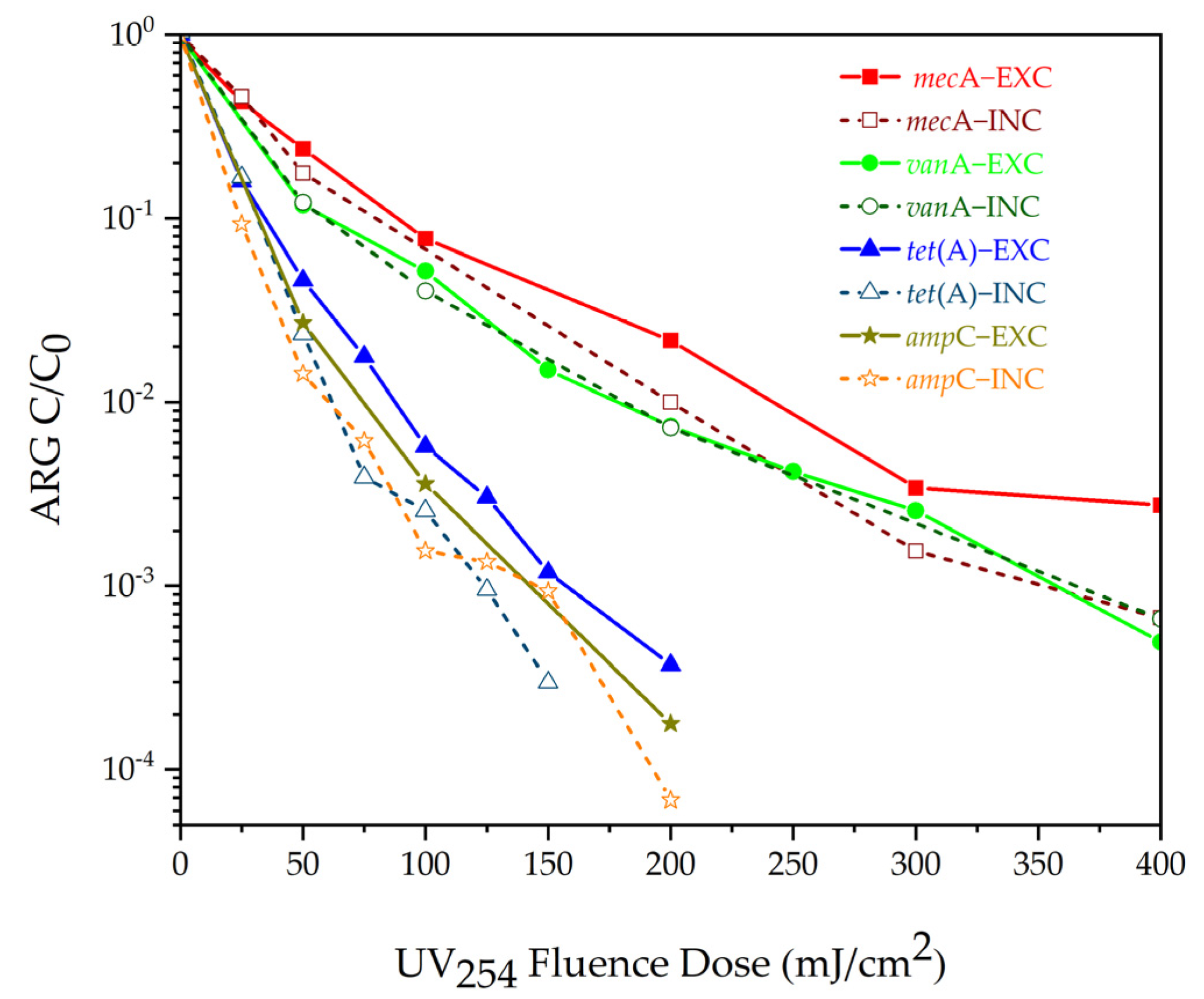{kind=link}
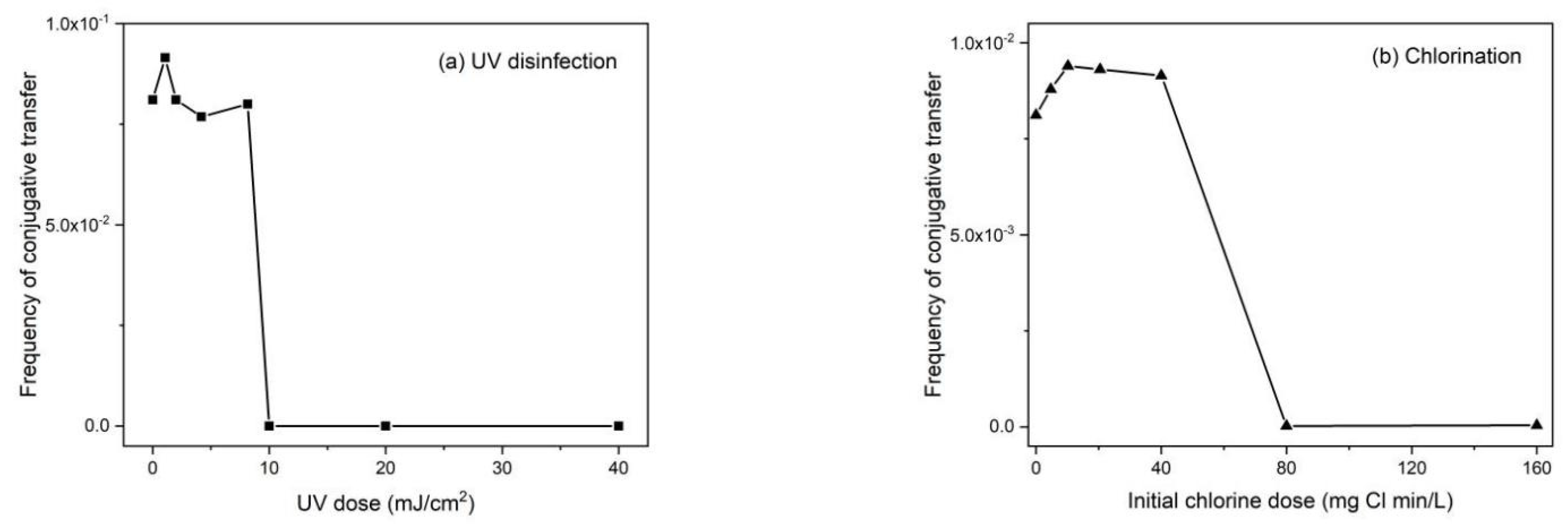{kind=link}
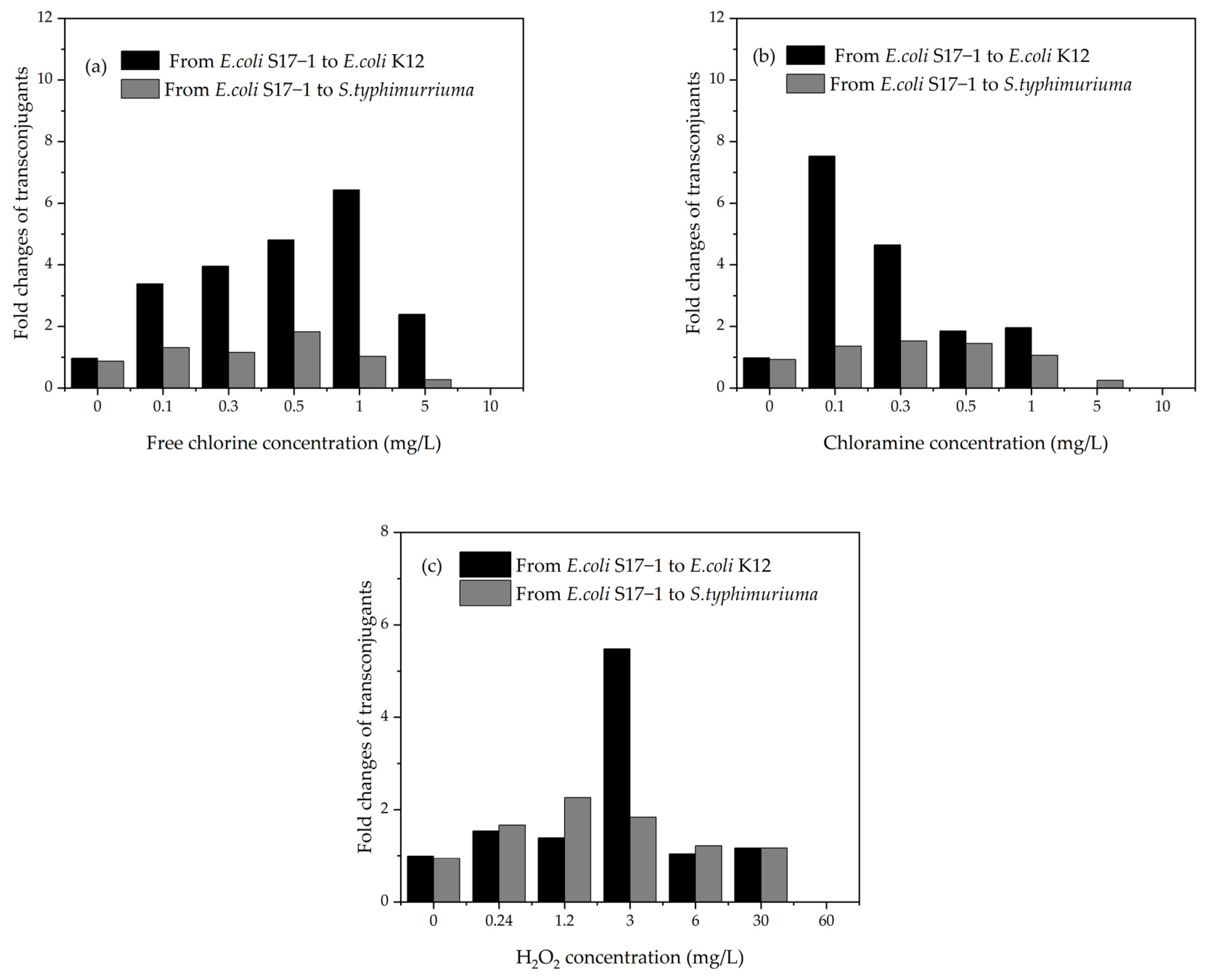{kind=link}
Table 1.
Microorganism discharge limitations based on the IMO D-2 Standard (IMO, 2004) [48].
Table 1.
Microorganism discharge limitations based on the IMO D-2 Standard (IMO, 2004) [48].
| Discharge Limitation Colony form Ingunit (cfu) | ||
|---|---|---|
| Indicator microbes | Toxicogenic Vibrio cholerae | <1 cfu per 100 mL |
| Escherichia coli (E.coli) | <250 cfu per 100 mL | |
| Intestinal Enterococci | <100 cfu per 100 mL | |
| Size of microorganism | ≥50 μm | <10 viable organisms per m3 |
| ≥10 μm and <50 μm | <10 viable organisms per mL |
Table 2.
Common types of antibiotics and representative drugs [58].
Table 2.
Common types of antibiotics and representative drugs [58].
| Category | Effect | Representative Drug |
|---|---|---|
| Tetracyclines | It is an antibiotic with a broad spectrum that destroys a variety of bacteria. Its primary function is to inhibit peptide chain growth and bacterial protein synthesis. | Tetracycline, oxytetracycline and chlortetracycline, etc. |
| Sulfonamides | It is a synthetic antibacterial drug with a broad antibacterial spectrum and stable properties. Its main function is to affect bacterial nucleoprotein synthesis by hindering the formation of dihydrofolic acid, thereby inhibiting bacterial reproduction. This kind of drug is a derivative of p-aminobenzene sulfonamide as a skeleton structure. | Sulfadiazine, Sulfathiazole, Sulfamethoxazole, etc. |
| Quinolones | It is an artificial, synthetic antibacterial agent with a broad antibacterial spectrum and strong antibacterial activity. It uses the 4-quinolone ring as the basic skeleton structure, and acts on bacterial deoxyribonucleic acid (DNA) helicase to damage chromosomes and kill bacteria. | Enrofloxacin, Norfloxacin, Ciprofloxacin, etc. |
| β-lactams | It mainly prevents the development of cell walls. It has the greatest diversity and is capable of treating most diseases. | Penicillins and cephalosporins, etc. |
| Aminoglycosides | It mainly inhibits the production of proteins by bacteria, increases the permeability of cell membranes, and causes chemicals within the cells to flow out. It removes bacteria. | Streptomycin, gentamicin, neomycin, kanamycin, etc. |
| Macrolides | It mainly inhibits bacterial protein synthesis. | Erythromycin, Tylosin, etc. |
| Amide alcohols | Broad-spectrum antibiotics produce an irreversible bond with the 50 S component of the bacterial ribosome, which then destroys microorganisms. This inhibits the formation of DNA white matter, acyl transfer, and peptide chain extension. | Chloramphenicol, Florfenicol, Thiamphenicol, etc. |
Table 3.
Types and distribution of ARGs and MGEs in marine environments.
| The Marine Environment | ARGs | MGEs | Ref. |
|---|---|---|---|
| Bohai Sea | blaTEM, sul1, sul2, qnrA, tetX, qnrS | intI1 | [65,66] |
| Yellow Sea | sul1, sul2, tetG, tetX | intI1 | [67,68] |
| East China Sea | sul1, sul2, sul3, tetC, tetW, dfrA1, dfrA13, floR, blaPSE-1 | intI1 | [69] |
| South China Sea | macB, acrB, tetX, bavA, arnA | [65,70] | |
| Manila Bay | sul1, sul2, sul3 | [71] | |
| Estuary | aacC, aadA5, mphA, oprD, oprJ, qacED1, tetG, aadA1 | intI1 | [72,73] |
| Mariculture | bacA, mexF, mexB, sul1, sul2, tetA, tetB, tetC, tetD, tetW, tetQ, tetO, floR, qnrA, qnrB, qnrD, aadA, blaTEM | intI1, intI2 | [62,63,64,74,75,76,77,78,79] |
| Beaches | tetA, tetB, qnrS, sul1, dfrA5, dfrA7 | intI1 | [80,81,82,83] |
| Polar | sul1, sul2, sul3, qnrB, tetD, tetG, aadA2, qacEdlta1 | intI1, cInl1, tnpA | [84,85,86] |
Table 4.
Advantages and disadvantages of different ballast water disinfection methods and removal effect of ARGs.
Table 4.
Advantages and disadvantages of different ballast water disinfection methods and removal effect of ARGs.
| Classification | Advantage | Shortcoming | Removal Effect of ARGs | Ref. |
|---|---|---|---|---|
| Ultraviolet disinfection | No byproducts, no chemical residue, noncorrosive water treatment. | There is photoreactivation. It is difficult to achieve a higher UV dose in actual production. | At a UV dose of 5 mJ cm−2, erythromycin ARGs and tetracycline ARGs decreased by an average of 3.0 ± 0.1 log and 1.9 ± 0.1 log, respectively. UV dose of 200~400 mJ cm−2 reduced ARGs (mecA, vanA, tetA and ampC) by 3~4 log; weak effect on tetX, sul1, tetG, and intI1. At 60 min, the removal efficiency of sul1-qPCR is greater than 3.50 log, and that of intI1-qPCR is greater than 4.00 log. | [41,122,123,124] |
| Chlorination disinfection | Simple operation, low cost. | It produces toxic byproducts, causes chemical residues. | Reduce erythromycin ARGs of 0.42 ± 0.12 log and tetracycline ARGs of 0.10 ± 0.02 log. sul1, tetX, tetG, and intI1 can remove 1.30~1.49 log. When the chlorine dosage is from 5–20 mg L−1, the removal of ARGs increases slowly, and a larger free chlorine dosage will lead to higher gene removal efficiency. | [42,122,123] |
| Ozonation | Strong oxidizability, fast reaction speed, no residue. | Ozone is easy to consume. It produces harmful byproducts. | sul1 and tetG decreased by 1.65~2.28 log. Under high ozone concentration, the removal rate of blactx was slightly higher than that of qnrS gene. | [125,126] |
| Fenton oxidation | Strong oxidation, simple operation, and low cost. | Reagent consumption is high. Excessive Fe2+ will affect the effluent quality and cause secondary pollution. | With an increase in Fenton reagent concentration, the band strength of tetM gene decreased gradually. The gel electrophoresis band strength of tetM decreased with an increase in oxidant dose; under the action of Fe2+/H2O2, the concentrations of sul1, tetX, and tetG decreased by 2.58~3.79 log, while under the action of UV/H2O2, they decreased by 2.8~3.5 log. | [43,127,128] |
| Photocatalytic oxidation | Tasteless and nontoxic, strong oxidation ability and complete sterilization, mild reaction conditions. | With the progress of the reaction, the catalyst will be deactivated, and the utilization rate of the light source will not be high. | 5.8 log mecA and 4.7 log ampC were removed. The degradation efficiency of floR, tetC, sul1, and intI1 was 97.82, 20.66, 99.45, and 93.67% respectively. The synthesized graphene-based photocatalyst successfully removed ampC and significantly reduced the ecfX abundance of Pseudomonas aeruginosa. | [44,45,67,129,130] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guo, J.; Jiang, B.; Sumita; Wu, C.; Zhang, Y.; Li, C. Review of the Distribution and Influence of Antibiotic Resistance Genes in Ballast Water. Water 2022, 14, 3501. https://doi.org/10.3390/w14213501
AMA Style
Guo J, Jiang B, Sumita, Wu C, Zhang Y, Li C. Review of the Distribution and Influence of Antibiotic Resistance Genes in Ballast Water. Water. 2022; 14(21):3501. https://doi.org/10.3390/w14213501
Chicago/Turabian StyleGuo, Jiaqi, Bo Jiang, Sumita, Chengzhang Wu, Yunshu Zhang, and Cong Li. 2022. "Review of the Distribution and Influence of Antibiotic Resistance Genes in Ballast Water" Water 14, no. 21: 3501. https://doi.org/10.3390/w14213501
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.