
The Occurrence and Role of Tetrasphaera in Enhanced Biological Phosphorus Removal Systems


Faculty of Civil and Environmental Engineering, Gdansk University of Technology, Narutowicza 11/12, 80-233 Gdansk, Poland
*
Author to whom correspondence should be addressed.
Water 2022, 14(21), 3428; https://doi.org/10.3390/w14213428
Submission received: 28 August 2022
/
Revised: 21 October 2022
/
Accepted: 22 October 2022
/
Published: 28 October 2022
(This article belongs to the Section Wastewater Treatment and Reuse)
Abstract
:The application of enhanced biological phosphorus removal (EBPR) in wastewater treatment plants (WWTPs) has commonly been utilized worldwide. However, the optimum efficiency has not been realized over the past decades, prompting many studies and publications. The limitations, especially comprehension of the abundance and actual potential of polyphosphate-accumulating organisms (PAOs), are not fully understood. Recently identified putative PAOs, Tetrasphaera, present a vast metabolic versatility compared to Candidatus Accumulibacter. The characterisation of Tetrasphaera unique abilities to utilize various carbon substrates, volatile fatty acids production and consistent high abundance, presents potential boosts towards the process efficiency improvement. This paper provides the existing knowledge on the physiology, morphology and genetic description of PAOs with a special attention to the current state of research on Tetrasphaera and its potential. In addition, process conditions and their influence on the microbial activities in EBPR systems are discussed.
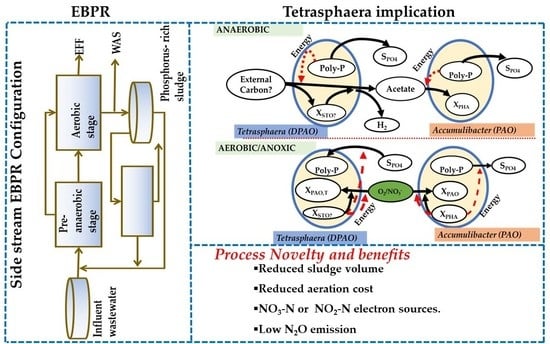
1. Introduction
Enhanced biological phosphorus removal (EBPR) has emerged as the most powerful phosphorus (P) removal process during municipal and industrial wastewater treatment. For a long time, the EBPR has been considered as one of the most complex processes involved in the metabolic activity of activated sludge systems and has shown promise in terms of the cost, reliability and sustainability [1]. Recent years have brought many research contributions to expand knowledge and improve the process efficiency based on the recognition of the pathways and microorganisms involved.
The EBPR in biological nutrient removal (BNR) systems is mainly carried out by a group of microorganisms known as polyphosphate-accumulating organisms (PAOs). Conventionally, P removal via PAO activity is achieved by triggering anaerobic–aerobic conditions, which considerably increase operational costs related to energy consumption by aerators. The focus has recently been on P removal by denitrifying PAOs (DPAOs) under anaerobic–anoxic conditions to reduce the costs. The DPAOs are capable of using alternative electron sources (nitrate or nitrite) to metabolize intracellular organic compounds under anoxic conditions, and P uptake and denitrification is performed simultaneously [2,3,4,5].
Due to the growing interest in the implementation of P removal under anaerobic-anoxic conditions, a special attention has been paid to the microorganisms responsible for that process. Representatives of the genus Tetrasphaera are among the recently confirmed putative denitrifying PAO attracting attention of the scientific community. Members of Tetrasphaera are able to perform either denitrification or aerobic respiration, depending on the local environmental conditions [6]. All currently characterized Tetrasphaera isolates have proven the capability of reducing nitrate only to nitrite, whereas some members revealed the ability to reduce nitric acid to nitrous oxide. Moreover, the Tetrasphaera group is capable of carrying out the complete physiological EBPR process compared to other known PAOs, whose activity is more dependent on interspecies relationships [7].
The key aspects of Tetrasphaera have been studied intensively, including their classification and taxonomy, development of methods for detection [8], abundance in wastewater treatment plants (WWTPs) [9,10], functions in EBPR and biochemistry [6,11,12]. The ubiquitous occurrence of Tetrasphaera in diverse ecological niches, the use of various carbon sources and the ability to produce volatile fatty acids (VFAs) show their extraordinary metabolic potential [4,13].
Interactions and competition between DPAOs and other functional microbial groups in the anaerobic-aerobic cycle enable P removal optimization in activated sludge systems [14,15,16]. In addition, the interest in DPAO ecophysiology, in particular in the context of Tetrasphaera activity related to nitrous oxide emissions, was significant [3]. An emerging approach to enhance the full-scale EBPR is optimization by the application of mathematical modelling. The conventional models are thought to favor Ca. Accumulibacter over other PAOs, such as Ca. Halomonas phosphatis, Tessaracoccus, as well Tetrasphaera [8]. To ensure the highest prediction accuracy of the model, it is strongly recommended to extend and develop currently available models for multiple PAO groups, differentiated in terms of the growth rate and physiology.
This study aims to consolidate the existing knowledge on the role of Tetrasphaera by reviewing their physiological and metabolic characteristics, the occurrence and abundance in WWTPs and factors influencing their growth. Additionally, it highlights the knowledge gaps and research challenges in the field of EBPR microbiology as well as presents scientific approaches to overcome these limitations, including the meta-analysis and models. Furthermore, this study investigates the abundance of Tetrasphaera and their response to the local process conditions, such as dissolved oxygen (DO) concentration, pH, temperature and influent characteristics.
2. Historical Perspective of Microorganisms Involved in EBPR
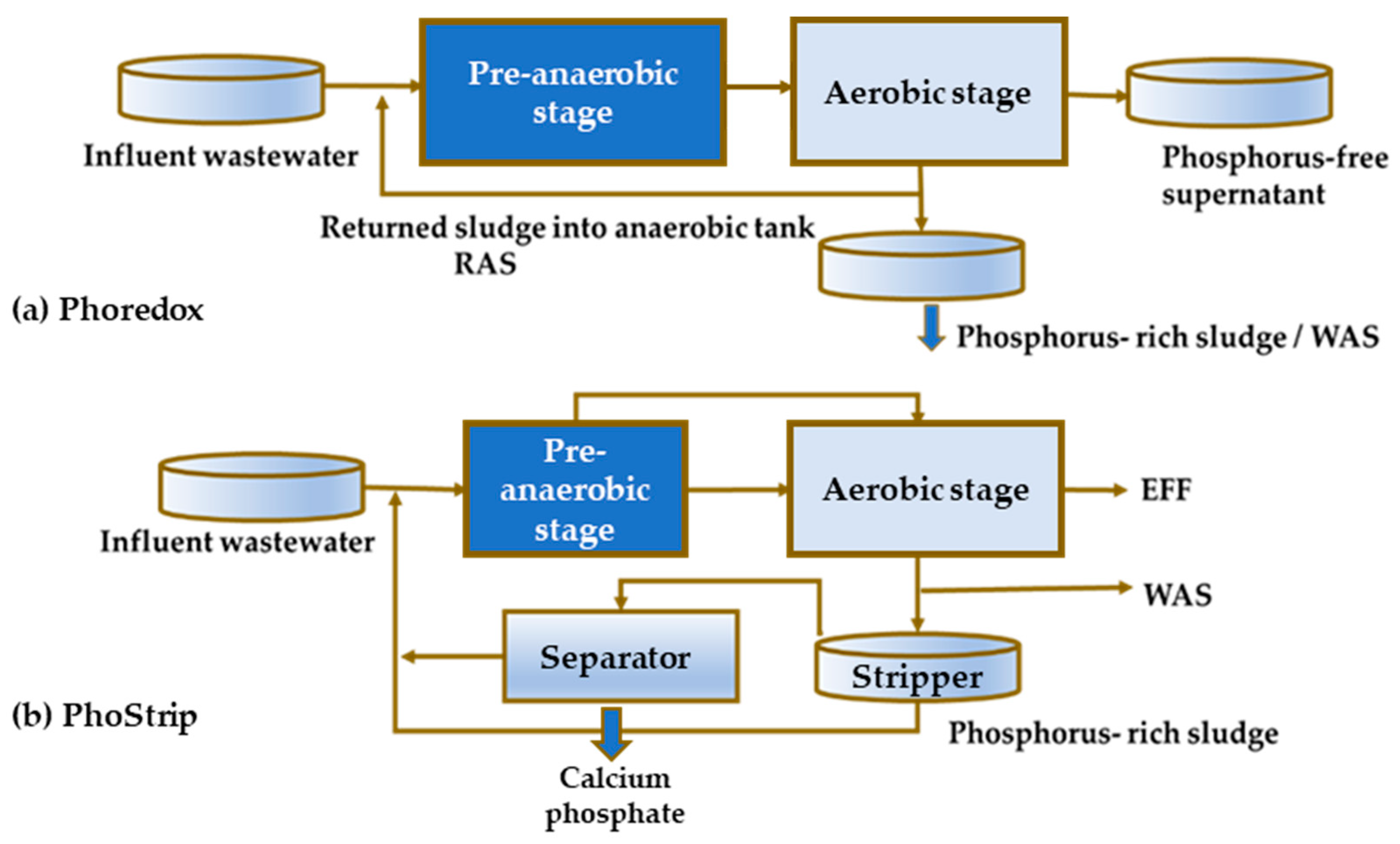
The first observations of EBPR date back to the mid-20th century and the findings obtained in laboratory scale experiments [17] and to a minor extent at full scale plants [18]. The principles of EBPR were formulated by Barnard [19,20], whose experiments clarified the need for anaerobic contact between activated sludge and influent wastewater prior to aerobic treatment to accomplish P removal. Subsequently, Barnard [21] used the term Phoredox to represent any process with an anaerobic/aerobic sequence to promote the EBPR technology concept (Figure 1a).
The performance and start-up process of the first full scale Phoredox system, which was launched in 1973, was briefly reported by Levin et al. [22]. In parallel, an alternative side-stream P removal technology, called PhoStrip, [1]. was developed based on the separation of the enriched side stream liquor treated with lime (Figure 1b).
While the nature of P removal was initially considered as chemical, Fuhs and Chen [23] found Acinetobacter as the primary microorganisms responsible for EBPR. These organisms responded to VFA in the influent wastewater under anaerobic conditions by releasing stored phosphate. Bacteria affiliated to Acinetobacter were considered as the key PAO responsible for the EBPR, mainly due to the limitations of the cultivation techniques applied for the microbial characterization at that time [11]. Significant advances in the microbial research have been achieved over the years and novel bacterial groups involved in the P metabolism were identified. Those microorganisms were able to store P in their cells in the form of energy-rich polyphosphates, resulting in the P content as high as 20 to 30 percent by dry weight [1]. The anaerobic zone free of nitrate and DO was found to favour the PAOs activity over other heterotrophs. In the following years, Betaproteobacteria were determined as the dominant bacterial group in the P metabolism [24], as well as the presence of Rhodocyclus-related bacteria was observed and linked with EBPR [25,26]. Subsequently, modern microbial tools without the cultivation step, established Ca. Accumulibacter as the most important member of PAOs, with the share ranging from 0.6 to 33.1% [27,28]. Insights into the biochemical characteristics of the Ca. Accumulibacter were applied to propose mathematical models of the EBPR within Activated Sludge Model [29]. Basic metabolic models are based on the assumption that PAOs exhibit intracellular phosphorus and energy storage in the form of poly-P and polyhydroxyalkanoates (PHA), respectively [30]. The stored PHA provide energy for the growth of PAOs when exposed to anoxic condition due to the capability of simultaneous denitrification and P uptake.
The novel microbial tools comprise deoxyribonucleic acid (DNA) and ribonucleic acid (RNA) polymorphism analyses (e.g., 16S rRNA high-throughput gene sequencing, metagenomics, fluorescence in situ hybridization (FISH) and their modification) as well as flow cytometry and Raman spectroscopy. Along with the development of those tool, the contribution of particular bacterial groups in EBPR could be revised greatly [4,28,31].
The next advance in understanding the microbiology of EBPR was the discovery of new PAO and capability of simultaneous P and nitrogen (N) removal under anoxic conditions. Genus Tetrasphaera is the most recently confirmed putative PAO. From the first characterization of the isolated Tetrasphaera strain [32,33], it was found that representatives of this genera show a large and often predominant number in full-scale WWTPs [7,34]. Moreover, versatile metabolic capabilities of Tetrasphaera have been recognised, including the capability of fermenting glucose and amino acids to produce VFA in the anaerobic zone, thereby enhancing a pool of the available substrates for EBPR. The main difference with respect to the typical PAO is that Tetrasphaera are capable of storing other (than PHA) intracellular compounds and use nitrate, but not nitrite, in addition to DO as an electron acceptor [1]. The characterization of Tetrasphaera and their assignment to PAO drew attention to this group, especially in terms of the competition and interaction with Ca. Accumulibacter [3,35].
In the recent years, several new genera have been proposed as potential PAOs, including Dechloromonas or Candidatus Microthrix, but only members of the betaproteobacterial genera Ca. Accumulibacter [7] and the actinobacterial genus Tetrasphaera were consistently found in high abundances in full-scale EBPR plants [34]. For instance, it was proven that approximately 24–70% of total P removed in Danish WWTPs was directly attributed to Ca. Accumulibacter and Tetrasphaera [7]. The relative abundances of other PAO within activated sludge are usually significantly lower. In the study by Seviour and McIlroy [36], the relative abundance of Acinetobacter reached 1.2%, whereas the relative abundances below 1% were reported for Dechloromonas, another newly characterized PAO group [37].
3. Morphology, Physiology and Phylogeny of Tetrasphaera
Tetrasphaera is a bacterial genus that belongs to the Intrasporangiaceae family within the Actinomycetia class and initially contained eight proposed species [38]. Zhang and Kinyua [8] listed the following representatives of the genus Tetrasphaera: Tetrasphaera japonica, Tetrasphaera australiensis, Tetrasphaera elongata, Tetrasphaera jenkinsii, Tetrasphaera vanveenii, Tetrasphaera veronensis, Tetrasphaera duodecadis and Tetrasphaera remsis. Based on the distinct morphological, biochemical characteristics (the capability of PHA storage) and probes applied for detection via FISH), particular Tetrasphaera representatives were divided into three clades: clade I including T. elongata and T. duodecadis; clade II including T. jenkinsii, T. australiensis, T. veronensis and the filamentous Candidatus Nostocoida limicola, clade III containing uncultured clones [39]. However, the phylogenetic classification of the genus Tetrasphaera has not been definitively clarified and is subject to continuous revision. For instance, important updates to representatives of clade III were provided by Singleton et al. [40], who postulated separation of the two novel genera Ca. Phosphoribacter and Ca. Lutibacillus from the genus Tetrasphaera. Similarly, the whole genome sequence analyses by Nouioui et al. [41] revealed the need to reclassify T. duodecadis, T. remsis, and T. elongata into Phycicoccus duodecadis, Knoellia remsis and Phycicoccus elongatus, respectively.
First important insights into the morphology and biochemical characteristics of Tetrasphaera have been provided by Maszenan et al. [33]. Representatives of Tetrasphaera were characterized as an aerobic, Gram-positive cocci, mostly clustered in tetrads and less often in pairs. Due to the lack of flagella, Tetrasphaera were considered as a non-motile. Despite P removal activity, members of T. japonica and T. australiensis did not reflect the ability to store PHA granules, which suggested the role of other intracellular compounds in their EBPR metabolism.
Further cultivation studies by Hanada et al. [32] on biomass from a EBPR system led to the characterization of new species T. elongata., which showed a versatile morphology (oval to rod-shaped) and capability to metabolize wide groups of complex organic compounds, including sugars, alcohols and organic acids. By a positive result of Neisser staining, intracellular polyphosphate granules occurrence has been confirmed and provide evidence of the characterized isolate contribution to P metabolism. Further studies focused on pure cultures of T. elongata have demonstrated archetypical PAO characteristics [8].
A consensus is yet to be built on generally accepted biochemical transformation models for EBPR by Tetrasphaera, in particular recognition of the storage compounds involved in EBPR. To date, the capability of intracellular PHA storage has been identified in several Tetrasphaera species, including T. japonica T. jenkinsii T. vanveenii T. veronensis [14]. However, the ability of Tetrasphaera species for anaerobic P release, aerobic/anoxic uptake patterns, and accumulation of intracellular poly-P granules remain inconclusive [12].
Important updates into the characterization of the Tetrasphaera metabolism have been by Close et al. [42], who conducted their study on an enriched Tetrasphaera culture. In that study, more complex nature of the intracellular compounds cycling within EBPR has been highlighted, where PHA storage was accompanied with the complex amino acids’ metabolism, mainly related to aspartic and glutamic acid accumulation within the cell. Moreover, the authors identified that polyhydroxyvalerate (PHV), rather than polyhydroxybutyrate (PHB) (typical for PAO) seems to be more specific feature of Tetrasphaera metabolism within EBPR. Despite PHA and intracellular amino acids, members of Tetrasphaera have shown the ability to store glycogen [11,12]. In contrast to the other PAOs, Tetrasphaera synthesize glycogen and release phosphate under anaerobic conditions, then metabolize glycogen as an energy source to produce polyphosphate after transition to the aerobic/anoxic environment [2]. Those metabolic features makes this bacterial group versatile in terms of ecophysiology [38].
As postulated by Barnard et al. [2], Tetrasphaera exhibits an alternative P depletion mechanism compared to Ca. Accumulibacter and other “traditional” PAOs with a potentially more effective P removal efficiency. There is scarce evidence concerning the ability of Tetrasphaera to perform denitrification within EBPR. The existing studies emphasize the fact that Tetrasphaera are less capable of removing P under anoxic conditions compared to Ca. Accumulibacter [6]. On the other hand, Tetrasphaera showed a high performance of N removal (>80%), indicating the ability of Tetrasphaera to reduce NO3 -N to NO2 -N similar to their counterpart DPAOs. In the studies by Marques et al. [6], a strong denitrifying activity by Tetrasphaera was observed in a mixed culture, where Tetrasphaera contributed to 60% of the general bacterial population. Tetrasphaera had the capability of denitrification, even though the specific P-uptake rate under anoxic conditions was insignificant compared to Ca. Accumulibacter [14].
3.1. Carbon Sources
Currently, the research interest has focused on the identification of preferable carbon sources that can be metabolized by Tetrasphaera to release P. The most frequently tested substrates were acetate, glucose, glutamate, glycine and lactate [14]. Moreover, Tetrasphaera exhibit the ability to assimilate a wider range of carbons sources, including amino acids, sugars, higher VFAs under anaerobic conditions [11,38,43]. Other processes have been demonstrated to produce an energy source for aerobic P uptake by Tetrasphaera, including fermentation of amino acids and sugars, the subsequent storage of either amino acids or glycogen anaerobically, and the use of internally stored substrates [11,12,39]. For instance, laboratory experiments with an enriched culture of Tetrasphaera, fed with casein hydrolysate as the sole carbon source, showed that Tetrasphaera can metabolize amino acids and were the main microorganisms responsible for aerobic P removal [39]. Moreover, P uptake and release has been detected in addition to the typical substrates (acetate and glucose) with other organics, including formic acid, propionate, butyric acid, pyruvate, lactate, ethanol, glucose, oleic acid, aspartic acid, glutamic acid, leucine, glycine, thymidine and mixed amino acids [43]. In some cases, Tetrasphaera isolates revealed different preferences for carbon sources in achieving P release. For instance, in the studies by Nguyen et al. [12], the application of glucose and glutamic acid was performed under anaerobic condition with Tetrasphaera isolates, despite substrate utilization no P release was observed. Species T. australiensis, T. japonica and T. elongate responded positively to acetate, whereas propionate in addition to acetate favored T. jenkinsii, T. vanveenii and T. veronensis [33]. In the studies by Marques et al., [39], glucose, aspartate, glutamate and glycine were explored with an enriched Tetrasphaera culture. The results from fluorescence in situ fluorescence in situ hybridization with microradiography FISH-MAR showed that Tetrasphaera could perform P-release anaerobically with each of those carbon sources used solely.
In terms of the intracellular compounds, glycogen has been considered an important energy storage compound in Tetrasphaera [11]. Kong et al. [43] and Nguyen et al. [38] demonstrated that mixed cultures predominated by Tetrasphaera were able to consume glucose anaerobically to promote P uptake aerobically. Other experiments with a pure culture of T. elongate showed the typical PAO phenotype with glucose as a preferable carbon source [11]. Moreover, glycogen production in the anaerobic phase was also observed in another study not only with glucose, but also glutamate and aspartate supplied as an external carbon source [39].
3.2. Metabolic Models of EBPR
In contrary to Ca. Accumulibacter, Tetrasphaera are able to synthesize and store anaerobically wider range of the extracellular compounds. Thus, metabolic pathways of P removal by Tetrasphaera differ from the metabolic models established for the typical PAOs with acetate as the main substrate. Moreover, due to the capability to decompose complex organic compounds, Tetrasphaera form substrate dependencies with other PAOs, including Ca. Accumulibacter.
Initially, EBPR has not been linked with denitrification, e.g., in the Activated Sludge Model No 2 (ASM2) [29]. Using the theory of Mino et al. [44], Smolders et al. [45] developed the anaerobic metabolic model of PAO fed with acetate as a single carbon source. According to that theory, PAO transported acetate across the cell membrane and convert it into acetyl-CoA with the process energy of cleaving poly-P and releasing phosphate from the cell. The parameter represents the ATP required for the transport of 1 C-mmol acetate across the cell membrane [45,46]. Acetyl-CoA was found to be linearly dependent on pH. The origin of the reducing power (i.e., nicotinamide adenine dinucleotide (NADH)), required for PHA synthesis, has been debated by many authors, with Mino et al. [44] supporting its origin from internal glycolysis.
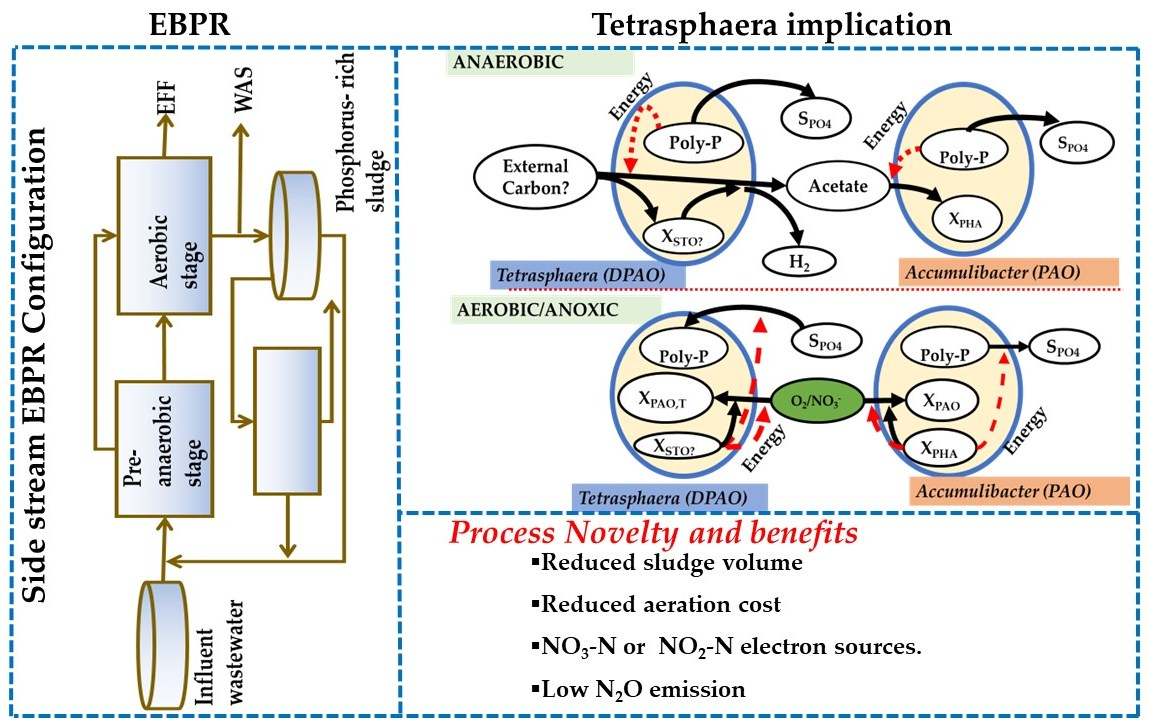
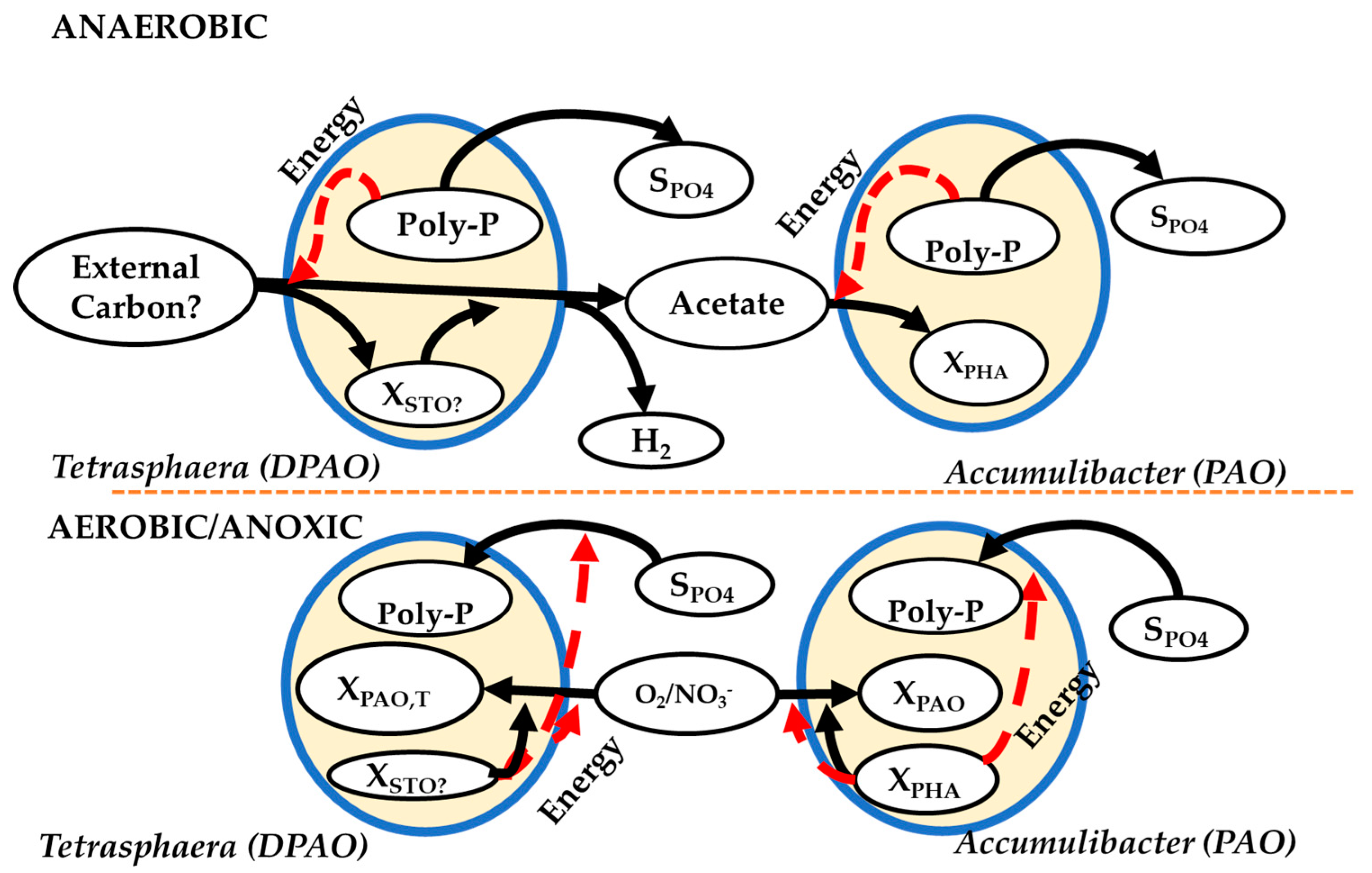
However, Vlekke et al. [47] had earlier demonstrated the capacity of DPAO which led to an update of the ASM2 to the ASM2d [29]. The characterization of the Tetrasphaera metabolism, especially in terms of the capability of complex carbon compounds utilization, provided further insights into the potential pathways within EBPR (Figure 2).
During the anaerobic phase, complex carbon compounds, such as, glucose, are taken up and either stored as glycogen or fermented to acetate. The energy required for glycogen synthesis is supplied by fermentation and/or polyphosphate decomposition to orthophosphate. In the aerobic/anoxic phase, the stored glycogen is degraded, supplying energy for the growth and enhanced P uptake, followed by replenishing the polyphosphate storage.
Various experimental studies have highlighted metabolic models regarding P removal by Tetrasphaera [11,12,39]. The majority of those models have been based on the applied carbon sources and microbial cultures (mixed or pure culture) [49]. Kristiansen et al. [11] proposed a model describing the metabolism of T. elongata as a representative in EBPR with glucose as the substrate. Under anaerobic conditions, Tetrasphaera take up glucose using poly-P as an energy source and glucose can be stored as glycogen. Under the subsequent aerobic conditions, the stored glycogen can be used for the growth and replenishing supplies of poly-P. A model of Nguyen et al. [12] incorporated glycine as a carbon source with no glycogen. The intracellular glycine was accumulated under anaerobic conditions along with small amounts of glutamine, serine, and alanine. These intracellular metabolites could subsequently be used to support the aerobic P uptake. Moreover, Tetrasphaera share some key metabolic pathways with Ca. Accumulibacter, such as tricarboxylic acid cycle (TCA) and poly-P degradation/synthesis. Overall, representatives of Tetrasphaera are extremely versatile, capable of surviving in highly dynamic environments and highly abundant in WWTPs. Therefore, a reliable generic model is still missing, and a consensus has not yet been built on general metabolic models for Tetrasphaera within EBPR.
Metabolic modelling and prediction of bacterial activity can provide useful information for the process optimization and design purposes [27]. For instance, modelling studies by Oehmen et al., [50] enabled to distinguish two subgroups of PAOs, PAO I able to denitrify from nitrate to N2 gas and PAO II performing denitrification with nitrite, as the preferable N source. In addition, the authors found that despite anaerobic kinetic parameters for all PAO (PAO I and PAO II) and GAO subgroups are constant, the actual process rates were strongly dependent on the activity of each specific bacterial group.
With regard to Tetrasphaera, research on their metabolism is still ongoing, particularly on storage products in the anaerobic phase [49]. Tetrasphaera models are considered directly relevant to the models of the EBPR incorporated in ASM and metabolic models [51]. However, an update of the currently available models is necessary as the knowledge of Tetrasphaera biochemical properties and interactions with other bacterial functional groups is continuously increasing.
4. Occurrence of Tetrasphaera in EBPR Systems
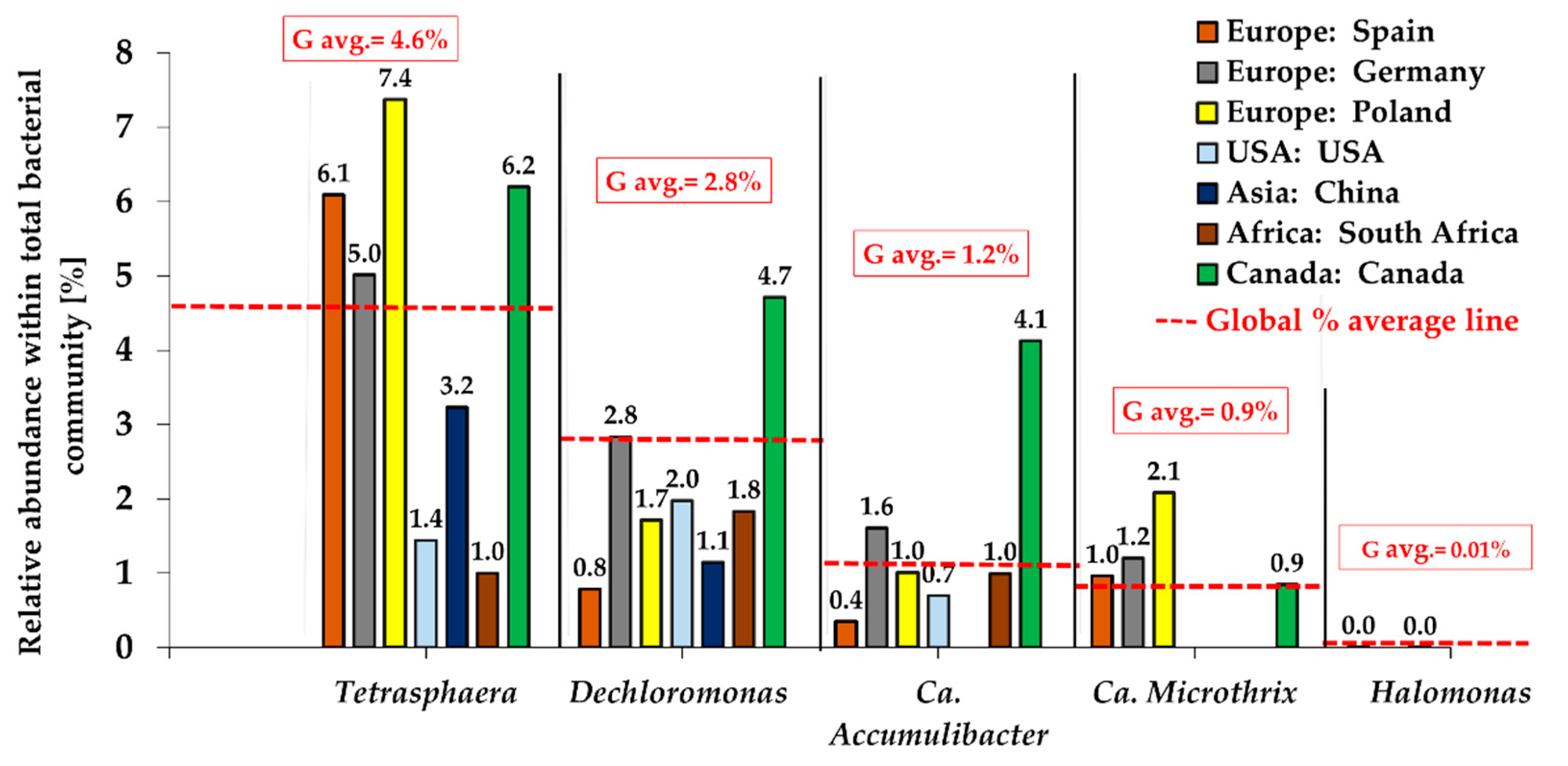
Tetrasphaera abundance has been consistent and demonstrated their predominant role in various studies. Full-scale EBPR systems had higher abundances of Tetrasphaera representatives than Ca. Accumulibacter in many previous studies, where their maximum contribution was estimated at 30% of the total biomass [12,34,38]. Stokholm-Bjerregaard et al. [34] detected Tetrasphaera in large amounts, i.e., up to 35% of the bacterial population, significantly outcompeting Ca. Accumulibacter. Moreover, using 16S rRNA amplicon sequencing and quantitative FISH, Herbst et al. [52] found that Tetrasphaera the most abundant genus in a Danish WWTP, accounting for 30% of the activated sludge community. These findings were confirmed by a survey of 32 full-scale EBPR plants in 12 countries, where higher abundances of Tetrasphaera were reported in most cases [3] by 16S rRNA high-throughput gene sequencing. In that study, the Tetrasphaera abundance in EBPR systems was in the range from 1.3% to 11.9%. According to the recent global survey over the bacterial community structure in EBPR systems across 12 countries from 5 continents (MIDAS project), the highest average abundance of Tetrasphaera was indicated in terms of both DPAOs subpopulation and a general bacterial community. The average Tetrasphaera abundance constituted 4.60%, and prevailed other DPAOs from Dechloromonas (2.84%), Ca. Accumulibacter (1.19%), Ca. Microthrix 0.85% and Halomonas at 0.01% (Figure 3). The predominance of Tetrasphaera in EBPR systems was confirmed also with the application of other than next generation sequencing techniques. For instance, the use of Raman spectroscopy technology for in situ intracellular compound quantification, detected a higher abundance of Tetrasphaera than Ca. Accumulibacter in full-scale WWTPs in Denmark [7]. Singleton et al. [40] showed that representatives of the former Tetrasphaera clade III, in particular the newly established genus Ca. Phosphoribacter, were the dominant PAOs in EPBR systems in Denmark.
However, as suggested by Close et al. [42], the overall abundance and contribution of Tetrasphaera within the total bacterial community is not unequivocally related to the potentially obtained P removal rates. The authors suggested that in case of this issue, the composition of Tetrasphaera clades played more important role.
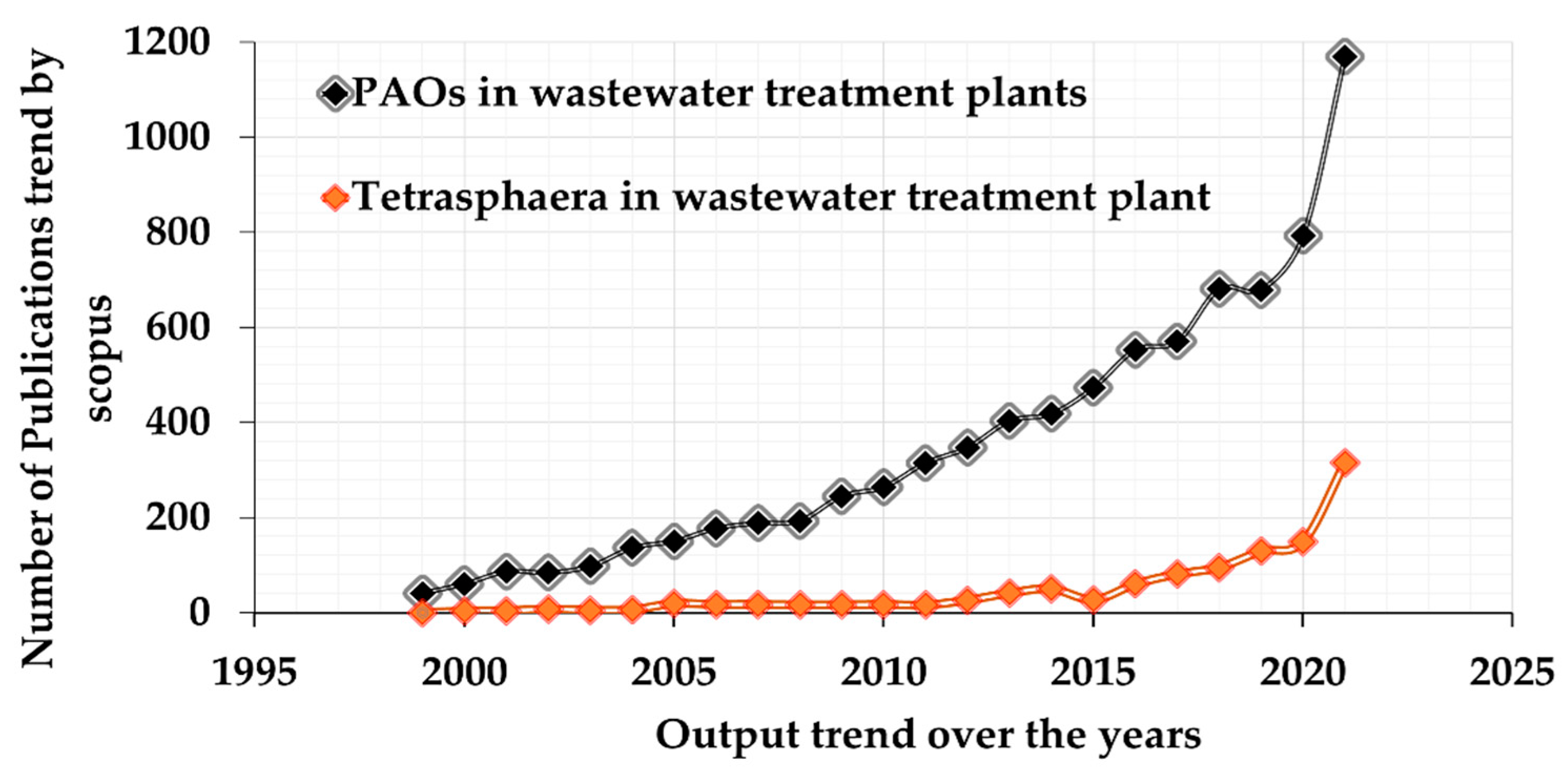
An increased scientific interest in Tetrasphaera has been observed in terms of the number of publications with the key words “Tetrasphaera in wastewater treatment plants” and “PAO in wastewater treatment plants” based on the Scopus database between 1999 to 2021 (Figure 4).
5. Implications of Tetrasphaera on EBPR Configuration and Operation
The identification of PAOs/DPAOs as well their relationship is a crucial step in optimizing P removal efficiency in WWTPs [14]. Maximizing the P removal fraction achieved in anoxic conditions can significantly reduce the operational costs of EBPR systems. Currently, several different process configurations are available, in which both P and N removal are combined [27].
The studies by Meinhold et al. [53] highlighted two different groups of PAO, including aerobic PAOs (APAOs) and denitrifying PAOs (DPAOs). The APAOs can use only DO as an electron acceptor, whereas DPAO can use either DO or nitrate as an electron acceptor. Oehmen et al. [50] postulated that DPAOs have different denitrification capabilities and can be classified based on their reduction abilities towards nitrate or nitrite. Type I Accumulibacter (DPAO I) showed the ability to reduce nitrate, whereas Type II Accumulibacter (DPAO II) had the ability to reduce nitrite only (Table 1). The representatives of Tetrasphaera reflect differential preferences for electron donors and acceptors. While genera T. australiensis, T. japonica, T. elongata show similar metabolic properties to DPAO I with acetate as the key carbon source, whereas the aerobic pathway and preference of propionate are specific features for of T. jenkinsii, T. vanveenii and T. veronensis [14].
According to Mino et al. [54], the denitrification ability of PAO is essential for design of activated sludge system configurations. Two stage anaerobic-anoxic configurations have been considered with the focus of DPAOs activity with EBPR, and their potential role in the anoxic P uptake and available carbon source for denitrification. The main idea of the modern EBPR systems, is to ensure more space to the anoxic zone as compared to the aerobic zone. However, even with these modifications, the DPAOs contributions to denitrification and the total P uptake usually vary in the range of 0–25% and 0–62%, respectively.
This suggests that despite DPAOs capability to grow under both anoxic and aerobic conditions, there is still a lower efficiency in the utilization of stored intracellular compounds, such as PHA or glycogen. It is apparent that DPAOs may have lower performance than APAO in terms of the usage of readily biodegradable COD [48].
The presence of other microorganisms, such as glycogen accumulating organisms (GAO) in EBPR systems, is an additional, important issue that can affect the EBPR performance [55,56]. With relative abundance of 36.0–42.6% [57,58]. Defluviicoccus and 15.3–24.9% of Candidatus Competibacter, are considered as the most dominant GAOs in WWTPs. The predominance of Defluviicoccus is strongly associated with a higher relative ratio of propionate over acetate in the feed. Propionate uptake rates by these bacteria are comparable to Candidatus Accumulibacter and much higher than Candidatus Competibacter [59]. Notably, it remains unclear which factors govern the changes in the known GAO community composition as well competition with PAOs, including solids retention time (SRT) [11,60], C/P ratios [61] and available carbon sources [62], which have been evidenced to be associated with GAO shifts. The challenge of identifying PAO and GAO metabolisms is merely based on the functional genes associated with phenotypes characteristics, such as the ability of cycling of phosphorus, polyhydroxyalkanoates (PHA) and/or glycogen, as intracellular storage materials within bacterial cells [13,63]. On the other hand, investigations by Tu and Schuler [64] led to the conclusion that the role of GAOs in full scale EBPR systems had been overestimated. Most of the previous microbial characterizations had been conducted in laboratory or pilot scale, where a common practice was application of the enormous concentrations of acetate, which favoured the growth of GAO. Similar suppositions have been formulated by Nielsen et al. [4]. The occurrence of GAOs was related to the excess of available carbon, which does not normally occur in typical EBPR system.
Configurations of the EBPR Systems
In general, EBPR systems can be categorized as main- and side stream configurations. The common feature of conventional side-stream configurations was sole treatment of return sludge anaerobically combined with chemical precipitation, whereas in mainstream configurations all mixed liquors flow through a sequence of anaerobic, anoxic and/or aerobic conditions for P removal [1].
Modern mainstream EBPR systems by design, are meant to avail conditions that sustain the parallel processes of N and P removal. Achieving optimal operating conditions for the biological processes, such as phosphorus release and uptake, nitrification and denitrification, requires consideration of specific environmental conditions within anaerobic, aerobic and anoxic zones. Due to the limited resources and stricter operational regulations, continuous modernization and revaluation of the BNR configurations has been observed [65].
A successful EBPR process is dependent on the presence of readily biodegradable organic carbon and phosphorus, anaerobic zone prior to aerobic zone and sufficient amount of nutrients since it relies on growth and selection of PAOs which are capable of storing orthophosphate in excess.
Earlier configurations achieved P removal with >90% efficiency [19,20,66]. By changing the sequential steps in the specific configuration, a low P removal efficiency was achieved, leading to an understanding of necessary conditions of EBPR [2]. The availability of COD favours heterotrophic activities (PAO, denitrifiers) under anaerobic/anoxic conditions, whereas nitrification takes place in the aerobic zone. P removal depends on several conditions that are essential for microbial metabolism of PAO [67]. However, simultaneous N and P removal are not straightforward as the addition of the anaerobic zone, which favours the PAO growth only [68]. In most systems, nitrification and denitrification may cause detrimental impact on EBPR due to the presence of nitrite and nitrate in the external recycle stream which enters the anaerobic zone, leading to a process failure. The presence of electron acceptors, such as nitrate and nitrite, under anaerobic conditions potentially sparks heterotrophic denitrifying bacteria growth and outcompete PAO [69]. Moreover, as indicated by Conidi et al. [70], the most common cause of instability in EBPR systems is the underestimated size of anaerobic zones, often less than 10% by mass of solids, whereas 15–25% is recommended for a stable operation.
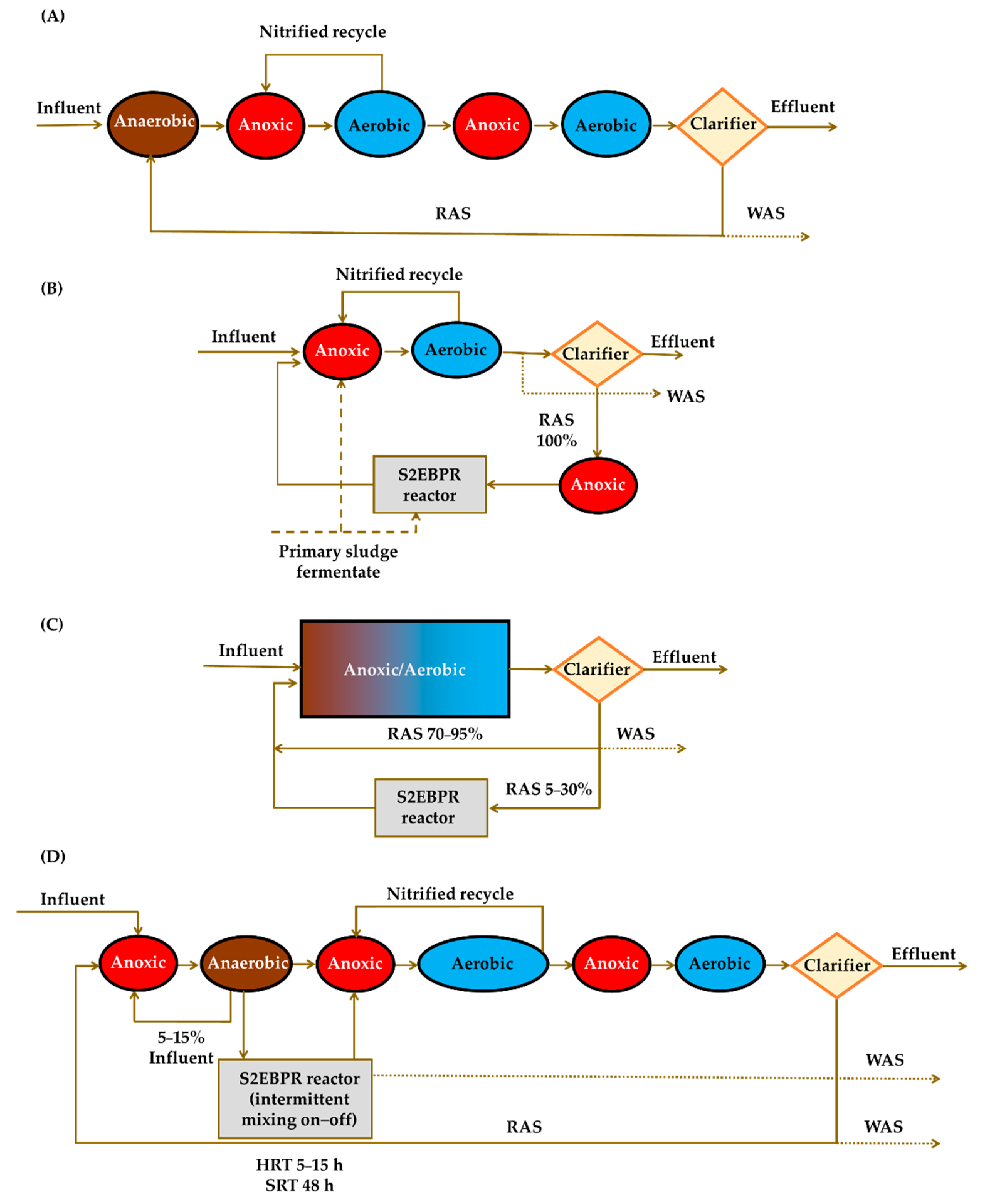
Currently, the main direction towards increasing EBPR stability is the implementation of the novel sidestream EBPR (S2EBPR) configurations. With respect to the existing bioreactor configurations, providing a side stream fermentation zone (S2EBPR reactor) of recirculated activated sludge (RAS) or mixed liquor fermentation have emerged as a perspective solution to solve this issue. The main potential advantages offered by these solutions are increased anaerobic mass fraction and potential of the selective GAO suppression [71].
Due to P metabolism combined with the capability of fermenting a wide range of organic compounds and denitrification with simultaneous P uptake, Tetrasphaera are considered the key players of the S2EBPR systems. However, it should be noted that currently available data from the S2EBPR operations are limited, basically due to a narrow range of the operational conditions studied to date. As mentioned by Dold and Conidi [72], more extensive data are highly required to develop models specifically dedicated for S2EBPR systems.
6. Factors Affecting the Occurrence and Activity of Tetrasphaera and Other DPAOs in EBPR Systems
The recognition of dependencies between activity and occurrence of Tetrasphaera in relation to the changes of the environmental and operational conditions (pH, temperature, influent wastewater characteristics and DO or nitrate presence), as well interactions with other functional bacterial groups (especially GAO) are crucial for the development and optimization of EBPR [60,74,75,76]. However, in practice, it is not possible to achieve stable and fully controlled operational parameters, such as temperature and pH [77]. In general, other factors, such as the availability of the readily biodegradable organic compounds, P load, cation (specifically K+ and M2+) concentration, pH, and the food-to-microorganism ratio should be considered [16].
6.1. Temperature
Temperature is known to affect the operational efficiency of EBPR systems [78]. A study on psychrophilic nature of PAOs indicated their functionality at the temperatures around 20 °C or lower [4]. The increasing temperatures between 5 and 24 °C, improve the process efficiency. At elevated temperatures, the mesophilic GAOs seem to outcompete PAOs in terms of the carbon sources uptake, so the process efficiency becomes problematic at tropical temperatures of (25–32 °C) [4,27,31,57,77].
Other studies also draw attention to the deterioration of the efficiency of the EBPR process at high temperatures (above 20 °C) [51,79,80,81]. This indicates that the threshold temperature for tropical EBPR failure ranges from 35 to 40 °C, and above a significant deterioration in the PAO activity is observed. In addition, Panswad et al. [79] found that under elevated temperature, the competition between GAOs and PAOs is believed to enhance in terms of uptake of the available VFAs under anaerobic conditions. There is a substantial effect of temperature on lowering the relative abundance of PAOs with the increasing operating temperature from 20 to 35 °C. According to Whang and Park [81] and Lopez -Vazquez et al., [51], GAOs had faster VFA uptake rates at higher temperatures, leading to their excessive proliferation in the system. Ca. Accumulibacter and Tetrasphaera reflect higher abundances in the systems operated below 20 °C [27,51,82], whereas Ca. Competibacter become more numerous at higher temperatures. However, studies by Qiu et al. [83] and Wang et al. [84] showed the possibility of achieving highly efficient EBPR at the temperatures ranging from 28 to 32 °C.
The effect of temperature on the occurrence of Tetrasphaera was studied by Liu et al. [14] with the correlation analysis of experimental data from over 60 references. The results of the analysis revealed that the highest abundances of Tetrasphaera were mainly recorded in the temperature range from 10 to 20 °C. In contrast, in the countries with the warm climate and temperatures ranging from 20 to 30 °C, such as Australia or Spain, the abundance of Tetrasphaera was significantly lower. This finding is not fully consistent with the characteristics of pure Tetrasphaera cultures, where the temperature of 25 °C was considered suboptimal [32,33]. The lowered occurrence of Tetrasphaera at elevated temperatures, should therefore be explained by outcompetition from other microbial groups, in particular GAOs.
6.2. Influent Wastewater Characteristics
Influent wastewater characteristics have the potential to affect the abundance of Tetrasphaera and alter the metabolic pathways. The availability of preferred carbon sources and electron acceptors are essential to promote P-removal physiology of Tetrasphaera. As pointed out by Kong et al. [43], industrial WWTPs reflect usually higher abundances of Tetrasphaera compared to the systems treating mainly domestic wastewater. Mielczarek et al. [85] studied the correlation of abundance of Tetrasphaera and wastewater characteristics, process design, and operation, with the data of over 3 years from 28 Danish WWTPs. Their results showed weak correlations with the increased amount of industrial wastewater in the influent. As reported by Lopez-Vazquez et al., [51], the diversity of carbon compounds in the influent (the ratios of acetate to propionate of 75–25% and 50–50%) provided more favorable conditions for PAO than GAO, despite the elevated temperature (30 °C) in contrary to the experimental trials when single carbon source was used.
The capability to metabolize and internally store many carbon compounds provides Tetrasphaera with enormous adaptability to the dynamic conditions of substrate availability in WWTPs [52]. Such physiological plasticity may gain Tetrasphaera advantage over Ca. Accumulibacter, which is highly dependent on the acetate availability under anaerobic conditions [56].
6.3. pH
It is considered that, pH set point and its control are critical for enhancing typical PAO and Tetrasphaera activity in P removal [86,87].
Experimental results showed that P removal linearly increased with initial pH increasing from 6.6 to 7.8, but slightly decreased when initial pH increased from 7.8 to 8.2. Initial optimal pH of 7.8 favored P removal offering approximately 1.7 times compared to the pH at 6.6. Additionally, the modeling studies have showed that the biomass cultured at initial pH 7.8 contained elevated abundances of PAOs [88]. A recent study by Kang et al. [89] showed that the significant effect of pH control at a pH greater than or equal to 7.5 improved the P removal efficiency from 90.8% to 99.6%, whereas a pH below 7.0 ensured only about 63.1%.
The results of technological studies cover expectations from the characteristics of cultivable Tetrasphaera strains, for which the pH range for growth is 6.0–9.0 with the optimum at 7.0 [32]. Filipe et al. [89] observed the improved phosphate removal in the studied EBPR system when the pH was allowed to increase to a maximum of 7.5.
Another critical issue related to the pH influence is the competition between PAOs and GAOs, where pH is considered to affect anaerobic uptake kinetics of organic carbon. Several studies confirmed that pH in the range of 7–8.5 leads to an increased abundance of PAOs, whereas GAOs were inhibited or their proliferation rate was reduced under those conditions [27,51,56]. There is now strong evidence that the stability of EBPR systems can be improved by increasing the pH in the anaerobic zone. This allows for creating conditions under which PAOs are able to uptake acetate faster than GAOs, which leads to the positive shift in the composition of activated sludge biomass.
On the other hand, researchers seek clarity on how alkaline conditions can inhibit GAO proliferation and induce PAO activity [10]. The main explanation of this observation is that metabolism of the bacterial cells and enzymes activity is highly dependent on pH, thus under unfavourable pH more energy is needed for the substrate uptake and P release. It is postulated that carbon source uptake and phosphorus release/uptake are provided by proton motive force (PMF), strongly dependent on the extracellular pH and related to cation release [45,90].
In the study of Schuler and Jenkins [56], it was found that pH significantly affects the anaerobic phase of EBPR performed by Ca. Accumulibacter. However, different correlations were observed between pH and acetate uptake rates as well as between pH and anaerobic P release rates. For acetate uptake rates, a continuous positive correlation was found with the increasing pH, but the increase in acetate uptake slowed down above pH 7.2. On the contrary, in the case of P release rate, a plateau was gained at pH 7.2 and above. This suggests that at elevated pH, the energetic cost of acetate transport could not be compensated by phosphate lysis. Based on the behaviour of cations, correlated with pH during anaerobic carbon uptake, Saunders [91] postulated a secondary transport model as the dominant mechanism of EBPR by PAOs. In that model, acetate was co-transported with cations (typically H+), which started to accumulate inside the bacterial cell and decrease PMF, thus slowing down the P release rate.
A recent study by Belka [92] showed that similar patterns were observed for pH-dependent anaerobic P release by Tetrasphaera as in previous studies with Ca. Accumulibacter. However, the anaerobic carbon uptake by Tetrasphaera was not correlated with pH and did not slow down at pH higher than 7.2. Due to more versatile metabolic properties (i.e., capability of accumulation of free organic carbon solutes and anaerobic amino acids uptake), Tetrasphaera could gain an important advantage over Ca. Accumulibacter under highly dynamic feeding and pH conditions.
6.4. Presence of DO and Nitrate
The presence of nitrate and DO in the anaerobic zone are considered inhibitors of the PAO activity. When the anaerobic zone is free of nitrate and DO, the PAO activity is favoured over any other rapidly growing heterotrophs that may use DO or nitrate as electron acceptors. The DO concentration of DO has been found to strongly affect EBPR performance and PAO dominance. Izadi et al. [10] achieved efficient P-removal at low DO concentrations (in the range of 0.5 or 0.8 mg/L), which was attributed to the favoured growth of PAOs compared to GAOs. The presence of nitrate and/or DO reduces the amount of VFAs available for the PAO activity, while hampering P removal [93]. At low DO levels, PAOs have an advantage over Ca. Competibacter due to a higher DO affinity. Excessive aeration may induce instable P-removal as a result of GAO competition. Chen et al. [94] found that maintaining the DO level at 0.5 mgO2/L promoted a higher efficiency EBPR.
Regardless of the dynamic conditions associated with the transition between anaerobic, anoxic and aerobic phases, Tetrasphaera representatives were found to be the dominant genus in most of the European WWTPs [34]. Herbst et al. [52] provided important insights into the characteristics of metabolic traits, which favored Tetrasphaera over most other functional bacterial groups under such dynamic conditions. By the use of label-free quantitative proteomics and nuclear magnetic resonance (NMR), the physiology of T. longate str. LP2 isolate was verified under dynamic shifts between anaerobic and aerobic conditions. Unlike the reference bacterial strain of Escherichia coli, Tetrasphaera reflected a stable proteome profile during transitions, which suggested that specific metabolic pathways for anaerobic processes remained induced under aerobic conditions. This metabolic property ensures a physiological advantage of Tetrasphaera over more specialized microbial groups. Moreover, it is suggested that by enlarging the anaerobic zone in EBPR systems, with a reduced oxidation-reduction potential (ORP), Tetrasphaera may gain a completive advantage over other heterotrophic bacteria [14].
7. Conclusions
The implementation of P-removal technologies based on application of the anaerobic–anoxic conditions has been recognized as the perspective approach for the energy efficient wastewater treatment. In such systems, the development of Tetrasphaera population and control of its activity may become a critical step. On the contrary, EBPR systems dominated by Ca. Accumulibacter, which are strongly dependent on acetate availability, are considered more susceptible to failure under dynamic conditions in WWTPs.
The development of Tetrasphaera-based EPBR systems is strongly determined by understanding the operational conditions, available carbon source and the presence of electron acceptors in the anaerobic zone. In addition, interactions with other functional bacterial groups (especially GAOs and other PAOs) have to be understood for further EBPR development and optimization.
The availability of complex carbon sources in the influent, moderate temperature (10–20 °C), elevated pH (>7.5) and increased sizes of the anaerobic zones are currently recognized as the possible main factors favouring Tetrasphaera abundance over typical PAOs (Ca. Accumulibacter). The conclusiveness of operational conditions such as pH and temperature specifically to promote the growth of Tetrasphaera over Accumulibacter is not definite at present. The known advantages exhibited by Tetrasphaera therefore, are attributable to versatile metabolism in line with their denitrification, fermentation and polyphosphate accumulation ability. Due to these versatile metabolic properties, Tetrasphaera are considered the key microorganisms for the novel S2EBPR systems.
However, there are still issues that need to be resolved in the future, including a phylogeny of Tetrasphaera genus, more complex characterization of metabolic traits of individual Tetrasphaera species, as well as kinetic studies based on Tetrasphaera-enriched cultures from full-scale systems.
Author Contributions
Conceptualization, J.O., J.M. and P.K.; J.O.; resources, J.O.; data curation, J.O.; writing—original draft preparation, J.O., writing—review and editing, P.K.; visualization, P.K. and J.O.; supervision, J.M. and P.K.; project administration, J.O. and P.K.; funding acquisition, P.K. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Polish National Science Center under project no. UMO-2019/03/X/NZ9/01257663.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Metcalf and Eddy Inc. Wastewater Engineering Treatment and Reuse, 5th ed.; McGraw-Hill: New York, NY, USA, 2014. [Google Scholar]
- Barnard, J.L.; Dunlap, P.; Steichen, M. Rethinking the Mechanisms of Biological Phosphorus Removal. Water Environ. Res. 2017, 89, 2043–2054. [Google Scholar] [CrossRef]
- Nielsen, P.H.; Saunders, A.M.; Hansen, A.A.; Larsen, P.; Nielsen, J.L. Microbial communities involved in enhanced biological phosphorus removal from wastewater—A model system in environmental biotechnology. Curr. Opin. Biotechnol. 2012, 23, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.H.; McIlroy, S.J.; Albertsen, M.; Nierychlo, M. Re-evaluating the microbiology of the enhanced biological phosphorus removal process. Curr. Opin. Biotechnol. 2019, 57, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Rincón, F.; Lopez-Vazquez, C.; Welles, L.; van Loosdrecht, M.; Brdjanovic, D. Cooperation between Candidatus Competibacter and Candidatus Accumulibacter clade I, in denitrification and phosphate removal processes. Water Res. 2017, 120, 156–164. [Google Scholar] [CrossRef]
- Marques, R.; Ribera-Guardia, A.; Santos, J.; Carvalho, G.; Reis, M.A.; Pijuan, M.; Oehmen, A. Denitrifying capabilities of Tetrasphaera and their contribution towards nitrous oxide production in enhanced biological phosphorus removal processes. Water Res. 2018, 137, 262–272. [Google Scholar] [CrossRef]
- Fernando, E.Y.; McIlroy, S.J.; Nierychlo, M.; Herbst, F.-A.; Petriglieri, F.; Schmid, M.C.; Wagner, M.; Nielsen, J.L.; Nielsen, P.H. Resolving the individual contribution of key microbial populations to enhanced biological phosphorus removal with Raman–FISH. ISME J. 2019, 13, 1933–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Kinyua, M.N. Identification and classification of the Tetrasphaera genus in enhanced biological phosphorus removal process: A review. Rev. Environ. Sci. Technol. 2020, 19, 699–715. [Google Scholar] [CrossRef]
- Cydzik-Kwiatkowska, A.; Zielińska, M. Bacterial communities in full-scale wastewater treatment systems. World J. Microbiol. Biotechnol. 2016, 32, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izadi, P.; Eldyasti, A. Understanding microbial shift of Enhanced Biological Phosphorus Removal process (EBPR) under different Dissolved Oxygen (DO) concentrations and Hydraulic Retention Time (HRTs). Biochem. Eng. J. 2021, 166, 107833. [Google Scholar] [CrossRef]
- Kristiansen, R.; Nguyen, H.T.T.; Saunders, A.M.; Nielsen, J.L.; Wimmer, R.; Le, V.Q.; McIlroy, S.J.; Petrovski, S.; Seviour, R.J.; Calteau, A.; et al. A metabolic model for members of the genus Tetrasphaera involved in enhanced biological phosphorus removal. ISME J. 2013, 7, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.T.; Kristiansen, R.; Vestergaard, M.; Wimmer, R.; Nielsen, P.H. Intracellular Accumulation of Glycine in Polyphosphate-Accumulating Organisms in Activated Sludge, a Novel Storage Mechanism under Dynamic Anaerobic-Aerobic Conditions. Appl. Environ. Microbiol. 2015, 81, 4809–4818. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Guanglei, Q.; Zuniga-Montanez, R.; Williams, R.B.; Wuertz, S. Recent advances in understanding the ecophysiology of enhanced biological phosphorus removal. Curr. Opin. Biotechnol. 2021, 67, 166–174. [Google Scholar] [CrossRef]
- Liu, R.; Hao, X.; Chen, Q.; Li, J. Research advances of Tetrasphaera in enhanced biological phosphorus removal: A review. Water Res. 2019, 166, 115003. [Google Scholar] [CrossRef] [PubMed]
- Welles, L.; Tian, W.; Saad, S.; Abbas, B.; Lopez-Vazquez, C.; Hooijmans, C.; van Loosdrecht, M.; Brdjanovic, D. Accumulibacter clades Type I and II performing kinetically different glycogen-accumulating organisms metabolisms for anaerobic substrate uptake. Water Res. 2015, 83, 354–366. [Google Scholar] [CrossRef]
- Wisniewski, K.; Kowalski, M.; Makinia, J. Modeling nitrous oxide production by a denitrifying-enhanced biologically phosphorus removing (EBPR) activated sludge in the presence of different carbon sources and electron acceptors. Water Res. 2018, 142, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Levin, G.V.; Shapiro, J. Metabolic Uptake of Phosphorus by Wastewater Organisms. Water Pollut. Control. Fed. 1965, 37, 800–821. [Google Scholar]
- Srinath, E.G.; Sastry, C.A.; Pillai, S.C. Rapid removal of phosphorus from sewage by activated sludge. Experientia 1959, 15, 339–340. [Google Scholar] [CrossRef]
- Barnard, J.L. Cut P and N without chemicals. Water Wastes Eng. Part 1 1974, 11, 33–36. [Google Scholar]
- Barnard, J.L. Cut P and N without chemicals. Water Wastes Eng. Part 2 1974, 11, 41–43. [Google Scholar]
- Barnard, J.L. Nutrient removal in biological systems. Water Pollut. Control 1975, 74, 143–154. [Google Scholar]
- Levin, G.V.; Topol, G.J.; Tarnay, A.G. Operation of full-scale biological phosphorus removal plant. Water Pollut. Control Fed. 1975, 47, 577–590. [Google Scholar]
- Fuhs, G.W.; Chen, M. Microbiological basis of phosphate removal in the activated sludge process for the treatment of wastewater. Microb. Ecol. 1975, 2, 119–138. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Loy, A.; Nogueira, R.; Purkhold, U.; Valeeva, A.V.; Daims, H. Microbial community composition and function in wastewater treatment plants. Antonie Leeuwenhoek 2002, 81, 665–680. [Google Scholar] [CrossRef]
- Bond, P.L.; Hugenholtz, P.; Keller, J.; Blackall, L.L. Bacterial community structures of phosphate-removing and non-phosphate-removing activated sludges from sequencing batch reactors. Appl. Environ. Microbiol. 1995, 61, 1910–1916. [Google Scholar] [CrossRef] [Green Version]
- Bond, P.L.; Keller, J.; Blackall, L.L. Characterisation of enhanced biological phosphorus removal activated sludges with dissimilar phosphorus removal performances. Water Sci. Technol. 1998, 37, 567–571. [Google Scholar] [CrossRef]
- Oehmen, A.; Lemos, P.C.; Carvalho, G.; Yuan, Z.; Keller, J.; Blackall, L.L.; Reis, M.A. Advances in enhanced biological phosphorus removal: From micro to macro scale. Water Res. 2007, 41, 2271–2300. [Google Scholar] [CrossRef] [PubMed]
- McIlroy, S.J.; Saunders, A.; Albertsen, M.; Nierychlo, M.; McIlroy, B.; Hansen, A.A.; Karst, S.M.; Nielsen, J.L.; Nielsen, P.H. MiDAS: The field guide to the microbes of activated sludge. Database 2015, 2015, bav062. [Google Scholar] [CrossRef]
- Henze, M.; Gujer, W.; Mino, T.; van Loosdrecht, M. Activated Sludge Models, ASM1, ASM2, ASM2d and ASM3. Scientific and Technical Report (Volume 5); IWA Publishing: London, UK, 2000. [Google Scholar] [CrossRef]
- Jenkins, D.; Wanner, J. Activated Sludge—100 Years and Counting; IWA Publishing: Glasgow, UK, 2014. [Google Scholar] [CrossRef]
- Bertanza, G.; Menoni, L.; Capoferri, G.U.; Pedrazzani, R. Promoting biological phosphorus removal in a full scale pre-denitrification wastewater treatment plant. J. Environ. Manag. 2020, 254, 109803. [Google Scholar] [CrossRef]
- Hanada, S.; Liu, W.-T.; Shintani, T.; Kamagata, Y.; Nakamura, K. Tetrasphaera elongata sp. nov., a polyphosphate-accumulating bacterium isolated from activated sludge. Int. J. Syst. Evol. Microbiol. 2002, 52, 883–887. [Google Scholar] [CrossRef]
- Maszenan, A.; Seviour, R.; Patel, B.; Schumann, P.; Burghardt, J.; Tokiwa, Y.; Stratton, H. Three isolates of novel polyphosphate-accumulating gram-positive cocci, obtained from activated sludge, belong to a new genus, Tetrasphaera gen. nov., and description of two new species, Tetrasphaera japonica sp. nov. and Tetrasphaera australiensis sp. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 593–603. [Google Scholar] [CrossRef] [Green Version]
- Stokholm-Bjerregaard, M.; McIlroy, S.J.; Nierychlo, M.; Karst, S.M.; Albertsen, M.; Nielsen, P.H. A Critical Assessment of the Microorganisms Proposed to be Important to Enhanced Biological Phosphorus Removal in Full-Scale Wastewater Treatment Systems. Front. Microbiol. 2017, 8, 718. [Google Scholar] [CrossRef] [Green Version]
- Seviour, R.J.; Mino, T.; Onuki, M. The microbiology of biological phosphorus removal in activated sludge systems. FEMS Microbiol. Rev. 2003, 27, 99–127. [Google Scholar] [CrossRef] [Green Version]
- Seviour, R.J.; McIlroy, S.J. The microbiology of phosphorus removal in activated sludge processes-the current state of play. J. Microbiol. 2008, 46, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Petriglieri, F.; Singleton, C.; Peces, M.; Petersen, J.F.; Nierychlo, M.; Nielsen, P.H. “Candidatus Dechloromonas phosphatis” and “Candidatus Dechloromonas phosphovora”, two novel polyphosphate accumulating organisms abundant in wastewater treatment systems. BioRxiv 2020. [Google Scholar] [CrossRef]
- Nguyen, H.T.T.; Le-Quy, V.; Hansen, A.A.; Nielsen, J.L.; Nielsen, P.H. High diversity and abundance of putative polyphosphate-accumulating Tetrasphaera-related bacteria in activated sludge systems. FEMS Microbiol. Ecol. 2011, 76, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Marques, R.; Santos, J.; Nguyen, H.; Carvalho, G.; Noronha, J.; Nielsen, P.H.; Reis, M.A.; Oehmen, A. Metabolism and ecological niche of Tetrasphaera and Ca. Accumulibacter in enhanced biological phosphorus removal. Water Res. 2017, 122, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Singleton, C.M.; Petriglieri, F.; Wasmund, K.; Nierychlo, M.; Kondrotaite, Z.; Petersen, J.F.; Peces, M.; Dueholm, M.S.; Wagner, M.; Nielsen, P.H. The novel genus, ‘Candidatus Phosphoribacter’, previously identified as Tetrasphaera, is the dominant polyphosphate accumulating lineage in EBPR wastewater treatment plants worldwide. ISME J. 2022, 16, 1605–1616. [Google Scholar] [CrossRef]
- Nouioui, I.; Carro, L.; García-López, M.; Meier-Kolthoff, J.P.; Woyke, T.; Kyrpides, N.C.; Pukall, R.; Klenk, H.-P.; Goodfellow, M.; Göker, M. Genome-Based Taxonomic Classification of the Phylum Actinobacteria. Front. Microbiol. 2018, 9, 2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Close, K.; Marques, R.; Carvalho, V.C.; Freitas, E.B.; Reis, M.A.; Carvalho, G.; Oehmen, A. The storage compounds associated with Tetrasphaera PAO metabolism and the relationship between diversity and P removal. Water Res. 2021, 204, 117621. [Google Scholar] [CrossRef]
- Kong, Y.; Nielsen, J.L.; Nielsen, P.H. Identity and Ecophysiology of Uncultured Actinobacterial Polyphosphate-Accumulating Organisms in Full-Scale Enhanced Biological Phosphorus Removal Plants. Appl. Environ. Microbiol. 2005, 71, 4076–4085. [Google Scholar] [CrossRef] [Green Version]
- Mino, T.; Arun, V.; Tsuzuki, Y.; Matsuo, T. Effect of phosphorus accumulation on acetate metabolism in the biological phosphorus removal process. In Biological Phosphate Removal from Wastewaters, Proceedings of an IAWPRC Specialized Conference held in Rome, Italy, 28–30 September 1987; Ramadori, R., Ed.; Elsevier: Amsterdam, The Netherlands, 1987; pp. 27–38. [Google Scholar] [CrossRef]
- Smolders, G.J.F.; van der Meij, J.; van Loosdrecht, M.C.M.; Heijnen, J.J. Model of the anaerobic metabolism of the biological phosphorus removal process: Stoichiometry and pH influence. Biotechnol. Bioeng. 1994, 43, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Filipe, C.D.M.; Daigger, G.T.; Grady, C.P.L. A metabolic model for acetate uptake under anaerobic conditions by glycogen accumulating organisms: Stoichiometry, kinetics, and the effect of pH. Biotechnol. Bioeng. 2001, 76, 17–31. [Google Scholar] [CrossRef]
- Vlekke, G.; Comeau, Y.; Oldham, W. Biological phosphate removal from wastewater with oxygen or nitrate in sequencing batch reactors. Environ. Technol. Lett. 1988, 9, 791–796. [Google Scholar] [CrossRef]
- Mąkinia, J.; Zaborowska, E. Mathematical Modelling and Computer Simulation of Activated Sludge Systems; International Water Association Publishing: London, UK, 2020. [Google Scholar] [CrossRef]
- Izadi, P.; Eldyasti, A. A review of biochemical diversity and metabolic modeling of EBPR process under specific environmental conditions and carbon source availability. J. Environ. Manag. 2021, 288, 112362. [Google Scholar] [CrossRef]
- Oehmen, A.; Carvalho, G.; Lopez-Vazquez, C.; van Loosdrecht, M.; Reis, M. Incorporating microbial ecology into the metabolic modelling of polyphosphate accumulating organisms and glycogen accumulating organisms. Water Res. 2010, 44, 4992–5004. [Google Scholar] [CrossRef]
- Lopez-Vazquez, C.M.; Oehmen, A.; Hooijmans, C.M.; Brdjanovic, D.; Gijzen, H.J.; Yuan, Z.; van Loosdrecht, M.C. Modeling the PAO–GAO competition: Effects of carbon source, pH and temperature. Water Res. 2009, 43, 450–462. [Google Scholar] [CrossRef]
- Herbst, F.-A.; Dueholm, M.S.; Wimmer, R.; Nielsen, P.H. The Proteome of Tetrasphaera elongata is adapted to Changing Conditions in Wastewater Treatment Plants. Proteomes 2019, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Meinhold, J.; Filipe, C.D.; Daigger, G.T.; Isaacs, S.H. Characterization of the denitrifying fraction of phosphate accumulating organisms in biological phosphate removal. Water Sci. Technol. 1999, 39, 31–42. [Google Scholar] [CrossRef]
- Mino, T.; van Loosdrecht, M.; Heijnen, J. Microbiology and biochemistry of the enhanced biological phosphate removal process. Water Res. 1998, 32, 3193–3207. [Google Scholar] [CrossRef]
- Meng, Q.; Zeng, W.; Wang, B.; Fan, Z.; Peng, Y. New insights in the competition of polyphosphate-accumulating organisms and glycogen-accumulating organisms under glycogen accumulating metabolism with trace Poly-P using flow cytometry. Chem. Eng. J. 2020, 385, 123915. [Google Scholar] [CrossRef]
- Schuler, A.J.; Jenkins, D. Enhanced biological phosphorus removal from wastewater by biomass with different phosphorus contents, Part I: Experimental results and comparison with metabolic models. Water Environ. Res. 2003, 75, 485–498. [Google Scholar] [CrossRef]
- Winkler, M.-K.; Bassin, J.; Kleerebezem, R.; de Bruin, L.; Brand, T.V.D.; van Loosdrecht, M. Selective sludge removal in a segregated aerobic granular biomass system as a strategy to control PAO–GAO competition at high temperatures. Water Res. 2011, 45, 3291–3299. [Google Scholar] [CrossRef]
- Wong, M.-T.; Tan, F.M.; Ng, W.J.; Liu, W.-T. Identification and occurrence of tetrad-forming Alphaproteobacteria in anaerobic–aerobic activated sludge processes. Microbiology 2004, 150, 3741–3748. [Google Scholar] [CrossRef] [Green Version]
- Carvalheira, M.; Oehmen, A.; Carvalho, G.; Reis, M.A. The effect of substrate competition on the metabolism of polyphosphate accumulating organisms (PAOs). Water Res. 2014, 64, 149–159. [Google Scholar] [CrossRef]
- Onnis-Hayden, A.; Majed, N.; Li, Y.; Rahman, S.M.; Drury, D.; Risso, L.; Gu, A.Z. Impact of solid residence time (SRT) on functionally relevant microbial populations and performance in full-scale enhanced biological phosphorus removal (EBPR) systems. Water Environ. Res. 2020, 92, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Majed, N.; Gu, A.Z. Phenotypic dynamics in polyphosphate and glycogen accumulating organisms in response to varying influent C/P ratios in EBPR systems. Sci. Total Environ. 2020, 743, 140603. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Zhou, Y. Enhanced biological phosphorus removal with different carbon sources. Appl. Microbiol. Biotechnol. 2016, 100, 4735–4745. [Google Scholar] [CrossRef]
- Oyserman, B.; Noguera, D.R.; del Rio, T.G.; Tringe, S.G.; McMahon, K.D. Metatranscriptomic insights on gene expression and regulatory controls in Candidatus Accumulibacter phosphatis. ISME J. 2016, 10, 810–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, Y.; Schuler, A.J. Low Acetate Concentrations Favor Polyphosphate-Accumulating Organisms over Glycogen-Accumulating Organisms in Enhanced Biological Phosphorus Removal from Wastewater. Environ. Sci. Technol. 2013, 47, 3816–3824. [Google Scholar] [CrossRef]
- Izadi, P.; Eldyasti, A. Design, operation and technology configurations for enhanced biological phosphorus removal (EBPR) process: A review. Rev. Environ. Sci. Technol. 2020, 19, 561–593. [Google Scholar] [CrossRef]
- Barnard, J.L. A review of biological phosphorus removal in the activated sludge process. Water SA 1976, 2, 136–144. [Google Scholar]
- Alasino, N.; Mussati, M.C.; Scenna, N.; Aguirre, P. Combined nitrogen and phosphorus removal. Model-based process optimization. Comput. Aided Chem. Eng. 2008, 25, 163–168. [Google Scholar] [CrossRef]
- Guerrero, J.; Guisasola, A.; Baeza, J.A. A novel control strategy for efficient biological phosphorus removal with carbon-limited wastewaters. Water Sci. Technol. 2014, 70, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, J.; Flores-Alsina, X.; Guisasola, A.; Baeza, J.A.; Gernaey, K.V. Effect of nitrite, limited reactive settler and plant design configuration on the predicted performance of simultaneous C/N/P removal WWTPs. Bioresour. Technol. 2013, 136, 680–688. [Google Scholar] [CrossRef] [Green Version]
- Conidi, D.; Andalib, M.; Andres, C.; Bye, C.; Umble, A.; Dold, P. Modeling quaternary ammonium compound inhibition of biological nutrient removal activated sludge. Water Sci. Technol. 2018, 79, 41–50. [Google Scholar] [CrossRef]
- Onnis-Hayden, A.; Srinivasan, V.; Tooker, N.B.; Li, G.; Wang, D.; Barnard, J.L.; Bott, C.; Dombrowski, P.; Schauer, P.; Menniti, A.; et al. Survey of full-scale sidestream enhanced biological phosphorus removal (S2EBPR) systems and comparison with conventional EBPRs in North America: Process stability, kinetics, and microbial populations. Water Environ. Res. 2020, 92, 403–417. [Google Scholar] [CrossRef]
- Dold, P.; Conidi, D. Achieving Enhanced Biological P Removal: Have we forgotten how to design a bioP plant? In Proceedings of the 92nd Annual Water Environment Federation Technical Exhibition and Conference [CD-ROM], Chicago, IL, USA, 21–25 September 2019; pp. 1452–1466. [Google Scholar]
- Gu, A.Z.; Tooker, N.; Onnis-Hayden, A.; Wang, D.; Srinivasan, V.; Li, G.; Takács, I.; Vargas, E. Optimization and Design of a Side-Stream EBPR Process as a Sustainable Approach for Achieving Stable and Efficient Phosphorus Removal; Project No. U1R13/4869; WRF: Alexandria, VA, USA, 2019. [Google Scholar]
- Ferrera, I.; Sánchez, O. Insights into microbial diversity in wastewater treatment systems: How far have we come? Biotechnol. Adv. 2016, 34, 790–802. [Google Scholar] [CrossRef] [Green Version]
- Guo, G.; Wu, D.; Hao, T.; Mackey, H.R.; Wei, L.; Chen, G. Denitrifying sulfur conversion-associated EBPR: The effect of pH on anaerobic metabolism and performance. Water Res. 2017, 123, 687–695. [Google Scholar] [CrossRef]
- Mulkerrins, D.; Dobson, A.; Colleran, E. Parameters affecting biological phosphate removal from wastewaters. Environ. Int. 2004, 30, 249–259. [Google Scholar] [CrossRef]
- Gebremariam, S.Y.; Beutel, M.W.; Christian, D.; Hess, T.F. Research Advances and Challenges in the Microbiology of Enhanced Biological Phosphorus Removal—A Critical Review. Water Environ. Res. 2011, 83, 195–219. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, M.; Wang, Y.; Liu, F.; Qin, M.; Zhang, Y.; Zhou, W. The condition optimization and mechanism of aerobic phosphorus removal by marine bacterium Shewanella sp. Chem. Eng. J. 2018, 345, 611–620. [Google Scholar] [CrossRef]
- Panswad, T.; Doungchai, A.; Anotai, J. Temperature effect on microbial community of enhanced biological phosphorus removal system. Water Res. 2003, 37, 409–415. [Google Scholar] [CrossRef]
- Weissbrodt, D.G.; Neu, T.R.; Kuhlicke, U.; Rappaz, Y.; Holliger, C. Assessment of bacterial and structural dynamics in aerobic granular biofilms. Front. Microbiol. 2013, 4, 175. [Google Scholar] [CrossRef] [Green Version]
- Whang, L.-M.; Park, J.K. Competition between Polyphosphate- and Glycogen-Accumulating Organisms in Enhanced-Biological-Phosphorus-Removal Systems: Effect of Temperature and Sludge Age. Water Environ. Res. 2006, 78, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Poh, P.K.; Ong, Y.H.; Arumugam, K.; Nittami, T.; Yeoh, H.K.; Bessarab, I.; William, R.; Chua, A.S.M. Tropical-based EBPR process: The long-term stability, microbial community and its response towards temperature stress. Water Environ. Res. 2021, 93, 2598–2608. [Google Scholar] [CrossRef] [PubMed]
- Qiu, G.; Zuniga-Montanez, R.; Law, Y.; Thi, S.S.; Nguyen, T.Q.N.; Eganathan, K.; Liu, X.; Nielsen, P.H.; Williams, R.B.; Wuertz, S. Polyphosphate-accumulating organisms in full-scale tropical wastewater treatment plants use diverse carbon sources. Water Res. 2019, 149, 496–510. [Google Scholar] [CrossRef]
- Wang, L.; Shen, N.; Oehmen, A.; Zhou, Y. The impact of temperature on the metabolism of volatile fatty acids by polyphosphate accumulating organisms (PAOs). Environ. Res. 2020, 188, 109729. [Google Scholar] [CrossRef] [PubMed]
- Mielczarek, A.T.; Nguyen, H.T.; Nielsen, J.L.; Nielsen, P.H. Population dynamics of bacteria involved in enhanced biological phosphorus removal in Danish wastewater treatment plants. Water Res. 2013, 47, 1529–1544. [Google Scholar] [CrossRef]
- Liau, K.F.; Shoji, T.; Ong, Y.H.; Chua, A.S.M.; Yeoh, H.K.; Ho, P.Y. Kinetic and stoichiometric characterization for efficient enhanced biological phosphorus removal (EBPR) process at high temperatures. Bioprocess Biosyst. Eng. 2015, 38, 729–737. [Google Scholar] [CrossRef]
- Xu, Y.; Hu, H.; Liu, J.; Luo, J.; Qian, G.; Wang, A. pH dependent phosphorus release from waste activated sludge: Contributions of phosphorus speciation. Chem. Eng. J. 2015, 267, 260–265. [Google Scholar] [CrossRef]
- Wang, D.; Tooker, N.B.; Srinivasan, V.; Li, G.; Fernandez, L.A.; Schauer, P.; Menniti, A.; Maher, C.; Bott, C.B.; Dombrowski, P.; et al. Side-stream enhanced biological phosphorus removal (S2EBPR) process improves system performance—A full-scale comparative study. Water Res. 2019, 167, 115109. [Google Scholar] [CrossRef] [PubMed]
- Kang, A.J.; Munz, G.; Yuan, Q. Influence of pH control on material characteristics, bacterial community composition and BNR performance of mature aerobic granules. Process Saf. Environ. Prot. 2019, 124, 158–166. [Google Scholar] [CrossRef]
- Filipe, C.D.; Daigger, G.T.; Grady, C.L. pH as a key factor in the competition between glycogen-accumulating organisms and phosphorus-accumulating organisms. Water Environ. Res. 2001, 73, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Saunders, A.M.; Mabbett, A.N.; McEwan, A.G.; Blackall, L.L. Proton motive force generation from stored polymers for the uptake of acetate under anaerobic conditions. FEMS Microbiol. Lett. 2007, 274, 245–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belka, D. The Effect of pH on Organic Carbon Uptake and Biological Phosphorus Removal by Tetrasphaera Polyphosphate Accumulating Organisms. 2021. Available online: https://digitalrepository.unm.edu/ce_etds/259 (accessed on 8 October 2022).
- Zuthi, M.; Guo, W.; Ngo, H.; Nghiem, L.; Hai, F. Enhanced biological phosphorus removal and its modeling for the activated sludge and membrane bioreactor processes. Bioresour. Technol. 2013, 139, 363–374. [Google Scholar] [CrossRef]
- Chen, H.-B.; Wang, D.-B.; Li, X.-M.; Yang, Q.; Luo, K.; Zeng, G.-M. Temperature influence on biological phosphorus removal induced by aerobic/extended-idle regime. Environ. Sci. Pollut. Res. 2014, 21, 6034–6043. [Google Scholar] [CrossRef]
Figure 1.
A typical configuration for P removal in the mainstream the mainstream EBPR (Phoredox process) (a), sidestream EBPR (PhoStrip process) (b) (abbreviations: EFF—effluent, RAS—recirculated activated sludge, WAS—waste activated sludge).
Figure 1.
A typical configuration for P removal in the mainstream the mainstream EBPR (Phoredox process) (a), sidestream EBPR (PhoStrip process) (b) (abbreviations: EFF—effluent, RAS—recirculated activated sludge, WAS—waste activated sludge).

Figure 2.
Comparison of the metabolic pathways and biochemical transformation of P between Tetrasphaera and Ca. Accumulibacter in full-scale EBPR. SPO4—external phosphate, XPHA—internal polyhydroxyalkanoates, XPAO—biomass of the PAO, XSTO?—not yet characterized internal carbon storage material, XPAO, T—biomass of Tetrasphaera. Adapted from Makinia and Zaborowska [48].
Figure 2.
Comparison of the metabolic pathways and biochemical transformation of P between Tetrasphaera and Ca. Accumulibacter in full-scale EBPR. SPO4—external phosphate, XPHA—internal polyhydroxyalkanoates, XPAO—biomass of the PAO, XSTO?—not yet characterized internal carbon storage material, XPAO, T—biomass of Tetrasphaera. Adapted from Makinia and Zaborowska [48].

Figure 3.
Overview of the global abundance of the denitrifying PAOs within the general microbial community in EBPR systems (based on data from [4,34]).

Figure 4.
Comparison of PAO and Tetrasphaera mentions in publications related to EBPR. Summary of the research trends over the past two decades against the number of scientific papers with the mention of PAOs and Tetrasphaera.
Figure 4.
Comparison of PAO and Tetrasphaera mentions in publications related to EBPR. Summary of the research trends over the past two decades against the number of scientific papers with the mention of PAOs and Tetrasphaera.

Figure 5.
Examples of the mainstream EBPR system configurations ((A)—Bardenpho) and sidestream application ((B)—sidestream RAS plus carbon [SSRC], (C)—sidestream RAS [SSR], (D)—Sidestream Mixed Liquor Suspended Solids [SSM]). RAS—recirculated activated sludge, WAS—waste activated sludge, HRT—hydraulic retention time, SRT—sludge retention time.
Figure 5.
Examples of the mainstream EBPR system configurations ((A)—Bardenpho) and sidestream application ((B)—sidestream RAS plus carbon [SSRC], (C)—sidestream RAS [SSR], (D)—Sidestream Mixed Liquor Suspended Solids [SSM]). RAS—recirculated activated sludge, WAS—waste activated sludge, HRT—hydraulic retention time, SRT—sludge retention time.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of preferable electron acceptors by the main functional bacterial groups involved in EBPR.
Table 1.
Summary of preferable electron acceptors by the main functional bacterial groups involved in EBPR.
| Bacterial Functional Group | Electron Acceptors |
|---|---|
| APAO | DO |
| DPAO I | NO3−→NO2−→N2O→N2 |
| DPAO II * | NO2−→N2O→N2 |
| GAO | NO3−→NO2−→N2O→N2 |
| T. australiensis T. japonica T. elongata | NO3−→NO2−→N2O→N2 |
| T. jenkinsii, T. vanveenii, T. veronensis | DO |
Note(s): * Reduction abilities towards nitrate or nitrite of the EBPR microbes.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Otieno, J.; Kowal, P.; Mąkinia, J. The Occurrence and Role of Tetrasphaera in Enhanced Biological Phosphorus Removal Systems. Water 2022, 14, 3428. https://doi.org/10.3390/w14213428
AMA Style
Otieno J, Kowal P, Mąkinia J. The Occurrence and Role of Tetrasphaera in Enhanced Biological Phosphorus Removal Systems. Water. 2022; 14(21):3428. https://doi.org/10.3390/w14213428
Chicago/Turabian StyleOtieno, Jeremiah, Przemysław Kowal, and Jacek Mąkinia. 2022. "The Occurrence and Role of Tetrasphaera in Enhanced Biological Phosphorus Removal Systems" Water 14, no. 21: 3428. https://doi.org/10.3390/w14213428
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.