

Combined Effects of Polystyrene Nanoplastics and Enrofloxacin on the Life Histories and Gut Microbiota of Daphnia magna
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Chemicals
2.3. Experimental System
2.4. Experimental Design
2.4.1. Experimental Protocol
2.4.2. Life History Parameters
2.4.3. Metabolomic Diversity of Gut Microbiota
2.4.4. Taxonomic Diversity of Gut Microbiota
2.5. Statistical Analysis
3. Results
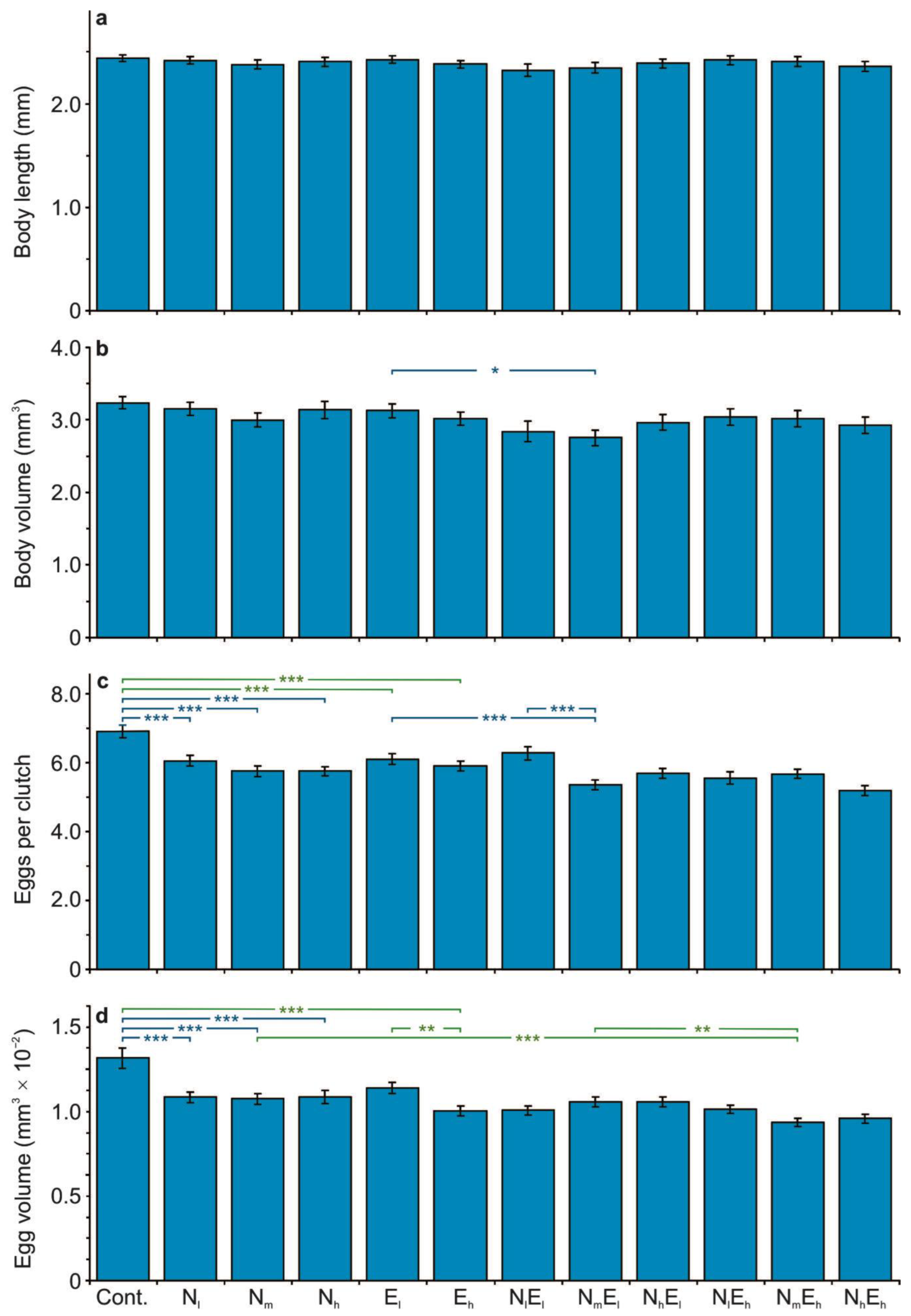
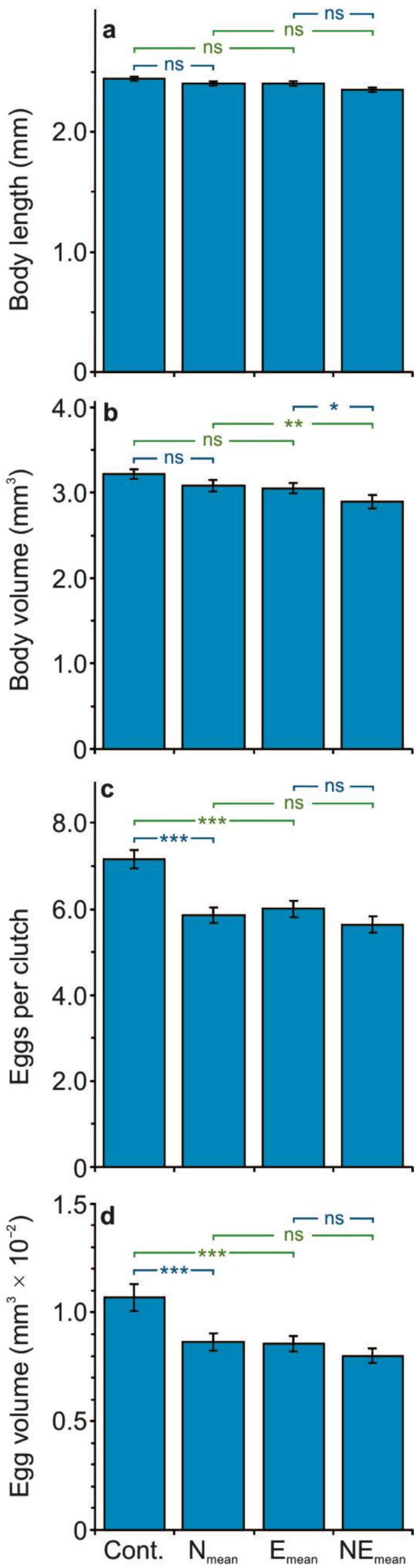
3.1. The Effect of the Stressors on the Life History Parameters
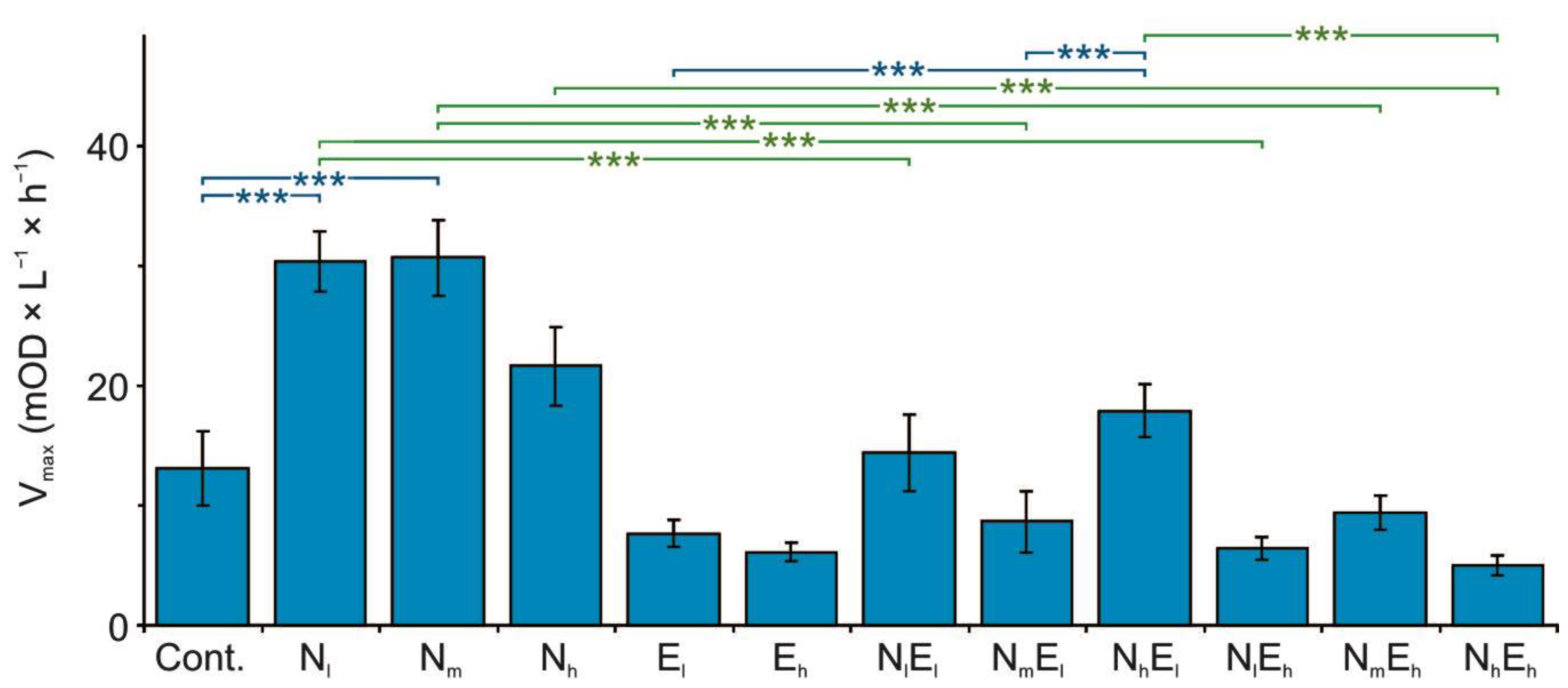
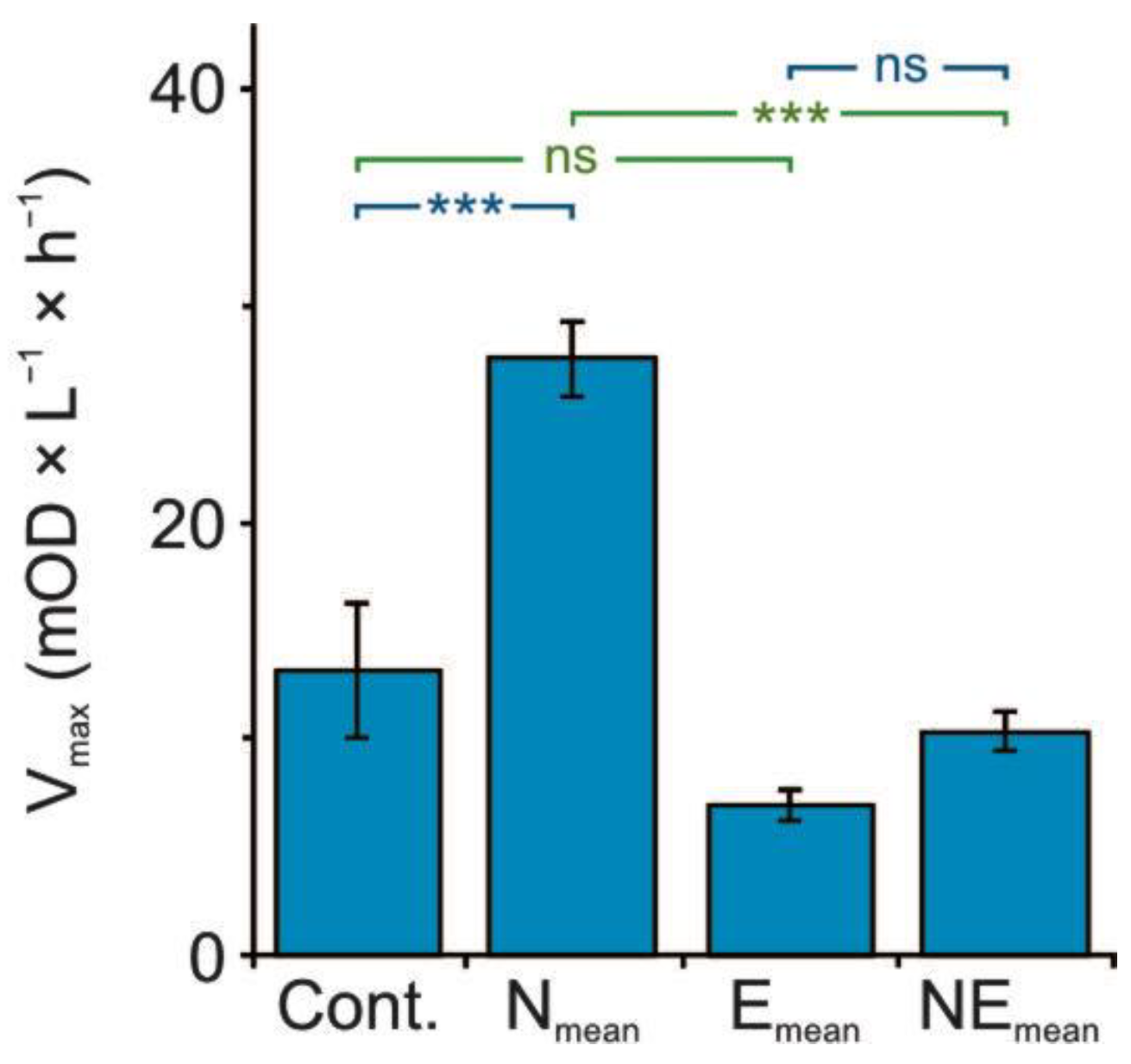
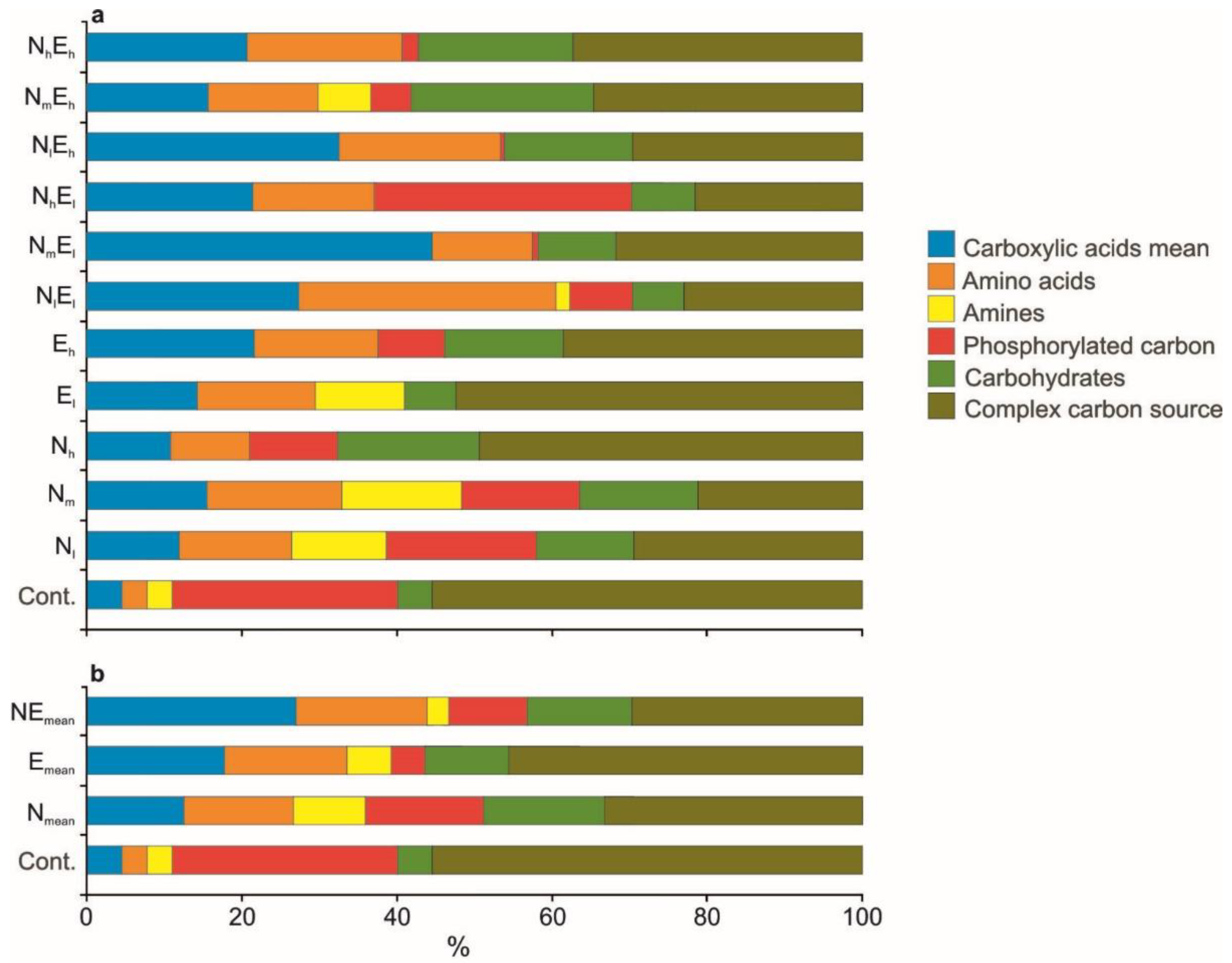
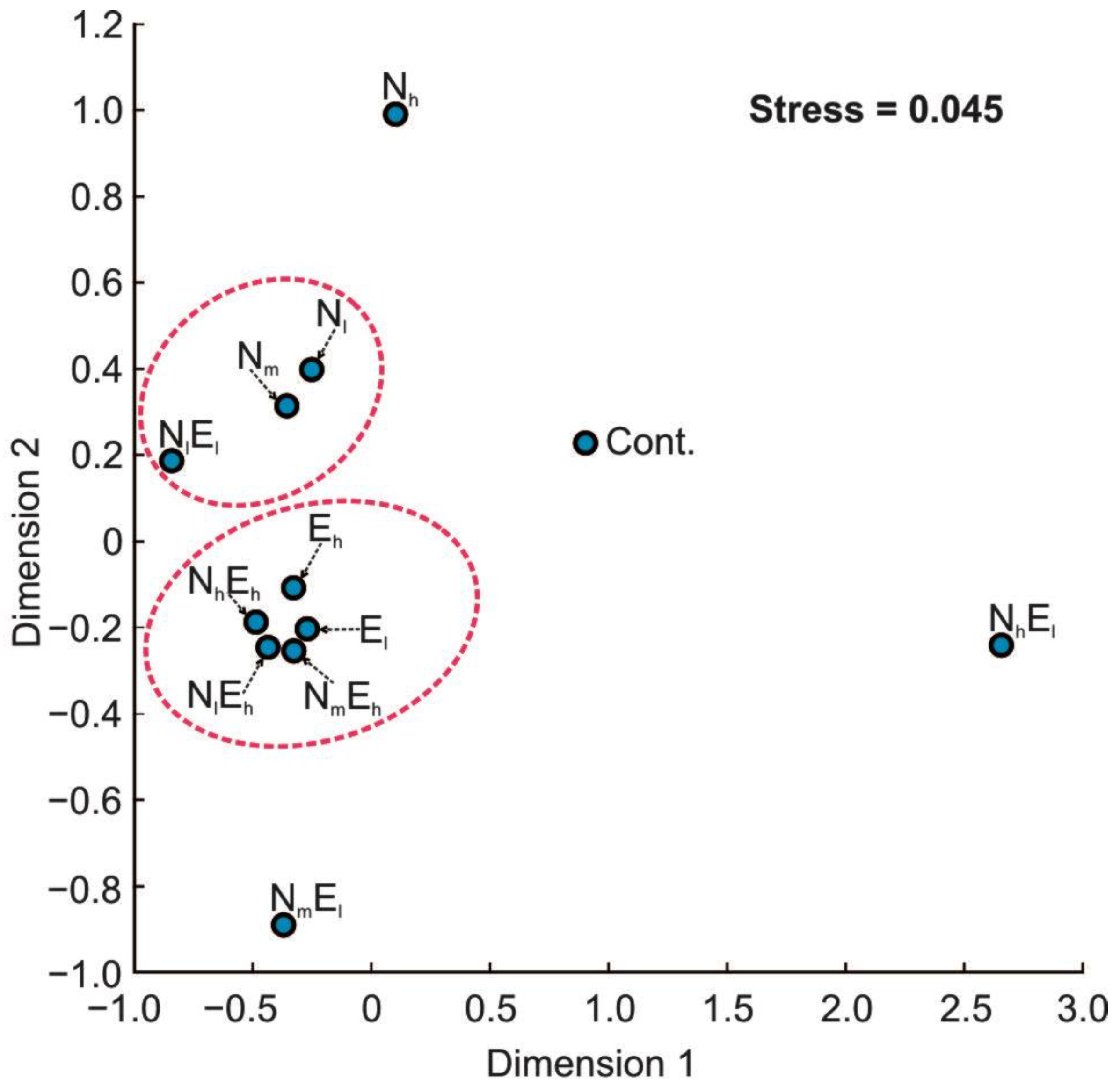
3.2. Metabolomic Diversity of Gut Microbiota
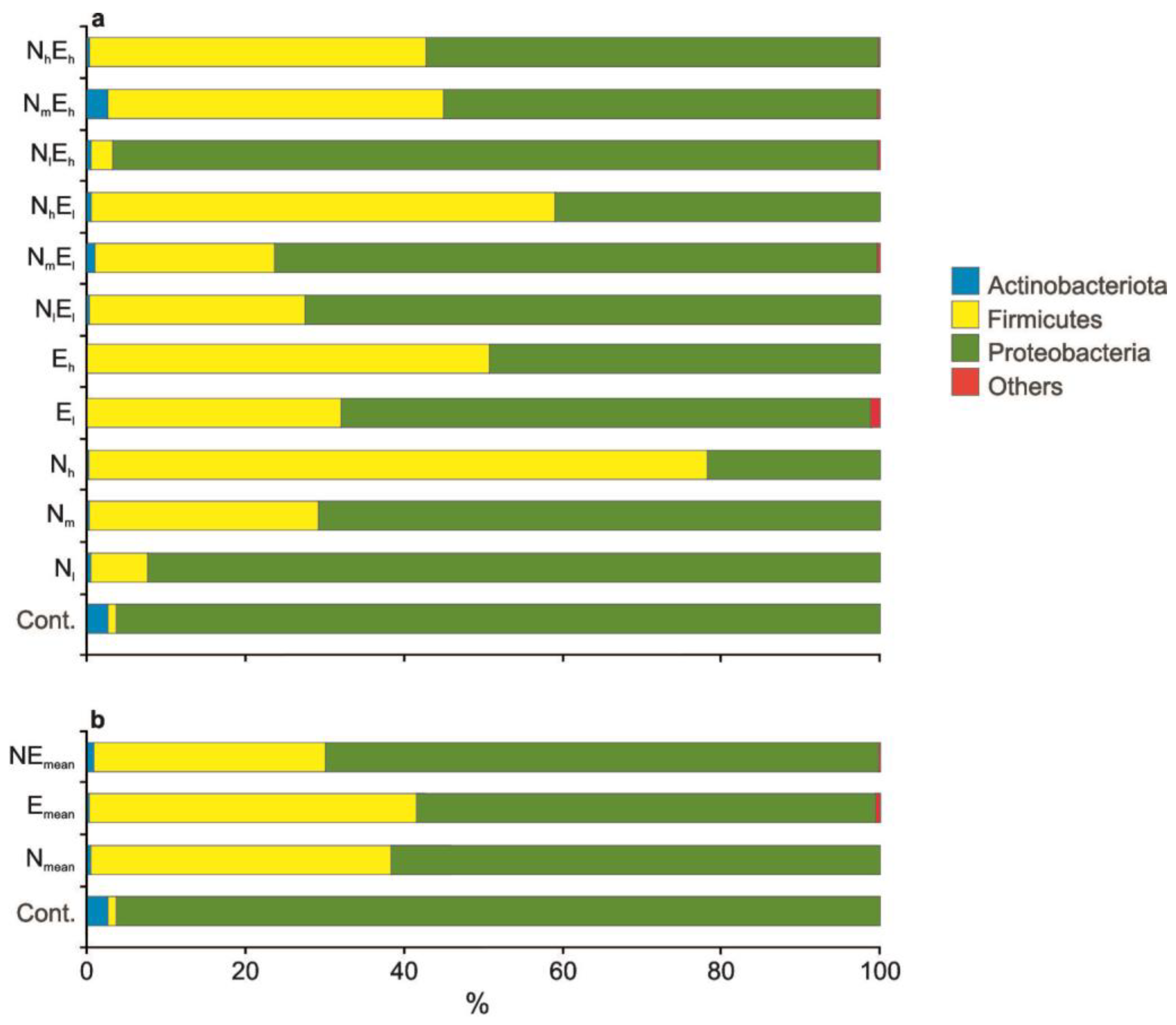
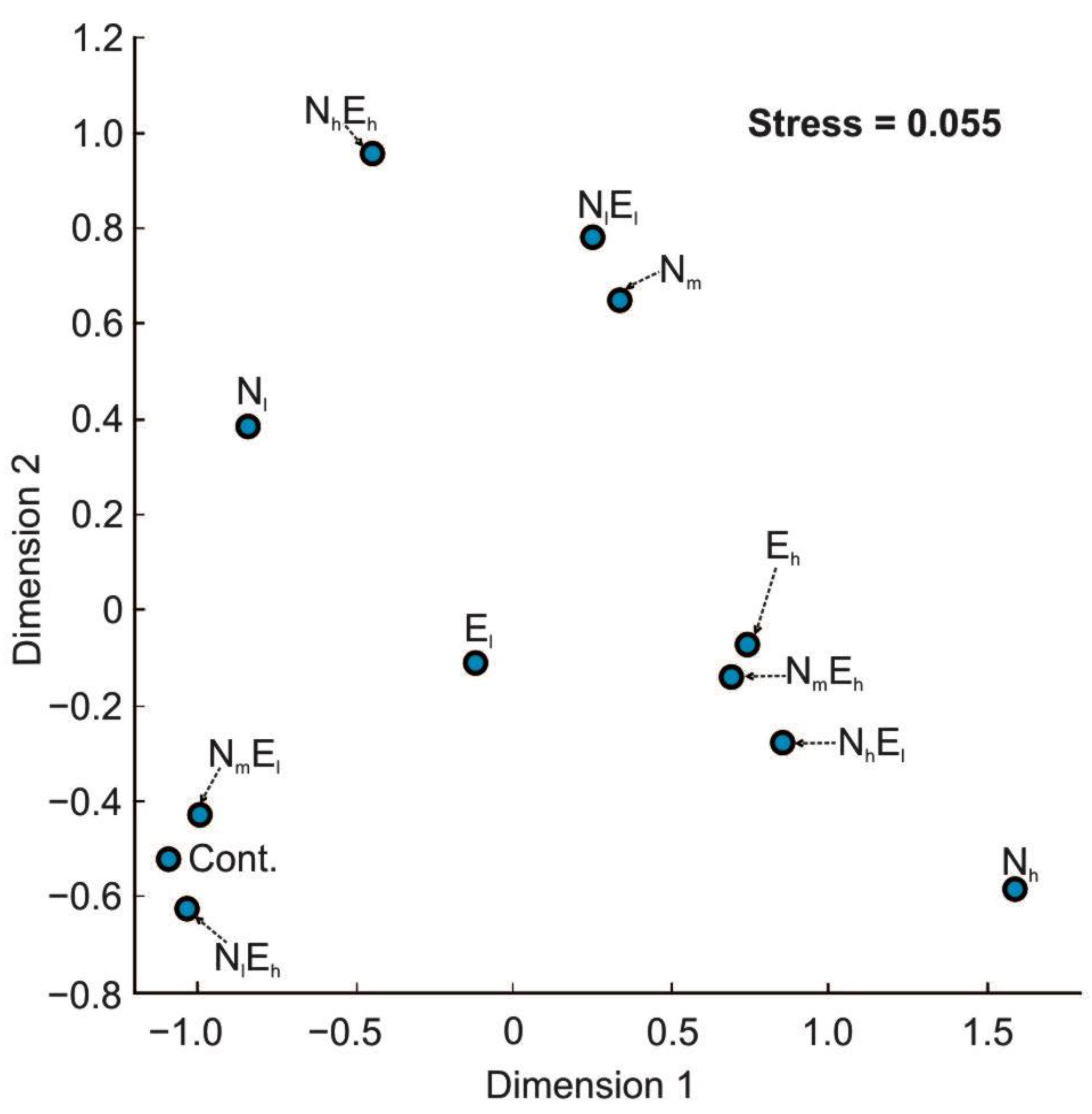
3.3. Taxonomic Diversity of Gut Microbiota
4. Discussion
4.1. General Effect
4.2. The Effect of Single and Combined Stressors on the Life History Parameters
4.3. The Effect of Single and Combined Stressors on the Metabolomic Diversity of Gut Microbiota
4.4. The Effect of Single and Combined Stressors on the Taxonomic Diversity of Gut Microbiota
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- MacLeod, M.; Arp, H.P.H.; Tekman, M.B.; Jahnke, A. The global threat from plastic pollution. Science 2021, 373, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.K.; Galgani, F.; Thompson, R.C.; Barlaz, M. Accumulation and fragmentation of plastic debris in global environments. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1985–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kedzierski, M.; Frčre, D.; Le Maguer, G.; Bruzaud, S. Why is there plastic packaging in the natural environment? Understanding the roots of our individual plastic waste management behaviours. Sci. Total Environ. 2020, 740, 139985. [Google Scholar] [CrossRef] [PubMed]
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gigault, J.; Ter Halle, A.; Baudrimont, M.; Pascal, P.Y.; Gauffre, F.; Phi, T.L.; Reynaud, S. Current opinion: What is a nanoplastic? Environ. Pollut. 2018, 235, 1030–1034. [Google Scholar] [CrossRef]
- Besseling, E.; Redondo-Hasselerharm, P.; Foekema, E.M.; Koelmans, A.A. Quantifying ecological risks of aquatic micro-and nanoplastic. Crit. Rev. Environ. Sci. Technol. 2019, 49, 32–80. [Google Scholar] [CrossRef] [Green Version]
- Egessa, R.; Nankabirwa, A.; Ocaya, H.; Pabire, W.G. Microplastic pollution in surface water of Lake Victoria. Sci. Total Environ. 2020, 741, 140201. [Google Scholar] [CrossRef]
- Huang, D.; Chen, H.; Shen, M.; Tao, J.; Chen, S.; Yin, L.; Li, R. Recent advances on the transport of microplastics/nanoplastics in abiotic and biotic compartments. J. Hazard. Mater. 2022, 438, 129515. [Google Scholar] [CrossRef]
- Koelmans, A.A.; Besseling, E.; Shim, W.J. Nanoplastics in the aquatic environment. Critical review. Mar. Anthropog. Litter 2015, 325–340. [Google Scholar]
- Ter Halle, A.; Jeanneau, L.; Martignac, M.; Jardé, E.; Pedrono, B.; Brach, L.; Gigault, J. Nanoplastic in the North Atlantic subtropical gyre. Environ. Sci. Technol. 2017, 51, 13689–13697. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Lin, S.; Turner, J.P.; Ke, P.C. Physical adsorption of charged plastic nanoparticles affects algal photosynthesis. J. Phys. Chem. C 2010, 114, 16556–16561. [Google Scholar] [CrossRef]
- Booth, A.M.; Hansen, B.H.; Frenzel, M.; Johnsen, H.; Altin, D. Uptake and toxicity of methylmethacrylate-based nanoplastic particles in aquatic organisms. Environ. Toxicol. Chem. 2016, 35, 1641–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maszczyk, P.; Pijanowska, J.; Mrówka, P.; Babkiewicz, E. Effects of nanoplastics on aquatic organisms. In Environmental Nanopollutants: Sources, Occurrence, Analysis and Fate; CRC Press: Boca Raton, FL, USA, 2022. [Google Scholar]
- El Hadri, H.; Gigault, J.; Maxit, B.; Grassl, B.; Reynaud, S. Nanoplastic from mechanically degraded primary and secondary microplastics for environmental assessments. NanoImpact 2020, 17, 100206. [Google Scholar] [CrossRef]
- Gaylarde, C.C.; Neto, J.A.B.; da Fonseca, E.M. Nanoplastics in aquatic systems-are they more hazardous than microplastics? Environ. Pollut. 2021, 272, 115950. [Google Scholar] [CrossRef] [PubMed]
- Schrank, I.; Trotter, B.; Dummert, J.; Scholz-Böttcher, B.M.; Löder, M.G.; Laforsch, C. Effects of microplastic particles and leaching additive on the life history and morphology of Daphnia magna. Environ. Pollut. 2019, 255, 113233. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Na, J.; An, D.; Jung, J. Role of benzophenone-3 additive in chronic toxicity of polyethylene microplastic fragments to Daphnia magna. Sci. Total Environ. 2021, 800, 149638. [Google Scholar] [CrossRef]
- Zhao, H.J.; Xu, J.K.; Yan, Z.H.; Ren, H.Q.; Zhang, Y. Microplastics enhance the developmental toxicity of synthetic phenolic antioxidants by disturbing the thyroid function and metabolism in developing zebrafish. Environ. Int. 2020, 140, 105750. [Google Scholar] [CrossRef]
- Li, Z.; Li, W.; Zha, J.; Chen, H.; Martyniuk, C.J.; Liang, X. Transcriptome analysis reveals benzotriazole ultraviolet stabilizers regulate networks related to inflammation in juvenile zebrafish (Danio rerio) brain. Environ. Toxicol. 2019, 34, 112–122. [Google Scholar] [CrossRef]
- Gauthier, P.T.; Norwood, W.P.; Prepas, E.E.; Pyle, G.G. Metal–PAH mixtures in the aquatic environment: A review of co-toxic mechanisms leading to more-than-additive outcomes. Aquat. Toxicol. 2014, 154, 253–269. [Google Scholar] [CrossRef]
- Trevisan, R.; Voy, C.; Chen, S.; Di Giulio, R.T. Nanoplastics decrease the toxicity of a complex PAH mixture but impair mitochondrial energy production in developing zebrafish. Environ. Sci. Technol. 2019, 53, 8405–8415. [Google Scholar] [CrossRef]
- Menéndez-Pedriza, A.; Jaumot, J. Interaction of environmental pollutants with microplastics: A critical review of sorption factors, bioaccumulation and ecotoxicological effects. Toxics 2020, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Li, D.; Wang, F.; Zhang, C.; Yue, C.; HuangF, Y.; Mu, J. Parental exposure to sulfamethazine and nanoplastics alters the gut microbial communities in the offspring of marine madaka (Oryzias melastigma). J. Hazard. Mater. 2022, 423, 127003. [Google Scholar] [CrossRef]
- Liu, S.; Yan, L.; Zhang, Y.; Junaid, M.; Wang, J. Polystyrene nanoplastics exacerbated the ecotoxicological and potential carcinogenic effects of tetracycline in juvenile grass carp (Ctenopharyngodon idella). Sci. Total Environ. 2022, 803, 150027. [Google Scholar] [CrossRef] [PubMed]
- Canniff, P.M.; Hoang, T.C. Microplastic ingestion by Daphnia magna and its enhancement on algal growth. Sci. Total Environ. 2018, 633, 500–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevisan, R.; Ranasinghe, P.; Jayasundara, N.; Di Giulio, R.T. Nanoplastics in Aquatic Environments: Impacts on Aquatic Species and Interactions with Environmental Factors and Pollutants. Toxics 2022, 10, 326. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yin, D.; Jia, Y.; Schiwy, S.; Legradi, J.; Yang, S.; Hollert, H. Enhanced uptake of BPA in the presence of nanoplastics can lead to neurotoxic effects in adult zebrafish. Sci. Total Environ. 2017, 609, 1312–1321. [Google Scholar] [CrossRef]
- Mendes, L.A.; Barreto, A.; Santos, J.; Amorim, M.J.; Maria, V.L. Co-Exposure of Nanopolystyrene and Other Environmental Contaminants—Their Toxic Effects on the Survival and Reproduction of Enchytraeus crypticus. Toxics 2022, 10, 193. [Google Scholar] [CrossRef]
- Booth, A.; Aga, D.S.; Wester, A.L. Retrospective analysis of the global antibiotic residues that exceed the predicted no effect concentration for antimicrobial resistance in various environmental matrices. Environ. Int. 2020, 141, 105796. [Google Scholar] [CrossRef]
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Med. Chem. 2014, 6, S14459. [Google Scholar] [CrossRef] [Green Version]
- Zainab, S.M.; Junaid, M.; Xu, N.; Malik, R.N. Antibiotics and antibiotic resistant genes (ARGs) in groundwater: A global review on dissemination, sources, interactions, environmental and human health risks. Water Res. 2020, 187, 116455. [Google Scholar] [CrossRef]
- Hill, D.A.; Hoffmann, C.; Abt, M.C.; Du, Y.; Kobuley, D.; Kirn, T.J.; Artis, D. Metagenomic analyses reveal antibiotic-induced temporal and spatial changes in intestinal microbiota with associated alterations in immune cell homeostasis. Mucosal Immunol. 2010, 3, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Limbu, S.M.; Shen, M.; Zhai, W.; Qiao, F.; He, A.; Zhang, M. Environmental concentrations of antibiotics impair zebrafish gut health. Environ. Pollut. 2018, 235, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Kovalakova, P.; Cizmas, L.; McDonald, T.J.; Marsalek, B.; Feng, M.; Sharma, V.K. Occurrence and toxicity of antibiotics in the aquatic environment: A review. Chemosphere 2020, 251, 126351. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Cui, H.; Jia, X.; Huang, X. Occurrence and ecotoxicity of sulfonamides in the aquatic environment: A review. Sci. Total Environ. 2022, 820, 153178. [Google Scholar] [CrossRef]
- Baggio, D.; Ananda-Rajah, M.R. Fluoroquinolone antibiotics and adverse events. Aust. Prescr. 2021, 44, 161. [Google Scholar] [CrossRef]
- Brown, S.A. Fluoroquinolones in animal health. J. Vet. Pharmacol. Ther. 1996, 19, 1–14. [Google Scholar] [CrossRef]
- Migliore, L.; Cozzolino, S.; Fiori, M. Phytotoxicity to and uptake of flumequine used in intensive aquaculture on the aquatic weed, Lythrum salicaria L. Chemosphere 2000, 40, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, G.A. Mechanisms of resistance to quinolones. Clin. Infect. Dis. 2005, 41 (Suppl. 2), S120–S126. [Google Scholar] [CrossRef] [Green Version]
- Välitalo, P.; Kruglova, A.; Mikola, A.; Vahala, R. Toxicological impacts of antibiotics on aquatic micro-organisms: A mini-review. Int. J. Hyg. Environ. Health 2017, 220, 558–569. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Jung, J.; Kim, M.; Park, J.; Boxall, A.B.; Choi, K. Prioritizing veterinary pharmaceuticals for aquatic environment in Korea. Environ. Toxicol. Pharmacol. 2008, 26, 167–176. [Google Scholar] [CrossRef]
- Kern, W.V. New plasmid-borne quinolone-resistance determinant in Escherichia coli. Future Microbiol. 2007, 2, 473–475. [Google Scholar] [CrossRef]
- Ding, G.; Chen, G.; Liu, Y.; Li, M.; Liu, X. Occurrence and risk assessment of fluoroquinolone antibiotics in reclaimed water and receiving groundwater with different replenishment pathways. Sci. Total Environ. 2020, 738, 139802. [Google Scholar] [CrossRef] [PubMed]
- Wagil, M.; Kumirska, J.; Stolte, S.; Puckowski, A.; Maszkowska, J.; Stepnowski, P.; Białk-Bielińska, A. Development of sensitive and reliable LC-MS/MS methods for the determination of three fluoroquinolones in water and fish tissue samples and preliminary environmental risk assessment of their presence in two rivers in northern Poland. Sci. Total Environ. 2014, 493, 1006–1013. [Google Scholar] [CrossRef]
- Ma, Y.; Li, M.; Wu, M.; Li, Z.; Liu, X. Occurrences and regional distributions of 20 antibiotics in water bodies during groundwater recharge. Sci. Total Environ. 2015, 518, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yan, X.; Shen, Y.; Di, M.; Wang, J. Antibiotics in surface water and sediments from Hanjiang River, Central China: Occurrence, behavior and risk assessment. Ecotoxicol. Environ. Saf. 2018, 157, 150–158. [Google Scholar] [CrossRef] [PubMed]
- You, X.; Cao, X.; Zhang, X.; Guo, J.; Sun, W. Unraveling individual and combined toxicity of nano/microplastics and ciprofloxacin to Synechocystis sp. at the cellular and molecular levels. Environ. Int. 2021, 157, 106842. [Google Scholar] [CrossRef]
- Feng, L.J.; Shi, Y.; Li, X.Y.; Sun, X.D.; Xiao, F.; Sun, J.W.; Yuan, X.Z. Behavior of tetracycline and polystyrene nanoparticles in estuaries and their joint toxicity on marine microalgae Skeletonema costatum. Environ. Pollut. 2020, 263, 114453. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Cai, Y.; Ma, C.; Han, L.; Yang, Z. Combined toxicity of micro/nano scale polystyrene plastics and ciprofloxacin to Corbicula fluminea in freshwater sediments. Sci. Total Environ. 2020, 789, 147887. [Google Scholar] [CrossRef]
- Almeida, M.; Martins, M.A.; Soares, A.M.; Cuesta, A.; Oliveira, M. Polystyrene nanoplastics alter the cytotoxicity of human pharmaceuticals on marine fish cell lines. Environ. Toxicol. Pharmacol. 2019, 69, 57–65. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Chen, H.; He, S.; Wang, F.; Liu, Y.; Chen, M.; Mu, J. Subchronic toxicity of dietary sulfamethazine and nanoplastics in marine medaka (Oryzias melastigma): Insights from the gut microbiota and intestinal oxidative status. Ecotoxicol. Environ. Saf. 2021, 226, 112820. [Google Scholar] [CrossRef]
- Baysal, A.; Saygin, H. Co-occurence of antibiotics and micro (nano) plastics: A systematic review between 2016–2021. J. Environ. Sci. Health Part A 2022, 57, 519–539. [Google Scholar] [CrossRef] [PubMed]
- Zebrowski, M.; Babkiewicz, E.; Błażejewska, A.; Pukos, S.; Wawrzeńczak, J.; Wilczynski, W.; Zebrowski, J.; Slusarczyk, M.; Maszczyk, P. The effect of microplastics on the interspecific competition of Daphnia. Environ. Pollut. 2022, 313, 120121. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum Press: New York, NY, USA, 1975; pp. 29–60. [Google Scholar]
- Pessoni, L.; Veclin, C.; El Hadri, H.; Cugnet, C.; Davranche, M.; Pierson-Wickmann, A.C.; Reynaud, S. Soap-and metal-free polystyrene latex particles as a nanoplastic model. Environ. Sci. Nano 2019, 6, 2253–2258. [Google Scholar] [CrossRef]
- Jiménez-Lamana, J.; Marigliano, L.; Allouche, J.; Grassl, B.; Szpunar, J.; Reynaud, S. A novel strategy for the detection and quantification of nanoplastics by single particle inductively coupled plasma mass spectrometry (ICP-MS). Anal. Chem. 2020, 92, 11664–11672. [Google Scholar] [CrossRef] [PubMed]
- Bruijning, M.; ten Berge, A.C.; Jongejans, E. Population-level responses to temperature, density and clonal differences in Daphnia magna as revealed by integral projection modelling. Funct. Ecol. 2018, 32, 2407–2422. [Google Scholar] [CrossRef] [Green Version]
- Ge, L.; Na, G.; Zhang, S.; Li, K.; Zhang, P.; Ren, H.; Yao, Z. New insights into the aquatic photochemistry of fluoroquinolone antibiotics: Direct photodegradation, hydroxyl-radical oxidation, and antibacterial activity changes. Sci. Total Environ. 2015, 527, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Bartosiewicz, M.; Jabłoński, J.; Kozłowski, J.; Maszczyk, P. Brood space limitation of reproduction may explain growth after maturity in differently sized Daphnia species. J. Plankton Res. 2015, 37, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Dickerson, T.L.; Williams, H.N. Functional diversity of bacterioplankton in three North Florida freshwater lakes over an Annual Cycle. Microb. Ecol. 2011, 67, 34–44. [Google Scholar] [CrossRef]
- Kiersztyn, B.; Chróst, R.; Kaliński, T.; Siuda, W.; Bukowska, A.; Kowalczyk, G.; Grabowska, K. Structural and functional microbial diversity along a eutrophication gradient of interconnected lakes undergoing anthropopressure. Sci. Rep. 2019, 9, 11144. [Google Scholar] [CrossRef] [Green Version]
- Macke, E.; Callens, M.; De Meester, L.; Decaestecker, E. Host-genotype dependent gut microbiota drives zooplankton tolerance to toxic cyanobacteria. Nat. Commun. 2017, 8, 1608. [Google Scholar] [CrossRef] [Green Version]
- Callens, M.; Watanabe, H.; Kato, Y.; Miura, J.; Decaestecker, E. Microbiota inoculum composition affects holobiont assembly and host growth in Daphnia. Microbiome 2018, 6, 56. [Google Scholar] [CrossRef] [PubMed]
- Hegg, A.; Radersma, R.; Uller, T. A field experiment reveals seasonal variation in the Daphnia gut microbiome. Oikos 2021, 130, 2191–2201. [Google Scholar] [CrossRef]
- Bulteel, L.; Houwenhuyse, S.; Declerck, S.A.; Decaestecker, E. The role of microbiome and genotype in Daphnia magna upon parasite re-exposure. Genes 2021, 12, 70. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Knight, R. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckers, B.; Op De Beeck, M.; Thijs, S.; Truyens, S.; Weyens, N.; Boerjan, W.; Vangronsveld, J. Performance of 16s rDNA primer pairs in the study of rhizosphere and endosphere bacterial microbiomes in metabarcoding studies. Front. Microbiol. 2016, 7, 650. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Caporaso, J.G. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Kay, M.; Elkin, L.A.; Higgins, J.J.; Wobbrock, J.O. ARTool: Aligned Rank Transform for Nonparametric Factorial ANOVAs, R package version 0.11.1; Association for Computing Machinery: New York, NY, USA, 2021; Available online: https://github.com/mjskay/ARTool (accessed on 27 April 2021). [CrossRef]
- Wobbrock, J.O.; Findlater, L.; Gergle, D.; Higgins, J.J. The aligned rank transform for nonparametric factorial analyses using only ANOVA procedures. In Proceedings of the ACM Conference on Human Factors in Computing Systems (CHI 2011), Vancouver, BC, Canada, 7–12 May 2011; ACM Press: New York, NY, USA, 2011; pp. 143–146. Available online: https://depts.washington.edu/acelab/proj/art/ (accessed on 7 May 2011). [CrossRef] [Green Version]
- Elkin, L.A.; Kay, M.; Higgins, J.J.; Wobbrock, J.O. An aligned rank transform procedure for multifactor contrast tests. In Proceedings of the ACM Symposium on User Interface Software and Technology (UIST 2021), Virtual Event, 10–14 October 2021; ACM Press: New York, NY, USA, 2021; pp. 754–768. [Google Scholar] [CrossRef]
- Hammer, Ř.; Harper, D.A.; Ryan, D.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Akbar, S.; Gu, L.; Sun, Y.; Zhou, Q.; Zhang, L.; Lyu, K.; Yang, Z. Changes in the life history traits of Daphnia magna are associated with the gut microbiota composition shaped by diet and antibiotics. Sci. Total Environ. 2020, 705, 135827. [Google Scholar] [CrossRef]
- Rist, S.; Baun, A.; Hartmann, N.B. Ingestion of micro- and nanoplastics in Daphnia magna—Quantification of body burdens and assessment of feeding rates and reproduction. Environ. Pollut. 2017, 228, 398–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besseling, E.; Wang, B.; Lurling, M.; Koelmans, A.A. Nanoplastic affects growth of S. obliquus and reproduction of D. magna. Environ. Sci. Technol. 2014, 48, 12336–12343. [Google Scholar] [CrossRef] [PubMed]
- Bergami, E.; Pugnalini, S.; Vannuccini, M.L.; Manfra, L.; Faleri, C.; Savorelli, F.; Corsi, I. Long-term toxicity of surface-charged polystyrene nanoplastics to marine planktonic species Dunaliella tertiolecta and Artemia franciscana. Aquat. Toxicol. 2017, 189, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Venâncio, C.; Ferreira, I.; Martins, M.A.; Soares, A.M.; Lopes, I.; Oliveira, M. The effects of nanoplastics on marine plankton: A case study with polymethylmethacrylate. Ecotoxicol. Environ. Saf. 2019, 184, 109632. [Google Scholar] [CrossRef] [PubMed]
- Sökmen, T.Ö.; Sulukan, E.; Türkoğlu, M.; Baran, A.; Özkaraca, M.; Ceyhun, S.B. Polystyrene nanoplastics (20 nm) are able to bioaccumulate and cause oxidative DNA damages in the brain tissue of zebrafish embryo (Danio rerio). Neurotoxicology 2020, 77, 51–59. [Google Scholar] [CrossRef]
- Brun, N.R.; Koch, B.E.; Varela, M.; Peijnenburg, W.J.; Spaink, H.P.; Vijver, M.G. Nanoparticles induce dermal and intestinal innate immune system responses in zebrafish embryos. Environ. Sci. Nano 2018, 5, 904–916. [Google Scholar] [CrossRef]
- Redondo-Hasselerharm, P.E.; Vink, G.; Mitrano, D.M.; Koelmans, A.A. Metal-doping of nanoplastics enables accurate assessment of uptake and effects on Gammarus pulex. Environ. Sci. Nano 2021, 8, 1761–1770. [Google Scholar] [CrossRef]
- Fernandez, R.; Colás-Ruiz, N.R.; Bolívar-Anillo, H.J.; Anfuso, G.; Hampel, M. Occurrence and Effects of Antimicrobials Drugs in Aquatic Ecosystems. Sustainability 2021, 13, 13428. [Google Scholar] [CrossRef]
- Kostich, M.S.; Lazorchak, J.M. Risks to aquatic organisms posed by human pharmaceutical use. Sci. Total Environm. 2008, 389, 329–339. [Google Scholar] [CrossRef]
- Qiu, W.; Fang, M.; Magnuson, J.T.; Greer, J.B.; Chen, Q.; Zheng, Y.; Xiong, Y.; Luo, S.; Zheng, C.; Schlenk, D. Maternal exposure to environmental antibiotic mixture during gravid period predicts gastrointestinal effects in zebrafish offspring. J. Hazard. Mater. 2020, 399, 123009. [Google Scholar] [CrossRef]
- Nunes, B.; Leal, C.; Rodrigues, S.; Antunes, S.C. Assessment of ecotoxicological effects of ciprofloxacin in Daphnia magna: Life-history traits, biochemical and genotoxic effects. Water Sci. Technol. 2018, 2017, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Sheng, G.D.; Chen, Q.L.; O’Connor, P. Do combined nanoscale polystyrene and tetracycline impact on the incidence of resistance genes and microbial community disturbance in Enchytraeus crypticus? J. Hazard. Mater. 2020, 387, 122012. [Google Scholar] [CrossRef] [PubMed]
- Motiei, A.; Brindefalk, B.; Ogonowski, M.; El-Shehawy, R.; Pastuszek, P.; Ek, K.; Gorokhova, E. Disparate effects of antibiotic-induced microbiome change and enhanced fitness in Daphnia magna. PLoS ONE 2020, 15, e0214833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobritz, M.A.; Belenky, P.; Porter, C.B.; Gutierrez, A.; Yang, J.H.; Schwarz, E.G.; Collins, J.J. Antibiotic efficacy is linked to bacterial cellular respiration. Proc. Natl. Acad. Sci. USA 2015, 112, 8173–8180. [Google Scholar] [CrossRef] [Green Version]
- Stokes, J.M.; Lopatkin, A.J.; Lobritz, M.A.; Collins, J.J. Bacterial metabolism and antibiotic efficacy. Cell Metab. 2019, 30, 251–259. [Google Scholar] [CrossRef]
- Siuda, W.; Kiersztyn, B.; Chrost, R.J. The dynamics of protein decomposition in lakes of different trophic status-reflections on the assessment of the real proteolytic activity in situ. J. Microbiol. Biotechnol. 2007, 17, 897–904. [Google Scholar]
- Kataria, R.; Ruhal, R. Microbiological metabolism under chemical stress. In Microbial Biodegradation and Bioremediation; Elsevier: Amsterdam, The Netherlands, 2014; pp. 497–509. [Google Scholar]
- Miao, L.; Guo, S.; Liu, Z.; Liu, S.; You, G.; Qu, H.; Hou, J. Effects of nanoplastics on freshwater biofilm microbial metabolic functions as determined by BIOLOG ECO microplates. Int. J. Environ. Res. Public Health 2019, 16, 4639. [Google Scholar] [CrossRef] [Green Version]
- Cabral, D.J.; Penumutchu, S.; Reinhart, E.M.; Zhang, C.; Korry, B.J.; Wurster, J.I.; Belenky, P. Microbial metabolism modulates antibiotic susceptibility within the murine gut microbiome. Cell Metab. 2019, 30, 800–823. [Google Scholar] [CrossRef]
- Zuo, Y.; Xie, W.; Pang, Y.; Li, T.; Li, Q.; Li, Y. Bacterial community composition in the gut content of Lampetra japonica revealed by 16S rRNA gene pyrosequencing. PLoS ONE 2017, 12, e0188919. [Google Scholar] [CrossRef] [Green Version]
- Zheng, F.; Zhu, D.; Giles, M.; Daniell, T.; Neilson, R.; Zhu, Y.G.; Yang, X.R. Mineral and organic fertilization alters the microbiome of a soil nematode Dorylaimus stagnalis and its resistome. Sci. Total Environ. 2019, 680, 70–78. [Google Scholar] [CrossRef]
- Berg, M.; Stenuit, B.; Ho, J.; Wang, A.; Parke, C.; Knight, M.; Alvarez-Cohen, L.; Shapira, M. Assembly of the Caenorhabditis elegans gut microbiota from diverse soil microbial environments. ISME J. 2016, 10, 1998–2009. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.B.; Nie, L.; Chen, J. Current understanding on the roles of gut microbiota in fish disease and immunity. Zool. Res. 2019, 40, 70–76. [Google Scholar] [PubMed] [Green Version]
- Wolever, T.; Brighenti, F.; Royall, D.; Jenkins, A.L.; Jenkins, D.J. Effect of rectal infusion of short chain fatty acids in human subjects. Am. J. Gastroenterol. 1989, 84, 1027–1033. [Google Scholar] [PubMed]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Zhu, D.; An, X.L.; Chen, Q.L.; Yang, X.R.; Christie, P.; Ke, X.; Wu, L.H.; Zhu, Y.G. Antibiotics disturb the microbiome and increase the incidence of resistance genes in the gut of a common soil collembolan. Environ. Sci. Technol. 2018, 52, 3081–3090. [Google Scholar] [CrossRef]
- Saygin, H.; Baysal, A. Similarities and discrepancies between bio-based and conventional submicron-sized plastics: In relation to clinically important bacteria. Bull. Environ. Contam. Toxicol. 2020, 105, 26–35. [Google Scholar] [CrossRef]
- Zhu, D.; Li, G.; Wang, H.T.; Duan, G.L. Effects of nano-or microplastic exposure combined with arsenic on soil bacterial, fungal, and protistan communities. Chemosphere 2021, 281, 130998. [Google Scholar] [CrossRef]
- Auguste, M.; Lasa, A.; Balbi, T.; Pallavicini, A.; Vezzulli, L.; Canesi, L. Impact of nanoplastics on hemolymph immune parameters and microbiota composition in Mytilus galloprovincialis. Mar. Environ. Res. 2020, 159, 105017. [Google Scholar] [CrossRef]
- Kang, H.M.; Byeon, E.; Jeong, H.; Kim, M.S.; Chen, Q.; Lee, J.S. Different effects of nano-and microplastics on oxidative status and gut microbiota in the marine medaka Oryzias melastigma. J. Hazard. Mater. 2021, 405, 124207. [Google Scholar] [CrossRef]
- Gu, W.; Liu, S.; Chen, L.; Liu, Y.; Gu, C.; Ren, H.Q.; Wu, B. Single-cell RNA sequencing reveals size-dependent effects of polystyrene microplastics on immune and secretory cell populations from zebrafish intestines. Environ. Sci. Technol. 2020, 54, 3417–3427. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Zhou, A.; Wei, T.; Li, S.; Yang, B.; Xu, G.; Zou, J. Nanoplastics induce more serious microbiota dysbiosis and inflammation in the gut of adult zebrafish than microplastics. Bull. Environ. Contam. Toxicol. 2021, 107, 640–650. [Google Scholar] [CrossRef]
- Yoon, J.H.; Kang, S.J.; Schumann, P.; Oh, T.K. Yonghaparkia alkaliphila gen. nov., sp nov., a novel member of the family Microbacteriaceae isolated from an alkaline soil. Int. J. Syst. Evol. Microbiol. 2006, 56, 2415–2420. [Google Scholar] [CrossRef] [PubMed]
- Pishgar, R.; Dominic, J.A.; Sheng, Z.; Tay, J.H. Denitrification performance and microbial versatility in response to different selection pressures. Bioresour. Technol. 2019, 281, 72–83. [Google Scholar] [CrossRef]
- Oh, S.; Choi, D. Microbial community enhances biodegradation of bisphenol A through selection of Sphingomonadaceae. Microb. Ecol. 2019, 77, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.B.; Zhang, X.X.; Wu, B.; Xian, Q.M. Metagenomic insights into tetracycline effects on microbial community and antibiotic resistance of mouse gut. Ecotoxicology 2015, 24, 2125–2132. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbohydrates | D-cellobiose |
| α-D-lactose | |
| β-methyl-D-glucoside | |
| D-xylose | |
| Erythritol | |
| D-mannitol | |
| N-acetyl-D-glucosamine | |
| D-galactonic acid γ-lactone | |
| Phosphorylated carbons | glucose-1-phosphate |
| D,L-α-glycerol phosphate | |
| Amines | phenylethylamine |
| putrescine | |
| Carboxylic acids | D-glucosaminic acid |
| D-galacturonic acid | |
| γ-hydroxybutyric acid | |
| itaconic acid | |
| α-ketobutyric acid | |
| D-malic acid | |
| pyruvic acid methyl ester | |
| 2-hydroxy benzoic acid | |
| 4-hydroxy benzoic acid | |
| Complex carbon | Tween 40 |
| Tween 80 | |
| α-cyclodextrin | |
| glycogen | |
| Amino acids | L-arginine |
| L-asparagine | |
| L-phenylalanine | |
| L-serine | |
| L-threonine | |
| glycyl-L-glutamic acid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maszczyk, P.; Kiersztyn, B.; Gozzo, S.; Kowalczyk, G.; Jimenez-Lamana, J.; Szpunar, J.; Pijanowska, J.; Jines-Muñoz, C.; Zebrowski, M.L.; Babkiewicz, E. Combined Effects of Polystyrene Nanoplastics and Enrofloxacin on the Life Histories and Gut Microbiota of Daphnia magna. Water 2022, 14, 3403. https://doi.org/10.3390/w14213403
Maszczyk P, Kiersztyn B, Gozzo S, Kowalczyk G, Jimenez-Lamana J, Szpunar J, Pijanowska J, Jines-Muñoz C, Zebrowski ML, Babkiewicz E. Combined Effects of Polystyrene Nanoplastics and Enrofloxacin on the Life Histories and Gut Microbiota of Daphnia magna. Water. 2022; 14(21):3403. https://doi.org/10.3390/w14213403
Chicago/Turabian StyleMaszczyk, Piotr, Bartosz Kiersztyn, Sebastiano Gozzo, Grzegorz Kowalczyk, Javier Jimenez-Lamana, Joanna Szpunar, Joanna Pijanowska, Cristina Jines-Muñoz, Marcin Lukasz Zebrowski, and Ewa Babkiewicz. 2022. "Combined Effects of Polystyrene Nanoplastics and Enrofloxacin on the Life Histories and Gut Microbiota of Daphnia magna" Water 14, no. 21: 3403. https://doi.org/10.3390/w14213403