
Sulfide Treatment Alters Antioxidant Response and Related Genes Expressions in Rice Field Eel (Monopterus albus)
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Exposure
2.2. Sampling
2.3. Biochemical Parameter Analysis
2.4. Genes Expressions Analysis
2.5. Integrated Biomarker Response (IBR)
2.6. Statistical Analysis
3. Results
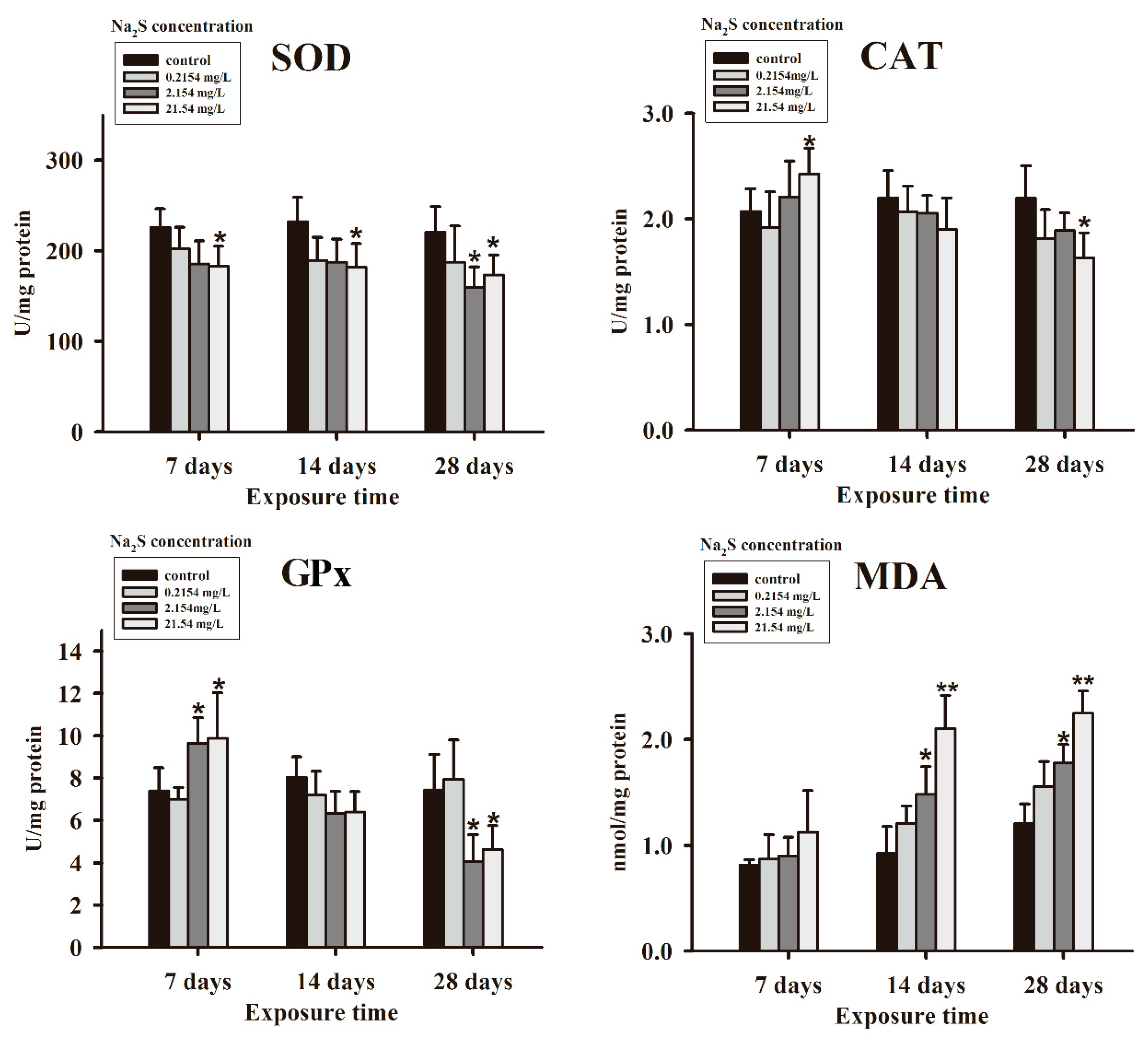
3.1. Antioxidant Parameters
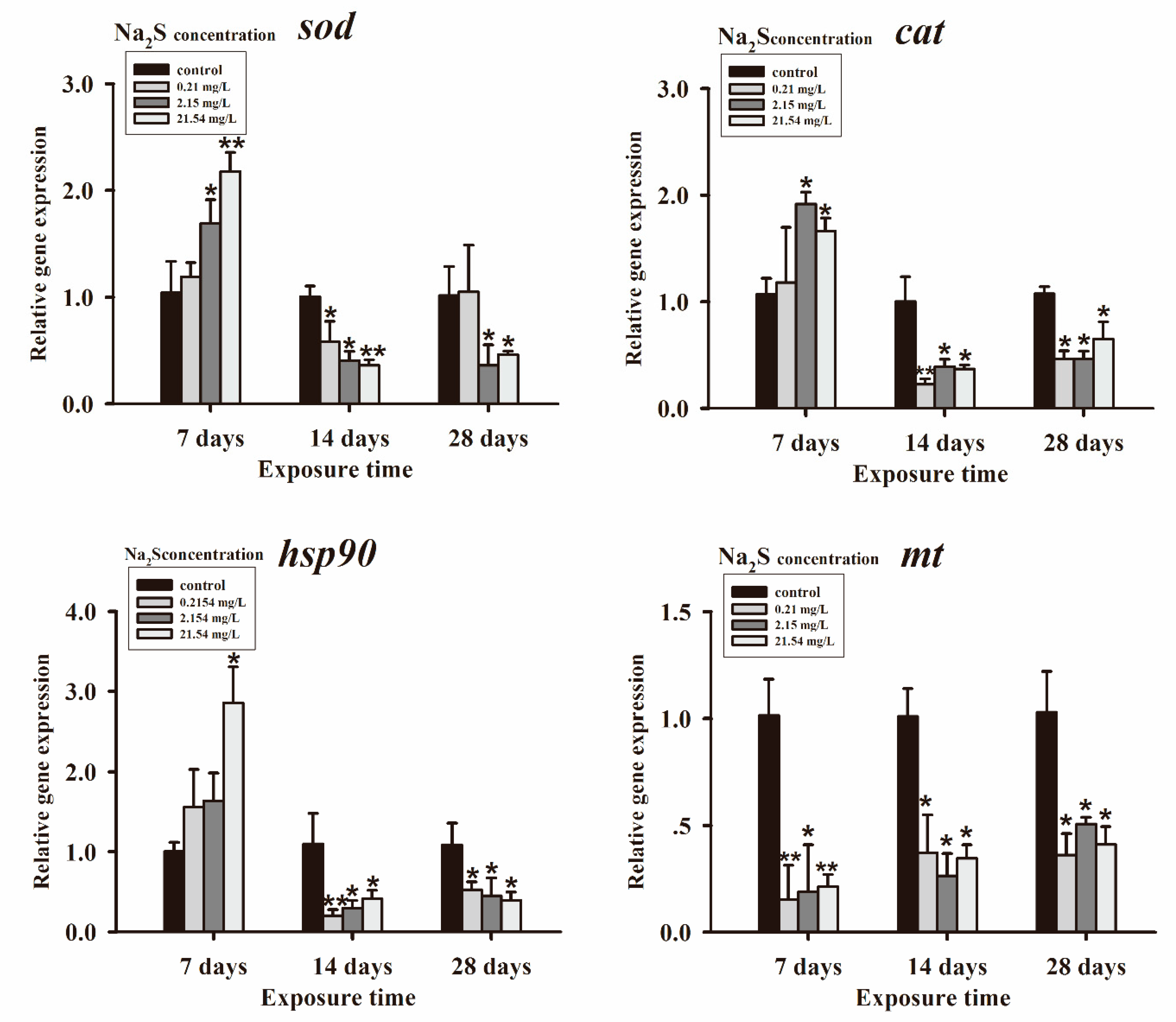
3.2. Genes Expressions
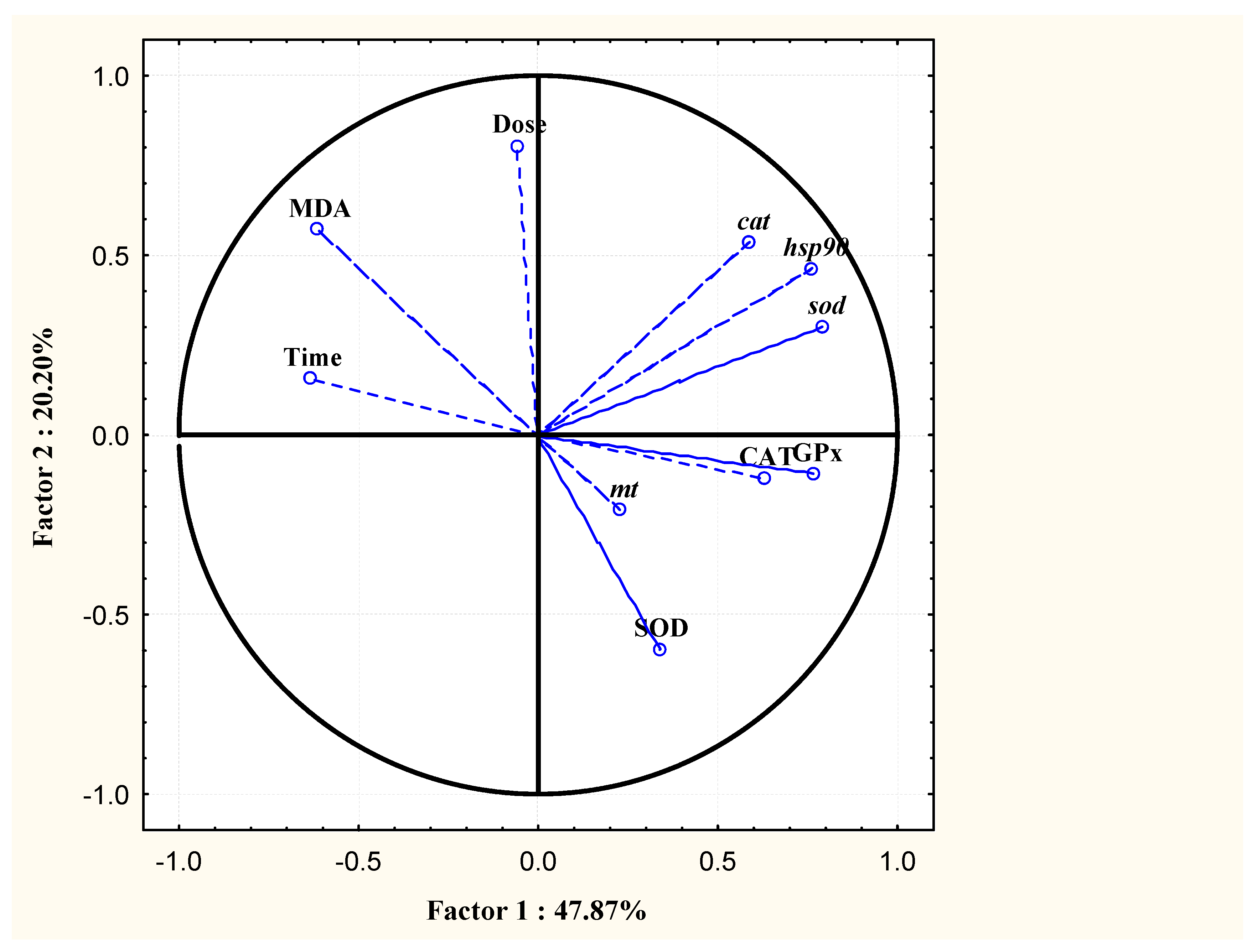
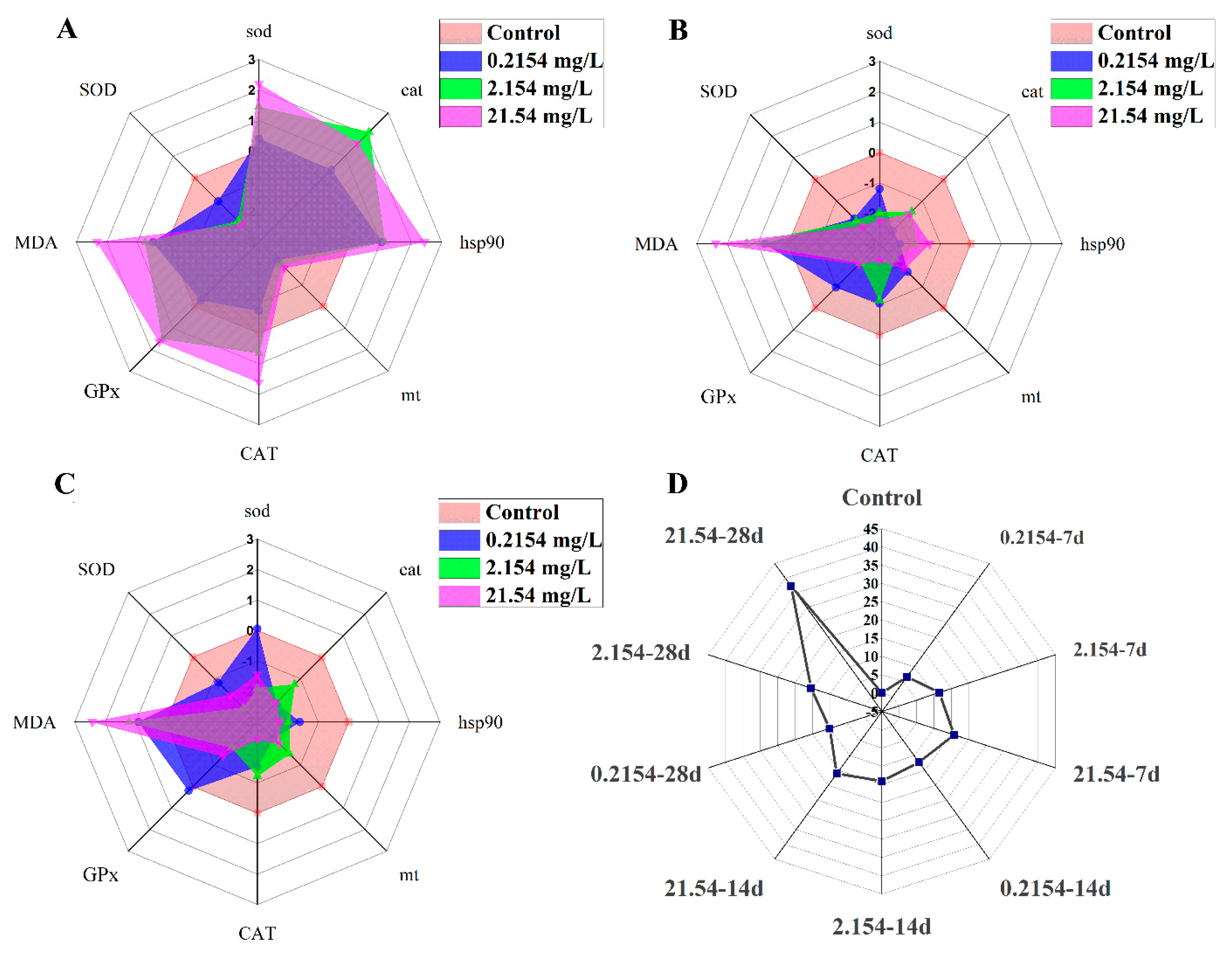
3.3. PCA and IBR Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bagarinao, T. Sulfide as an environmental factor and toxicant: Tolerance and adaptations in aquatic organisms. Aquat. Toxicol. 1992, 24, 21–62. [Google Scholar] [CrossRef]
- Shen, Y.Y.; Chen, J.Q.; Shen, W.L.; Chen, C.F.; Lin, Z.H.; Li, C.H. Molecular characterization of a novel sulfide: Quinone oxidoreductase from the razor clam Sinonovacula constricta and its expression response to sulfide stress. Comp. Biochem. Phys. B 2020, 239, 110367. [Google Scholar] [CrossRef]
- Shao, Y.; Chen, Z.L.; Wu, L.L. Oxidative Stress Effects of Soluble Sulfide on Human Hepatocyte Cell Line LO2. Int. J. Environ. Res. Public Health 2019, 16, 1662. [Google Scholar] [CrossRef] [Green Version]
- Petruci, J.F.D.; Cardoso, A.A. A new palladium chelate compound for determination of sulfide. Microchem. J. 2013, 106, 368–372. [Google Scholar] [CrossRef]
- Wu, L.L.; Shao, Y.; Hu, Z.J.; Gao, H.W. Effects of soluble sulfide on zebrafish (Danio rerio) embryonic development. Environ. Toxicol. Pharmcol. 2016, 42, 183–189. [Google Scholar] [CrossRef]
- Li, T.Y.; Li, E.C.; Suo, Y.T.; Xu, Z.X.; Jia, Y.Y.; Qin, J.G.; Chen, L.Q.; Gu, Z.M. Energy metabolism and metabolomics response of Pacific white shrimp Litopenaeus vannamei to sulfide toxicity. Aquat. Toxicol. 2017, 183, 28–37. [Google Scholar] [CrossRef]
- Forgan, L.G.; Forster, M.E. Oxygen consumption, ventilation frequency and cytochrome c oxidase activity in blue cod (Parapercis colias) exposed to hydrogen sulphide or isoeugenol. Comp. Biochem. Phys. C 2010, 151, 57–65. [Google Scholar] [CrossRef]
- Khan, A.A.; Schuler, M.M.; Prior, M.G.; Yong, S.; Coppock, R.W.; Florence, L.Z.; Lillie, L.E. Effects of hydrogen sulfide exposure on lung mitochondrial respiratory chain enzymes in rats. Toxicol. Appl. Pharmcol. 1990, 103, 482–490. [Google Scholar] [CrossRef]
- Suo, Y.T.; Li, E.C.; Li, T.Y.; Jia, Y.Y.; Qin, J.G.; Gu, Z.M.; Chen, L.Q. Response of gut health and microbiota to sulfide exposure in Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2017, 63, 87–96. [Google Scholar] [CrossRef]
- Guo, K.; Ruan, G.L.; Fan, W.H.; Fang, L.; Wang, Q.; Luo, M.Z.; Yi, T.L. The effect of nitrite and sulfide on the antioxidant capacity and microbial composition of the intestines of red swamp crayfish, Procambarus clarkii. Fish Shellfish Immunol. 2020, 96, 290–296. [Google Scholar] [CrossRef]
- Tay, A.S.; Chew, S.F.; Ip, Y.K. The swamp eel Monopterus albus reduces endogenous ammonia production and detoxifies ammonia to glutamine during 144 h of aerial exposure. J. Exp. Biol. 2003, 206, 2473–2486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.M.; Zhang, J.; Guo, F.; Wei, L.L.; Zhou, Q.B. Oxidative stress and inflammatory responses in the liver of swamp eel (Monopterus albus) exposed to carbon tetrachloride. Aquaculture 2018, 496, 232–238. [Google Scholar] [CrossRef]
- Ma, X.; Hu, Y.; Wang, X.Q.; Ai, Q.H.; He, Z.G.; Feng, F.X.; Lu, X.Y. Effects of practical dietary protein to lipid levels on growth, digestive enzyme activities and body composition of juvenile rice field eel (Monopterus albus). Aquac. Int. 2014, 22, 749–760. [Google Scholar] [CrossRef]
- Xiang, J.H.; Qin, L.; Zhao, D.; Xiong, F.; Wang, G.T.; Zou, H.; Li, W.X.; Li, M.; Song, K.B.; Wu, S.G. Growth performance, immunity and intestinal microbiota of swamp eel (Monopterus albus) fed a diet supplemented with house fly larvae (Musca domestica). Aquac. Nutr. 2020, 26, 693–704. [Google Scholar] [CrossRef]
- Yue, H.M.; Huang, X.Q.; Ruan, R.; Ye, H.; Li, Z.; Li, C.J. Effects of dietary protein levels on the growth, body composition, serum biochemistry and digestive enzyme activity in Chinese rice field eel (Monopterus albus) fingerlings. Aquac. Res. 2020, 51, 400–409. [Google Scholar] [CrossRef]
- Schröder, P.; Krutmann, J. Environmental Oxidative Stress—Environmental Sources of ROS. In Reactions, Processes: Oxidants and Antioxidant Defense Systems; Grune, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 19–31. [Google Scholar]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Guemouri, L.; Artur, Y.; Herbeth, B.; Jeandel, C.; Cuny, G.; Siest, G. Biological variability of superoxide dismutase, glutathione peroxidase, and catalase in blood. Clin. Chem. 1991, 37, 1932–1937. [Google Scholar] [CrossRef]
- Aliahmat, N.S.; Noor, M.R.M.; Yusof, W.J.W.; Makpol, S.; Ngah, W.Z.W.; Yusof, Y.A.M. Antioxidant enzyme activity and malondialdehyde levels can be modulated by Piper betle, tocotrienol rich fraction and Chlorella vulgaris in aging C57BL/6 mice. Clinics 2012, 67, 1447–1454. [Google Scholar] [CrossRef]
- Camkurt, M.A.; Findikli, E.; Bakacak, M.; Tolun, F.I.; Karaaslan, M.F. Evaluation of Malondialdehyde, Superoxide Dismutase and Catalase Activity in Fetal Cord Blood of Depressed Mothers. Clin. Psychopharmacol. Neurosci. 2017, 15, 35–39. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Bremner, I. Oxygen free radicals and metallothionein. Free Radic. Biol. Med. 1993, 14, 325–337. [Google Scholar] [CrossRef]
- Jiang, L.; Feng, J.X.; Ying, R.; Yin, F.M.; Pei, S.R.; Lu, J.G.; Cao, Y.T.; Guo, J.L.; Li, Z.F. Individual and combined effects of ammonia-N and sulfide on the immune function and intestinal microbiota of Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 92, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.F.; Dong, H.B.; Wang, Y.; Li, H.; Liu, Q.S.; Zhang, Y.; Zhang, J.S. Intestine oxidative stress and immune response to sulfide stress in Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2017, 63, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Bradford. A rapid and sensitive method for the quantitation of microgam quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zhong, L.; Yuan, L.; Rao, Y.; Li, Z.; Zhang, X.; Liao, T.; Xu, Y.; Dai, H.; Zhong, L.; Yuan, L. Distribution of vitellogenin in zebrafish (Danio rerio) tissues for biomarker analysis. Aquat. Toxicol. 2014, 149, 1–7. [Google Scholar] [CrossRef]
- Zhong, L.; Yuan, L.; Rao, Y.; Li, Z.; Gu, Q.; Long, Y.; Zhang, X.; Cui, Z.; Xu, Y.; Dai, H. Investigation of effect of 17α-ethinylestradiol on vigilin expression using an isolated recombinant antibody. Aquat. Toxicol. 2014, 156, 1–9. [Google Scholar] [CrossRef]
- Beliaeff, B.; Burgeot, T. Integrated biomarker response: A useful tool for ecological risk assessment. Environ. Toxicol. Chem. 2002, 21, 1316–1322. [Google Scholar] [CrossRef]
- Sanchez, W.; Burgeot, T.; Porcher, J.M. A novel “Integrated Biomarker Response” calculation based on reference deviation concept. Environ. Sci. Pollut. Res. 2013, 20, 2721–2725. [Google Scholar] [CrossRef]
- Jordan, J.; Galindo, M.F.; Calvo, S.; Gonzalez-Garcia, C.; Cena, V. Veratridine induces apoptotic death in bovine chromaffin cells through superoxide production. Br. J. Pharmacol. 2000, 130, 1496–1504. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Kim, C.Y.; Lee, J.; Lee, S.K.; Jeon, J.S. OsWRKY42 Represses OsMT1d and Induces Reactive Oxygen Species and Leaf Senescence in Rice. Mol. Cells 2014, 37, 532–539. [Google Scholar] [CrossRef]
- Kirschvink, N.; de Moffarts, B.; Lekeux, P. The oxidant/antioxidant equilibrium in horses. Vet. J. 2008, 177, 178–191. [Google Scholar] [CrossRef]
- van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.G.; Lam, J.C.W.; Tang, L.Z.; Hu, C.Y.; Liu, M.Y.; Lam, P.K.S.; Zhou, B.S. Probiotic Modulation of Lipid Metabolism Disorders Caused by Perfluorobutanesulfonate Pollution in Zebrafish. Environ. Sci. Technol. 2020, 54, 7494–7503. [Google Scholar] [CrossRef]
- Chen, L.G.; Lam, J.C.W.; Hu, C.Y.; Tsui, M.M.P.; Wang, Q.; Giesy, J.P.; Lam, P.K.S. Perfluorobutanesulfonate Exposure Causes Durable and Transgenerational Dysbiosis of Gut Microbiota in Marine Medaka. Environ. Sci. Tech. Lett. 2018, 5, 731–738. [Google Scholar] [CrossRef]
- Livingstone, D.R. Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organisms. Mar. Pollut. Bull. 2001, 42, 656–666. [Google Scholar] [CrossRef]
- Rubright, S.L.M.; Pearce, L.L.; Peterson, J. Environmental toxicology of hydrogen sulfide. Biol. Chem. 2017, 71, 1–13. [Google Scholar] [CrossRef]
- Fu, L.H.; Wei, Z.Z.; Hu, K.D.; Hu, L.Y.; Li, Y.H.; Chen, X.Y.; Han, Z.; Yao, G.F.; Zhang, H. Hydrogen sulfide inhibits the growth of Escherichia coli through oxidative damage. J. Microbiol. 2018, 56, 238–245. [Google Scholar] [CrossRef]
- Eghbal, M.A.; Pennefather, P.S.; O’Brien, P.J. H2S cytotoxicity mechanism involves reactive oxygen species formation and mitochondrial depolarisation. Toxicology 2004, 203, 69–76. [Google Scholar] [CrossRef]
- Tanaka, Y.; Hibino, T.; Hayashi, Y.; Tanaka, A.; Kishitani, S.; Takabe, T.; Yokota, S.; Takabe, T. Salt tolerance of transgenic rice overexpressing yeast mitochondrial Mn-SOD in chloroplasts. Plant Sci. 1999, 148, 131–138. [Google Scholar] [CrossRef]
- Li, Z.H.; Velisek, J.; Zlabek, V.; Grabic, R.; Machova, J.; Kolarova, J.; Li, P.; Randak, T. Chronic toxicity of verapamil on juvenile rainbow trout (Oncorhynchus mykiss): Effects on morphological indices, hematological parameters and antioxidant responses. J. Hazard. Mater. 2011, 185, 870–880. [Google Scholar] [CrossRef]
- Li, Z.H.; Li, P.; Shi, Z.C. Chronic Exposure to Tributyltin Induces Brain Functional Damage in Juvenile Common Carp (Cyprinus carpio). PLoS ONE 2015, 10, e0123091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.C.; Sharma, A.; Mishra, M.; Mishra, R.K.; Chowdhuri, D.K. Heat shock proteins in toxicology: How close and how far? Life Sci. 2010, 86, 377–384. [Google Scholar] [CrossRef]
- Padmini, E.; Rani, M.U. Seasonal influence on heat shock protein 90 alpha and heat shock factor 1 expression during oxidative stress in fish hepatocytes from polluted estuary. J. Exp. Mar. Biol. Ecol. 2009, 372, 1–8. [Google Scholar] [CrossRef]
- Richter, K.; Haslbeck, M.; Buchner, J. The Heat Shock Response: Life on the Verge of Death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef]
- Liu, Q.P.; Huang, S.T.; Deng, C.A.; Xiong, L.; Gao, X.; Chen, Y.; Niu, C.Q.; Liu, Y. Molecular characterization of heat-shock protein 90 gene and its expression in Gobiocypris rarus juveniles exposed to pentachlorophenol. Fish Physiol. Biochem. 2015, 41, 1279–1291. [Google Scholar] [CrossRef]
- Taylor, D.A.; Thompson, E.L.; Nair, S.V.; Raftos, D.A. Differential effects of metal contamination on the transcript expression of immune- and stress-response genes in the Sydney Rock oyster, Saccostrea glomerata. Environ. Pollut. 2013, 178, 65–71. [Google Scholar] [CrossRef]
- Werner, I.; Hinton, D.E. Field validation of hsp70 stress proteins as biomarkers in Asian clam (Potamocorbula amurensis): Is downregulation an indicator of stress? Biomarkers 1999, 4, 473–484. [Google Scholar]
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.M. Metallothionein: The multipurpose protein. Cell. Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef]
- Ruttkay-Nedecky, B.; Nejdl, L.; Gumulec, J.; Zitka, O.; Masarik, M. The Role of Metallothionein in Oxidative Stress. Int. J. Mol. Sci. 2013, 14, 6044–6066. [Google Scholar] [CrossRef] [Green Version]
- Cosson, R.P. Bivalve metallothionein as a biomarker of aquatic ecosystem pollution by trace metals: Limits and perspectives. Cell Mol. Biol. 2000, 46, 295–309. [Google Scholar]
- Falfushynska, H.; Horyn, O.; Fedoruk, O.; Khoma, V.; Rzymski, P. Difference in biochemical markers in the gibel carp (Carassius auratus gibelio) upstream and downstream of the hydropower plant. Environ. Pollut. 2019, 255, 113213. [Google Scholar] [CrossRef] [PubMed]
- Falfushynska, H.I.; Gnatyshyna, L.L.; Stoliar, O.B.; Nam, Y.K. Various responses to copper and manganese exposure of Carassius auratus gibelio from two populations. Comp. Biochem. Phys. C 2011, 154, 242–253. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence of Primer (5′-3′) | Product Length (bp) | Efficiency (%) | Gene Bank ID |
|---|---|---|---|---|
| β-actin-F | GAGGTATCCTGACCCTGAAGTA | 105 | 96.09 | XM_020621264 |
| β-actin-R | CGACTCTCAGCTCATTGTAGAAG | |||
| sod-F | CTCTCTCTCTCGGCACCTATTA | 133 | 104.6 | XM_020602040 |
| sod-R | GACTCAAGGCTACAAAGGTCAA | |||
| cat-F | CTGCACAGTGTCAGGTCTAAA | 111 | 92.11 | XM_020624985 |
| cat-R | TGACCTCTGAGACCCTCTATTC | |||
| mt-F | TACACACCACTGGCTCTTTG | 94 | 93.59 | XM_020604384 |
| mt-R | CGCAAGGGTCCATCTCTTT | |||
| hsp90-F | TAAGGAGGTTGAGGAGGATGAG | 95 | 95.36 | XM_020622865 |
| hsp90-R | CCTCAGCTGTGAAGTGGATATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, L.; Yao, F.; Zhang, H.; Xie, H.; Ru, H.; Wei, N.; Ni, Z.; Li, Z.; Li, Y. Sulfide Treatment Alters Antioxidant Response and Related Genes Expressions in Rice Field Eel (Monopterus albus). Water 2022, 14, 3230. https://doi.org/10.3390/w14203230
Zhong L, Yao F, Zhang H, Xie H, Ru H, Wei N, Ni Z, Li Z, Li Y. Sulfide Treatment Alters Antioxidant Response and Related Genes Expressions in Rice Field Eel (Monopterus albus). Water. 2022; 14(20):3230. https://doi.org/10.3390/w14203230
Chicago/Turabian StyleZhong, Liqiao, Fan Yao, He Zhang, Huaxiao Xie, Huijun Ru, Nian Wei, Zhaohui Ni, Zhong Li, and Yunfeng Li. 2022. "Sulfide Treatment Alters Antioxidant Response and Related Genes Expressions in Rice Field Eel (Monopterus albus)" Water 14, no. 20: 3230. https://doi.org/10.3390/w14203230