
Effect of Seawater and Surface-Sediment Variables on Epipelic Diatom Diversity and Abundance in the Coastal Area of Negeri Sembilan, Malaysia

Abstract
:1. Introduction
2. Materials and Methods
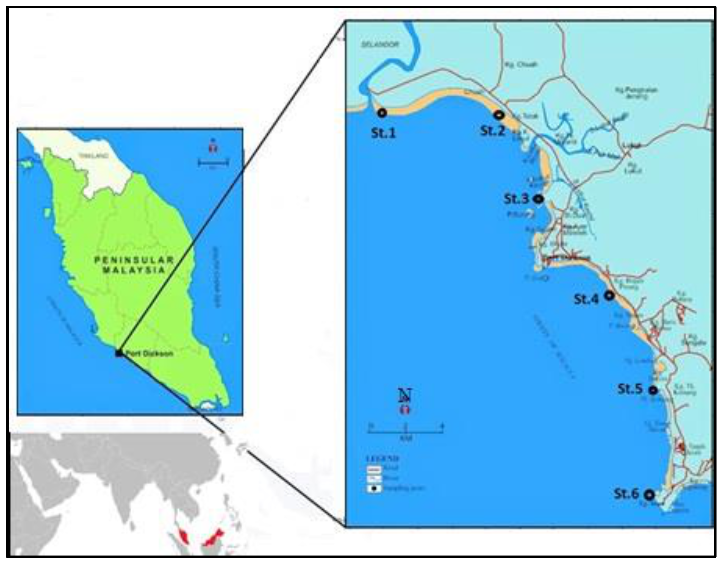
2.1. Study Area
2.2. Epipelic Diatom Extraction and Counting
2.3. Environmental Parameters
2.4. Data Analysis
3. Results
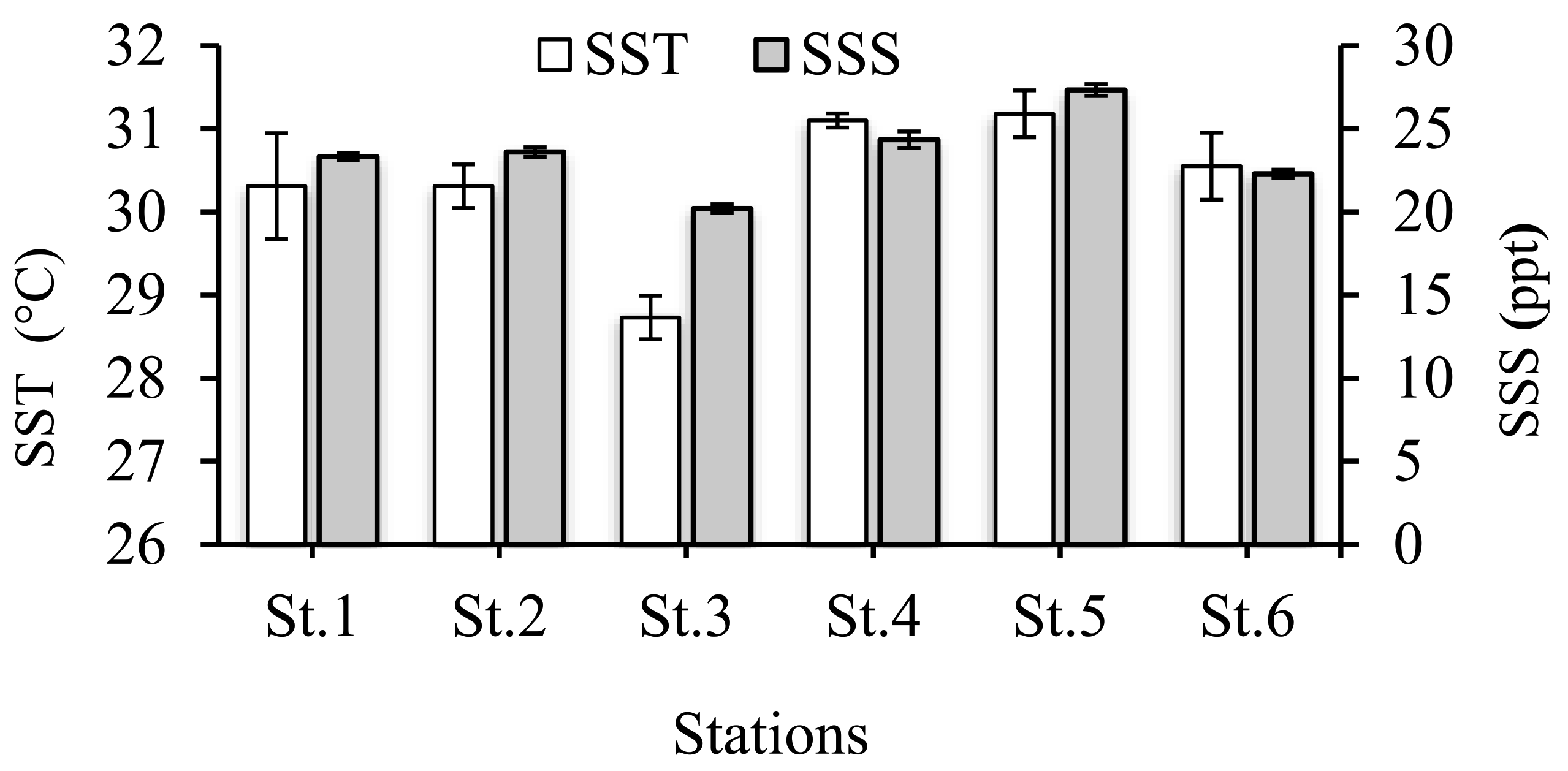
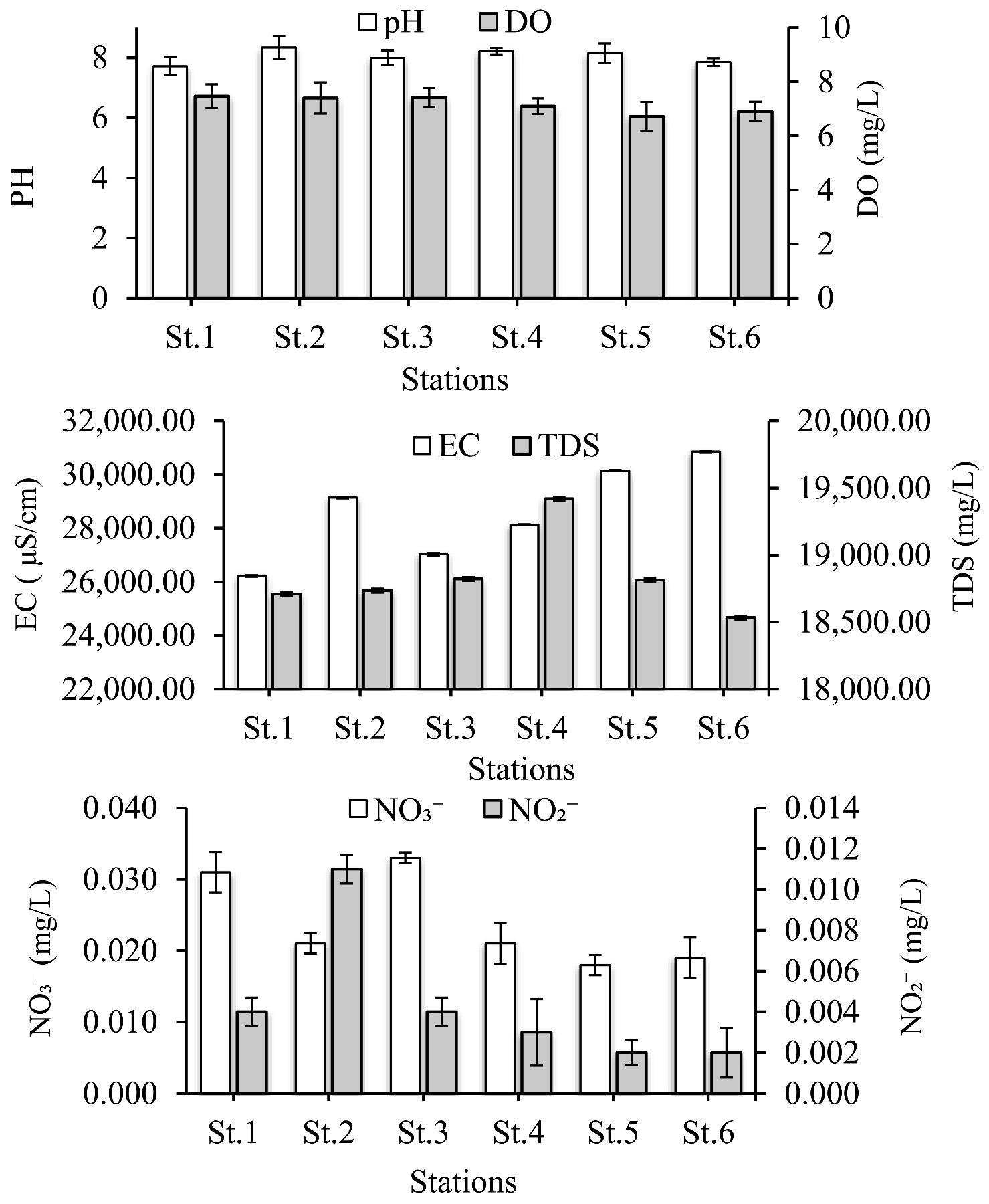
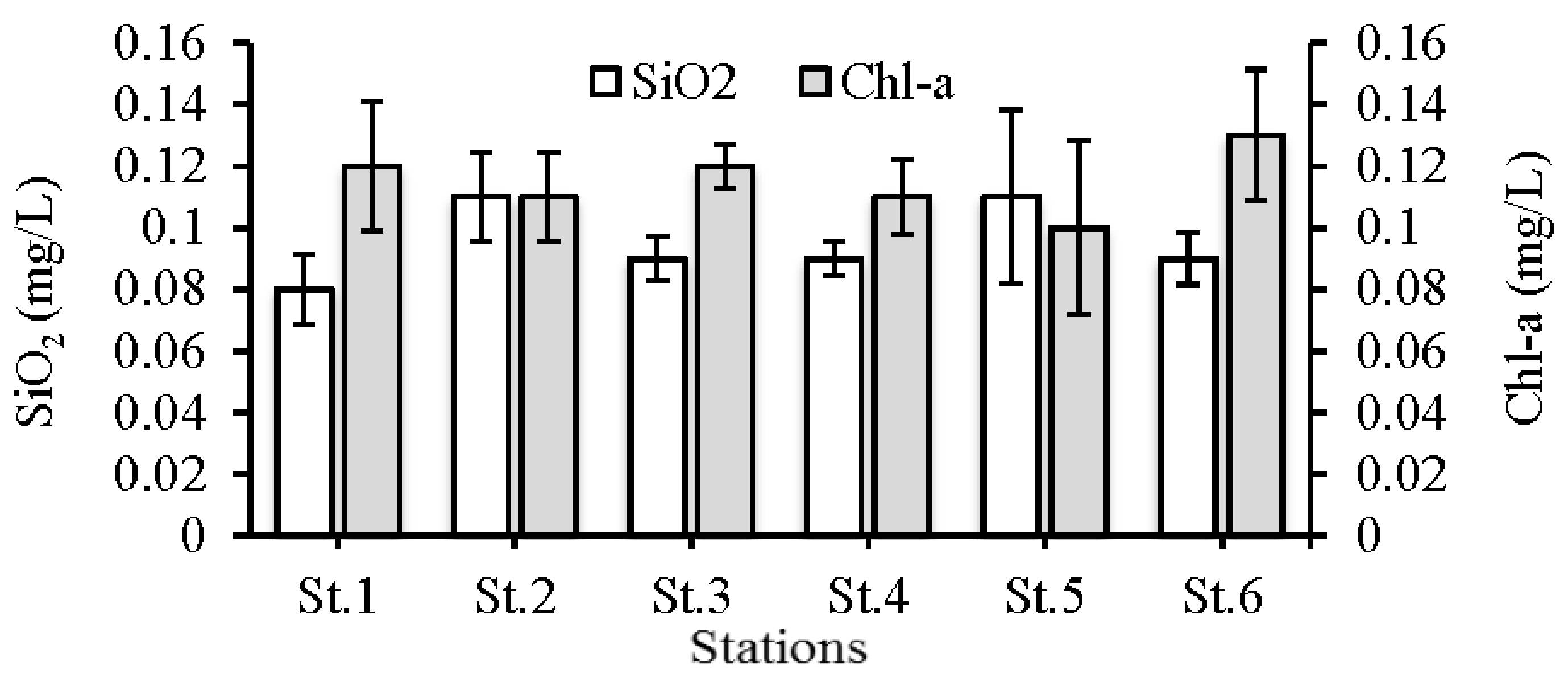
3.1. Environmental Conditions
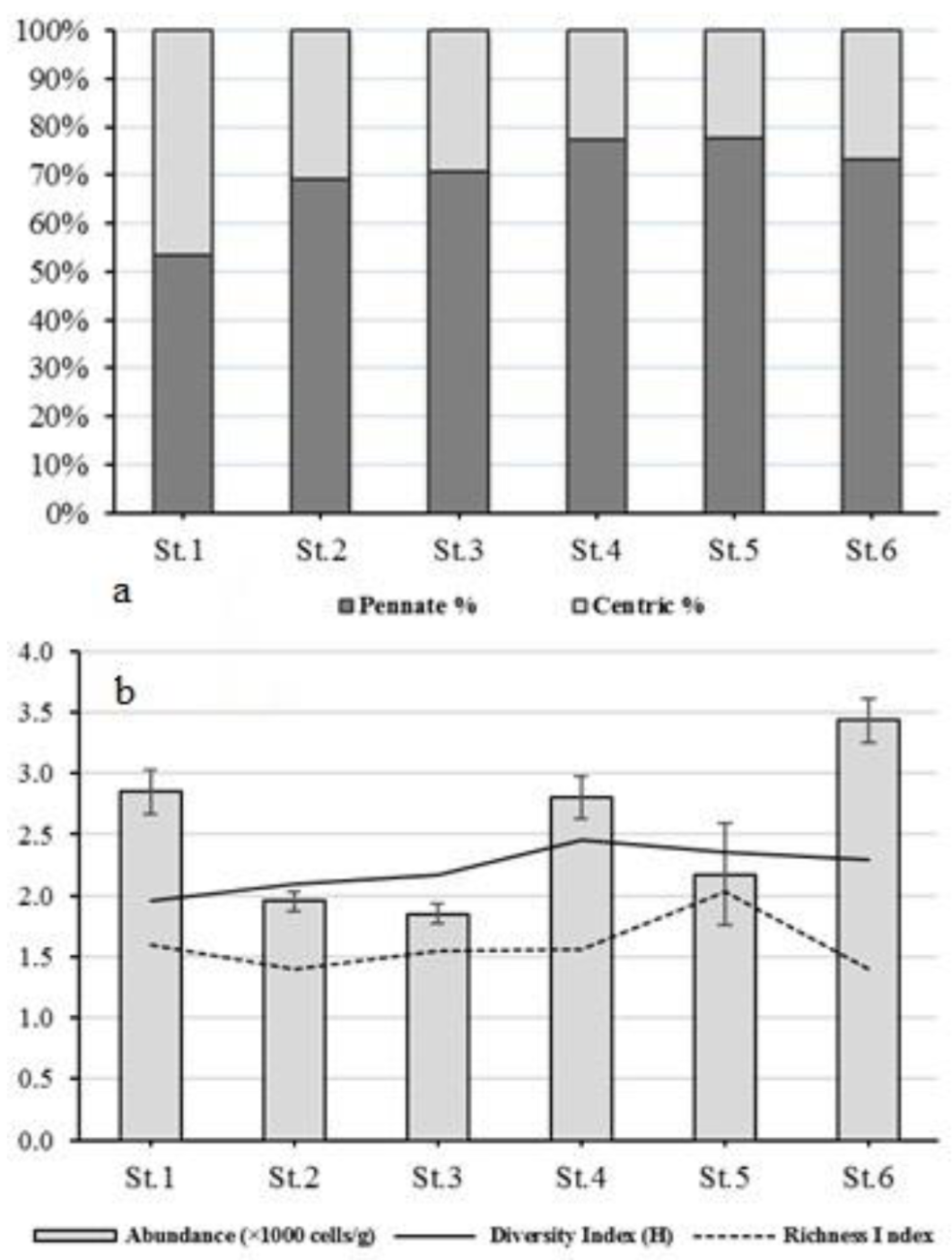
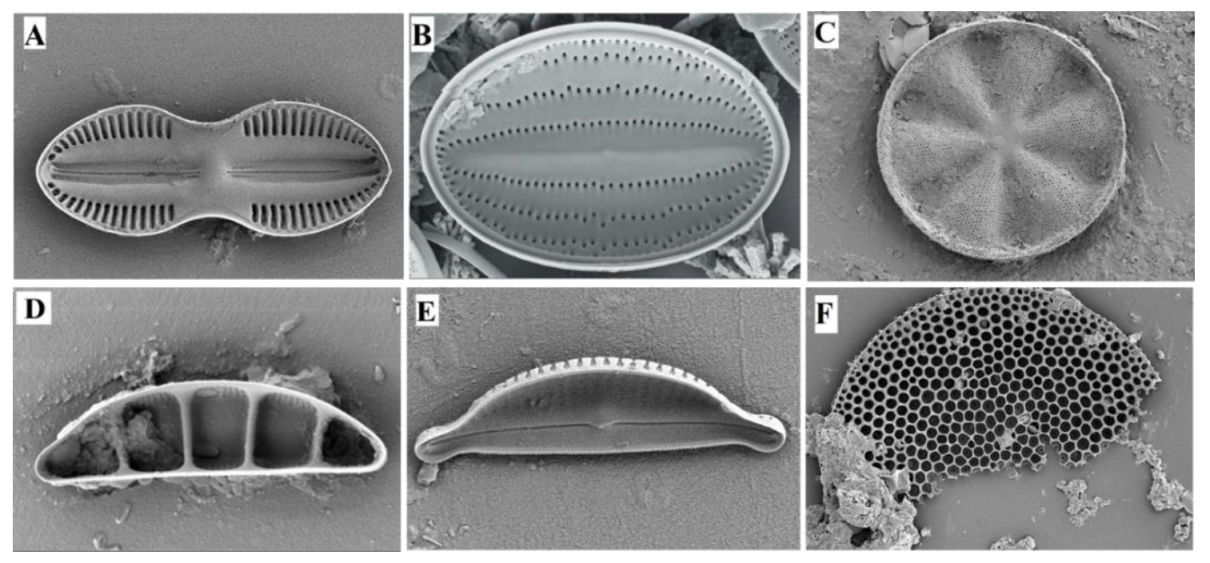
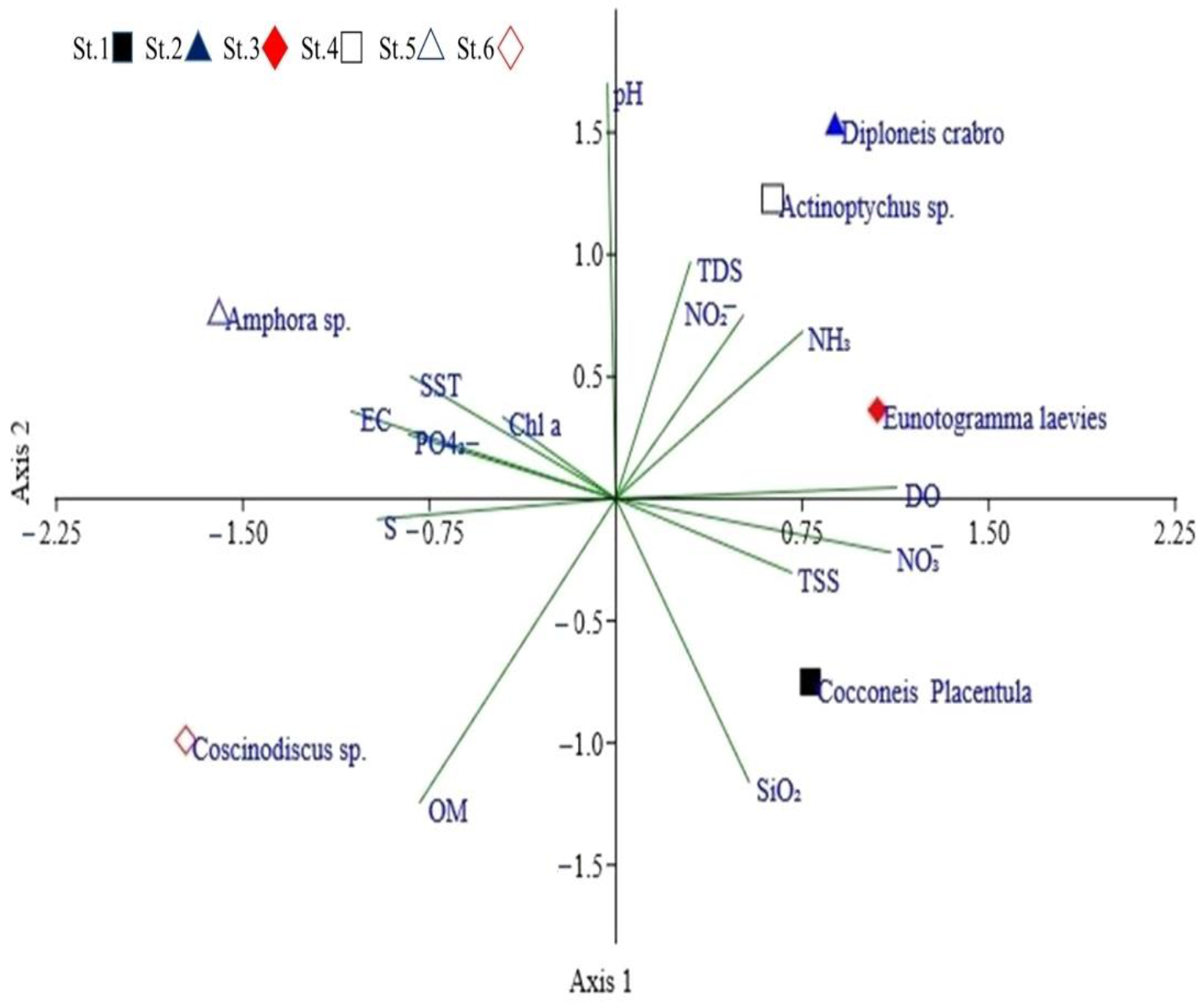
3.2. Dynamics of Epipelic Diatoms
4. Discussion
4.1. Environmental Conditions
4.2. Dynamics of Epipelic Diatoms
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ducklow, H.W.; Steinberg, D.K.; Buesseler, K.O. Upper ocean carbon export and the Biological Pump. Oceanography 2001, 14, 50–58. [Google Scholar] [CrossRef]
- Muller-Karger, F.E.; Varela, R.; Thunell, R.; Luerssen, R.; Hu, C.; Walsh, J.J. The importance of continental margins in the global carbon cycle. Geophys. Res. Lett. 2005, 32, 10–13. [Google Scholar] [CrossRef] [Green Version]
- Kasai, A.; Horie, H.; Sakamoto, W. Selection of food sources by Ruditapes philippinarum and Mactra veneriformis (Bivalva: Mollusca) determined from stable isotope analysis. Fish. Sci. 2004, 70, 11–20. [Google Scholar] [CrossRef]
- Sanger, D.; Blair, A.; Didonato, G.T.; Washburn, S.; Jones, G.; Riekerk, E.; Wirth, J.; Stewart, D.; White, L.; Vandiver, A.F. Impacts of coastal development on the ecology of tidal creek ecosystems of the US Southeast including consequences to humans. Estuaries Coasts 2015, 38, 49–66. [Google Scholar] [CrossRef] [PubMed]
- Vincent, F.; Bowler, C. Diatoms are selective segregators in global ocean planktonic communities. Msystems 2020, 5, e00444-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.S.; Lee, K.W.; Jung, S.W.; Lee, T.K.; Joo, H.M. Winter distribution of diatom assemblages along the coastline of Korea in 2020. Acta Oceanol. Sin. 2022, 6, 1–10. [Google Scholar]
- Kasim, M.; Mukai, H. Contribution of Benthic and Epiphytic Diatoms to Clam and Oyster Production in the Akkeshi-ko Estuary. J. Oceanogr. 2006, 62, 267–281. [Google Scholar] [CrossRef]
- Thornton, D.C.O.; Dong, L.F.; Underwood, G.J.C.; Nedwell, D.B. Factors affecting microphytobenthic biomass, species composition and production in the Colne estuary (UK). Aquat. Microb. Ecol. 2002, 27, 285–300. [Google Scholar] [CrossRef] [Green Version]
- Facca, C.; Sfriso, A. Epipelic diatom spatial and temporal distribution and relationship with the main environmental parameters in coastal waters. Estuar. Coast. Shelf Sci. 2007, 75, 135–149. [Google Scholar] [CrossRef]
- Cochero, J.; Licursi, M.; Gómez, N. Changes in the epipelic diatom assemblage in nutrient rich streams owing to the variations of simultaneous stressors. Limnologica 2015, 51, 15–23. [Google Scholar] [CrossRef]
- Méléder, V.; Rincé, Y.; Barillé, L.; Gaudin, P.; Rosa, P. Spatiotemporal changes in microphytobenthos assemblages in a macrotidal flat (Bourgneuf Bay, France). J. Phycol. 2007, 43, 1177–1190. [Google Scholar] [CrossRef]
- Ribeiro, L.; Brotas, V.; Rincé, Y.; Jesus, B. Structure and diversity of intertidal benthic diatom assemblages in contrasting shores: A case study from the Tagus estuary. J. Phycol. 2013, 49, 258–270. [Google Scholar] [CrossRef]
- Jabbari, M.; Salahi, M.; Ghorbani, R. Spatio-temporal influence of physicochemical parameters on phytoplankton assemblage in coastal brackish lagoon: Gomishan Lagoon, Caspian Sea, Iran. Biodiversitas 2018, 19, 2020–2027. [Google Scholar] [CrossRef]
- Vajravelu, M.; Martin, Y.; Ayyappan, S.; Mayakrishnan, M. Seasonal influence of physico-chemical parameters on phytoplankton diversity, community structure and abundance at Parangipettai coastal waters, Bay of Bengal, south east coast of India. Oceanologia 2018, 60, 114–127. [Google Scholar] [CrossRef]
- Zimba, P.V.; Hill, E.M.; Withers, K. Benthic microalgae serve as the major food resource for porcelain crabs (Petrolisthes spp.) in oyster reefs: Digestive track content and pigment evidence. J. Exp. Mar. Biol. Ecol. 2016, 483, 53–58. [Google Scholar] [CrossRef]
- McMinn, A.; Sellah, S.; Llah, W.A.W.; Mohammad, M.; Merican, F.M.S.; Omar, W.W.; Samad, F.; Cheah, W.; Idris, I.; Sim, Y.K.; et al. Quantum yield of the marine benthic microflora of near-shore coastal Penang, Malaysia. Mar. Freshw. Res. 2005, 7, 1047–1053. [Google Scholar] [CrossRef] [Green Version]
- Nursuhayati, A.S.; Yusoff, F.M.; Shariff, M. Spatial and temporal distribution of phytoplankton in Perak estuary, Malaysia, during monsoon season. J. Fish. Aquat. Sci. 2013, 8, 480–493. [Google Scholar] [CrossRef] [Green Version]
- Salleh, A.; Wakid, S.A.; Bahnan, I.S. Diversity of phytoplankton collected during the scientific expedition to Pulau Perak, Pulau Jarak and the Sembilan Group of Islands. Malays. J. Sci. 2008, 27, 33–45. [Google Scholar]
- Ke, Z.; Tan, Y.; Huang, L. Spatial variation of phytoplankton community from Malacca Strait to southern South China Sea in May of 2011. Acta Ecol. Sin. 2016, 36, 154–159. [Google Scholar] [CrossRef]
- Siswanto, E.; Tanaka, K. Phytoplankton biomass dynamics in the Strait of Malacca within the period of the Sea WiFS full mission: Seasonal cycles, interannual variations and decadal-scale trends. Remote Sens. 2012, 6, 2718–2742. [Google Scholar] [CrossRef] [Green Version]
- Salleh, A.; Wakid, S.A.; Bahnan, I.S.; Rahman, K.A.A.; Nasrodin, S. Diversity of phytoplankton at Langkawi Island, Malaysia. Malays. J. Sci. 2005, 24, 43–55. [Google Scholar]
- Salleh, A.; Ruslan, N.D. Phytoplankton composition and distribution in the Coastal Area of Bachok Kelantan. Malays. J. Sci. 2010, 29, 19–29. [Google Scholar]
- Lim, H.C.; Teng, S.T.; Leaw, C.P.; Wataki, M.; Lim, P. Phytoplankton assemblage of the Merambong Shoal, Tebrau Straits with note on potentially harmful species. Malay Nat. J. 2014, 66, 198–221. [Google Scholar]
- Joon, H.L.; Choon, W.L. Short-timescale Variation of Phytoplankton Abundance and Diversity at Redang Island. Malays. J. Sci. 2015, 34, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.H.; Lee, C.W. Effects of eutrophication on diatom abundance, biovolume and diversity in tropical coastal waters. Environ. Monit. Assess. 2017, 189, 1–10. [Google Scholar] [CrossRef]
- Desianti, N.; Potapova, M.; Enache, M.; Belton, T.J.; Velinsk, D.J.; Thomas, R.; Mead, J. Sediment diatoms as environmental indicators in New Jersey coastal lagoons. J. Coast. Res. 2017, 78, 127–140. [Google Scholar] [CrossRef]
- Sha, L.; Jiang, H.; Knudsen, K.L. Diatom evidence of climatic change in Holsteinsborg Dyb, west of Greenland, during the last 1200 years. Holocene 2012, 22, 347–358. [Google Scholar] [CrossRef]
- Benito, X.; Trobajo, R.; Ibáñez, C. Benthic diatoms in a Mediterranean delta: Ecological indicators and a conductivity transfer function for paleoenvironmental studies. J. Paleolimnol. 2015, 2, 171–188. [Google Scholar] [CrossRef]
- Kobayasi, H.; Idei, M.; Mayama, S.; Nagumo, T.; Osada, K.; Kobayasi’s, H. Atlas of Japanese Diatoms based on Electron Microscopy, 1st ed.; Uchida Rokakuho Publishing Co., Ltd.: Tokyo, Japan, 2006; p. 531. [Google Scholar]
- Giffen, M.H. Contributions to the diatom flora of South Africa IV. Nova Hedwig. Beih. 1963, 31, 259–312. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Daiber, F.C. Light, nitrogen, and phosphorus limitation of edaphic algae in a Delaware salt marsh. J. Exp. Mar. Biol. Ecol. 1975, 18, 79–88. [Google Scholar] [CrossRef]
- Washington, H.G. Diversity, biotic and similarity indices: A review with special relevance to aquatic ecosystems. Water Res. 1984, 18, 653–694. [Google Scholar] [CrossRef]
- Margalef, R. Temporal succession and spatial heterogeneity in phytoplankton. In Perspectives in Marine Biology; Buzzati-Traverso, A.A., Ed.; University of California Press: Oakland, CA, USA, 1958; pp. 323–349. [Google Scholar]
- Lim, J.H.; Wong, Y.Y.; Lee, C.W.; Bong, C.W.; Kudo, I. Long-term comparison of dissolved nitrogen species in tropical estuarine and coastal water systems. Estuar. Coast. Shelf Sci. 2019, 222, 103–111. [Google Scholar] [CrossRef]
- Mantoura, R.F.C.; Llewellyn, C.A. The rapid determination of algal chlorophyll and carotenoid pigments and their breakdown products in natural waters by reverse-phase high-performance liquid chromatography. Anal. Chim. Acta 1983, 151, 297–314. [Google Scholar] [CrossRef]
- D.O.E. (Department of Environment Malaysia). Malaysian Marine Water Quality Standards Report 2017; DOE: Putrajaya, Malaysia, 2017; pp. 1–135. [Google Scholar]
- Heiri, O.; Lotter, A.F.; Lemcke, G. Loss on ignition as a method for estimating organic and carbonate content in sediments: Reproducibility and comparability of results. J. Paleolimnol. 2000, 25, 101–110. [Google Scholar] [CrossRef]
- Lee, C.W.; Bong, C.W. Bacterial abundance and production and their relation to primary production in tropical coastal waters of Peninsular Malaysia. Mar. Freshw. Res. 2008, 59, 10–21. [Google Scholar] [CrossRef]
- Yun, M.S.; Lee, S.H.; Chung, I.K. Photosynthetic activity of benthic diatoms in response to different temperatures. J. Appl. Phycol. 2010, 22, 559–562. [Google Scholar] [CrossRef]
- Dalu, T.; Richoux, N.B.; Froneman, P.W. Distribution of benthic diatom communities in a permanently open temperate estuary in relation to physico-chemical variables. S. Afr. J. Bot. 2016, 107, 31–38. [Google Scholar] [CrossRef]
- Bagazi, Z.A.; Baakdah, M.A.; Affan, M.A. Seasonal dynamics and diversity of phytoplankton in Sharm Yanbu Lagoon, Red Sea, Saudi Arabia. J. King Abdulaziz Univ. Mar. Sci. 2018, 28, 9–26. [Google Scholar]
- Hamzah, A.; Kipli, S.H.; Ismail, S.R.; Una, R.; Sarmani, S. Microbiological study in coastal water of Port Dickson, Malaysia. Sains Malays. 2011, 40, 93–99. [Google Scholar]
- Park, G.S. The role and distribution of total suspended solids in the macrotidal coastal waters of Korea. Environ. Monit. Assess. 2007, 135, 153–162. [Google Scholar] [CrossRef]
- Liu, Q.; Liang, Y.; Cai, W.J.; Wang, K.; Wang, J.; Yin, K. Changing riverine organic C: N ratios along the Pearl River: Implications for estuarine and coastal carbon cycles. Sci. Total Environ. 2020, 709, 136052. [Google Scholar] [CrossRef] [PubMed]
- Chaikaew, P.; Chavanic, S. Spatial Variability and Relationship of Mangrove Soil Organic Matter to Organic Carbon. Appl. Environ. Soil Sci. 2017, 20, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Litchman, E. Resource Competition and The Ecological Success of Phytoplankton. In Evolution of Primary Producers in the Sea, 1st ed.; Falkowski, P.G., Knoll, A.H., Eds.; Elsevier Science Publishing Co. Inc.: San Diego, CA, USA, 2007; pp. 351–375. [Google Scholar]
- Praveena, S.M.; Aris, A.Z. A baseline study of tropical coastal water quality in Port Dickson, Strait of Malacca, Malaysia. Mar. Pollut. Bull. 2013, 67, 196–199. [Google Scholar] [CrossRef] [PubMed]
- Shaari, F.; Mustapha, M.A. Factors influencing the distribution of Chl-a along coastal waters of east Peninsular Malaysia. Sains Malays. 2017, 46, 1191–1200. [Google Scholar] [CrossRef]
- Leleyter, L.; Baraud, F.; Gil, O.; Gouali, S.; Lemoine, M.; Orvain, F. Aluminium Impact on The Growth of Benthic Diatom. In Marine Sediments: Formation, Distribution and Environmental Impacts, 1st ed.; Williams, W., Ed.; Novinka: New York, NY, USA, 2016; pp. 1–19. [Google Scholar]
- Agatz, M.; Asmus, R.M.; Deventer, B. Structural changes in the benthic diatom community along a eutrophication gradient on a tidal flat. Helgol. Mar. Res. 1999, 53, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Bilcke, G.; Van Craenenbroeck, L.; Castagna, A.; Osuna-Cruz, C.M.; Vandepoele, K.; Sabbe, K.; DeVeylder, L.; Vyverman, W. Light intensity and spectral composition drive reproductive success in the marine benthic diatom Seminavis robusta. Sci. Rep. 2021, 1, 17560. [Google Scholar]
- Balasubramaniam, J.; Prasath, D.; Jayaraj, K.A. Microphytobenthic biomass, species composition and environmental gradients in the mangrove intertidal region of the Andaman Archipelago, India. Environ. Monit. Assess. 2017, 189, 1–9. [Google Scholar] [CrossRef]
- Dodds, W.K.; Smith, V.H.; Lohman, K. Nitrogen and phosphorus relationships to benthic algal biomass in temperate streams. Can. J. Fish Aquat. Sci. 2002, 59, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Cartaxana, P.; Vieira, S.; Ribeiro, L.; Rocha, R.J.; Cruz, S.; Calado, R.; da Silva, J.M. Effects of elevated temperature and CO2 on intertidal microphytobenthos. BMC Ecol. 2015, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Admiraal, W.; Peletier, H. Influence of organic compounds and light limitation on the growth rate of estuarine benthic diatoms. Brit. Phycol. J. 1979, 14, 197–206. [Google Scholar] [CrossRef]
- Virta, L.; Gammal, J.; Järnström, M.; Bernard, G.; Soininen, J.; Norkko, J.; Norkko, A. The diversity of benthic diatoms affects ecosystem productivity in heterogeneous coastal environments. Ecology 2019, 100, 1–11. [Google Scholar] [CrossRef]
- Jézéquel, V.M.; Hildebrand, M.; Brzezinski, M.A. Silicon metabolism in diatoms: Implications for growth. J. Phycol. 2000, 36, 821–840. [Google Scholar] [CrossRef]
- Domingues, R.B.; Barbosa, A.B.; Sommer, U.; Galvão, H.M. Ammonium, nitrate and phytoplankton interactions in a freshwater tidal estuarine zone: Potential effects of cultural eutrophication. Aquat. Sci. 2011, 73, 331–343. [Google Scholar] [CrossRef]
- Underwood, G.; Phillips, J.; Saunders, K. Distribution of estuarine benthic diatom species along salinity and nutrient gradients. Eur. J Phycol. 1998, 33, 173–183. [Google Scholar] [CrossRef]
- Round, F.E.; Round, R.M.; Mann, D.G. The Diatoms, Biology and Morphology of the Genera, 1st ed.; Cambridge University Press: New York, NY, USA, 1990; pp. 1–744. [Google Scholar]
- Vrieling, E.G.; Poort, L.; Beelen, T.P.; Giesked, W.W. Growth and silica content of the diatoms Thalassiosira weissflogii and Navicula salinarum at different salinities and enrichments with aluminium. Eur. J. Phycol. 1999, 34, 307–316. [Google Scholar] [CrossRef]
- Leterme, S.C.; Prime, E.; Mitchell, J.; Brown, M.H.; Ellis, A.V. Diatom adaptability to environmental change: A case study of two Cocconeis species from high-salinity areas. Diatom Res. 2013, 28, 29–35. [Google Scholar] [CrossRef]
- Gnanamoorthy, P.; Karthikeyan, V.; Prabu, V.A. Field emission scanning electron microscopy (FESEM) characterisation of the porous silica nanoparticulate structure of marine diatoms. J. Porous Mat. 2014, 21, 225–233. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| St.1 | St.2 | St.3 | St.4 | St.5 | St.6 | %Pr | Class | |
|---|---|---|---|---|---|---|---|---|
| Actinoptychus sp. | 9 | 3 | 15 | 50 | C | |||
| A. undulates (J.W.Bailey) Ralfs, 1861. | 3 | 3 | 5 | 50 | C | |||
| Amphora arenaria Donkin, 1858. | 13 | 8 | 4 | 50 | C | |||
| Amphora sp. | 2 | 5 | 28 | 50 | C | |||
| Auliscus elegans Auliscus elegans var. californica (Grunow in Schmidt et al.) Rattray, 1888. | 4 | 5 | 8 | 50 | C | |||
| Caloneis sp. | 8 | 3 | 33 | R | ||||
| Campylodiscus sp. | 7 | 3 | 6 | 50 | C | |||
| Cocconeis placentula Ehrenberg, 1838. | 41 | 14 | 5 | 1 | 67 | F | ||
| C. radiatus Ehrenberg, 1840. | 12 | 17 | X | |||||
| C. gigas var. praetexta (Janisch) Hustedt, 1930. | 7 | 9 | 11 | 50 | C | |||
| Coscinodiscus sp. | 6 | 15 | 33 | R | ||||
| Cyclotella striata Grunow in Van Heurck, 1882. | 6 | 8 | 13 | 50 | C | |||
| Diploneis crabro Ehrenberg, 1854. | 24 | 5 | 2 | 50 | C | |||
| Diploneis obliqua (Brun) Hustedt, 1937. | 11 | 17 | X | |||||
| Eunotogramma laevis Grunow, 1883. | 34 | 5 | 6 | 50 | C | |||
| Eolimna minima (Grunow) Lange-Bertalot & W.Schiller, 1997. | 5 | 6 | 8 | 7 | 67 | F | ||
| Gyrosigma eximium (Thwaites) Boyer, 1927. | 3 | 6 | 33 | R | ||||
| Lyrella clavata (Gregory) D.G.Mann, 1990. | 10 | 17 | X | |||||
| Lyrella sp. | 5 | 2 | 33 | R | ||||
| Mastogloia angulata Lewis, 1861. | 5 | 6 | 4 | 50 | C | |||
| Melosira sp. | 3 | 3 | 11 | 50 | C | |||
| Navicula sp. | 4 | 17 | X | |||||
| N. longa (Gregory) Ralfs ex Pritchard, 1861. | 9 | 17 | X | |||||
| N. peregrine | 4 | 4 | 33 | R | ||||
| Nitzschia sigma (Hantzsch) Grunow, 1878. | 7 | 17 | X | |||||
| Odontella sp. | 5 | 10 | 33 | R | ||||
| O. mobiliensis (J.W.Bailey) Grunow, 1884. | 3 | 17 | X | |||||
| Paralia sulcate (Ehrenberg) Cleve, 1873. | 5 | 1 | 33 | R | ||||
| Petroneis granulate (Bailey) D.G.Mann, 1990. | 4 | 8 | 1 | 50 | C | |||
| Pinnularia sp. | 7 | 17 | X | |||||
| P. aestuarii Cleve, 1895. | 3 | 17 | X | |||||
| Pleurosigma sp. | 10 | 4 | 33 | R | ||||
| P. naviculaceum Brébisson, 1854 | 4 | 4 | 17 | X | ||||
| pseudo-nitzschia sp. | 2 | 17 | X | |||||
| Surirella sp. | 7 | 17 | X | |||||
| S. fastuosa (Ehrenberg) Ehrenberg, 1843. | 6 | 17 | X | |||||
| S. spiralis Kützing, 1844 | 2 | 17 | X | |||||
| Thalassiosira sp. | 3 | 8 | 8 | 50 | C | |||
| Triceratium sp. | 13 | 17 | X |
| Physicochemical Variables | Abundances of Epipelic Diatom Communities (cells/g) | Diversity of Epipelic Diatom Communities (H’) | ||
|---|---|---|---|---|
| r | p Value | r | p Value | |
| SST (°C) | 0.63 | 0.03 * | 0.58 | 0.04 * |
| pH | −0.58 | 0.04 * | 0.43 | 0.16 |
| TSS (mg/L) | 0.53 | 0.08 * | −0.85 | <0.01 * |
| NH3; (mg/L) | −0.67 | 0.02 * | 0.07 | 0.83 |
| SiO2 (mg/L) | 0.36 | 0.25 | −0.63 | 0.03 * |
| Chl-a (mg/L) | 0.74 | <0.01 * | −0.11 | 0.73 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sas, A.A.; Suriyanti, S.N.P.; Das, S.K.; Cob, Z.C. Effect of Seawater and Surface-Sediment Variables on Epipelic Diatom Diversity and Abundance in the Coastal Area of Negeri Sembilan, Malaysia. Water 2022, 14, 3187. https://doi.org/10.3390/w14193187
Sas AA, Suriyanti SNP, Das SK, Cob ZC. Effect of Seawater and Surface-Sediment Variables on Epipelic Diatom Diversity and Abundance in the Coastal Area of Negeri Sembilan, Malaysia. Water. 2022; 14(19):3187. https://doi.org/10.3390/w14193187
Chicago/Turabian StyleSas, Ahmed Awadh, Su Nyun Pau Suriyanti, Simon Kumar Das, and Zaidi Che Cob. 2022. "Effect of Seawater and Surface-Sediment Variables on Epipelic Diatom Diversity and Abundance in the Coastal Area of Negeri Sembilan, Malaysia" Water 14, no. 19: 3187. https://doi.org/10.3390/w14193187