
The Influence of Distinct Seasons on the Succession and Diversity of Bacteria on the Anticorrosive Coatings Surfaces in a Marine Environment
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling
2.2. Samples Collection and DNA Extraction
2.3. Processing Sequencing Data
2.4. Bioinformatics Analysis
2.5. Statistical Analysis
3. Results
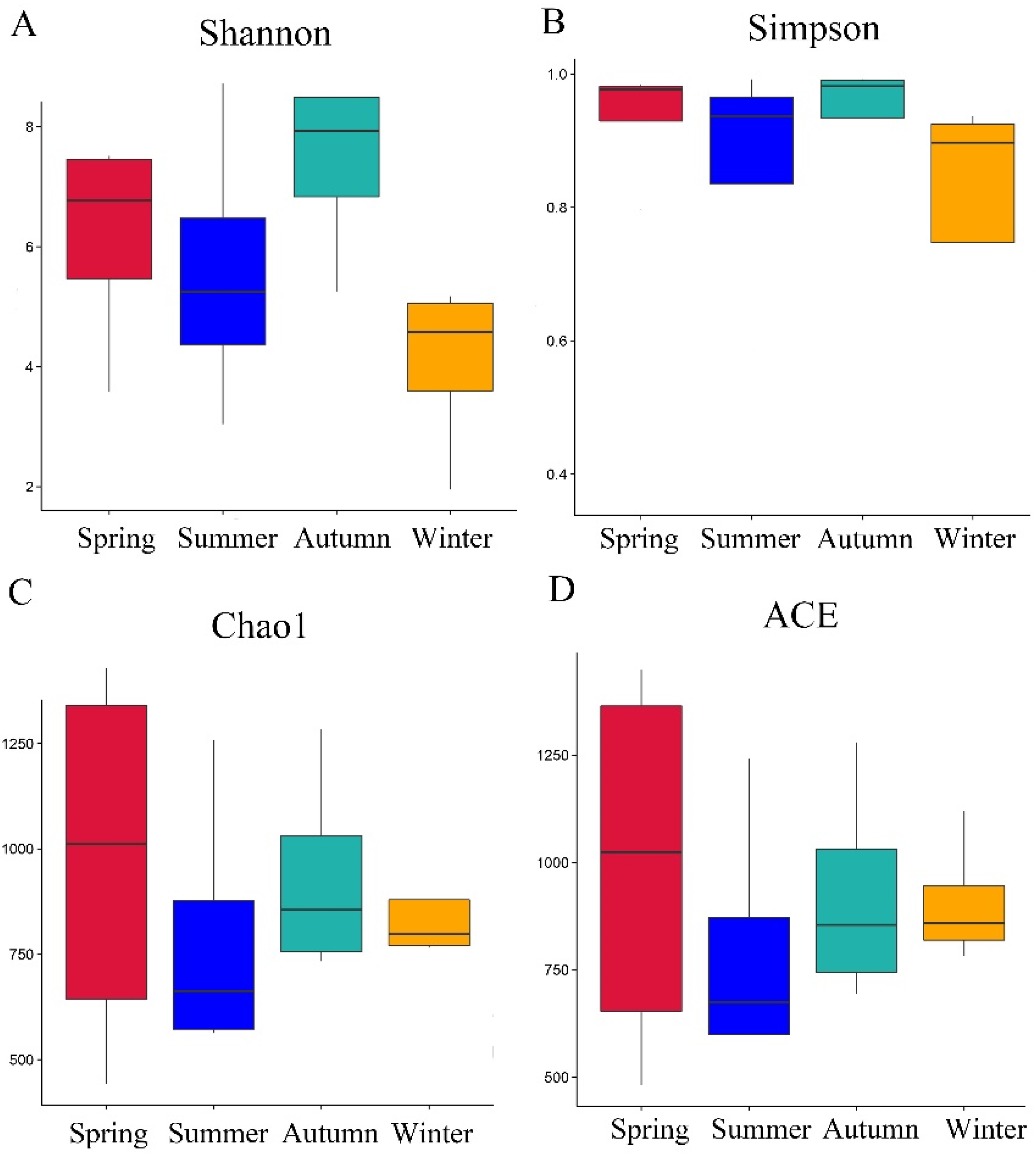
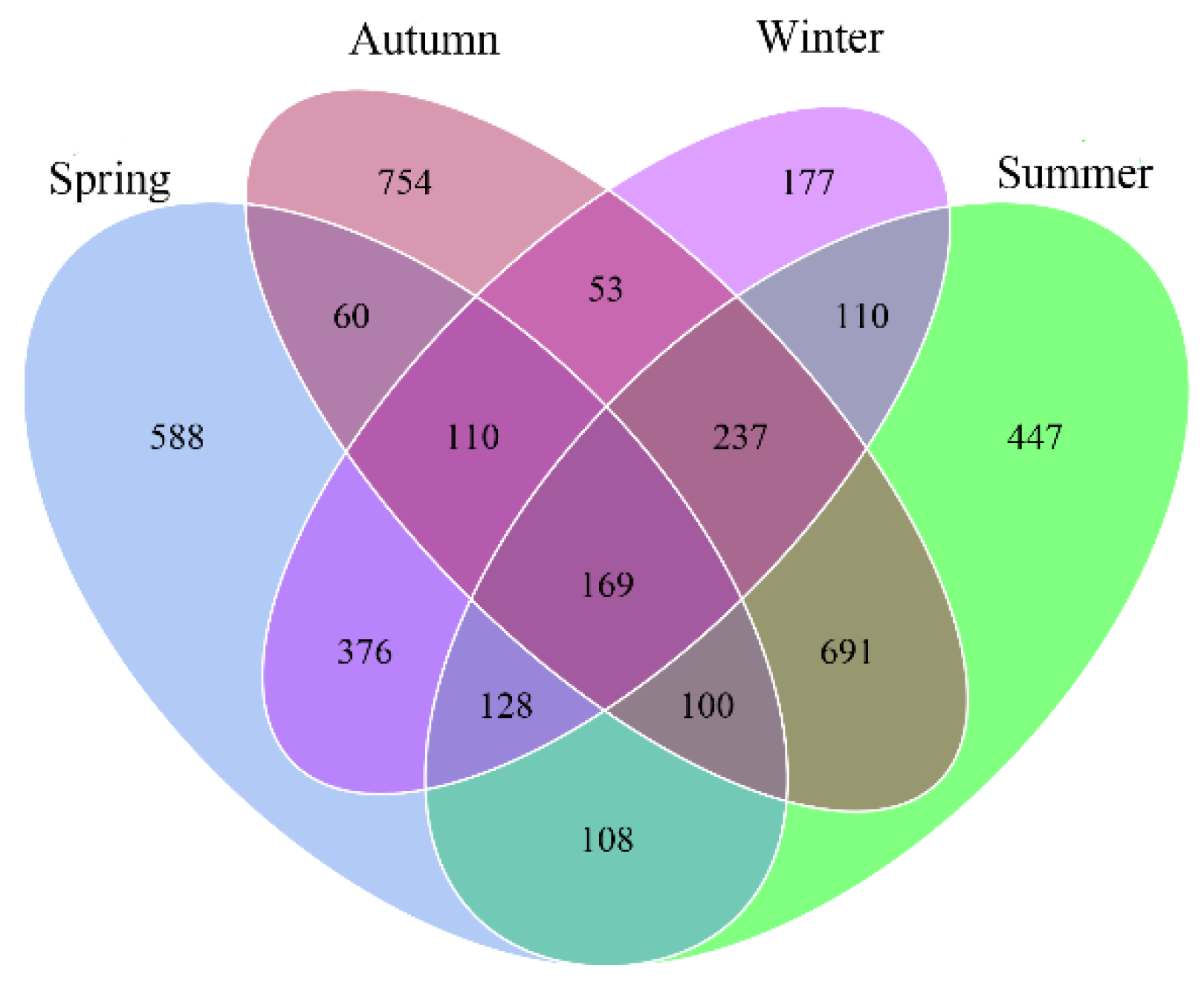
3.1. Diversity Analysis of Bacteria in Different Seasons
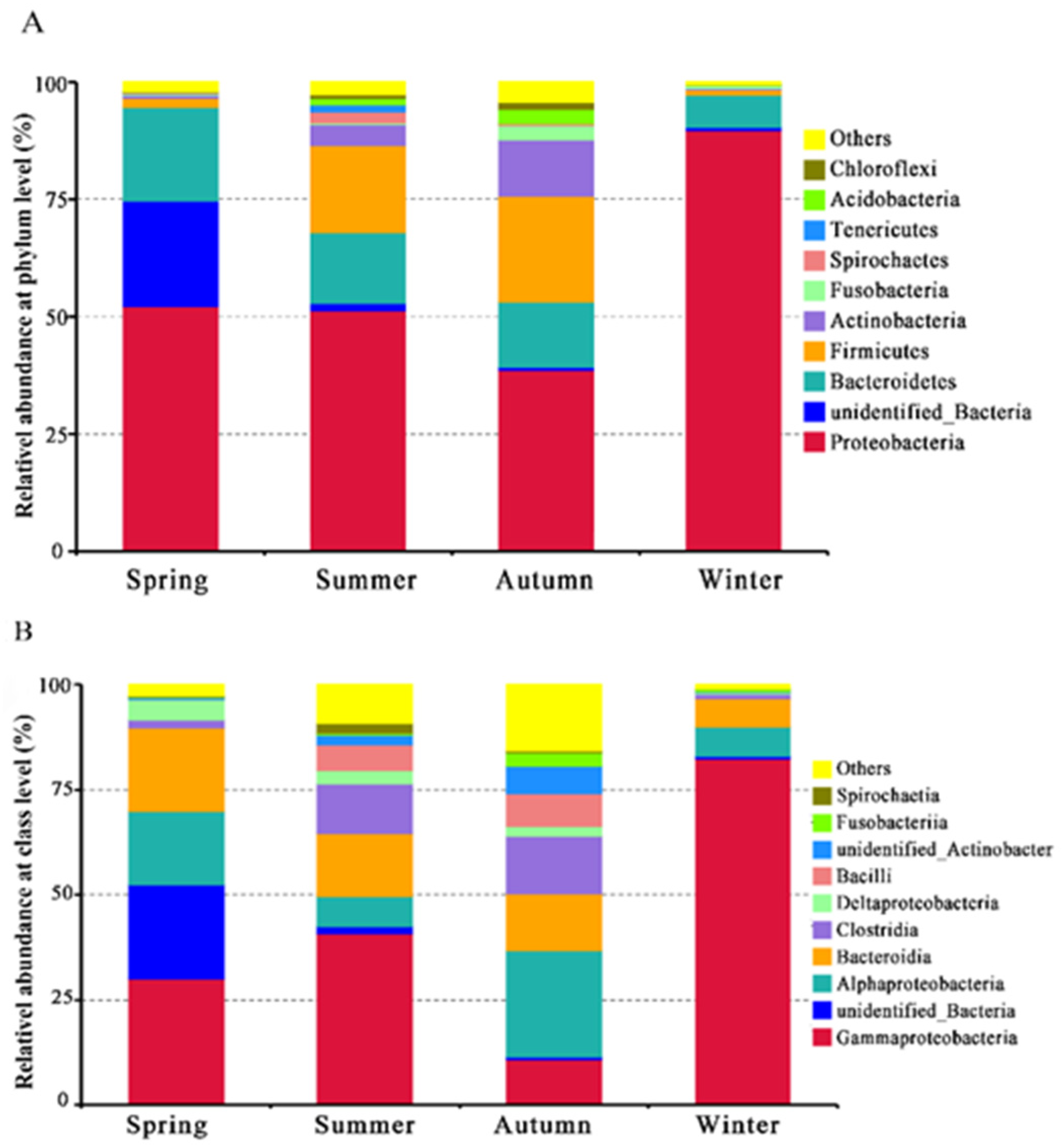
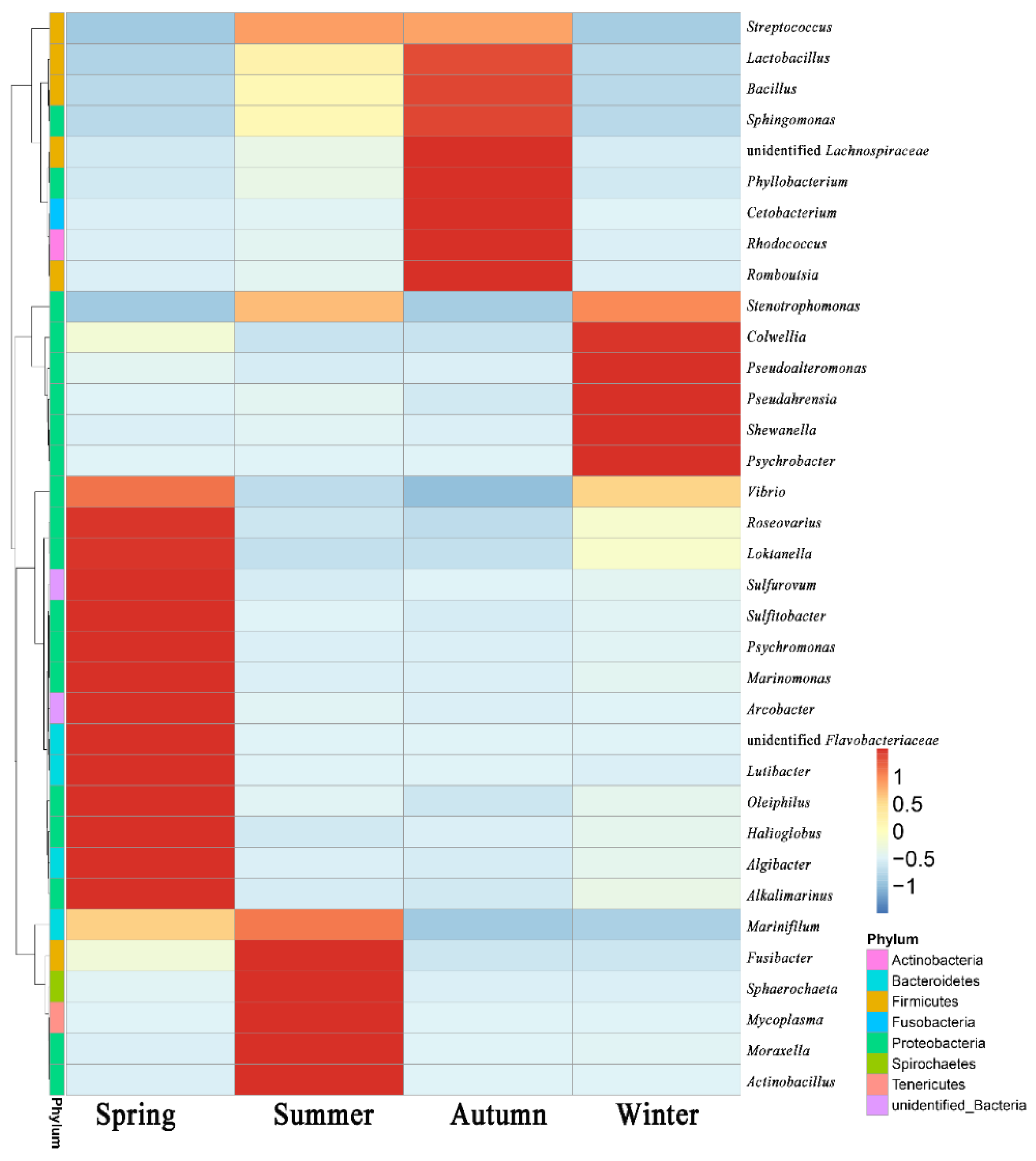
3.2. Community Diversity Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hou, B.; Li, X.; Ma, X.; Du, C.; Zhang, D.; Zheng, M.; Xu, W.; Lu, D.; Ma, F. The cost of corrosion in China. Npj Mater. Degrad. 2017, 1, 4. [Google Scholar] [CrossRef]
- Enning, D.; Garrelfs, J. Corrosion of iron by sulfate-reducing bacteria: New views of an old problem. Appl. Environ. Microbiol. 2014, 80, 1226–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capao, A.; Moreira-Filho, P.; Garcia, M.; Bitati, S.; Procopio, L. Marine bacterial community analysis on 316L stainless steel coupons by Illumina MiSeq sequencing. Biotechnol. Lett. 2020, 42, 1431–1448. [Google Scholar] [CrossRef] [PubMed]
- Procopio, L. Changes in microbial community in the presence of oil and chemical dispersant and their effects on the corrosion of API 5L steel coupons in a marine-simulated microcosm. Appl. Microbiol. Biotechnol. 2020, 104, 6397–6411. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, Y.; Zhang, R.; Guan, F.; Hou, B.; Duan, J. Microbiologically influenced corrosion of marine steels within the interaction between steel and biofilms: A brief view. Appl. Microbiol. Biotechnol. 2020, 104, 515–525. [Google Scholar] [CrossRef]
- Li, K.; Whitfield, M.; Van Vliet, K.J. Beating the bugs: Roles of microbial biofilms in corrosion. Corros. Rev. 2013, 31, 73–84. [Google Scholar] [CrossRef]
- Moura, V.; Ribeiro, I.; Moriggi, P.; Capao, A.; Salles, C.; Bitati, S.; Procopio, L. The influence of surface microbial diversity and succession on microbiologically influenced corrosion of steel in a simulated marine environment. Arch. Microbiol. 2018, 200, 1447–1456. [Google Scholar] [CrossRef]
- Zuo, R. Biofilms: Strategies for metal corrosion inhibition employing microorganisms. Appl. Microbiol. Biotechnol. 2007, 76, 1245–1253. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, H.; Karn, S.K.; Duan, J.; Guan, F.; Zhai, X.; Zhao, S.; Li, K.; Hou, B. Corrosion behavior of Q235 carbon steel in presence of H2S producing Bacillus sp. and a consortium of microbes isolated from inner rust layer. Int. J. Electrochem. Sci. 2017, 12, 2315–2328. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, Y.; Duan, J.; Li, X.; Wang, J.; Hou, B. Analysis of marine microbial communities colonizing various metallic materials and rust layers. Biofouling 2019, 35, 429–442. [Google Scholar] [CrossRef]
- Li, X.; Xiao, H.; Zhang, W.; Li, Y.; Tang, X.; Duan, J.; Yang, Z.; Wang, J.; Guan, F.; Ding, G. Analysis of cultivable aerobic bacterial community composition and screening for facultative sulfate-reducing bacteria in marine corrosive steel. J. Oceanol. Limnol. 2018, 37, 600–614. [Google Scholar] [CrossRef]
- Saravanan, P.; Duraibabu, D.; Jayamoorthy, K.; Suresh, S.; Kumar, S.A. Twin applications of tetra-functional epoxy monomers for anticorrosion and antifouling studies. Silicon 2018, 10, 555–565. [Google Scholar] [CrossRef]
- Quan, X.; Wang, J.; Zhao, S.; Cai, W.; Wang, Z.; Wang, S.; Cui, X. Improved antibacterial, antifouling and corrosion protective performance of epoxy coatings with poly(m-aminophenol). Prog. Org. Coat. 2018, 115, 9–17. [Google Scholar] [CrossRef]
- Vermeirssen, E.L.M.; Dietschweiler, C.; Werner, I.; Burkhardt, M. Corrosion protection products as a source of bisphenol A and toxicity to the aquatic environment. Water. Res. 2017, 123, 586–593. [Google Scholar] [CrossRef]
- Kausar, A. Performance of corrosion protective epoxy blend-based nanocomposite coatings: A review. Polym. Plast. Technol. Eng. 2019, 59, 658–673. [Google Scholar] [CrossRef]
- David, R.; Raja, V.S.; Singh, S.K.; Gore, P. Development of anti-corrosive paint with improved toughness using carboxyl terminated modified epoxy resin. Prog. Org. Coat. 2018, 120, 58–70. [Google Scholar] [CrossRef]
- Chen, C.-L.; Maki, J.S.; Rittschof, D.; Teo, S.L.-M. Early marine bacterial biofilm on a copper-based antifouling paint. Int. Biodeterior. Biodegrad. 2013, 83, 71–76. [Google Scholar] [CrossRef]
- Kim, H.J.; Jung, S.W.; Lim, D.I.; Jang, M.C.; Lee, T.K.; Shin, K.; Ki, J.S. Effects of temperature and nutrients on changes in genetic diversity of bacterioplankton communities in a semi-closed bay, South Korea. Mar. Pollut. Bull. 2016, 106, 139–148. [Google Scholar] [CrossRef]
- Briand, J.F.; Barani, A.; Garnier, C.; Réhel, K.; Urvois, F.; LePoupon, C.; Bouchez, A.; Debroas, D.; Bressy, C. Spatio-temporal variations of marine biofilm communities colonizing artificial substrata including antifouling coatings in contrasted French coastal environments. Microb. Ecol. 2017, 74, 585–598. [Google Scholar] [CrossRef]
- Antunes, J.T.; Sousa, A.G.G.; Azevedo, J.; Rego, A.; Leao, P.N.; Vasconcelos, V. Distinct temporal succession of bacterial communities in early marine biofilms in a Portuguese Atlantic Port. Front. Microbiol. 2020, 11, 1938. [Google Scholar] [CrossRef]
- Ponsonnet, L.; Boureanu, M.; Jaffrezic, N.; Othmane, A.; Dorel, C.; Lejeune, P. Local pH variation as an initial step in bacterial surface-sensing and biofilm formation. Mater. Sci. Eng. C 2008, 28, 896–900. [Google Scholar] [CrossRef]
- Antunes, J.; Leão, P.; Vasconcelos, V. Marine biofilms: Diversity of communities and of chemical cues. Environ. Microbiol. Rep. 2019, 11, 287–305. [Google Scholar] [CrossRef] [PubMed]
- Lanneluc, I.; Langumier, M.; Sabot, R.; Jeannin, M.; Refait, P.; Sablé, S. On the bacterial communities associated with the corrosion product layer during the early stages of marine corrosion of carbon steel. Int. Biodeterior. Biodegrad. 2015, 99, 55–65. [Google Scholar] [CrossRef]
- Procópio, L. Microbial community profiles grown on 1020 carbon steel surfaces in seawater-isolated microcosm. Ann. Microbiol. 2020, 70, 13. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Yan, B.; Duan, J.; Hou, B. Biofilm inhibition effect of an ivermectin/silyl acrylate copolymer coating and the colonization dynamics. Sci. Total. Environ. 2020, 736, 139599. [Google Scholar] [CrossRef]
- Garcia, M.; Procopio, L. Distinct profiles in microbial diversity on carbon steel and different welds in simulated marine microcosm. Curr. Microbiol. 2020, 77, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mugge, R.L.; Lee, J.S.; Brown, T.T.; Hamdan, L.J. Marine biofilm bacterial community response and carbon steel loss following deepwater horizon spill contaminant exposure. Biofouling 2019, 35, 870–882. [Google Scholar] [CrossRef]
- Techtmann, S.M.; Fitzgerald, K.S.; Stelling, S.C.; Joyner, D.C.; Uttukar, S.M.; Harris, A.P.; Alshibli, N.K.; Brown, S.D.; Hazen, T.C. Colwellia psychrerythraea strains from distant deep sea basins show adaptation to local conditions. Front. Environ. Sci. 2016, 4, 33. [Google Scholar] [CrossRef]
- Oliveira, V.M.; Lopes-Oliveira, P.F.; Passarini, M.R.; Menezes, C.B.; Oliveira, W.R.; Rocha, A.J.; Sette, L.D. Molecular analysis of microbial diversity in corrosion samples from energy transmission towers. Biofouling 2011, 27, 435–447. [Google Scholar] [CrossRef]
- Moradi, M.; Song, Z.; Yang, L.; Jiang, J.; Jin, H. Effect of marine Pseudoalteromonas sp. on the microstructure and corrosion behaviour of 2205 duplex stainless steel. Corros. Sci. 2014, 84, 103–112. [Google Scholar] [CrossRef]
- Liu, T.; Pan, S.; Wang, Y.; Guo, Z.; Wang, W.; Zhao, Q.; Zeng, Z.; Guo, N. Pseudoalteromonas lipolytica accelerated corrosion of low alloy steel by the endogenous electron mediator pyomelanin. Corros. Sci. 2020, 162, 108215. [Google Scholar] [CrossRef]
- McBeth, J.M.; Little, B.J.; Ray, R.I.; Farrar, K.M.; Emerson, D. Neutrophilic iron-oxidizing “Zetaproteobacteria” and mild steel corrosion in nearshore marine environments. Appl. Environ. Microbiol. 2011, 77, 1405–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Duan, J.; Hui, X.; Li, Y.; Liu, H.; Fang, G.; Zhai, X. Analysis of bacterial community composition of corroded steel immersed in sanya and xiamen seawaters in china via method of Illumina MiSeq sequencing. Front. Microbiol. 2017, 8, 1737. [Google Scholar] [CrossRef]
- Procopio, L. The era of ‘omics’ technologies in the study of microbiologically influenced corrosion. Biotechnol. Lett. 2020, 42, 341–356. [Google Scholar] [CrossRef]
- Iino, T.; Ito, K.; Wakai, S.; Tsurumaru, H.; Harayama, S. Iron corrosion induced by nonhydrogenotrophic nitrate-reducing Prolixibacter sp. strain MIC1-1. Appl. Environ. Microbiol. 2015, 81, 1839–1846. [Google Scholar] [CrossRef] [Green Version]
- Ibtissem, D.; Sophie, R.; Alexis, B.; Alain, D.; David, A.; Valérie, S.; Sophie, S.; Isabelle, L. Marine bacteria from the French Atlantic coast displaying high forming-biofilm abilities and different biofilm 3D architectures. BMC Microbiol. 2015, 15, 231. [Google Scholar]
- Procopio, L. The role of biofilms in the corrosion of steel in marine environments. World J. Microbiol. Biotechnol. 2019, 35, 73. [Google Scholar] [CrossRef]
- Elumalai, P.; Parthipan, P.; AlSalhi, M.S.; Huang, M.; Devanesan, S.; Karthikeyan, O.P.; Kim, W.; Rajasekar, A. Characterization of crude oil degrading bacterial communities and their impact on biofilm formation. Environ. Pollut. 2021, 286, 117556. [Google Scholar] [CrossRef]
- Elumalai, P.; Parthipan, P.; Narenkumar, J.; Anandakumar, B.; Madhavan, J.; Oh, B.-T.; Rajasekar, A. Role of thermophilic bacteria (Bacillus and Geobacillus) on crude oil degradation and biocorrosion in oil reservoir environment. Biotech 2019, 9, 79. [Google Scholar] [CrossRef]
- Yang, Y.; Masoumeh, M.; Zhou, E.; Liu, D.; Song, Y.; Xu, D.; Wang, F.; Smith, J.A. Streptococcus mutans biofilms induce metabolite-mediated corrosion of 316 L stainless steel in a simulated oral environment. Corros. Sci. 2021, 182, 109286. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Liu, J.; Li, Q.; Li, W.; Xiao, X.; Zhang, L. The Influence of Distinct Seasons on the Succession and Diversity of Bacteria on the Anticorrosive Coatings Surfaces in a Marine Environment. Water 2022, 14, 3183. https://doi.org/10.3390/w14193183
Li S, Liu J, Li Q, Li W, Xiao X, Zhang L. The Influence of Distinct Seasons on the Succession and Diversity of Bacteria on the Anticorrosive Coatings Surfaces in a Marine Environment. Water. 2022; 14(19):3183. https://doi.org/10.3390/w14193183
Chicago/Turabian StyleLi, Shuangwei, Jie Liu, Qian Li, Wenfang Li, Xinfeng Xiao, and Linlin Zhang. 2022. "The Influence of Distinct Seasons on the Succession and Diversity of Bacteria on the Anticorrosive Coatings Surfaces in a Marine Environment" Water 14, no. 19: 3183. https://doi.org/10.3390/w14193183