
New Data on Reproductive System and Spermatozoa Confirm Macrodasys as a Model in Comparative Reproductive Analysis in Macrodasyida (Gastrotricha)
 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
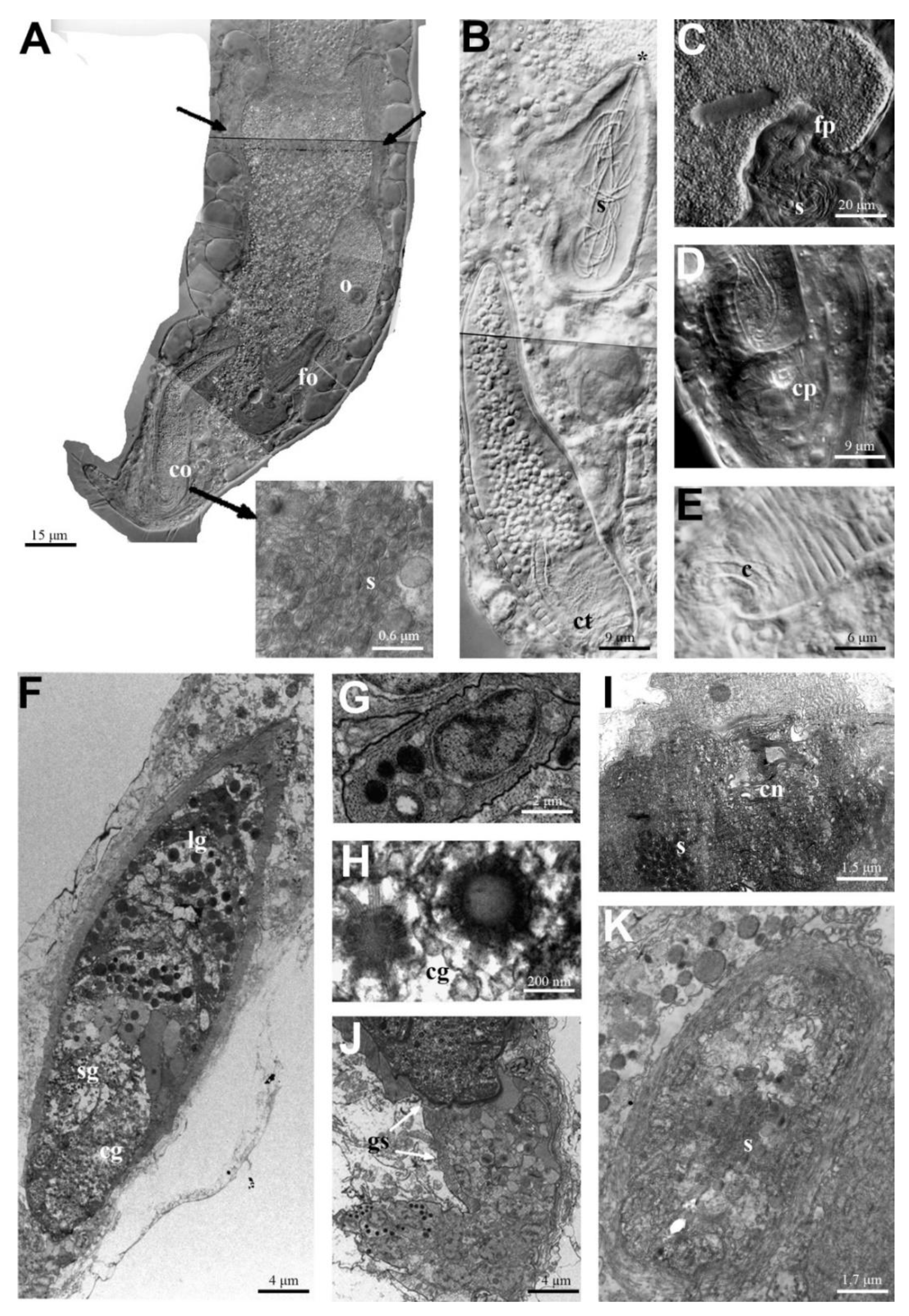
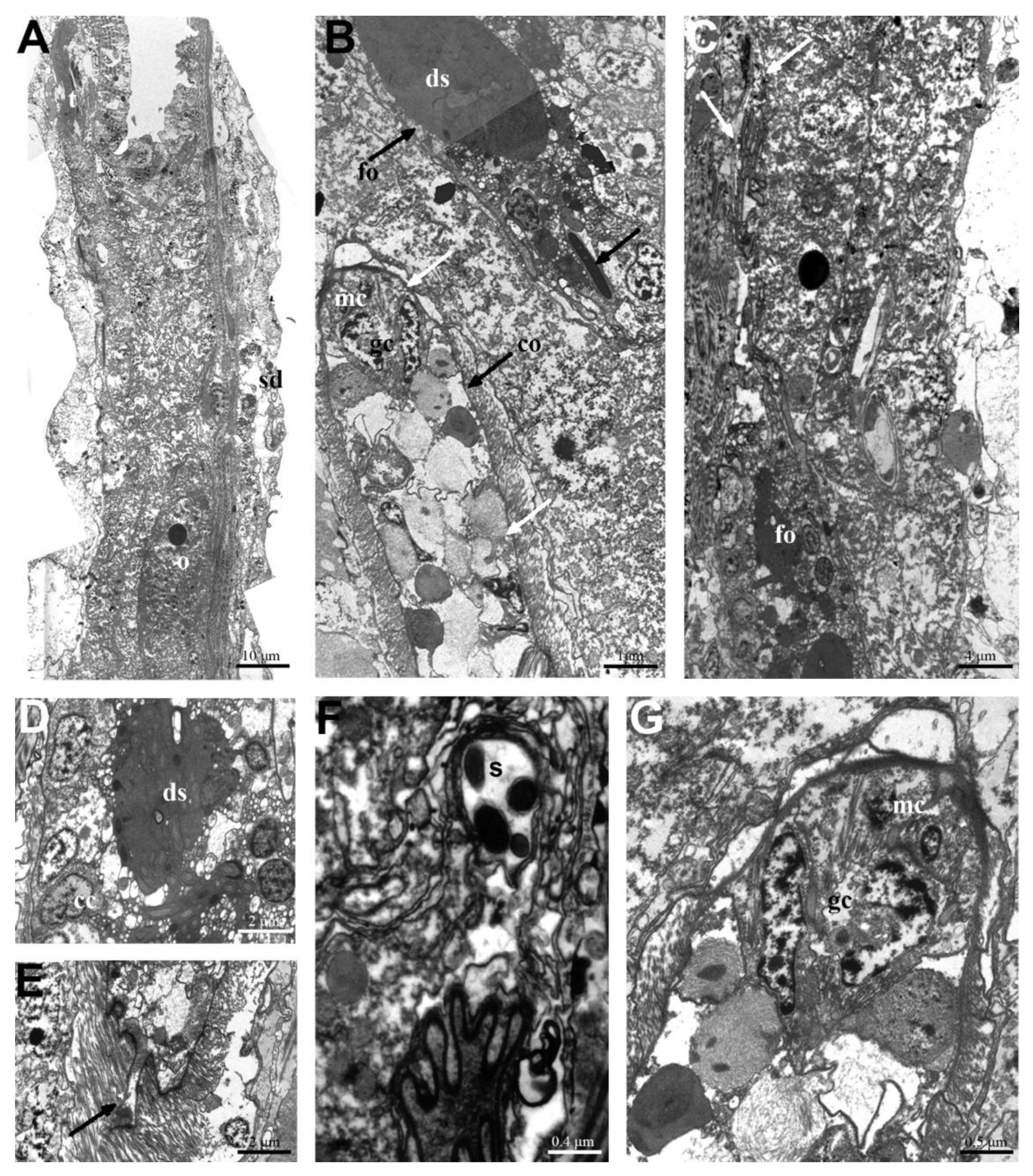
3.1. Macrodasys caudatus and Macrodasys sp. 3: Reproductive System
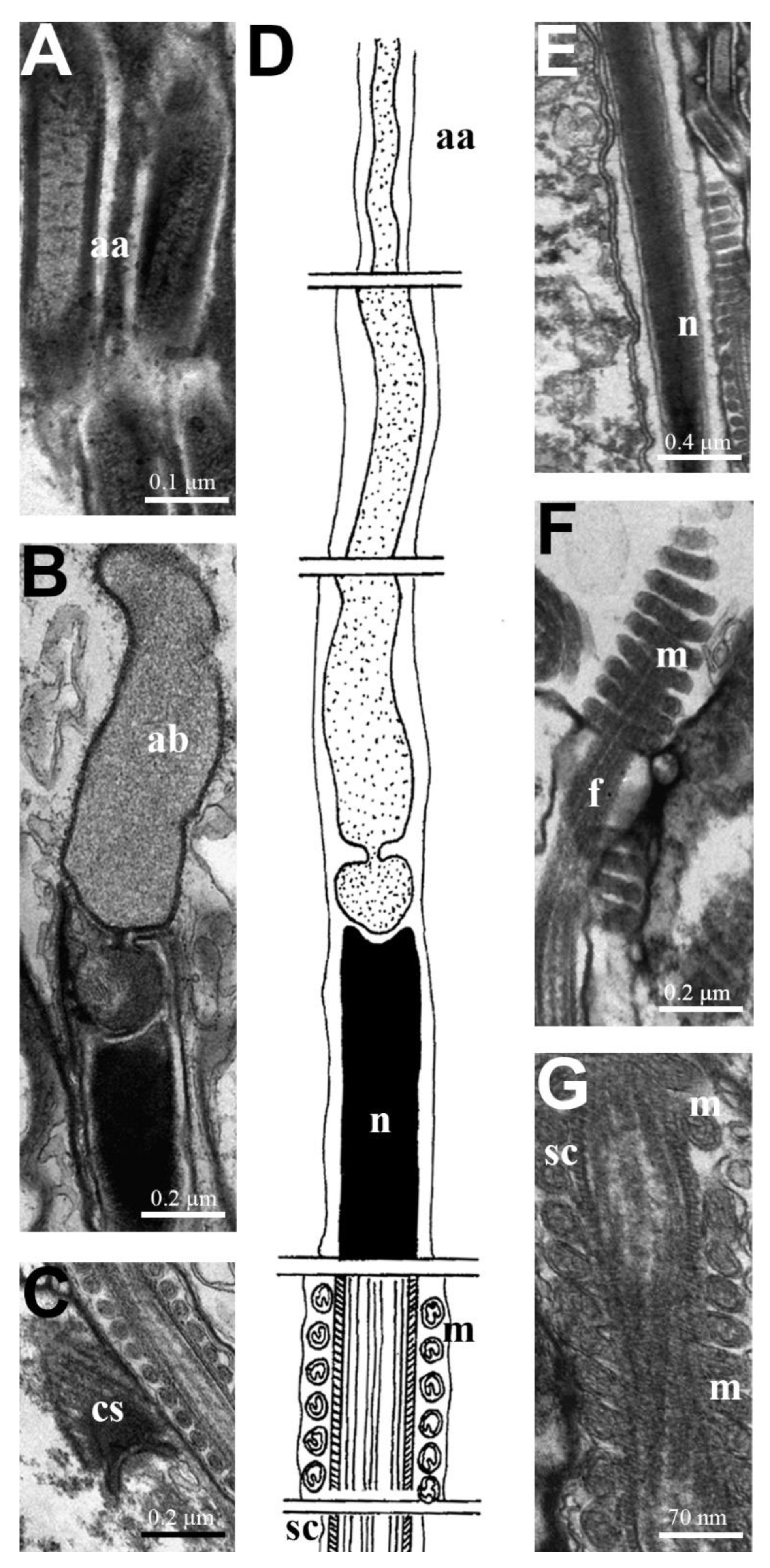
3.2. Macrodasys Spermatozoa
4. Discussion
4.1. Macrodasys caudatus and Macrodasys sp. 3: Reproductive System
4.2. Macrodasys Spermatozoa
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weiss, M.J. Widespread hermaphroditism in freshwater gastrotrichs. Invertebr. Biol. 2001, 120, 308–341. [Google Scholar] [CrossRef]
- Leasi, F.; Todaro, M.A. Meiofaunal cryptic species revealed by confocal microscopy: The case of Xenotrichula intermedia (Gastrotricha). Mar. Biol. 2009, 156, 1335–1346. [Google Scholar] [CrossRef]
- Kånneby, T.; Todaro, M.A.; Jondelius, U. Phylogeny of Chaetonotidae and other Paucitubulatina (Gastrotricha: Chaetonotida) and the colonization of aquatic ecosystems. Zool. Scr. 2013, 42, 88–105. [Google Scholar] [CrossRef]
- Ruppert, E.E. Gastrotricha. In Microscopic Anatomy of Invertebrates; Harrison, F.W., Ruppert, R.R., Eds.; Wiley-Liss: New York, NY, USA, 1991; Volume 4, pp. 41–109. [Google Scholar]
- Kieneke, A.; Schmidt-Rhaesa, A. Gastrotricha. In Handbook of Zoology; Schmidt-Rhaesa, A., Ed.; De Gruyter: Berlin, Germany, 2015; pp. 1–134. [Google Scholar]
- Guidi, L.; Todaro, M.A.; Ferraguti, M.; Balsamo, M. Reproductive system and spermatozoa ultrastructure support the phylogenetic proximity of Megadasys and Crasiella (Gastrotricha, Macrodasyida). Contrib. Zool. 2014, 83, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Todaro, M.A.; Dal Zotto, M.; Cesaretti, A. Marine gastrotrichs from Lanzarote, with a description of a phylogenetically relevant species of Urodasys (Gastrotricha, Macrodasyida). Mar. Biodiv. 2019, 49, 2109–2123. [Google Scholar] [CrossRef]
- Campos, A.; Todaro, M.A.; Garraffoni, A.R.S. A new species of Paraturbanella Remane, 1927 (Gastrotricha, Macrodasyida) from the Brazilian coast, and the molecular phylogeny of Turbanellidae Remane, 1926. Diversity 2020, 12, 42. [Google Scholar] [CrossRef] [Green Version]
- Todaro, M.A.; Dal Zotto, M.; Kånneby, T.; Hochberg, R. Integrated data analysis allows the establishment of a new, cosmopolitan genus of marine Macrodasyida (Gastrotricha). Sci. Rep. 2019, 9, 7989. [Google Scholar] [CrossRef] [Green Version]
- Ruppert, E.E. The reproductive system of gastrotrichs. II. Insemination in Macrodasys: A unique mode of sperm transfer in Metazoa. Zoomorphologie 1978, 89, 201–228. [Google Scholar]
- Kieneke, A.; Arbizu, P.M.; Ahlrichs, W.H. Anatomy and ultrastructure of the reproductive organs in Dactylopodola typhle (Gastrotricha: Macrodasyida) and their possible functions in sperm transfer. Invertebr. Biol. 2008, 127, 12–32. [Google Scholar]
- Todaro, M.A.; Leasi, F.; Hochberg, R. A new species, genus and family of marine Gastrotricha from Jamaica, with a phylogenetic analysis of Macrodasyida based on molecular data. Syst. Biodivers. 2014, 12, 473–488. [Google Scholar] [CrossRef]
- Remane, A. Neue Gastrotricha Macrodasyoidea. Zool. Jb. Syst. 1927, 54, 203–242. [Google Scholar]
- Roszczak, R. Die Psammitgastrotricha des polnischen Ostsee-strandes. Zool. Pol. 1939, 4, 1–24. [Google Scholar]
- Boaden, P.J.S. The interstitial fauna of some north Wales beaches. J. Mar. Biol. Assoc. U. K. 1963, 43, 79–96. [Google Scholar] [CrossRef]
- Ganapati, P.N.; Chandrasekhara Rao, G. On some marine interstitial gastrotrichs from the beach sands of Waltair coast. Proc. Indian Acad. Sci. 1967, 66, 214–225. [Google Scholar] [CrossRef]
- Thane-Fenchel, A. Interstitial gastrotrichs in some south Florida beaches. Ophelia 1970, 7, 113–137. [Google Scholar] [CrossRef]
- Marotta, R.; Guidi, L.; Pierboni, L.; Ferraguti, M.; Todaro, M.A.; Balsamo, M. Sperm ultrastructure of Macrodasys caudatus (Gastrotricha: Macrodasyida) and a sperm based phylogenetic analysis of Gastrotricha. Meiofauna Mar. 2005, 14, 9–21. [Google Scholar]
- Ermak, T.H.; Eakin, R.M. Fine structure of the cerebral and pygidial ocelli in Chone ecaudata (Polychaeta: Sabellidae). J. Ultrastruct. Res. 1976, 54, 243–260. [Google Scholar] [CrossRef]
- Ferraguti, M.; Balsamo, M. Sperm morphology and anatomy of the genital organs in Mesodasys laticaudatus Remane, 1951 (Gastrotricha: Macrodasyida). J. Submicrosc. Cytol. Pathol. 1994, 26, 21–28. [Google Scholar]
- Ruppert, E.E.; Shaw, K. The reproductive system of gastrotrichs. I. Introduction with morphological data for two new Dolichodasys species. Zool. Scr. 1977, 6, 185–195. [Google Scholar] [CrossRef]
- Ruppert, E.E. The reproductive system of gastrotrichs. III Genital organs of Thaumastodermatinae subfaM. n. and Diplodasyinae subfam. n. with discussion of reproduction in Macrodasyida. Zool. Scr. 1978, 7, 93–114. [Google Scholar] [CrossRef]
- Guidi, L.; Ferraguti, M.; Todaro, M.A.; Pierboni, L.; Balsamo, M. Unusual spermatozoa and reproductive modalities of Xenodasys eknomios (Gastrotricha: Xenodasyidae). It. J. Zool. 2009, 76, 165–172. [Google Scholar] [CrossRef]
- Hummon, W.D.; Todaro, M.A. Analytic taxonomy and notes on marine, brackish-water and estuarine Gastrotricha. Zootaxa 2010, 2392, 1–32. [Google Scholar] [CrossRef]
- Remane, A. Morphologie und Verwandschaftsbeziehungen der aberranten Gastrotrichen. I. Z. Morph. Oecol. Tiere 1926, 5, 625–754. [Google Scholar] [CrossRef]
- Schmidt, P. Interstitielle fauna von Galapagos. IV. Gastrotricha. Mikrofauna des Meeresbodens; Akademie der Wissenchaften und der Literatur: Mainz, Germany, 1974; pp. 1–76. [Google Scholar]
- Guidi, L.; Todaro, M.A.; Ferraguti, M.; Balsamo, M. Reproductive system of the genus Crasiella (Gastrotricha, Macrodasyida). Helgol. Mar. Res. 2011, 65, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Fischer, U. Ultrastructure of penetrated spermatozoa, ovary, and oogenesis of Dactylopodola baltica (Gastrotricha, Macrodasyida). Invertebr. Reprod. Dev. 1996, 29, 71–78. [Google Scholar] [CrossRef]
- Balsamo, M.; Ferraguti, M.; Guidi, L.; Todaro, M.A.; Tongiorgi, P. Reproductive system and spermatozoa of Paraturbanella tessieri (Gastrotricha, Macrodasyida): Implications for the sperm transfer modality in Turbanellidae. Zoomorphology 2002, 121, 235–241. [Google Scholar] [CrossRef]
- Evans, W.A. Morphological variability in warm-temperate and subtropical populations of Macrodasys (Gastrotricha: Macrodasyida: Macrodasyidae) with the description of seven new species. Proc. Biol. Soc. Wash. 1994, 107, 239. [Google Scholar]
- Balsamo, M.; Guidi, L.; Pierboni, L.; Marotta, R.; Todaro, M.A.; Ferraguti, M. Living without mitochondria: Spermatozoa and spermatogenesis in two species of Urodasys (Gastrotricha, Macrodasyida) from dysoxic sediments. Invertebr. Biol. 2007, 126, 1–9. [Google Scholar] [CrossRef]
- Guidi, L.; Balsamo, M.; Ferraguti, M.; Todaro, M.A. Reproductive organs and spermatogenesis of the peculiar spermatozoa of the genus Kryptodasys (Gastrotricha, Macrodasyida), with an appraisal of the occurrence and origin of the tail-less spermatozoa in Gastrotricha. J. Zool. Syst. Evol. Res. 2021, 59, 1673–1688. [Google Scholar] [CrossRef]
- Todaro, M.A.; Dal Zotto, M.; Leasi, F. An integrated morphological and molecular approach to the description and systematisation of a novel genus and species of Macrodasyida (Gastrotricha). PLoS ONE 2015, 10, e0130278. [Google Scholar] [CrossRef] [Green Version]
- Guidi, L.; Ferraguti, M.; Pierboni, L.; Balsamo, M. Spermiogenesis and spermatozoa in Acanthodasys aculeatus (Gastrotricha, Macrodasyida): An ultrastructural study. Acta Zool. 2003, 84, 77–85. [Google Scholar] [CrossRef]
- Guidi, L.; Pierboni, L.; Ferraguti, M.; Todaro, M.A.; Balsamo, M. Spermatology of the genus Lepidodasys Remane, 1926 (Gastrotricha, Macrodasyida): Towards a revision of the family Lepidodasyidae Remane 1927. Acta Zool. 2004, 85, 211–221. [Google Scholar] [CrossRef]
- Guidi, L.; Garraffoni, A.R.S.; Semprucci, F.; Balsamo, M. Spermatozoa ultrastructure, spermatogenesis, and reproductive system of Acanthodasys australis (Gastrotricha, Macrodasyida). Zool. Anz. 2020, 286, 108–116. [Google Scholar] [CrossRef]
- Todaro, M.A.; Guidi, L.; Ferraguti, M.; Balsamo, M. A fresh look at Dinodasys mirabilis (Gastrotricha, Macrodasyida), with focus on the reproductive apparatus and sperm ultrastructure. Zoomorphology 2012, 131, 115–125. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guidi, L.; Balsamo, M.; Grassi, E.; Semprucci, F.; Todaro, M.A. New Data on Reproductive System and Spermatozoa Confirm Macrodasys as a Model in Comparative Reproductive Analysis in Macrodasyida (Gastrotricha). Water 2022, 14, 3085. https://doi.org/10.3390/w14193085
Guidi L, Balsamo M, Grassi E, Semprucci F, Todaro MA. New Data on Reproductive System and Spermatozoa Confirm Macrodasys as a Model in Comparative Reproductive Analysis in Macrodasyida (Gastrotricha). Water. 2022; 14(19):3085. https://doi.org/10.3390/w14193085
Chicago/Turabian StyleGuidi, Loretta, Maria Balsamo, Eleonora Grassi, Federica Semprucci, and M. Antonio Todaro. 2022. "New Data on Reproductive System and Spermatozoa Confirm Macrodasys as a Model in Comparative Reproductive Analysis in Macrodasyida (Gastrotricha)" Water 14, no. 19: 3085. https://doi.org/10.3390/w14193085