
Alteration of Dissimilatory Nitrate Reduction Pathways in the Intertidal Sediment during Macroalgae Blooms
by
 ,
,
Yukiko Senga
1,*,
Tsubasa Sato
1,
Kanae Shibaki
1,
Megumi Kuroiwa
2,
Seiichi Nohara
3 and
Yuichi Suwa
4 1
Faculty of Sciences, Toho University, 2-2-1 Miyama, Funabashi 274-8510, Japan
2
Graduate School of Engineering, Tokyo University of Agriculture and Technology, Naka-cho 2-24-16, Koganei, Tokyo 184-8588, Japan
3
National Institute for Environmental Studies, 16-2 Onogawa, Ibaraki, Tsukuba 305-8506, Japan
4
Faculty of Science and Engineering, Chuo University, 1-13-27 Kasuga, Bunkyo-ku, Tokyo 112-8551, Japan
*
Author to whom correspondence should be addressed.
Water 2022, 14(19), 3022; https://doi.org/10.3390/w14193022
Submission received: 27 August 2022
/
Revised: 22 September 2022
/
Accepted: 23 September 2022
/
Published: 26 September 2022
(This article belongs to the Special Issue Material Cycling’s around the Sediment-Water Interface: Implications for Understanding, Assessment, Remediation of Eutrophicated and Contaminated Sediment)
Abstract
:To elucidate the effect of macroalgae blooms on dissimilatory nitrate reduction pathways (denitrification, anammox, and DNRA) in sediments of the hypereutrophic Yatsu tidal flat, eastern Japan, sediment denitrification, anammox, and DNRA rates were measured using a 15N tracer technique at two sites affected and unaffected by macroalgae (Ulva) blooms and in incubation experiments with and without Ulva. Anammox was insignificant at both sites and in both experiments. The denitrification rate was consistently higher than the DNRA rate, and its contributions to the total dissimilatory nitrate reduction were 82% and 85% at sites affected and unaffected by Ulva, respectively. In a sediment incubation experiment with Ulva, the contribution of DNRA had increased to approximately 30% on day 7, which is when the sulfide concentration was the highest. Sulfide produced by sulfate reduction during macroalgae blooms inhibited denitrification and did not change the DNRA, and consequently increased the DNRA contribution. On day 21, after reaching the peak sulfide concentration during the late macroalgae collapse, the DNRA contribution decreased to 15%. These results indicated that the DNRA contribution was greater during the macroalgae blooms than at the collapse, although denitrification dominated DNRA regardless of the macroalgal status. Therefore, vigorous macroalgae cover and sulfide production under the macroalgae cover had an important impact on the nitrogen dynamics.
1. Introduction
Blooms of drift green macroalgae in coastal areas have increased both in scale and frequency over the last few decades in response to elevated levels of coastal eutrophication [1,2]. The macroalgae blooms occur mainly in the North Temperate Zone, with America, Europe, and the Asia-Pacific area being the most seriously affected [1]. Macroalgae have diverse ecology and can be widely distributed [1,2,3]. For example, Ulva prolifera has four main propagation methods [1]. Additionally, opportunistic and drift macroalgae, such as Ulva sp., take up large amounts of nutrients, resulting in explosive blooms within short periods [1,2,4]. Macroalgae cover on surface sediment attenuates sunlight penetration and decreases sediment-attached periphytic algae. In addition, macroalgal decomposition promotes anoxic conditions and provides a source of organic carbon and sulfide [4,5,6]. Therefore, large mats of algae have the potential to greatly affect biogeochemical cycles in the coastal sediment, particularly nitrogen dynamics, which involve heterotrophic and autotrophic microorganisms.
Nitrogen is a limiting nutrient in coastal ecosystems. Under aerobic conditions, organic nitrogen is mineralized to ammonium (NH4+), which is continuously oxidized to nitrate (NO3−) via nitrification. Therefore, NO3− and NH4+ are generally the dominant forms of nitrogen nutrients that are delivered from rivers to coastal ecosystems [7]. Dissimilatory nitrate reduction processes, including denitrification, anaerobic ammonium oxidation (anammox), and dissimilatory nitrate reduction to ammonium (DNRA), play important roles in controlling the NO3− and NH4+ dynamics and fate in estuaries and coastal environments [8]. Denitrification generally results from the respiratory reduction of NO3− to nitrogen gas (N2) in response to the oxidation of electron donors such as organic matter [9,10]. However, some chemolithoautotrophic denitrifying organisms use inorganic matter, such as reduced sulfur compounds and hydrogen, instead of organic matter [11]. In anammox, NH4+ is oxidized to N2, with NO3− or NO2− serving as the electron acceptor under anaerobic conditions [12]. Therefore, the processes of denitrification and anammox remove NO3− from coastal ecosystems. In contrast to these processes, DNRA does not remove NO3− from coastal ecosystems in the form of N2 but instead reduces it to NH4+ [13,14]. DNRA is performed by both heterotrophic organisms, which use organic carbon as the electron donor (fermentative), and chemolithoautotrophic organisms, which use nitrate to oxidize sulfide or other reduced inorganic substrates [15]. The balance between denitrification, anammox, and DNRA determines whether bioavailable nitrogen is removed as nitrogen gas (N2) or recycled as NH4+ [16]. Therefore, an understanding of nitrogen transformation is required to assess the nitrogen fate and control nitrogen pollution in aquatic ecosystems [8]. Despite its importance, alterations in sediment denitrification, anammox, and DNRA during macroalgae blooms remain unclear.
The Yatsu tidal flat in eastern Japan, which extends over an area of approximately 40 ha, was isolated from Tokyo Bay by the construction of concrete dikes in the 1970s. Seawater containing excessive nutrients from intrudes in Tokyo Bay enters the Yatsu tidal flat via two small channels. As a result, eutrophication in the tidal flat has progressed over the past few decades [4,17,18]. Macroalgae blooms were first observed in the Yatsu tidal flat in 1995 and gradually continued to develop [19]. Recently, macroalgae blooms of Ulva spp. (Ulva), including Ulva pertusa and U. ohnoi, were observed [17,18,19,20]. Previous studies conducted in the Yatsu tidal flat reported that denitrification rather than anammox is the major nitrogen removal process [18], and Ulva blooms promote sulfide release via sulfate reduction and alter sulfur dynamics from sulfate reduction to sulfide oxidation [4].
In this study, to elucidate the effect of macroalgae blooms on dissimilatory nitrate reduction in sediments of the hypereutrophic Yatsu tidal flat, denitrification, anammox, and DNRA rates were measured in sediments that were affected and unaffected by Ulva blooms using a 15N tracer technique. In addition, in sediment incubation experiments with and without Ulva, the changes in denitrification, anammox, and DNRA rates were observed when the macroalgae status shifted from vigorous macroalgae cover to macroalgal collapse. Moreover, we hypothesized that sulfide production in the presence of macroalgae would influence the dissimilatory NO3− reduction pathways. To understand the effect of sulfide concentration on denitrification, the denitrifying activity was measured with different concentrations of sulfide using the acetylene block method.
2. Materials and Methods
2.1. Study Site and Sampling
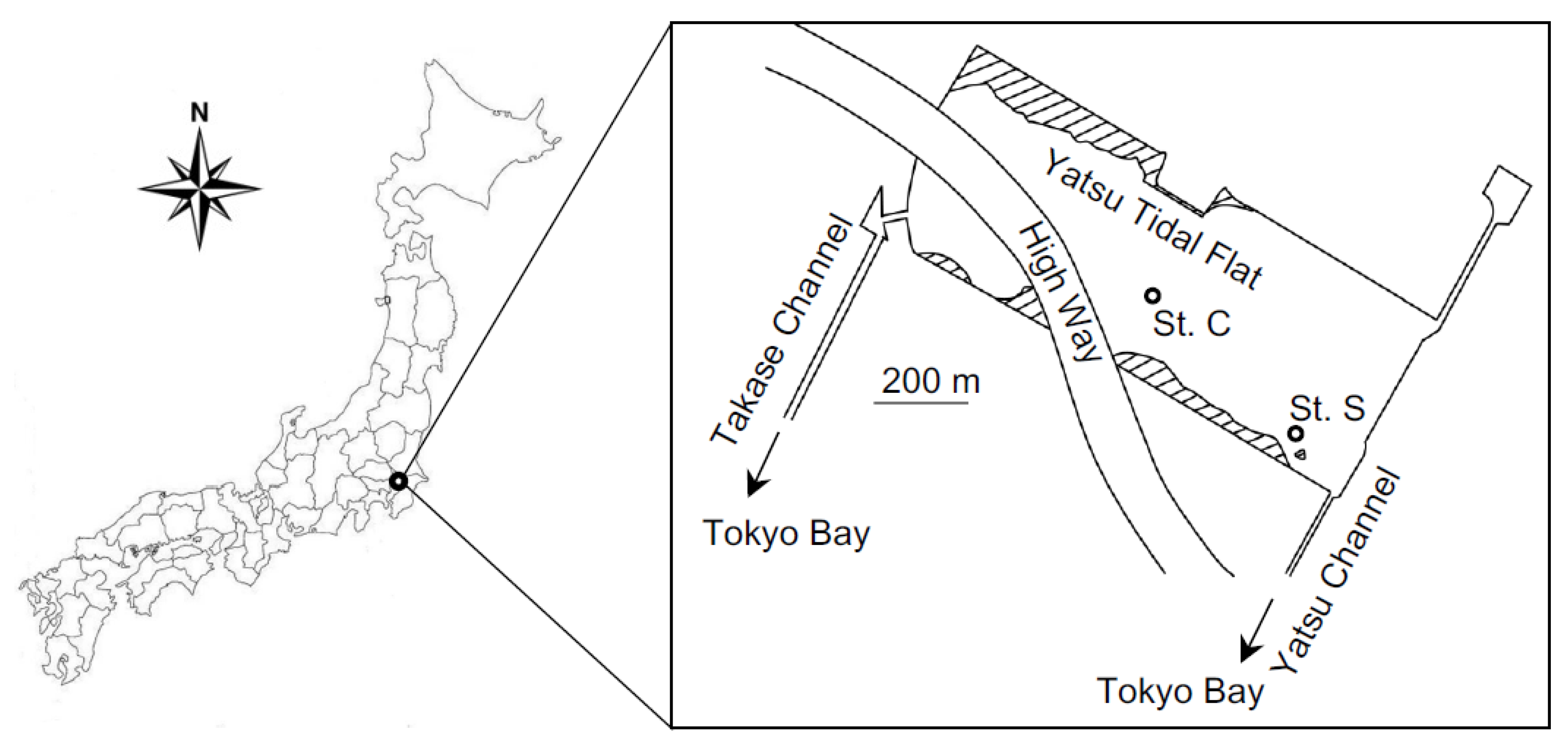
Sampling was performed monthly during low tide at sites C (35°40′34″ N, 140°00′22″ E) and S (35°40′29″ N, 140°00′27″ S) of the hypereutrophic Yatsu tidal flat, eastern Japan (Figure 1). At St. C near the center of the tidal flat, the surface sediment was frequently affected by Ulva cover, while at St. S, located at the southeastern edge of the tidal flat, the sediment was unaffected by the Ulva cover [18].
To examine the denitrification, anammox, and DNRA rates using a 15N tracer technique, surface sediments (0–5 cm depth) at Sts. C and S were collected in November 2017. Sediment sampled at St. C was also used for sediment incubation experiments with and without Ulva. In November 2017, there were almost no macroalgae at Sts. C and S of the Yatsu tidal flat. The sediment temperature and pH were measured using a sensor (IQ170, IQ Scientific Instruments, Carlsbad, CA, USA). Additionally, the surface sediments sampled at St. C in October 2017 were used for the effect of sulfide on the denitrifying activity. All seawater was collected in the tidal creek, and Ulva samples for sediment incubation experiments were collected near St. C.
2.2. Sediment Incubations
2.2.1. Denitrification, Anammox, and DNRA Rates in Sediments with and without Ulva
Ulva samples were rinsed with tap water and then washed several times with sterile seawater. Approximately 250 mL of sediment and 200 mL of seawater were transferred to 500 mL glass flasks. The flasks were mixed overnight to oxidize and remove the sulfide. A 35 g wet Ulva sample (0.6 kg DW m−2 of Ulva biomass) was added to each flask. Control flasks (without Ulva) were also prepared. The flasks with and without Ulva were prepared in triplicate. The flasks were incubated for 21 days at 25 °C under a 12 h:12 h light:dark cycle. Seawater and sediment were collected from the flasks at 0, 7, and 21 days to measure the denitrification, anammox, and DNRA rates using a 15N tracer technique. Nutrients and sulfide concentrations in the seawater were also measured.
2.2.2. Effect of Sulfide on Denitrifying Activity
The effect of sulfide on denitrification was examined using the acetylene block method [21,22]. This method is suitable for clarifying the effect of sulfide on denitrification only. Approximately 5 g of wet sediment and 15 mL of seawater were transferred to a 70 mL glass vial. The vial was then sealed with a butyl-rubber stopper and an aluminum cap, after which the air in the headspace was replaced with N2. A potassium nitrate (KNO3) solution and acetylene (C2H2) were then added to give final concentrations of 5 mg-N L−1 and 6 kPa, respectively.
A sodium sulfide (Na2S) solution was prepared by bubbling argon in water. The Na2S solution was added to the vials to attain concentrations ranging from 0 to 200 mg-S L−1 before adding a KNO3 solution and C2H2. Six vials were prepared for each sulfide concentration treatment. Sulfide concentrations in the slurry phase of three vials were measured immediately (initial sulfide concentration). The remaining three vials were incubated under anaerobic conditions for 20 h at 20 °C in the dark. After incubation, N2O in the gas phase of the vial was measured to determine the denitrifying activity. Additionally, the sulfide concentration was measured immediately after the N2O measurement (final sulfide concentration).
2.3. Chemical Analysis
Denitrification, Anammox, and DNRA Rates Measured Using a 15N Tracer Technique
The denitrification and anammox rates were measured using a 15N tracer technique [23,24,25,26,27]. A sediment slurry (10% w/v) was dispensed into glass vials after sterilizing the seawater via autoclaving at 121 °C for 15 min, and the vials were tightly capped with butyl-rubber stoppers and purged with helium. The vials were incubated at 20 °C in the dark, and the slurry was continuously stirred using a magnetic stirrer. The incubation was performed strictly anaerobically with combinations of 15N-labeled or unlabeled ammonium chloride (NH4Cl) and KNO3 as reactive substrates for N2 production. The sediment suspension was incubated with 5.6 mg-N L−1 NH4Cl and 28 mg-N L−1 KNO3 in the following combinations: (i) 15NH4Cl + KNO3, (ii) NH4Cl + K15NO3, (iii) 15NH4Cl without NO3− (negative control), and (iv) no NH4+ or NO3− (negative control). The N2 isotopologues (14N15N, 15N15N) in the headspace gas of each vial were collected periodically using a gas-tight syringe under a helium stream and were quantified using a quadrupole gas chromatography–mass spectrometry system (6890N-5973 inert, Agilent Technologies, Santa Clara, CA, USA). The denitrification and anammox rates were estimated by quantifying two N2 isotopologues. See Senga et al. [18] for a more detailed description of the procedure.
The DNRA rate was measured using 14NH4+/15NH4+ analyses via sequential conversion to N2O [28]. After measurements of denitrification and anammox rates, a water sample in a 20 mL vial was immediately collected via centrifugation (4 °C, 3000 rpm, 5 min). Then, 5 mL of water sample was filtered and transferred to a new vial. Magnesium oxide and a polytetrafluoroethylene (PTFE sealing tape, Sigma-Aldrich, St. Louis, MI, USA) envelope containing an acidified glass fiber filter (Whatman GF/D; GE Healthcare Bio-Sciences, Pittsburgh, PA, USA) were added to the vial. After sealing the vial with a butyl-rubber stopper and an aluminum cap, it was shaken at 150 rpm for 3 h at 4 °C to trap NH4+ in the water sample on the acidified glass fiber filter into the envelope as an ammonium salt. After shaking, the PTFE envelope was removed from the vial and opened to remove the glass fiber filter. The glass filter was transferred to an 11 mL screw cap test tube containing a persulfate-oxidizing reagent. The tube was autoclaved for 1 h at 121 °C to oxidize NH4+ to NO3−. After the NO3− converted from NH4+ was oxidized to N2O via the denitrifier method, the amounts of 45N2O and 46N2O in the headspace gas were determined using quadrupole GC/MS. The DNRA rate was calculated by quantifying the two N2O isotopologues. See Kuroiwa et al. [28] for a more detailed description of the procedure.
2.4. Other Quantifications
To determine the sediment water content, approximately 5 g of wet sediment was weighed (wet weight), dried for 24 h at 105 °C to a constant weight, and weighed again. The dry sediment was then homogenized and combusted at 450 °C for 12 h to determine the loss on ignition (ignition loss).
The pore water in the sediment at St. C was extracted from a sediment sample via centrifugation at 3000× g for 10 min. Insufficient pore water for analysis was extracted from the sediment at St. S due to the low water content. The supernatant was filtered through a pre-combusted (450 °C) glass fiber filter (Whatman GF/F, GE Healthcare) and analyzed for NO3−, NH4+, NO2−, and PO43− concentrations. Seawater was filtered through a pre-combusted glass fiber filter (Whatman GF/F) to determine the NO3−, NH4+, NO2−, and PO43− concentrations. The NO3−, NO2−, NH4+, and PO43− concentrations were measured using the methods described by Wood et al. [29], Bendschneider and Robinson [30], Sagi [31], and Murphy and Riley [32], respectively. The sulfide concentration in seawater was analyzed following the in situ extraction method of Davison and Lishman [33] and Sugahara et al. [34]. All analytical procedures were performed in a sealed system of glass syringes to prevent the volatilization of sulfide and sulfide oxidation by air. Approximately 20 mL of seawater was added to a graduated glass syringe, which was filtered using a 0.45 μm Millex filter (Millipore, Carrigtwohill, Ireland) into an empty 10 mL syringe (syringe–filter–syringe system). The sulfide concentration of the filtrate was spectrophotometrically determined using the methylene blue method [35]. Hereafter, sulfide refers to the sum of the dissolved H2S gas and its dissociated forms, namely, HS− and S2−.
The N2O in the acetylene block method was measured using a gas chromatograph with an electron capture detector (GC-ECD) (6890, Agilent Technologies, Palo Alto, CA, USA). The amount of dissolved N2O was calculated using the solubility formula of Weiss and Price [36].
3. Statistical Analysis
Significant changes in denitrification and DNRA rates; NH4+, NO2−, NO3−, and sulfide concentrations with and without Ulva; and denitrification activities with different sulfide concentrations were identified using analysis of variance (ANOVA). Student’s t-test was used to compare the initial and final sulfide concentrations in the experiment to determine the effect of sulfide on denitrification. The statistical analyses were performed using R statistical software [37].
4. Results
4.1. Denitrification, Anammox, and DNRA Rates in Sediments Affected and Unaffected by Ulva
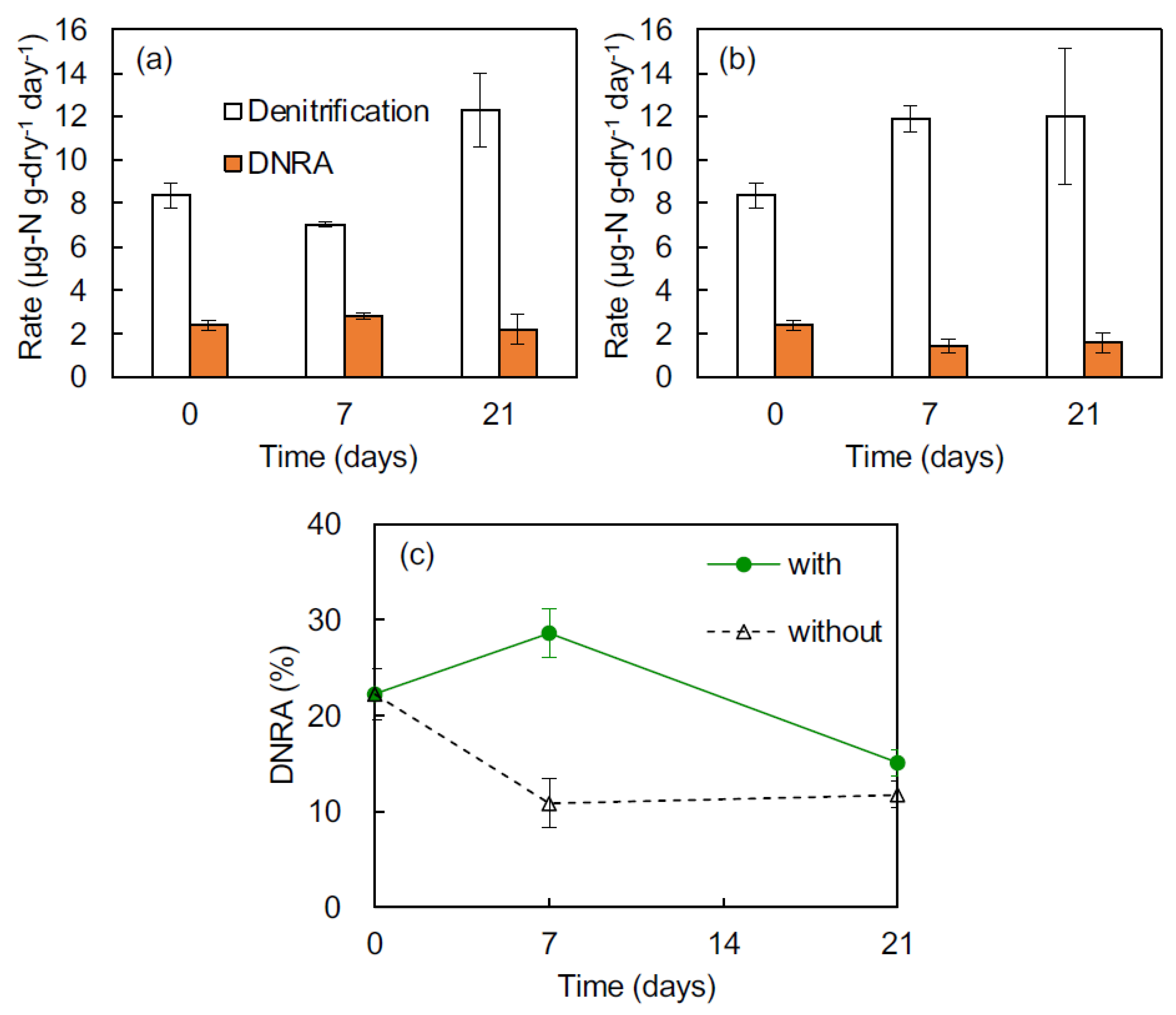
Anammox was not detected in November 2017 in the surface sediments at Sts. C and S (Table 1). The contributions of denitrification and DNRA to total dissimilatory NO3− reduction (i.e., denitrification + DNRA) at St. C were 82% and 18%, respectively. On the other hand, at St. S, the contributions of denitrification and DNRA were 85% and 15%, respectively. The DNRA rates at both sites did not exceed the denitrification rates. The denitrification and DNRA rates at St. C were higher than at St. S.
4.2. Denitrification, Anammox, and DNRA Rates in Sediment Incubations with and without Ulva
The biomass of Ulva samples added to the flasks (0.6 kg DW m−2) was similar to the maximum biomass of the blooms in May 2016 [18]. It was visually confirmed that the Ulva biomass decayed greatly after 7 days. The Ulva debris looked like sludge at 21 days (late Ulva collapse). Whitish and pink particles above the Ulva at the air/water interface were observed after day 7.
All concentrations in the seawater phase of the flasks, except for NH4+ and sulfide without Ulva, varied significantly with time (ANOVA, p < 0.05; Figure 2). The concentration of NH4+ with Ulva increased with time, while almost no NH4+ was detected in the flask without Ulva (Figure 2a). In the flask with Ulva, the NO2− concentration measured on day 0 (ca. 100 μg-N L−1) was completely consumed by day 7 (Figure 2b). The NO2− concentration in the flask without Ulva peaked on day 7 and then decreased to almost nil by day 21. In both the presence and absence of Ulva, the initial concentration of NO3− (ca. 260 μg-N L−1) on day 0 had largely decreased by day 7, and was completely consumed by day 21 (Figure 2c). The concentration of sulfide in the seawater phase of flasks containing Ulva peaked on day 7 and then decreased until day 21, while sulfide was not detected in the flask without Ulva (Figure 2d).
The rate of denitrification in the presence of Ulva varied significantly with time (ANOVA, p < 0.01; Figure 3a). The denitrification rate with Ulva decreased slightly on day 7 and then increased on day 21. The rates of denitrification in the absence of Ulva did not vary significantly with time (Figure 3b). The DNRA rates in the presence and absence of Ulva tended to slightly increase and decrease over time, respectively, but not significantly.
There were significant differences between times regarding the contributions of DNRA to the total dissimilatory NO3− reduction with and without Ulva (Figure 3c; ANOVA, p < 0.05). The contribution of DNRA with Ulva significantly increased from approximately 22% on day 0 to 29% on day 7, and then significantly decreased to 15% on day 21. On the other hand, the contribution of DNRA without Ulva significantly decreased to 11% on day 7 and was constant between days 7 and 21.
4.3. Effect of Sulfide on Denitrifying Activity
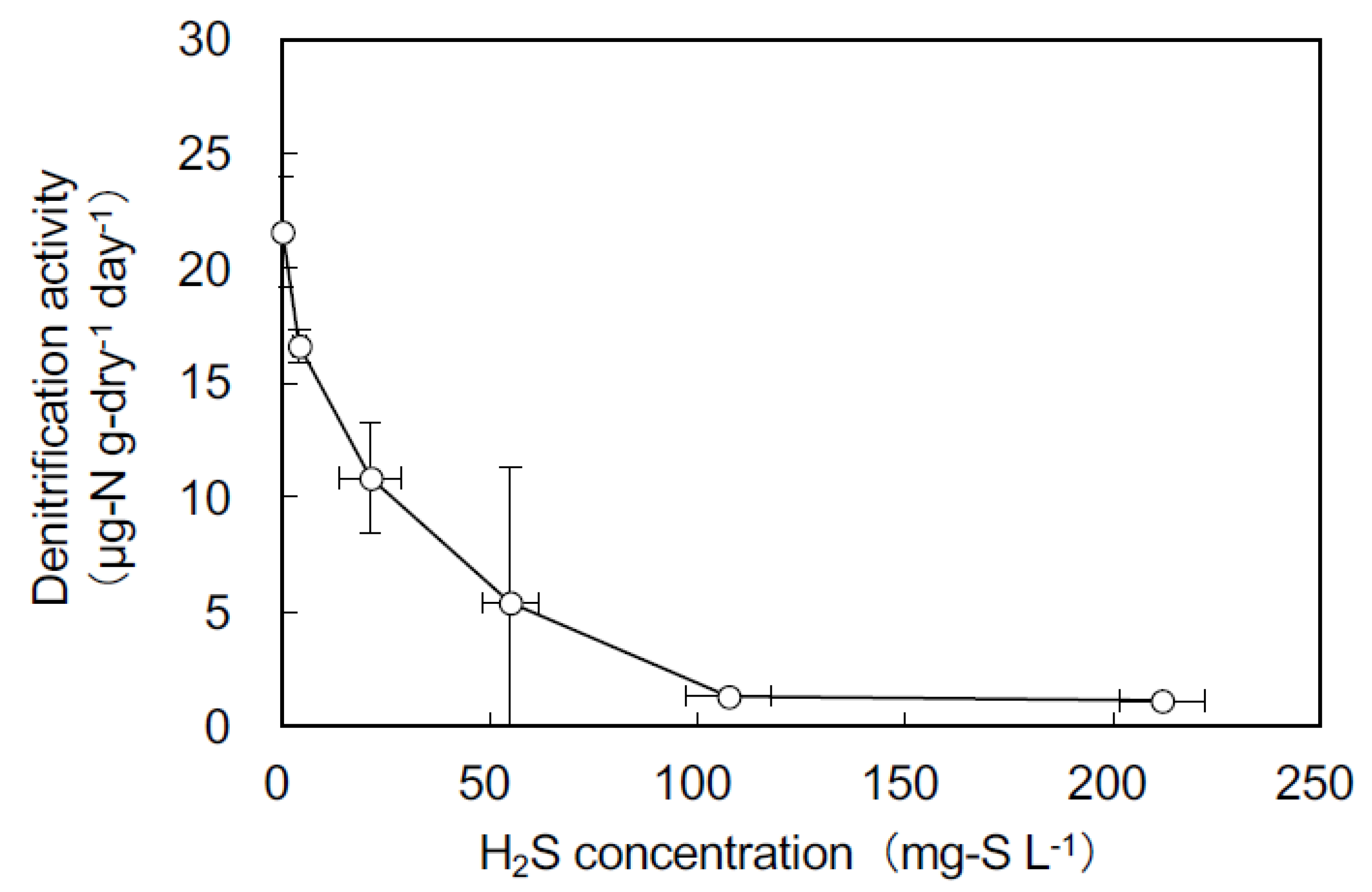
The effect of sulfide concentration on denitrification in the surface sediment at St. C was quantified using the acetylene block method. Although sulfide was added at 0, 5, 20, 50, 100, and 200 mg-S L−1, the sulfide concentrations measured in the seawater at the beginning of the incubation were 0, 4, 21, 55, 108, and 212 mg-S L−1, respectively (Figure 4 and Table 2). The denitrifying activities varied significantly with sulfide concentration (ANOVA, p < 0.001; Figure 4). The denitrifying activities drastically decreased between 0 and 108 mg-S L−1 and were constant between 108 and 212 mg-S L−1 sulfide concentrations (Figure 4). After the incubation, there were significant differences between the initial and final sulfide concentrations for all treatments (Student’s t-test, p < 0.01; Table 2). All of the final sulfide concentrations decreased by >40%.
5. Discussion
5.1. Dissimilatory NO3− Reduction Pathways in the Yatsu Tidal Flat
In this study, anammox was negligible at Sts. C and S. In a previous study, we reported that anammox at Sts. C and S was consistently low compared with denitrification, accounting for <7% of the total N2 production in September 2015 and November 2016 [18]. This result suggested that anammox played a limited role in NO3− removal. Therefore, the principal pathway of nitrogen removal in the surface sediment at the hypereutrophic Yatsu tidal flat was denitrification. Anammox generally plays a limited role in regulating NO3− reduction in eutrophic coastal ecosystems [6,7,38].
The rates of denitrification and DNRA were approximately twofold higher at St. C than at St. S (Table 1). Ulva cover on the surface sediment was frequently found at St. C, but not at St. S. Ignition loss, as a measure of organic content, at St. C was higher than at St. S, probably due to the presence of Ulva (Table 1). Additionally, at St. C, sulfide accumulation under macroalgal mats was reported [4]. There are two ways to enhance denitrification and DNRA: a heterotrophic (fermentative) process with organic carbon as an electron donor and a chemolithoautotrophic process with sulfide or Fe (II) as an electron donor [7,15,39]. Therefore, because there was a rich source of electron donors to drive denitrification and DNRA, the rates of denitrification and DNRA were higher at St. C than at St. S. From August 2012 to January 2017, the denitrifying activity found using the acetylene block method was also found to be higher at St. C than at St. S [18]. A key factor that regulates denitrification in the sandy sediment at St. S is the organic carbon content, while the key factor at St. C is nitrogen substrates due to competition with macroalgae for nitrogen.
There was no significant difference in the contribution of DNRA to the total dissimilatory NO3− reduction between Sts. C and S. Unfortunately, on November 2017 when sediment was collected from the Yatsu tidal flat, no macroalgae blooms were found at St. C. Therefore, the effect of macroalgae cover or its collapse on the dissimilatory NO3− reduction could not be determined. However, even in the absence of macroalgae, there were differences in the rates of denitrification and DNRA between Sts. C and S, but there were no significant differences in the ratios. The surface sediment at the Yatsu tidal flat had a 10–20% DNRA potential even without the impact of Ulva cover. The estimated contribution of DNRA in this study was in the range of values previously reported values for other intertidal sediments. In the intertidal permeable sediments of the eutrophied Wadden Sea, DNRA also accounted for 10–20% of the NO3− consumption [40]. The DNRA in the intertidal sediments of the Yangtze Estuary and Colne Estuary accounted for 3–45% and 11–60% of the total NO3− consumption, respectively [8,41]. The DNRA is of major importance in most shallow coastal sediments [15].
5.2. Influence of Macroalgae Blooms on Dissimilatory NO3− Reduction Pathways
Although sulfide in flasks without Ulva was not detected in seawater, the presence of Ulva accelerated the sulfide production via sulfate reduction in the sediment until day 7, and the sulfide concentration decreased thereafter (Figure 2d). Sulfide is oxidized both chemically and microbially [42,43]. Since whitish and pink particles were observed in the flasks containing Ulva after day 7, most of the sulfide was expected to have been oxidized microbially. Namely, these particles might be sulfide-oxidizing bacteria (purple sulfur bacteria) or elemental sulfur. These results are consistent with the results obtained in the incubation experiment of the previous study in which sulfate reduction would be shifted to sulfur oxidation after the peak of sulfide production [4]. In this study, the sulfide was produced largely by sulfate reduction in flasks containing Ulva on day 7 and was transformed to elemental sulfur by purple sulfur bacteria between days 7 and 21. Under such a change in sulfur speciation, the NO3− and NO2− concentrations were almost undetectable between days 7 and 21, while the NH4+ concentration increased over time (Figure 2a–c), suggesting that the anaerobic environment was conserved in flasks containing Ulva throughout the experiment, and there would be virtually no nitrification in the presence of macroalgae.
The rate of denitrification decreased in the flasks containing Ulva from days 0 to 7. The sulfide concentration was the highest on day 7. The denitrification rate was reduced with increasing sulfide concentration, as sulfide inhibited the reduction of N2O to N2 [39,44]. In the experiment using sediment from the Yatsu Tidal Flat, negative effects of >4 mg-S L−1 on denitrifying activity were observed (Figure 4). Therefore, sulfide production under macroalgae covers negatively affected denitrification in the Yatsu tidal flat. The DNRA rates were unchanged on day 7; therefore, DNRA was not affected by sulfide. Previous studies reported that treatment with high concentrations of sulfide results in either an increase or no difference in the DNRA rate [6,16,44,45]. In the surface sediment at the Yatsu tidal flat, sulfide produced in the presence of vigorous macroalgae negatively influenced denitrification but had no significant effect on the DNRA rate. Consequently, the DNRA contribution to total dissimilatory NO3− reduction increased from 20% on day 0 to 30% on day 7. A similar result was reported in the sediment of the invasive Spartina alterniflora marshland in the Yangtze River Estuary, China [46]. The invasion of Spartina alterniflora increased DNRA by altering the SO42− concentration and pH in the sediment.
The denitrification rate in the flasks with Ulva increased, while the DNRA rate was unchanged between days 7 and 21. On day 21, the sulfide concentration had decreased greatly from day 7, and elemental-sulfur-like particles were observed above the Ulva. In the experiment examining the effect of the sulfide concentration on the denitrifying activity, the sulfide concentration decreased by 40–100% after 20 h (Table 2). This result suggested that sulfide produced in the presence of Ulva might be consumed via sulfur oxidation by microorganisms. Autotrophic denitrification, sulfide oxidation coupled with nitrate reduction was found in the salt marsh at the Plum Island Ecosystem Long-Term Ecological Research site in Rowley, USA [16]; the Gulf of Thailand [47]; and the Yarra River Estuary, Australia [48]. We hypothesized that sulfide produced beneath Ulva mats inhibits general heterotrophic denitrification and accelerates sulfide-oxidizing denitrification, but does not affect DNRA. The macroalgae status alters sulfur dynamics and affects dissimilatory NO3− reduction pathways. The details of the relationship between sulfur dynamics and the dissimilatory NO3− reduction pathways in the Yatsu tidal flat need to be explored using molecular microbiology techniques.
The contribution of DNRA in the presence of Ulva decreased on day 21 to 15% due to an increase in the denitrification rate and was similar to the DNRA observed on day 21 without Ulva (12%). This result indicated that DNRA was greater during the blooms than at the collapse. The amount of NO3− removed by denitrification decreased and the proportion of N that remained as NH4+ consequently increased during the macroalgae blooms. This result suggested that macroalgae controlled the removal versus retention of bioavailable nitrogen within the sediment under anaerobic conditions. This is very important in terms of the physiology of Ulva because Ulva in the Yatsu tidal flat preferentially uptakes NH4+ over NO3− [4]. An increase in ammonification is considered to provide a strong advantage to the subsequent generation of Ulva blooms.
In the flasks containing Ulva, NH4+ was likely produced via DNRA, but also via Ulva decomposition, especially in the late experiment (Figure 2a). NH4+ is reported to account for a major proportion of the total nitrogen efflux during the decomposition of Ulva sp. [49]. NH4+ produced via decomposition is typically greater than that produced via DNRA in coastal ecosystems [50,51]. In the anaerobic sediment where DNRA and decomposition occur, nitrification was not able to progress; consequently, denitrification did not occur. Thus, it was expected that the large proportion of nitrogen present as NH4+ would not transform further and would be recycled by macroalgae such as Ulva in these ecosystems.
The behaviors of NH4+, NO2−, and NO3− indicated that nitrification occurred in the flasks without Ulva (Figure 2a–c). Sulfide, which inhibits denitrification and nitrification [52], was not produced in the flasks without Ulva (Figure 2d). Additionally, the rate of denitrification in the absence of Ulva was higher from days 7 to 21 than on day 0 (Figure 3b). Nitrogen removal pathways, i.e., nitrification–denitrification, appeared to proceed at a constant rate in the flasks without Ulva for 21 days. However, DNRA progressed slightly, and its contribution was 12% (Figure 3c). This contribution was in the range of those measured at Sts. C and S (Table 1). Therefore, in the sediment at the Yatsu tidal flat, the DNRA potential was in the range of 10–20% and increased to approximately 30% only when there were macroalgae blooms.
6. Conclusions
This study demonstrated the important role of the dissimilatory nitrate reduction pathway in nitrogen dynamics in the sediment hypereutrophic Yatsu tidal flat. Denitrification was a major pathway for dissimilatory nitrate reduction regardless of the macroalgal status. Macroalgae cover on the sediment accelerated sulfide production, which inhibited denitrification and enhanced the DNRA contribution (30%). During the late collapse of macroalgae blooms, sulfate reduction shifted to sulfide oxidation. Consequently, the denitrification rate increased and the DNRA contribution returned to its original value in the absence of macroalgae (10–20%). During macroalgae blooms, NH4+ tends to accumulate because NH4+ is produced by macroalgal decomposition and DNRA, and nitrification is not able to proceed in the reductive sediment under macroalgae mats. NH4+ accumulation was very important for sustaining the growth of macroalgae blooms, as well as for the progress of eutrophication.
Author Contributions
Y.S. (Yukiko Senga): Designing the study, measuring environmental factors, analyzing the data including statistics, and drafting the article. T.S.: Conducting experiments, measuring denitrification, anammox, and DNRA rates using a 15N tracer technique. K.S.: Conducting experiments, measuring denitrification activity using a C2H2 block method. S.N.: Set-up the samplings and analytical instruments. M.K.: Set-up the experiments and analytical instruments. Y.S. (Yuichi Suwa): Designing of the study, and drafting and final approval of the article. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Japan Society for the Promotion of Science KAKENHI grants (no. JP15K00525 and JP21K12224 to Senga).
Acknowledgments
The study site is located in a protected area of the National Natural Park under the control of the Ministry of the Environment of Japan. This study was conducted with the permission of the related organizations. We thank the Yatsu Higata Nature Observation Center for keeping this area open and providing assistance. We also thank the students in the Senga laboratory at Toho University for their assistance with the field surveys and chemical analyses.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ye, N.H.; Zhang, X.W.; Mao, Y.Z.; Liang, C.W.; Xu, D.; Zou, J.; Zhuang, Z.M.; Wang, Q.Y. Green tides are overwhelming the coastline of our blue planet: Taking the world’s largest example. Ecol. Res. 2011, 26, 477–485. [Google Scholar] [CrossRef]
- Valiela, I.; McLelland, J.; Hauxwell, J.; Behr, P.J.; Hersh, D.; Foreman, K. Macroalgal blooms in shallow estuaries: Controls and ecophysiological and ecosystem consequences. Limnol. Oceanogr. 1997, 42, 1105–1118. [Google Scholar] [CrossRef]
- Shah, Z.; Badshah, S.L.; Iqbal, A.; Shah, Z.; Emwas, A.-H.; Jaremko, M. Investigation of important biochemical compounds from selected freshwater macroalgae and their role in agriculture. Chem. Biol. Technol. Agric. 2022, 9, 9. [Google Scholar] [CrossRef]
- Senga, Y.; Kobayashi, W.; Mikawa, K.; Kitazawa, T.; Lee, S.; Shiraki, Y. Influences of green macroalgae blooms on nutrients and sulfide dynamics in hypereutrophic intertidal ecosystems. Limnology 2021, 22, 187–196. [Google Scholar] [CrossRef]
- Nedergaard, R.I.; Risgaard-Petersen, N.; Finster, K. The importance of sulfate reduction associated with Ulva lactuca thalli during decomposition: A mesocosm experiment. J. Exp. Mar. Biol. Ecol. 2002, 275, 15–29. [Google Scholar] [CrossRef]
- Wong, W.W.; Greening, C.; Shelley, G.; Lappan, R.; Leung, P.M.; Kessler, A.; Winfrey, B.; Poh, A.C.; Cook, P. Effects of drift algae accumulation and nitrate loading on nitrogen cycling in a eutrophic coastal sediment. Sci. Total Environ. 2021, 790, 147749. [Google Scholar] [CrossRef]
- Li, X.; Bai, D.; Deng, Q.; Cao, X.; Zhou, Y.; Song, C. DNRA was limited by sulfide and nrfA abundance in sediments of Xiamen Bay where heterotrophic sulfide-producing genus (Pelobacter) prevailed among DNRA bacteria. J. Soils Sediments 2021, 21, 3493–3504. [Google Scholar] [CrossRef]
- Deng, F.Y.; Hou, L.J.; Liu, M.; Zheng, Y.L.; Yin, G.Y.; Li, X.F.; Lin, X.B.; Chen, F.; Gao, J.; Jiang, X.F. Dissimilatory nitrate reduction processes and associated contribution to nitrogen removal in sediments of the Yangtze Estuary. J. Geophys. Res-Biogeo. 2015, 120, 1521–1531. [Google Scholar] [CrossRef]
- Payne, W. Reduction of nitrogenous oxides by microorganisms. Bacteriol. Rev. 1973, 37, 407–452. [Google Scholar] [CrossRef]
- Seitzinger, S.P. Denitrification in freshwater and coastal marine ecosystem: Ecological and geochemical significance. Limnol. Oceanogr. 1988, 33, 702–724. [Google Scholar] [CrossRef]
- Kumar, S.; Herrmann1, M.; Blohm, A.; Hilke, I.; Frosch, T.; Trumbore, S.E.; Kusel, K. Thiosulfate- and hydrogen-driven autotrophic denitrification by a microbial consortium enriched from groundwater of an oligotrophic limestone aquifer. FEMS Microbiol. Ecol. 2018, 94, fiy141. [Google Scholar] [CrossRef] [PubMed]
- Mulder, A.; Vandegraaf, A.A.; Robertson, L.A.; Kuenen, J.G. Anaerobic ammonium oxidation discovered in a denitrifying fluidized-bed reactor. FEMS Microbiol. Ecol. 1995, 16, 177–183. [Google Scholar] [CrossRef]
- Koike, I.; Hattori, A. Denitrification and ammonia formation in anaerobic coastal sediments. Appl. Environ. Microbiol. 1978, 35, 278–282. [Google Scholar] [CrossRef]
- Jørgensen, K.S. Annual pattern of denitrification and nitrate ammonification in estuarine sediment. Appl. Environ. Microbiol. 1989, 55, 1841–1847. [Google Scholar] [CrossRef]
- Giblin, A.E.; Tobias, C.R.; Song, B.; Weston, N.; Banta, G.T.; Rivera-Monroy, V.H. The importance of dissimilatory nitrate reduction to ammonium (DNRA) in the nitrogen cycle of coastal ecosystems. Oceanography 2013, 26, 124–131. [Google Scholar] [CrossRef]
- Murphy, A.E.; Bulseco, A.N.; Ackerman, R.; Vineis, J.H.; Bowen, J.L. Sulphide addition favours respiratory ammonification (DNRA) over complete denitrification and alters the active microbial community in salt marsh sediments. Environ. Microbiol. 2020, 22, 2124–2139. [Google Scholar] [CrossRef] [PubMed]
- Yauchi, E.; Hayami, T.; Imoto, T.; Gomyo, M. An analysis of the extraordinary proliferation of Ulva sp. in Yatsu tidal flat and an estimation of effects for environment. Proc. Coast Eng. JSCE 2006, 53, 1191–1196. (In Japanese) [Google Scholar]
- Senga, Y.; Sato, T.; Kuroiwa, M.; Nohara, S.; Suwa, Y. Anammox and denitrification in the intertidal sediment of the hypereutrophic Yatsu tidal flat, Japan. Estuar. Coasts 2019, 42, 665–674. [Google Scholar] [CrossRef]
- Yabe, T.; Ishii, Y.; Amano, Y.; Koga, T.; Hayashi, S.; Nohara, S.; Tatsumoto, H. Green tide formed by free-floating Ulva spp. at Yatsu tidal flat, Japan. Limnology 2009, 10, 239–245. [Google Scholar] [CrossRef]
- Ishii, Y.; Murakami, K.; Yauchi, E.; Ishii, T.; Taki, K. Effect of Ulva sp. on material circulation in tideland lake located in Tokyo inner bay. Proc. Coast Eng. JSCE 2001, 48, 1136–1140. (In Japanese) [Google Scholar]
- Yoshinari, T.; Knowles, R. Acetylene inhibition of nitrous oxide reduction by denitrifying bacteria. Biochem. Biophys. Res. Commun. 1976, 69, 705–710. [Google Scholar] [CrossRef]
- Yoshinari, T.; Hynes, R.; Knowles, R. Acetylene inhibition of nitrous oxide reduction and measurement of denitrification and nitrogen fixation in soil. Soil Biol. Biochem. 1977, 9, 177–183. [Google Scholar] [CrossRef]
- Thamdrup, B.; Dalsgaard, T. Production of N2 through anaerobic ammonium oxidation coupled to nitrate reduction in marine sediments. App. Environ. Microbiol. 2002, 68, 1312–1318. [Google Scholar] [CrossRef] [PubMed]
- Amano, T.; Yoshinaga, I.; Okada, K.; Yamagishi, T.; Ueda, S.; Obuchi, A.; Sako, Y.; Suwa, Y. Detection of anammox activity and diversity of anammox bacteria-related 16S rRNA genes in coastal marine sediment in Japan. Microbes Environ. 2007, 22, 232–242. [Google Scholar] [CrossRef]
- Amano, T.; Yoshinaga, I.; Yamagishi, T.; Thuoc, C.V.; Thu, P.T.; Ueda, S.; Kato, K.; Sako, Y.; Suwa, Y. Contribution of anammox bacteria to benthic nitrogen cycling in a mangrove forest and shrimp ponds, Haiphong, Vietnam. Microbes Environ. 2011, 26, 1–6. [Google Scholar] [CrossRef]
- Isobe, K.; Suwa, Y.; Ikutani, J.; Kuroiwa, M.; Makita, T.; Takebayashi, Y.; Yoh, M.; Otsuka, S.; Senoo, K.; Ohmori, M.; et al. Analytical techniques for quantifying 15N/14N of nitrate, nitrite, total dissolved nitrogen and ammonium in environmental samples using a gas chromatograph equipped with a quadrupole mass spectrometer. Microbes Environ. 2011, 26, 46–53. [Google Scholar] [CrossRef]
- Yoshinaga, I.; Amano, T.; Yamagishi, T.; Okada, K.; Ueda, S.; Sako, Y.; Suwa, Y. Distribution and diversity of anaerobic ammonium oxidation (anammox) bacteria in the sediment of a eutrophic freshwater lake, Lake Kitaura, Japan. Microbes Environ. 2011, 26, 189–197. [Google Scholar] [CrossRef]
- Kuroiwa, M.; Fukushima, K.; Hashimoto, K.; Senga, Y.; Sato, T.; Katsuyama, C.; Suwa, Y. Measurement of the potential rates of dissimilatory nitrate reduction to ammonium (DNRA) based on 14NH4+/15NH4+ analysis by sequential conversion and detection with quadrupole GC/MS. JoVE 2020, 164, e59562. [Google Scholar]
- Wood, E.D.; Armstrong, F.A.J.; Richards, F.A. Determination of nitrate in sea water by cadmium–copper reduction to nitrate. J. Mar. Biol. Assoc. UK 1967, 47, 23–31. [Google Scholar] [CrossRef]
- Bendschneider, K.; Robinson, R.J. A new spectrophotometric method for the determination of nitrite in seawater. J. Mar. Res. 1952, 11, 87–96. [Google Scholar]
- Sagi, T. Determination of ammonia in sea water by the indophenol method and its application to the coastal and off-shore waters. Oceanogr. Mag. 1966, 18, 43–51. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Davison, W.; Lishman, J.P. Rapid colorimetric procedure for the determination of acid volatile sulphide in sediments. Analyst 1983, 108, 1235–1239. [Google Scholar] [CrossRef]
- Sugahara, S.; Kamiya, H.; Suyama, Y.; Senga, Y.; Ayukawa, K.; Okumura, M.; Seike, Y. Influence of hypersaturated dissolved oxygenated water on the elution of hydrogen sulfide and methane from sediment in the dredged area in polyhaline Lake Nakaumi. Landsc. Ecol. Eng. 2015, 11, 269–282. [Google Scholar] [CrossRef]
- Cline, J.D. Spectrophotometric determination of hydrogen sulfide in natural water. Limnol. Oceanogr. 1969, 14, 454–458. [Google Scholar] [CrossRef]
- Weiss, R.F.; Price, B.A. Nitrous oxide solubility in water and seawater. Mar. Chem. 1980, 8, 347–359. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; ISBN 3-900051-07-0. Available online: http://www.R-project.org (accessed on 1 September 2020).
- Yin, G.; Hou, L.; Liu, M.; Li, X.; Zheng, Y.; Gao, J.; Jiang, X.; Wang, R.; Yu, C.; Lin, X. DNRA in intertidal sediments of the Yangtze Estuary. J. Geophys. Res. Biogeosci. 2017, 122, 1988–1998. [Google Scholar] [CrossRef]
- Brunet, R.C.; Garcia-Gil, L.J. Sulfide-induced dissimilatory nitrate reduction to ammonia in anaerobic freshwater sediments. FEMS Microbiol. Ecol. 1996, 21, 131–138. [Google Scholar] [CrossRef]
- Marchant, H.K.; Lavik, G.; Holtappels, M.; Kuypers, M.M.M. The fate of nitrate in intertidal permeable sediments. PLoS ONE 2014, 9, e104517. [Google Scholar] [CrossRef]
- Dong, L.F.; Smith, C.J.; Papaspyrou, S.; Osborn, A.M.; Nedwell, D.B. Changes in benthic denitrification, nitrate ammonification, and anammox process rates and nitrate and nitrite reductase gene abundances along an estuarine nutrient gradient (the Colne Estuary, United Kingdom). Appl. Environ. Microbiol. 2009, 75, 3171–3179. [Google Scholar] [CrossRef]
- Heijs, S.K.; van Gemerden, H. Microbiological and environmental variables involved in the sulfide buffering capacity along a eutrophication gradient in a coastal lagoon (Bassin d’Arcachon, France). Hydrobiologia 2000, 437, 121–131. [Google Scholar] [CrossRef]
- Sugahara, S.; Yurimoto, T.; Ayukawa, K.; Kimoto, K.; Senga, Y.; Okumura, M.; Seike, Y. A simple in situ extraction method for dissolved sulfide in sandy mud sediments followed by spectrophotometric determination and its application to the bottom sediment at the northeast of Ariake Bay. Bunseki Kagaku 2010, 59, 1155–1161, (In Japanese, with English abstract). [Google Scholar] [CrossRef]
- Senga, Y.; Mochida, K.; Fukumori, R.; Okamoto, N.; Seike, Y. N2O accumulation in estuarine and coastal sediments: The influence of H2S on dissimilatory nitrate reduction. Estuar. Coast. Shelf Sci. 2006, 67, 231–238. [Google Scholar] [CrossRef]
- Bonaglia, S.; Klawonn, I.; De Brabandere, L.; Deutsch, B.; Thamdrup, B.; Brüchert, V. Denitrification and DNRA at the Baltic Sea oxic–anoxic interface: Substrate spectrum and kinetics. Limnol. Oceanogr. 2016, 61, 1900–1915. [Google Scholar] [CrossRef]
- Li, N.; Nie, M.; Wu, M.; Wu, J. Spartina alterniflora Invasion Enhances Dissimilatory Nitrate Reduction to Ammonium (DNRA) Rates in the Yangtze River Estuary, China. J. Mar. Sci. Eng. 2022, 10, 655. [Google Scholar] [CrossRef]
- Dong, L.F.; Sobey, M.N.; Smith, C.J.; Rusmana, I.; Phillips, W.; Stott, A.; Osborn, A.M.; Nedwell, D.B. Dissimilatory reduction of nitrate to ammonium, not denitrification or anammox, dominates benthic nitrate reduction in tropical estuaries. Limnol. Oceanogr. 2011, 56, 279–291. [Google Scholar] [CrossRef]
- Kessler, A.J.; Wawryk, M.; Marzocchi, U.; Roberts, K.L.; Wong, W.W.; Risgaard-Petersen, N.; Meysman, F.J.; Glud, R.N.; Cook, P.L.M. Cable bacteria promote DNRA through iron sulfide dissolution. Limnol. Oceanogr. 2019, 64, 1228–1238. [Google Scholar] [CrossRef]
- Lomstein, B.A.; Guldberg, L.B.; Neubauer, A.T.A.; Hansen, J.; Donnelly, A.R.; Herbert, A.; Viaroli, P.; Giordani, G.; Azzoni, R.; de Wit, R.; et al. Benthic decomposition of Ulva lactuca: A controlled laboratory experiment. Aquat. Bot. 2006, 85, 271–281. [Google Scholar] [CrossRef]
- Tobias, C.R.; Anderson, I.C.; Canuel, E.A.; Macko, S.A. Nitrogen cycling through a fringing marsh-aquifer ecotone. Mar. Ecol. Prog. Ser. 2001, 210, 25–39. [Google Scholar] [CrossRef]
- Tobias, C.; Giblin, A.; McClelland, J.; Tucker, J.; Peterson, B. Sediment DIN fluxes and preferential recycling of benthic microalgal nitrogen in a shallow macrotidal estuary. Mar. Ecol. Prog. Ser. 2003, 257, 25–36. [Google Scholar] [CrossRef]
- Joye, S.B.; Hollibaugh, J.T. Influence of sulfide inhibition of nitrification on nitrogen regeneration in sediments. Science 1995, 270, 623–625. [Google Scholar] [CrossRef]
Figure 1.
Location of the Yatsu tidal flat, eastern Japan, and sampling Sts. C (35°40.34′ N, 140°00.22′ E) and S (35°40.29′ N, 140°00.27′ E) within the tidal flat.
Figure 1.
Location of the Yatsu tidal flat, eastern Japan, and sampling Sts. C (35°40.34′ N, 140°00.22′ E) and S (35°40.29′ N, 140°00.27′ E) within the tidal flat.

Figure 2.
Changes in NH4+ (a), NO2− (b), NO3− (c), and sulfide (d) concentrations in flasks with and without Ulva.
Figure 2.
Changes in NH4+ (a), NO2− (b), NO3− (c), and sulfide (d) concentrations in flasks with and without Ulva.

Figure 3.
Changes in denitrification and DNRA rates in flasks with Ulva (a) and without Ulva (b) and contributions of DNRA to the total dissimilatory NO3− reduction in each treatment (c).
Figure 3.
Changes in denitrification and DNRA rates in flasks with Ulva (a) and without Ulva (b) and contributions of DNRA to the total dissimilatory NO3− reduction in each treatment (c).

Figure 4.
The effects of different sulfide concentrations on denitrification activities in the sediment at St. C measured using the acetylene blockage method.
Figure 4.
The effects of different sulfide concentrations on denitrification activities in the sediment at St. C measured using the acetylene blockage method.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Temperature (°C); pH; water content (%); ignition loss (%); NH4+, NO2−, NO3−, and PO43− concentrations (μg-N or -P L−1); and denitrification, anammox, and DNRA rates (μg-N g-dry−1 day−1) in the surface sediments at the Yatsu tidal flat in November 2017.
Table 1.
Temperature (°C); pH; water content (%); ignition loss (%); NH4+, NO2−, NO3−, and PO43− concentrations (μg-N or -P L−1); and denitrification, anammox, and DNRA rates (μg-N g-dry−1 day−1) in the surface sediments at the Yatsu tidal flat in November 2017.
| St. C | St. S | |
|---|---|---|
| Temperature | 17.3 | 17.9 |
| pH | 7.63 | 6.83 |
| Water content | 39.7 | 18.4 |
| Ignition loss | 1.4 | 0.9 |
| NH4+ | 4549 | |
| NO2− | 5 | |
| NO3− | 5 | |
| PO43− | 164 | |
| Denitrification | 10.4 | 5.9 |
| Anammox | nd | nd |
| DNRA | 2.3 | 1.0 |
Notes: The NH4+, NO2−, NO3−, and PO43− concentrations at St. S could not be measured due to the low water content. The denitrification, anammox, and DNRA rates were the averages of three measurements.
Table 2.
Estimated, initial, and final sulfide concentrations (mg-S L−1) to determine the effect of sulfide on denitrification. The initial and final concentrations were measured on days 0 and 21, respectively.
Table 2.
Estimated, initial, and final sulfide concentrations (mg-S L−1) to determine the effect of sulfide on denitrification. The initial and final concentrations were measured on days 0 and 21, respectively.
| Estimated Concentration | Initial Concentration | Final Concentration |
|---|---|---|
| 0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 5 | 4.1 ± 1.7 | 0.0 ± 0.0 |
| 10 | 21.1 ± 7.3 | 0.0 ± 0.0 |
| 50 | 54.9 ± 6.8 | 15.1 ± 2.7 |
| 100 | 107.5 ± 10.4 | 65.5 ± 2.0 |
| 200 | 211.9 ± 10.5 | 95.8 ± 32.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Senga, Y.; Sato, T.; Shibaki, K.; Kuroiwa, M.; Nohara, S.; Suwa, Y. Alteration of Dissimilatory Nitrate Reduction Pathways in the Intertidal Sediment during Macroalgae Blooms. Water 2022, 14, 3022. https://doi.org/10.3390/w14193022
AMA Style
Senga Y, Sato T, Shibaki K, Kuroiwa M, Nohara S, Suwa Y. Alteration of Dissimilatory Nitrate Reduction Pathways in the Intertidal Sediment during Macroalgae Blooms. Water. 2022; 14(19):3022. https://doi.org/10.3390/w14193022
Chicago/Turabian StyleSenga, Yukiko, Tsubasa Sato, Kanae Shibaki, Megumi Kuroiwa, Seiichi Nohara, and Yuichi Suwa. 2022. "Alteration of Dissimilatory Nitrate Reduction Pathways in the Intertidal Sediment during Macroalgae Blooms" Water 14, no. 19: 3022. https://doi.org/10.3390/w14193022
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.