
Impact of Environmental Conditions and Seasonality on Ecosystem Transpiration and Evapotranspiration Partitioning (T/ET Ratio) of Pure European Beech Forest

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Information
2.2. Energy Balance
2.3. Eddy-Covariance and Meteorological Measurements
2.4. Sap Flow Measurements and Upscaling
2.5. Statistical Analysis
3. Results
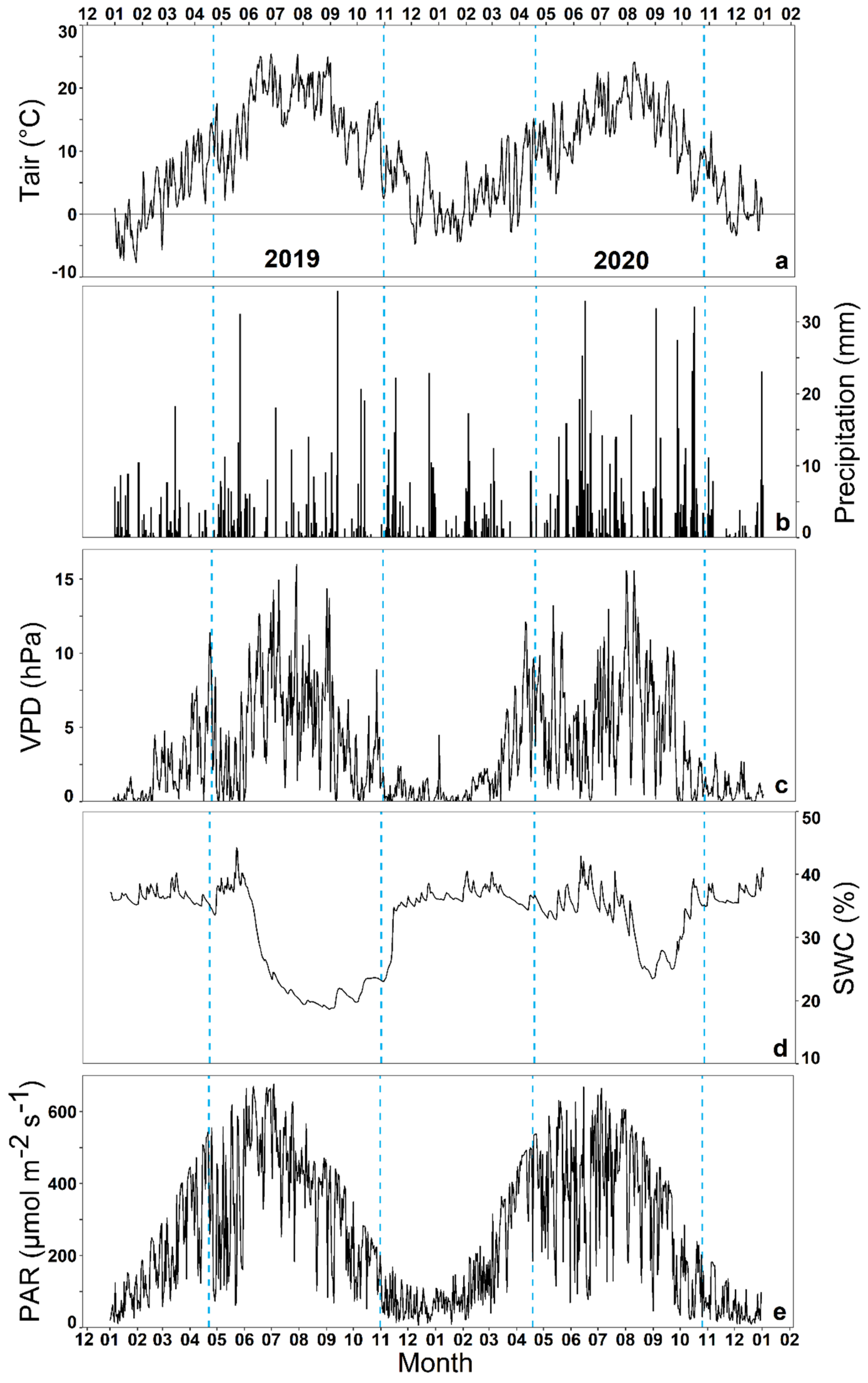
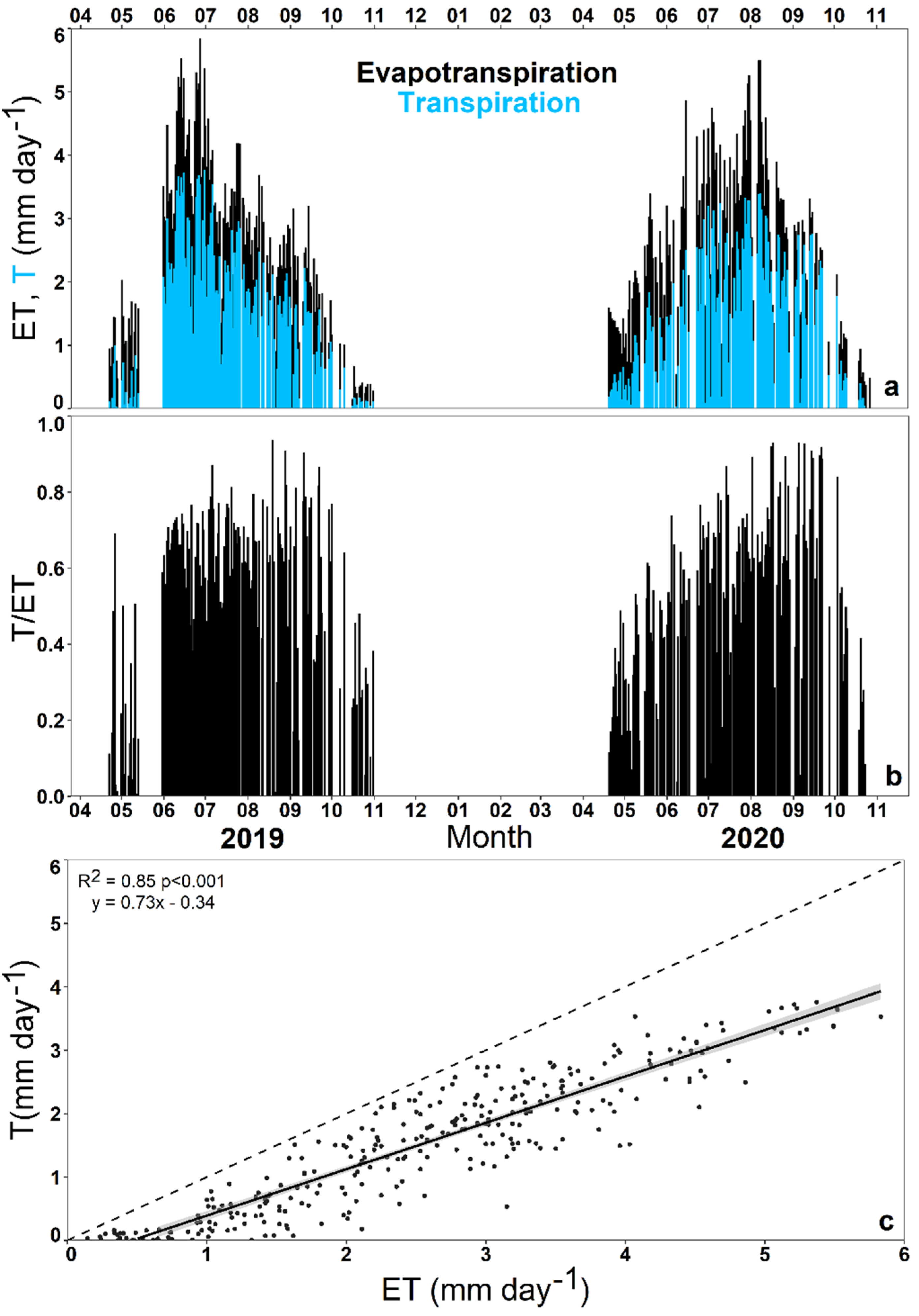
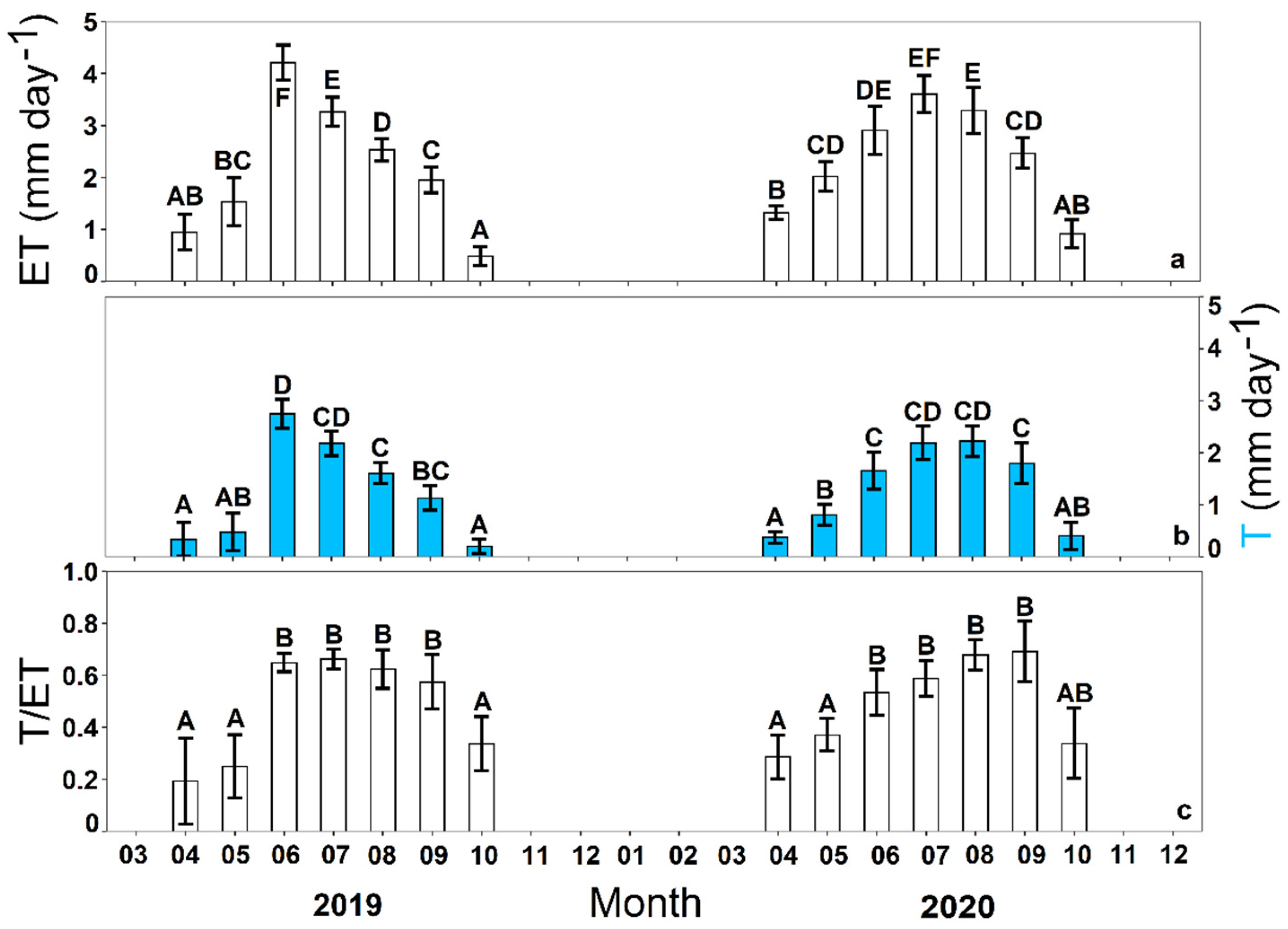
3.1. Seasonal Dynamics of Ecosystem Transpiration and Evapotranspiration
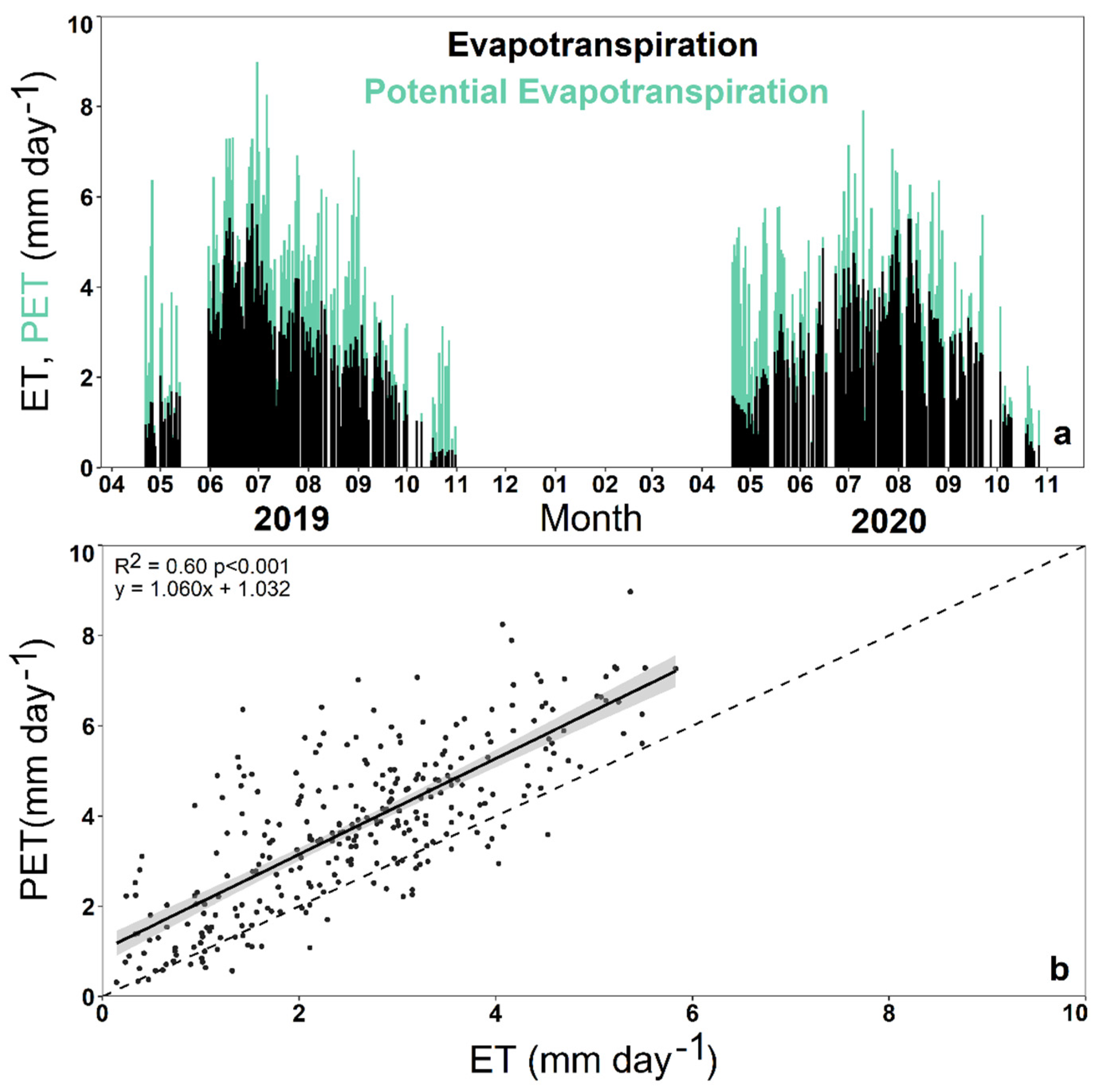
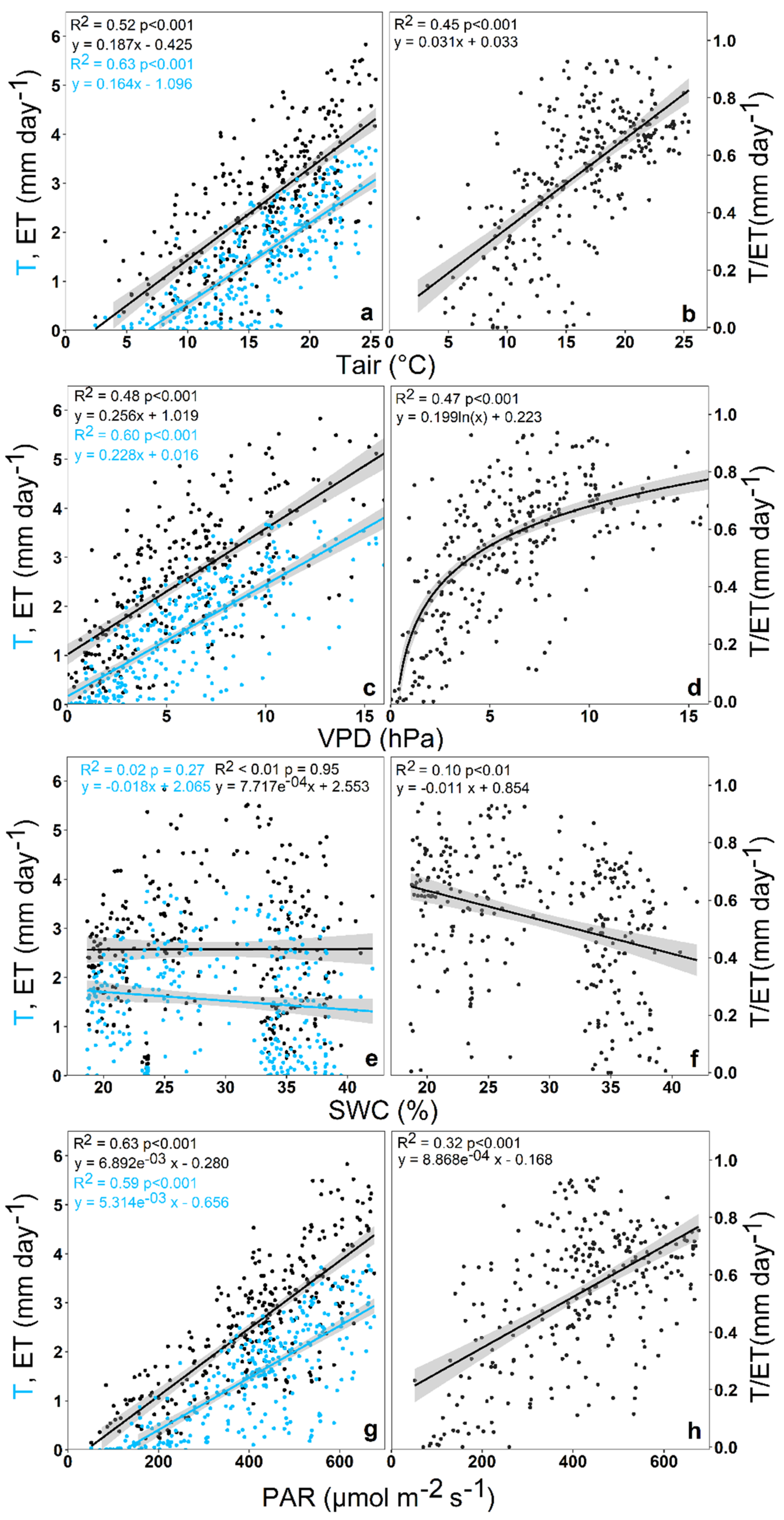
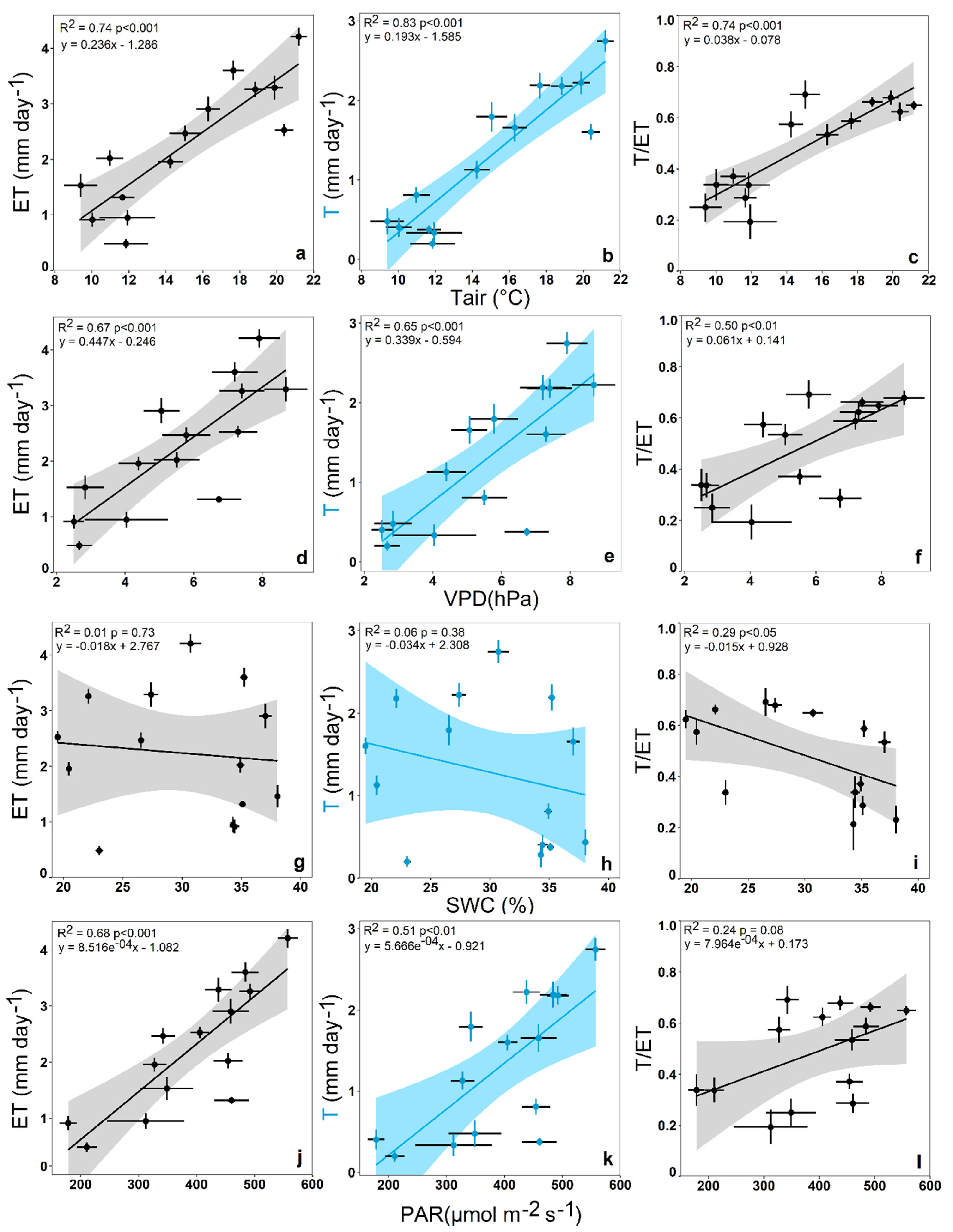
3.2. Impact of Environmental Variables on Ecosystem Transpiration and Evapotranspiration
4. Discussion
4.1. Seasonal Changes of Ecosystem Transpiration and Evapotranspiration Partitioning
4.2. Environmental Impact on Ecosystem Transpiration and Evapotranspiration
4.3. Environmental Impact on T/ET Ratio
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dimitriadou, S.; Nikolakopoulos, K.G. Evapotranspiration Trends and Interactions in Light of the Anthropogenic Footprint and the Climate Crisis: A Review. Hydrology 2021, 8, 163. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Pan, Y.; Hu, R.; Chen, N. Global Quantitative Synthesis of Effects of Biotic and Abiotic Factors on Stemflow Production in Woody Ecosystems. Global Ecol. Biogeogr. 2021, 30, 1713–1723. [Google Scholar] [CrossRef]
- Nelson, J.A.; Pérez-Priego, O.; Zhou, S.; Poyatos, R.; Zhang, Y.; Blanken, P.D.; Gimeno, T.E.; Wohlfahrt, G.; Desai, A.R.; Gioli, B.; et al. Ecosystem Transpiration and Evaporation: Insights from Three Water Flux Partitioning Methods across FLUXNET Sites. Glob. Chang. Biol. 2020, 26, 6916–6930. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J.B.; Valle Junior, L.C.G.; Faria, T.O.; Marques, J.B.; Dalmagro, H.J.; Nogueira, J.S.; Vourlitis, G.L.; Rodrigues, T.R. Temporal Variability in Evapotranspiration and Energy Partitioning over a Seasonally Flooded Scrub Forest of the Brazilian Pantanal. Agric. For. Meteorol. 2021, 308–309, 108559. [Google Scholar] [CrossRef]
- Paul-Limoges, E.; Revill, A.; Maier, R.; Buchmann, N.; Damm, A. Insights for the Partitioning of Ecosystem Evaporation and Transpiration in Short-Statured Croplands. JGR Biogeosci. 2022, 127, e2021JG006760. [Google Scholar] [CrossRef]
- Nie, C.; Huang, Y.; Zhang, S.; Yang, Y.; Zhou, S.; Lin, C.; Wang, G. Effects of Soil Water Content on Forest Ecosystem Water Use Efficiency through Changes in Transpiration/Evapotranspiration Ratio. Agric. For. Meteorol. 2021, 308–309, 108605. [Google Scholar] [CrossRef]
- Sun, H.; Wang, P.; Chen, Q.; Zhang, D.; Xing, Y. Coupling the Water Use of Populus Euphratica and Tamarix Ramosissima and Evapotranspiration Partitioning in a Desert Riparian Forest Ecosystem. Agric. For. Meteorol. 2022, 323, 109064. [Google Scholar] [CrossRef]
- Winbourne, J.B.; Jones, T.S.; Garvey, S.M.; Harrison, J.L.; Wang, L.; Li, D.; Templer, P.H.; Hutyra, L.R. Tree Transpiration and Urban Temperatures: Current Understanding, Implications, and Future Research Directions. BioScience 2020, 70, 576–588. [Google Scholar] [CrossRef]
- Grossiord, C.; Forner, A.; Gessler, A.; Granier, A.; Pollastrini, M.; Valladares, F.; Bonal, D. Influence of Species Interactions on Transpiration of Mediterranean Tree Species during a Summer Drought. Eur. J. For. Res. 2015, 134, 365–376. [Google Scholar] [CrossRef]
- Forrester, D.I. Transpiration and Water-Use Efficiency in Mixed-Species Forests versus Monocultures: Effects of Tree Size, Stand Density and Season. Tree Physiol. 2015, 35, 289–304. [Google Scholar] [CrossRef] [Green Version]
- Kirschbaum, M.U.F.; McMillan, A.M.S. Warming and Elevated CO2 Have Opposing Influences on Transpiration. Which Is More Important? Curr. For. Rep. 2018, 4, 51–71. [Google Scholar] [CrossRef]
- Alvarado-Barrientos, M.S.; Holwerda, F.; Geissert, D.R.; Muñoz-Villers, L.E.; Gotsch, S.G.; Asbjornsen, H.; Dawson, T.E. Nighttime Transpiration in a Seasonally Dry Tropical Montane Cloud Forest Environment. Trees 2015, 29, 259–274. [Google Scholar] [CrossRef]
- Höök, M.; Tang, X. Depletion of Fossil Fuels and Anthropogenic Climate Change—A Review. Energy Policy 2013, 52, 797–809. [Google Scholar] [CrossRef]
- Engineer, C.B.; Hashimoto-Sugimoto, M.; Negi, J.; Israelsson-Nordström, M.; Azoulay-Shemer, T.; Rappel, W.-J.; Iba, K.; Schroeder, J.I. CO2 Sensing and CO2 Regulation of Stomatal Conductance: Advances and Open Questions. Trends Plant Sci. 2016, 21, 16–30. [Google Scholar] [CrossRef]
- Šigut, L.; Holiova, P.; Klem, K.; Prtova, M.; Calfapietra, C.; Marek, M.V.; Punda, V.; Urban, O. Does Long-Term Cultivation of Saplings under Elevated CO2 Concentration Influence Their Photosynthetic Response to Temperature? Ann. Bot. 2015, 116, 929–939. [Google Scholar] [CrossRef]
- Monteiro, M.V.; Blanuša, T.; Verhoef, A.; Hadley, P.; Cameron, R.W.F. Relative Importance of Transpiration Rate and Leaf Morphological Traits for the Regulation of Leaf Temperature. Aust. J. Bot. 2016, 64, 32. [Google Scholar] [CrossRef]
- Markulj Kulundžić, A.; Kovačević, J.; Viljevac Vuletić, M.; Josipović, A.; Liović, I.; Mijić, A.; Lepeduš, H.; Matoša Kočar, M. Impact of Abiotic Stress on Photosynthetic Efficiency and Leaf Temperature in Sunflower. Poljoprivreda 2016, 22, 17–22. [Google Scholar] [CrossRef]
- Ruehr, N.K.; Gast, A.; Weber, C.; Daub, B.; Arneth, A. Water Availability as Dominant Control of Heat Stress Responses in Two Contrasting Tree Species. Tree Physiol. 2015, 36, 164–178. [Google Scholar] [CrossRef]
- Rossi, S.; Burgess, P.; Jespersen, D.; Huang, B. Heat-Induced Leaf Senescence Associated with Chlorophyll Metabolism in Bentgrass Lines Differing in Heat Tolerance. Crop Sci. 2017, 57, S-169–S-178. [Google Scholar] [CrossRef]
- Mariën, B.; Dox, I.; De Boeck, H.J.; Willems, P.; Leys, S.; Papadimitriou, D.; Campioli, M. Does Drought Advance the Onset of Autumn Leaf Senescence in Temperate Deciduous Forest Trees? Biogeosciences 2021, 18, 3309–3330. [Google Scholar] [CrossRef]
- Frank, D.C.; Poulter, B.; Saurer, M.; Esper, J.; Huntingford, C.; Helle, G.; Treydte, K.; Zimmermann, N.E.; Schleser, G.H.; Ahlström, A.; et al. Water-Use Efficiency and Transpiration across European Forests during the Anthropocene. Nat. Clim. Chang. 2015, 5, 579–583. [Google Scholar] [CrossRef]
- Novick, K.A.; Ficklin, D.L.; Stoy, P.C.; Williams, C.A.; Bohrer, G.; Oishi, A.C.; Papuga, S.A.; Blanken, P.D.; Noormets, A.; Sulman, B.N.; et al. The Increasing Importance of Atmospheric Demand for Ecosystem Water and Carbon Fluxes. Nat. Clim. Chang. 2016, 6, 1023–1027. [Google Scholar] [CrossRef]
- Martens, C.; Hickler, T.; Davis-Reddy, C.; Engelbrecht, F.; Higgins, S.I.; Maltitz, G.P.; Midgley, G.F.; Pfeiffer, M.; Scheiter, S. Large Uncertainties in Future Biome Changes in Africa Call for Flexible Climate Adaptation Strategies. Glob. Chang. Biol. 2021, 27, 340–358. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Serrano, S.M.; Miralles, D.G.; McDowell, N.; Brodribb, T.; Domínguez-Castro, F.; Leung, R.; Koppa, A. The Uncertain Role of Rising Atmospheric CO2 on Global Plant Transpiration. Earth-Sci. Rev. 2022, 230, 104055. [Google Scholar] [CrossRef]
- Ben-Gal, A.; Karlberg, L.; Jansson, P.-E.; Shani, U. Temporal Robustness of Linear Relationships between Production and Transpiration. Plant Soil 2003, 251, 211–218. [Google Scholar] [CrossRef]
- Tardieu, F.; Parent, B. Predictable ‘Meta-Mechanisms’ Emerge from Feedbacks between Transpiration and Plant Growth and Cannot Be Simply Deduced from Short-Term Mechanisms: Meta-Mechanisms in Plant Water Relations. Plant Cell Environ. 2017, 40, 846–857. [Google Scholar] [CrossRef]
- Quan, Q.; Zhang, F.; Tian, D.; Zhou, Q.; Wang, L.; Niu, S. Transpiration Dominates Ecosystem Water-Use Efficiency in Response to Warming in an Alpine Meadow. J. Geophys. Res. Biogeosci. 2018, 123, 453–462. [Google Scholar] [CrossRef]
- Fricke, W. Night-Time Transpiration—Favouring Growth? Trends Plant Sci. 2019, 24, 311–317. [Google Scholar] [CrossRef]
- Gu, C.; Tang, Q.; Zhu, G.; Ma, J.; Gu, C.; Zhang, K.; Sun, S.; Yu, Q.; Niu, S. Discrepant Responses between Evapotranspiration- and Transpiration-Based Ecosystem Water Use Efficiency to Interannual Precipitation Fluctuations. Agric. For. Meteorol. 2021, 303, 108385. [Google Scholar] [CrossRef]
- An, X. Responses of Water Use Efficiency to Climate Change in Evapotranspiration and Transpiration Ecosystems. Ecol. Indic. 2022, 141, 109157. [Google Scholar] [CrossRef]
- Köstner, B.; Granier, A.; Cermák, J. Sapflow Measurements in Forest Stands: Methods and Uncertainties. Ann. For. Sci. 1998, 55, 13–27. [Google Scholar] [CrossRef]
- Flo, V.; Martinez-Vilalta, J.; Steppe, K.; Schuldt, B.; Poyatos, R. A Synthesis of Bias and Uncertainty in Sap Flow Methods. Agric. For. Meteorol. 2019, 271, 362–374. [Google Scholar] [CrossRef]
- Poyatos, R.; Granda, V.; Flo, V.; Adams, M.A.; Adorján, B.; Aguadé, D.; Aidar, M.P.M.; Allen, S.; Alvarado-Barrientos, M.S.; Anderson-Teixeira, K.J.; et al. Global Transpiration Data from Sap Flow Measurements: The SAPFLUXNET Database. Earth Syst. Sci. Data 2021, 13, 2607–2649. [Google Scholar] [CrossRef]
- Xiao, W.; Wei, Z.; Wen, X. Evapotranspiration Partitioning at the Ecosystem Scale Using the Stable Isotope Method—A Review. Agric. For. Meteorol. 2018, 263, 346–361. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.; Shan, N.; Zhang, Z.; Wei, Z. Global Assessment of Partitioning Transpiration from Evapotranspiration Based on Satellite Solar-Induced Chlorophyll Fluorescence Data. J. Hydrol. 2022, 612, 128044. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Jasechko, S. Transpiration in the Global Water Cycle. Agric. For. Meteorol. 2014, 189–190, 115–117. [Google Scholar] [CrossRef]
- Kool, D.; Agam, N.; Lazarovitch, N.; Heitman, J.L.; Sauer, T.J.; Ben-Gal, A. A Review of Approaches for Evapotranspiration Partitioning. Agric. For. Meteorol. 2014, 184, 56–70. [Google Scholar] [CrossRef]
- Hadiwijaya, B.; Pepin, S.; Isabelle, P.-E.; Nadeau, D.F. The Dynamics of Transpiration to Evapotranspiration Ratio under Wet and Dry Canopy Conditions in a Humid Boreal Forest. Forests 2020, 11, 237. [Google Scholar] [CrossRef]
- Cao, R.; Huang, H.; Wu, G.; Han, D.; Jiang, Z.; Di, K.; Hu, Z. Spatiotemporal Variations in the Ratio of Transpiration to Evapotranspiration and Its Controlling Factors across Terrestrial Biomes. Agric. For. Meteorol. 2022, 321, 108984. [Google Scholar] [CrossRef]
- Raz-Yaseef, N.; Yakir, D.; Schiller, G.; Cohen, S. Dynamics of Evapotranspiration Partitioning in a Semi-Arid Forest as Affected by Temporal Rainfall Patterns. Agric. For. Meteorol. 2012, 157, 77–85. [Google Scholar] [CrossRef]
- Ungar, E.D.; Rotenberg, E.; Raz-Yaseef, N.; Cohen, S.; Yakir, D.; Schiller, G. Transpiration and Annual Water Balance of Aleppo Pine in a Semiarid Region: Implications for Forest Management. For. Ecol. Manag. 2013, 298, 39–51. [Google Scholar] [CrossRef]
- Zhou, S.; Yu, B.; Zhang, Y.; Huang, Y.; Wang, G. Water Use Efficiency and Evapotranspiration Partitioning for Three Typical Ecosystems in the Heihe River Basin, Northwestern China. Agric. For. Meteorol. 2018, 253–254, 261–273. [Google Scholar] [CrossRef]
- Gao, G.; Wang, D.; Zha, T.; Wang, L.; Fu, B. A Global Synthesis of Transpiration Rate and Evapotranspiration Partitioning in the Shrub Ecosystems. J. Hydrol. 2022, 606, 127417. [Google Scholar] [CrossRef]
- Jia, G.; Liu, Z.; Chen, L.; Yu, X. Distinguish Water Utilization Strategies of Trees Growing on Earth-Rocky Mountainous Area with Transpiration and Water Isotopes. Ecol. Evol. 2017, 7, 10640–10651. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, L.M.; Garg, A. Investigating Plant Transpiration-Induced Soil Suction Affected by Root Morphology and Root Depth. Comput. Geotech. 2018, 103, 26–31. [Google Scholar] [CrossRef]
- Wu, W.; Li, H.; Feng, H.; Si, B.; Chen, G.; Meng, T.; Li, Y.; Siddique, K.H.M. Precipitation Dominates the Transpiration of Both the Economic Forest (Malus pumila) and Ecological Forest (Robinia pseudoacacia) on the Loess Plateau after about 15 Years of Water Depletion in Deep Soil. Agric. For. Meteorol. 2021, 297, 108244. [Google Scholar] [CrossRef]
- Lüttschwager, D.; Jochheim, H. Drought Primarily Reduces Canopy Transpiration of Exposed Beech Trees and Decreases the Share of Water Uptake from Deeper Soil Layers. Forests 2020, 11, 537. [Google Scholar] [CrossRef]
- Nalevanková, P.; Ježík, M.; Sitková, Z.; Vido, J.; Leštianska, A.; Střelcová, K. Drought and Irrigation Affect Transpiration Rate and Morning Tree Water Status of a Mature European Beech ( Fagus Sylvatica L.) Forest in Central Europe: Ecophysiology of European Beech Exposed to Drought. Ecohydrology 2018, 11, e1958. [Google Scholar] [CrossRef]
- Leuschner, C.; Schipka, F.; Backes, K. Stomatal Regulation and Water Potential Variation in European Beech: Challenging the Iso/Anisohydry Concept. Tree Physiol. 2022, 42, 365–378. [Google Scholar] [CrossRef]
- Nalevanková, P.; Sitková, Z.; Kučera, J.; Střelcová, K. Impact of Water Deficit on Seasonal and Diurnal Dynamics of European Beech Transpiration and Time-Lag Effect between Stand Transpiration and Environmental Drivers. Water 2020, 12, 3437. [Google Scholar] [CrossRef]
- Zhu, X.; Yu, G.; Wang, Q.; Hu, Z.; Han, S.; Yan, J.; Wang, Y.; Zhao, L. Seasonal Dynamics of Water Use Efficiency of Typical Forest and Grassland Ecosystems in China. J. For. Res. 2014, 19, 70–76. [Google Scholar] [CrossRef]
- Nguyen, M.N.; Hao, Y.; Baik, J.; Choi, M. Partitioning Evapotranspiration Based on the Total Ecosystem Conductance Fractions of Soil, Interception, and Canopy in Different Biomes. J. Hydrol. 2021, 603, 126970. [Google Scholar] [CrossRef]
- Lai, J.; Liu, T.; Luo, Y. Evapotranspiration Partitioning for Winter Wheat with Shallow Groundwater in the Lower Reach of the Yellow River Basin. Agric. Water Manag. 2022, 266, 107561. [Google Scholar] [CrossRef]
- Jasechko, S.; Sharp, Z.D.; Gibson, J.J.; Birks, S.J.; Yi, Y.; Fawcett, P.J. Terrestrial Water Fluxes Dominated by Transpiration. Nature 2013, 496, 347–350. [Google Scholar] [CrossRef]
- Coenders-Gerrits, A.M.J.; van der Ent, R.J.; Bogaard, T.A.; Wang-Erlandsson, L.; Hrachowitz, M.; Savenije, H.H.G. Uncertainties in Transpiration Estimates. Nature 2014, 506, E1–E2. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.G.; Cable, W.; Hultine, K.; Hoedjes, J.C.B.; Yepez, E.A.; Simonneaux, V.; Er-Raki, S.; Boulet, G.; de Bruin, H.A.R.; Chehbouni, A.; et al. Evapotranspiration Components Determined by Stable Isotope, Sap Flow and Eddy Covariance Techniques. Agric. For. Meteorol. 2004, 125, 241–258. [Google Scholar] [CrossRef]
- Wei, Z.; Lee, X.; Wen, X.; Xiao, W. Evapotranspiration Partitioning for Three Agro-Ecosystems with Contrasting Moisture Conditions: A Comparison of an Isotope Method and a Two-Source Model Calculation. Agric. For. Meteorol. 2018, 252, 296–310. [Google Scholar] [CrossRef]
- Niu, Z.; He, H.; Zhu, G.; Ren, X.; Zhang, L.; Zhang, K.; Yu, G.; Ge, R.; Li, P.; Zeng, N.; et al. An Increasing Trend in the Ratio of Transpiration to Total Terrestrial Evapotranspiration in China from 1982 to 2015 Caused by Greening and Warming. Agric. For. Meteorol. 2019, 279, 107701. [Google Scholar] [CrossRef]
- Li, M.; Wu, P.; Ma, Z.; Pan, Z.; Lv, M.; Yang, Q.; Duan, Y. The Increasing Role of Vegetation Transpiration in Soil Moisture Loss across China under Global Warming. J. Hydrometeorol. 2022, 23, 253–274. [Google Scholar] [CrossRef]
- Mauder, M.; Cuntz, M.; Drüe, C.; Graf, A.; Rebmann, C.; Schmid, H.P.; Schmidt, M.; Steinbrecher, R. A Strategy for Quality and Uncertainty Assessment of Long-Term Eddy-Covariance Measurements. Agric. For. Meteorol. 2013, 169, 122–135. [Google Scholar] [CrossRef]
- Schotanus, P.; Nieuwstadt, F.T.M.; De Bruin, H.A.R. Temperature Measurement with a Sonic Anemometer and Its Application to Heat and Moisture Fluxes. Bound.-Layer Meteorol. 1983, 26, 81–93. [Google Scholar] [CrossRef]
- Horst, T.W.; Lenschow, D.H. Attenuation of Scalar Fluxes Measured with Spatially-Displaced Sensors. Bound.-Layer Meteorol. 2009, 130, 275–300. [Google Scholar] [CrossRef]
- Ibrom, A.; Dellwik, E.; Flyvbjerg, H.; Jensen, N.O.; Pilegaard, K. Strong Low-Pass Filtering Effects on Water Vapour Flux Measurements with Closed-Path Eddy Correlation Systems. Agric. For. Meteorol. 2007, 147, 140–156. [Google Scholar] [CrossRef]
- Moncrieff, J.; Clement, R.; Finnigan, J.; Meyers, T. Averaging, Detrending, and Filtering of Eddy Covariance Time Series. In Handbook of Micrometeorology; Lee, X., Massman, W., Law, B., Eds.; Atmospheric and Oceanographic Sciences Library; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2005; Volume 29, pp. 7–31. ISBN 978-1-4020-2264-7. [Google Scholar]
- Wilczak, J.M.; Oncley, S.P.; Stage, S.A. Sonic Anemometer Tilt Correction Algorithms. Bound.-Layer Meteorol. 2001, 99, 127–150. [Google Scholar] [CrossRef]
- McGloin, R.; Šigut, L.; Havránková, K.; Dušek, J.; Pavelka, M.; Sedlák, P. Energy Balance Closure at a Variety of Ecosystems in Central Europe with Contrasting Topographies. Agric. For. Meteorol. 2018, 248, 418–431. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration—Guidelines for computing crop water requirements -FAO Irrigation and drainage paper No. 56. Rome: Food and Agriculture Organization of the United Nations. Fao Rome 1998, 56, e156. [Google Scholar]
- Kučera, J.; Brito, P.; Jiménez, M.S.; Urban, J. Direct Penman–Monteith Parameterization for Estimating Stomatal Conductance and Modeling Sap Flow. Trees 2017, 31, 873–885. [Google Scholar] [CrossRef]
- Čermák, J.; Kučera, J.; Nadezhdina, N. Sap Flow Measurements with Some Thermodynamic Methods, Flow Integration within Trees and Scaling up from Sample Trees to Entire Forest Stands. Trees 2004, 18, 529–546. [Google Scholar] [CrossRef]
- Lindroth, A.; Cermak, J.; Kucera, J.; Cienciala, E.; Eckersten, H. Sap Flow by the Heat Balance Method Applied to Small Size Salix Trees in a Short-Rotation Forest. Biomass Bioenergy 1995, 8, 7–15. [Google Scholar] [CrossRef]
- Urban, J.; Rubtsov, A.V.; Urban, A.V.; Shashkin, A.V.; Benkova, V.E. Canopy Transpiration of a Larix Sibirica and Pinus Sylvestris Forest in Central Siberia. Agric. For. Meteorol. 2019, 271, 64–72. [Google Scholar] [CrossRef]
- Davi, H.; Dufrêne, E.; Granier, A.; Le Dantec, V.; Barbaroux, C.; François, C.; Bréda, N. Modelling Carbon and Water Cycles in a Beech Forest. Ecol. Model. 2005, 185, 387–405. [Google Scholar] [CrossRef] [Green Version]
- Urban, J.; Bednárová, E.; Plichta, R.; Gryc, V.; Vavrčík, H.; Hacura, J.; Fajstavr, M.; Kučera, J. Links between Phenology and Ecophysiology in a European Beech Forest. iForest 2015, 8, 438–447. [Google Scholar] [CrossRef]
- Lavrič, M.; Eler, K.; Ferlan, M.; Vodnik, D.; Gričar, J. Chronological Sequence of Leaf Phenology, Xylem and Phloem Formation and Sap Flow of Quercus Pubescens from Abandoned Karst Grasslands. Front. Plant Sci. 2017, 8, 314. [Google Scholar] [CrossRef] [PubMed]
- Hovenden, M.J. The Influence of Temperature and Genotype on the Growth and Stomatal Morphology of Southern Beech, Nothofagus Cunninghamii (Nothofagaceae). Aust. J. Bot. 2001, 49, 427. [Google Scholar] [CrossRef]
- Spiegelhalder, R.P.; Raissig, M.T. Morphology Made for Movement: Formation of Diverse Stomatal Guard Cells. Curr. Opin. Plant Biol. 2021, 63, 102090. [Google Scholar] [CrossRef]
- Wilson, K.B.; Hanson, P.J.; Mulholland, P.J.; Baldocchi, D.D.; Wullschleger, S.D. A Comparison of Methods for Determining Forest Evapotranspiration and Its Components: Sap-Flow, Soil Water Budget, Eddy Covariance and Catchment Water Balance. Agric. For. Meteorol. 2001, 106, 153–168. [Google Scholar] [CrossRef]
- Aranda, I.; Gil, L.; Pardos, J.A. Seasonal Changes in Apparent Hydraulic Conductance and Their Implications for Water Use of European Beech (Fagus sylvatica L.) and Sessile Oak [Quercus petraea (Matt.) Liebl] in South Europe. Plant Ecol. 2005, 179, 155–167. [Google Scholar] [CrossRef]
- Liu, J.; Cheng, F.; Munger, W.; Jiang, P.; Whitby, T.G.; Chen, S.; Ji, W.; Man, X. Precipitation Extremes Influence Patterns and Partitioning of Evapotranspiration and Transpiration in a Deciduous Boreal Larch Forest. Agric. For. Meteorol. 2020, 287, 107936. [Google Scholar] [CrossRef]
- Paul-Limoges, E.; Wolf, S.; Schneider, F.D.; Longo, M.; Moorcroft, P.; Gharun, M.; Damm, A. Partitioning Evapotranspiration with Concurrent Eddy Covariance Measurements in a Mixed Forest. Agric. For. Meteorol. 2020, 280, 107786. [Google Scholar] [CrossRef]
- Kubota, T.; Kagawa, A.; Shichi, K.; Ono, K. The Promotional Effect of Increased Growth on Transpiration Exceeds the Inhibitory Effect of Increased Water Use Efficiency over the Life History of Fagus Crenata Trees. J. For. Res. 2022, 27, 1–11. [Google Scholar] [CrossRef]
- Fisher, R.A.; Williams, M.; Do Vale, R.L.; Da Costa, A.L.; Meir, P. Evidence from Amazonian Forests Is Consistent with Isohydric Control of Leaf Water Potential. Plant Cell Environ. 2006, 29, 151–165. [Google Scholar] [CrossRef]
- Yi, K.; Dragoni, D.; Phillips, R.P.; Roman, D.T.; Novick, K.A. Dynamics of Stem Water Uptake among Isohydric and Anisohydric Species Experiencing a Severe Drought. Tree Physiol. 2017, 37, 1379–1392. [Google Scholar] [CrossRef] [PubMed]
- Hesse, B.D.; Gebhardt, T.; Hafner, B.D.; Hikino, K.; Häberle, K.-H.; Grams, T.E.E. High Resilience of Water Related Physiology after Five Years of Repeated Summer Drought of Mature Beech and Spruce. Copernic. Meet. 2022, No. EGU22-2835. [Google Scholar] [CrossRef]
- Liu, X.; Biondi, F. Transpiration Drivers of High-Elevation Five-Needle Pines (Pinus longaeva and Pinus flexilis) in Sky-Island Ecosystems of the North American Great Basin. Sci. Total Environ. 2020, 739, 139861. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Tian, Q.; He, Z.; Du, J.; Chen, L.; Lin, P.; Zhu, X. Response of Sap Flow in Qinghai Spruce (Picea crassifolia) to Environmental Variables in the Qilian Mountains of China. Trees 2022, 36, 1261–1272. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Y.; An, J.; Liu, Q.; Lang, Y. Sap Flow of Black Locust in Response to Environmental Factors in Two Soils Developed from Different Parent Materials in the Lithoid Mountainous Area of North China. Trees 2018, 32, 675–688. [Google Scholar] [CrossRef]
- Dolschak, K.; Gartner, K.; Berger, T.W. The Impact of Rising Temperatures on Water Balance and Phenology of European Beech (Fagus sylvatica L.) Stands. Model. Earth Syst. Environ. 2019, 5, 1347–1363. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, X.; Xu, M.; Wen, R.; Jia, Q.; Xie, Y.; Ma, H. Evapotranspiration Dynamics and Their Drivers in a Temperate Mixed Forest in Northeast China. PeerJ 2022, 10, e13549. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.; Ingwers, M.; McGuire, M.A.; Teskey, R.O. Stomatal Conductance Increases with Rising Temperature. Plant Signal. Behav. 2017, 12, e1356534. [Google Scholar] [CrossRef]
- Jiang, S.; Liang, C.; Cui, N.; Zhao, L.; Liu, C.; Feng, Y.; Hu, X.; Gong, D.; Zou, Q. Water Use Efficiency and Its Drivers in Four Typical Agroecosystems Based on Flux Tower Measurements. Agric. For. Meteorol. 2020, 295, 108200. [Google Scholar] [CrossRef]
- Zhu, X.-J.; Yu, G.-R.; Hu, Z.-M.; Wang, Q.-F.; He, H.-L.; Yan, J.-H.; Wang, H.-M.; Zhang, J.-H. Spatiotemporal Variations of T/ET (the Ratio of Transpiration to Evapotranspiration) in Three Forests of Eastern China. Ecol. Indic. 2015, 52, 411–421. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | T air | P | VPD | T air (3–11) | P (3–11) | VPD (3–11) |
|---|---|---|---|---|---|---|
| (°C) | (mm) | (hPa) | (°C) | (mm) | (hPa) | |
| 2019 | 9.91 | 653 | 3.36 | 13.88 | 427 | 4.66 |
| 2020 | 8.86 | 835 | 3.54 | 12.46 | 669 | 4.92 |
| 2009–2016 | 9.26 | 688 | 3.55 | 12.29 | 571 | 4.44 |
| DF | SumSQ | MeanSQ | F | p | |
|---|---|---|---|---|---|
| Evapotranspiration | |||||
| Month | 13 | 311.8 | 23.986 | 40.93 | <0.001 |
| Residuals | 277 | 162.3 | 0.586 | ||
| Transpiration | |||||
| Month | 13 | 184.4 | 14.19 | 33.78 | <0.001 |
| Residuals | 277 | 116.3 | 0.42 | ||
| Transpiration/Evapotranspiration | |||||
| Month | 13 | 6.474 | 0.498 | 15.92 | <0.001 |
| Residuals | 277 | 8.663 | 0.031 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrík, P.; Zavadilová, I.; Šigut, L.; Kowalska, N.; Petek-Petrik, A.; Szatniewska, J.; Jocher, G.; Pavelka, M. Impact of Environmental Conditions and Seasonality on Ecosystem Transpiration and Evapotranspiration Partitioning (T/ET Ratio) of Pure European Beech Forest. Water 2022, 14, 3015. https://doi.org/10.3390/w14193015
Petrík P, Zavadilová I, Šigut L, Kowalska N, Petek-Petrik A, Szatniewska J, Jocher G, Pavelka M. Impact of Environmental Conditions and Seasonality on Ecosystem Transpiration and Evapotranspiration Partitioning (T/ET Ratio) of Pure European Beech Forest. Water. 2022; 14(19):3015. https://doi.org/10.3390/w14193015
Chicago/Turabian StylePetrík, Peter, Ina Zavadilová, Ladislav Šigut, Natalia Kowalska, Anja Petek-Petrik, Justyna Szatniewska, Georg Jocher, and Marian Pavelka. 2022. "Impact of Environmental Conditions and Seasonality on Ecosystem Transpiration and Evapotranspiration Partitioning (T/ET Ratio) of Pure European Beech Forest" Water 14, no. 19: 3015. https://doi.org/10.3390/w14193015