
Effects of Temperature on Growth, Molting, Feed Intake, and Energy Metabolism of Individually Cultured Juvenile Mud Crab Scylla paramamosain in the Recirculating Aquaculture System
 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Design
2.2. Experimental Animal and Rearing Conditions
2.3. Sampling
2.4. Ecdysone Content and Molting-Related Genes
2.5. OCR and AER
2.6. Antioxidant Capacity
2.7. Hemolymph Indexes
2.8. Data Collection and Calculation
2.9. Statistics
3. Results
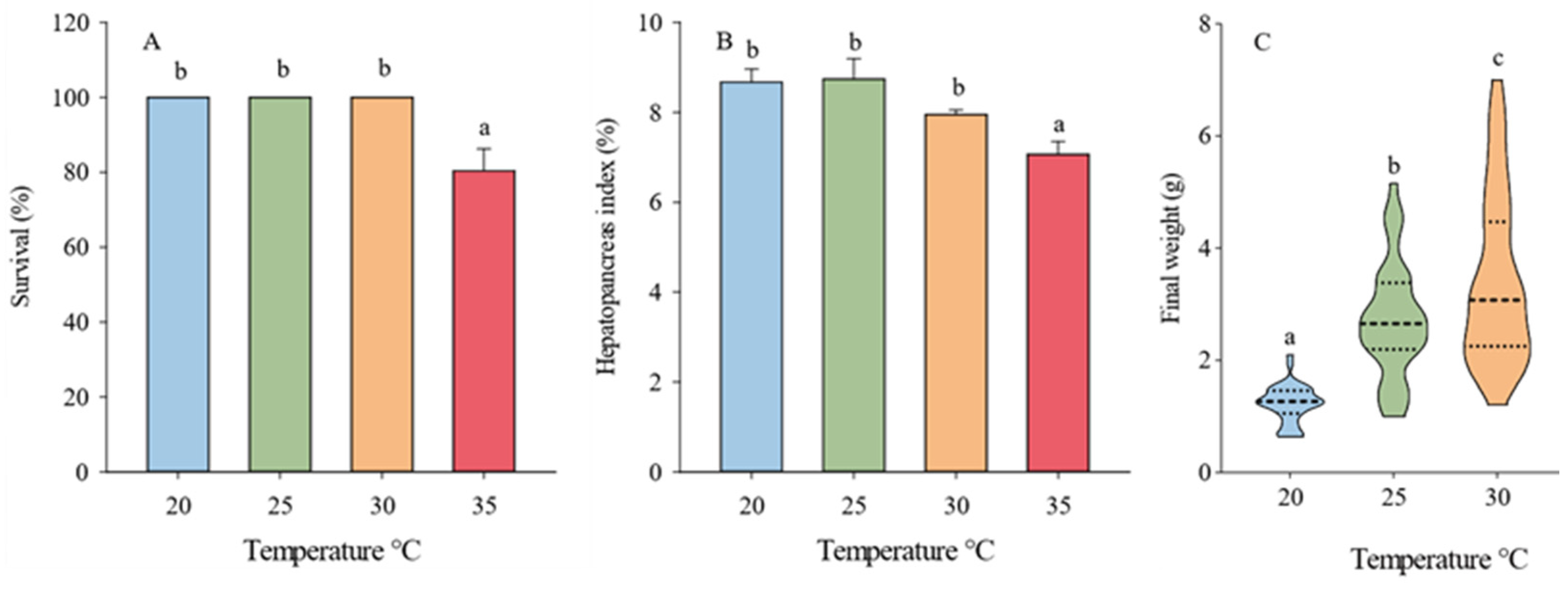
3.1. Survival, Growth Performance, Molting, and Feeding
3.2. Ecdysone Content and Expression of Molting-Related Genes
3.3. OCR, AER, and O: N
3.4. Antioxidant Capacity
3.5. Hemolymph Index
3.6. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keenan, C.P. Aquaculture of the mud crab, genus Scylla-past, present and future. In Aciar Proceedings; Australian Centre for International Agricultural: Washington, DC, USA, 1999; pp. 9–13. [Google Scholar]
- Le Vay, L. Ecology and Management of Mud Crab Scylla spp. Asian Fish. Sci. 2001, 14, 101–112. [Google Scholar] [CrossRef]
- Cowan, L. Crab farming in Japan, Taiwan and the Philippines. Qld. Dep. Prim. Ind. Inf. Ser. Q 1984, 85, 184009. [Google Scholar]
- Liu, X. Fisheries Bureau of Agriculture Ministry of China; China Agriculture Press: Beijing, China, 2021. [Google Scholar]
- Yu, K.; Shi, C.; Liu, X.; Ye, Y.; Wang, C.; Mu, C.; Song, W.; Ren, Z. Tank bottom area influences the growth, molting, stress response, and antioxidant capacity of juvenile mud crab Scylla paramamosain. Aquaculture 2022, 548, 737705. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, G.; Gao, G.; Wang, C.; Mu, C.; Ye, Y.; Shi, C. Effects of culture container color on growth, stress and carapace color of juvenile mud crabs Scylla paramamosain. J. Ningbo Univ. Sci. Technol. Ed. 2021, 34, 43–49. [Google Scholar]
- Beitinger, T.L.; Fitzpatrick, L.C. Physiological and Ecological Correlates of Preferred Temperature in Fish. Am. Zool. 1979, 19, 319–329. [Google Scholar] [CrossRef]
- Tropea, C.; Stumpf, L.; Greco, L.S.L. Effect of Temperature on Biochemical Composition, Growth and Reproduction of the Ornamental Red Cherry Shrimp Neocaridina heteropoda heteropoda (Decapoda, Caridea). PLoS ONE 2015, 10, e0119468. [Google Scholar] [CrossRef]
- Bastos, A.M.; Lima, J.F.; Tavares-Dias, M. Effect of increase in temperature on the survival and growth of Macrobrachium amazonicum (Palaemonidae) in the Amazon. Aquat. Living Resour. 2018, 31, 21. [Google Scholar] [CrossRef]
- Stoner, A.W.; Ottmar, M.L.; Copeman, L.A. Temperature effects on the molting, growth, and lipid composition of newly-settled red king crab. J. Exp. Mar. Biol. Ecol. 2010, 393, 138–147. [Google Scholar] [CrossRef]
- Hyde, C.J.; Elizur, A.; Ventura, T. The crustacean ecdysone cassette: A gatekeeper for molt and metamorphosis. J. Steroid Biochem. Mol. Biol. 2019, 185, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huang, D.; Jia, X.; Zou, Z.; Wang, Y.; Zhang, Z. Functional analysis of the promoter of the molt-inhibiting hormone (mih) gene in mud crab Scylla paramamosain. Gen. Comp. Endocrinol. 2018, 259, 131–140. [Google Scholar] [CrossRef]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Hernández-Sandoval, P.; Díaz-Herrera, F.; Díaz-Gaxiola, J.M.; Martínez-Valenzuela, C.; García-Guerrero, M. Effect of temperature on growth, survival, thermal behavior, and critical thermal maximum in the juveniles of Macrobrachium occidentale (Holthuis, 1950) (Decapoda: Caridea: Palaemonidae) from Mexico. J. Crustacean Biol. 2018, 38, 483–488. [Google Scholar] [CrossRef] [Green Version]
- Re, A.D.; Diaz, F.; Sierra, E.; Rodríguez, J.; Perez, E. Effect of salinity and temperature on thermal tolerance of brown shrimp Farfantepenaeus aztecus (Ives) (Crustacea, Penaeidae). J. Therm. Biol. 2005, 30, 618–622. [Google Scholar] [CrossRef]
- González, R.A.; Díaz, F.; Licea, A.; Re, A.D.; Sánchez, L.N.; García-Esquivel, Z. Thermal preference, tolerance and oxygen consumption of adult white shrimp Litopenaeus vannamei (Boone) exposed to different acclimation temperatures. J. Therm. Biol. 2010, 35, 218–224. [Google Scholar] [CrossRef]
- Thomas, C.W.; Crear, B.J.; Hart, P.R. The effect of temperature on survival, growth, feeding and metabolic activity of the southern rock lobster, Jasus edwardsii. Aquaculture 2000, 185, 73–84. [Google Scholar] [CrossRef]
- Frisk, M.; Steffensen, J.F.; Skov, P.V. The effects of temperature on specific dynamic action and ammonia excretion in pikeperch (Sander lucioperca). Aquaculture 2013, 404, 65–70. [Google Scholar] [CrossRef]
- Hu, Y.; Tan, B.; Mai, K.; Ai, Q.; Zheng, S.; Cheng, K. Growth and body composition of juvenile white shrimp, Litopenaeus vannamei, fed different ratios of dietary protein to energy. Aquac. Nutr. 2008, 14, 499–506. [Google Scholar] [CrossRef]
- Pascual, C.; Gaxiola, G.; Rosas, C. Blood metabolites and hemocyanin of the white shrimp, Litopenaeus vannamei: The effect of culture conditions and a comparison with other crustacean species. Mar. Biol. 2003, 142, 735–745. [Google Scholar] [CrossRef]
- Ciaramella, M.A.; Battison, A.L.; Horney, B. Measurement of tissue lipid reserves in the American lobster (Homarus americanus): Hemolymph metabolites as potential biomarkers of nutritional status. J. Crustac. Biol. 2014, 34, 629–638. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Madeira, D.; Narciso, L.; Cabral, H.N.; Vinagre, C.; Diniz, M.S. Influence of temperature in thermal and oxidative stress responses in estuarine fish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 166, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Paital, B.; Chainy, G.B.N. Effects of temperature on complexes I and II mediated respiration, ROS generation and oxidative stress status in isolated gill mitochondria of the mud crab Scylla serrata. J. Therm. Biol. 2014, 41, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Wang, G.; Li, S. Effects of low temperature acclimation on antioxidant defenses and ATPase activities in the muscle of mud crab (Scylla paramamosain). Aquaculture 2012, 370–371, 144–149. [Google Scholar] [CrossRef]
- Feng, G.; Shi, X.; Huang, X.; Zhuang, P. Oxidative Stress and Antioxidant Defenses after Long-term Fasting in Blood of Chinese Sturgeon (Acipenser sinensis). Procedia Environ. Sci. 2011, 8, 469–475. [Google Scholar] [CrossRef]
- Viarengo, A.; Canesi, L.; Pertica, M.; Poli, G.; Moore, M.N.; Orunesu, M. Heavy metal effects on lipid peroxidation in the tissues of mytilus gallopro vincialis lam. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1990, 97, 37–42. [Google Scholar] [CrossRef]
- Chen, S.; Migaud, H.; Shi, C.; Song, C.; Wang, C.; Ye, Y.; Mu, C. Light intensity impacts on growth, molting and oxidative stress of juvenile mud crab Scylla paramamosain. Aquaculture 2021, 545, 737159. [Google Scholar] [CrossRef]
- Gao, Y.; He, Z.; Zhao, B.; Li, Z.; He, J.; Lee, J.Y.; Chu, Z. Effect of stocking density on growth, oxidative stress and HSP 70 of pacific white shrimp Litopenaeus vannamei. Turk. J. Fish. Aquat. Sci. 2017, 17, 877–884. [Google Scholar] [CrossRef]
- Maniam, J.; Morris, M.J. The link between stress and feeding behaviour. Neuropharmacology 2012, 63, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Shi, C.; Migaud, H.; Song, C.; Mu, C.; Ye, Y.; Wang, C.; Ren, Z. Light Spectrum Impacts on Growth, Molting, and Oxidative Stress Response of the Mud Crab Scylla paramamosain. Front. Mar. Sci. 2022, 9, 840353. [Google Scholar] [CrossRef]
- Shi, C.; Wang, J.; Peng, K.; Mu, C.; Ye, Y.; Wang, C. The effect of tank colour on background preference, survival and development of larval swimming crab Portunus trituberculatus. Aquaculture 2019, 504, 454–461. [Google Scholar] [CrossRef]
- Luo, H.; Wu, Q.; Lin, J.; He, L.; Chen, X.; Niu, W.; Wang, Q.; Zheng, L. Interactive effects of temperature and body weight on oxygen consumption rate, ammonia excretion rate and asphyxiation point of Hippocampus erectus. Fish. Sci. Technol. Inf. 2022, 49, 67–72. [Google Scholar]
- Zhu, S.; Long, X.; Turchini, G.M.; Deng, D.; Cheng, Y.; Wu, X. Towards defining optimal dietary protein levels for male and female sub-adult Chinese mitten crab, Eriocheir sinensis reared in earthen ponds: Performances, nutrient composition and metabolism, antioxidant capacity and immunity. Aquaculture 2021, 536, 736442. [Google Scholar] [CrossRef]
- Nur Syafaat, M.; Mohammad, S.; Nor Azra, M.; Ma, H.; Abol-Munafi, A.B. Effect of water temperature on survival, growth and molting cycle during early crablet instar of mud crab, Scylla paramamosain (Estampador, 1950). Thalass. Int. J. Mar. Sci. 2020, 36, 543–551. [Google Scholar] [CrossRef]
- Phuong, N.T.; Ha, N.T.K.; Nguyen, T.E.; Do Thi, T.H. Effects of different temperatures on the growth, survival and digestive enzyme activities of mud crab Scylla paramamosain at juvenile stage. Aquac. Aquar. Conserv. Legis. 2021, 14, 2741–2750. [Google Scholar]
- Warren, C.E.; Davis, G.E. Laboratory Studies on the Feeding, Bioenergetics, and Growth of Fish. 1967. Available online: https://ir.library.oregonstate.edu/concern/administrative_report_or_publications/x920g185p?locale=en (accessed on 19 August 2022).
- Volkoff, H.; Rønnestad, I. Effects of temperature on feeding and digestive processes in fish. Temperature 2020, 7, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Imsland, A.K.; Foss, A.; Gunnarsson, S.; Berntssen, M.H.; FitzGerald, R.; Bonga, S.W.; Stefansson, S.O. The interaction of temperature and salinity on growth and food conversion in juvenile turbot (Scophthalmus maximus). Aquaculture 2001, 198, 353–367. [Google Scholar] [CrossRef]
- Hewitt, D.R.; Duncan, P.F. Effect of high water temperature on the survival, moulting and food consumption of Penaeus (Marsupenaeus) japonicus (Bate, 1888). Aquac. Res. 2001, 32, 305–313. [Google Scholar] [CrossRef]
- Handeland, S.O.; Imsland, A.K.; Stefansson, S.O. The effect of temperature and fish size on growth, feed intake, food conversion efficiency and stomach evacuation rate of Atlantic salmon post-smolts. Aquaculture 2008, 283, 36–42. [Google Scholar] [CrossRef]
- Zhou, L.; Li, S.; Wang, Z.; Li, F.; Xiang, J. An eclosion hormone-like gene participates in the molting process of Palaemonid shrimp Exopalaemon carinicauda. Dev. Genes Evol. 2017, 227, 189–199. [Google Scholar] [CrossRef]
- Tian, H.; Yang, C.; Yu, Y.; Yang, W.; Lu, N.; Wang, H.; Liu, F.; Wang, A.; Xu, X. Dietary cholesterol level affects growth, molting performance and ecdysteroid signal transduction in Procambarus clarkii. Aquaculture 2020, 523, 735198. [Google Scholar] [CrossRef]
- Liu, T.; Xu, H.; Han, T.; Wang, J.; Yin, F.; Wang, C. Effect of dietary egg yolk lecithin levels on survival, growth, lipid metabolism, and antioxidant capacity of early juvenile green mud crab Scylla paramamosain. Aquaculture 2021, 540, 736706. [Google Scholar] [CrossRef]
- Yuan, Q.; Qian, J.; Ren, Y.; Zhang, T.; Li, Z.; Liu, J. Effects of stocking density and water temperature on survival and growth of the juvenile Chinese mitten crab, Eriocheir sinensis, reared under laboratory conditions. Aquaculture 2018, 495, 631–636. [Google Scholar] [CrossRef]
- Cai, Y.; Summerfelt, R.C. Effects of temperature and size on oxygen consumption and ammonia excretion by walleye. Aquaculture 1992, 104, 127–138. [Google Scholar] [CrossRef]
- Clarke, A.; Fraser, K.P.P. Why does metabolism scale with temperature? Funct. Ecol. 2004, 18, 243–251. [Google Scholar] [CrossRef]
- Mayzaud, P.; Conover, R.J. O:N atomic ratio as a tool to describe zooplankton metabolism. Mar. Ecol. Prog. Ser. 1988, 45, 289–302. [Google Scholar] [CrossRef]
- Brito, R.; Chimal, M.E.; Gaxiola, G.; Rosas, C. Growth, metabolic rate, and digestive enzyme activity in the white shrimp Litopenaeus setiferus early postlarvae fed different diets. J. Exp. Mar. Biol. Ecol. 2000, 255, 21–36. [Google Scholar] [CrossRef]
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Mommsen, T.P.; Vijayan, M.M.; Moon, T.W. Cortisol in teleosts: Dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol. Fish. 1999, 9, 211–268. [Google Scholar] [CrossRef]
- Jia, Y.; Fan, Y.S.; Wang, Y.Z.; Chai, Y.H.; Gao, J.W.; Dou, Y.; Peng, S. Physiological and energy metabolism responses of chinese Loach Paramisgurnus dabryanus (Dabry de Thiersant, 1872) to waterless preservation during transport. Turk. J. Fish. Aquat. Sci. 2018, 19, 279–287. [Google Scholar]
- Schreck, C.B. Stress and fish reproduction: The roles of allostasis and hormesis. Gen. Comp. Endocrinol. 2010, 165, 549–556. [Google Scholar] [CrossRef]
- Pörtner, H.O. Climate variations and the physiological basis of temperature dependent biogeography: Systemic to molecular hierarchy of thermal tolerance in animals. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 132, 739–761. [Google Scholar] [CrossRef]
- Pörtner, H.-O. Oxygen- and capacity-limitation of thermal tolerance: A matrix for integrating climate-related stressor effects in marine ecosystems. J. Exp. Biol. 2010, 213, 881–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′ to 3′) | Size (bp) | Access Number |
|---|---|---|---|
| ecr | F: TAAGTGATGACGACTCGGATGC | 150 | KC354381 |
| R: ACGAGCAAGCCTTTAGCAGTG | |||
| mih | F: TATCAAGTGCAGGAACTCAG | 110 | EU869539 |
| R: GGAACATACAAGCCTAAACA | |||
| β-actin | F: CGAAACCTTCAACACTCCCG | 154 | FJ641977 |
| R: GATAGCGTGAGGAAGGGCATA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Shi, C.; Ye, Y.; Ma, Z.; Mu, C.; Ren, Z.; Wu, Q.; Wang, C. Effects of Temperature on Growth, Molting, Feed Intake, and Energy Metabolism of Individually Cultured Juvenile Mud Crab Scylla paramamosain in the Recirculating Aquaculture System. Water 2022, 14, 2988. https://doi.org/10.3390/w14192988
Liu J, Shi C, Ye Y, Ma Z, Mu C, Ren Z, Wu Q, Wang C. Effects of Temperature on Growth, Molting, Feed Intake, and Energy Metabolism of Individually Cultured Juvenile Mud Crab Scylla paramamosain in the Recirculating Aquaculture System. Water. 2022; 14(19):2988. https://doi.org/10.3390/w14192988
Chicago/Turabian StyleLiu, Jiahao, Ce Shi, Yangfang Ye, Zhen Ma, Changkao Mu, Zhiming Ren, Qingyang Wu, and Chunlin Wang. 2022. "Effects of Temperature on Growth, Molting, Feed Intake, and Energy Metabolism of Individually Cultured Juvenile Mud Crab Scylla paramamosain in the Recirculating Aquaculture System" Water 14, no. 19: 2988. https://doi.org/10.3390/w14192988