
The First Record of Non-Indigenous Cladoceran Evadne nordmanni Lovén, 1836 (Cladocera, Podonidae) in the Middle Part of the Caspian Sea
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
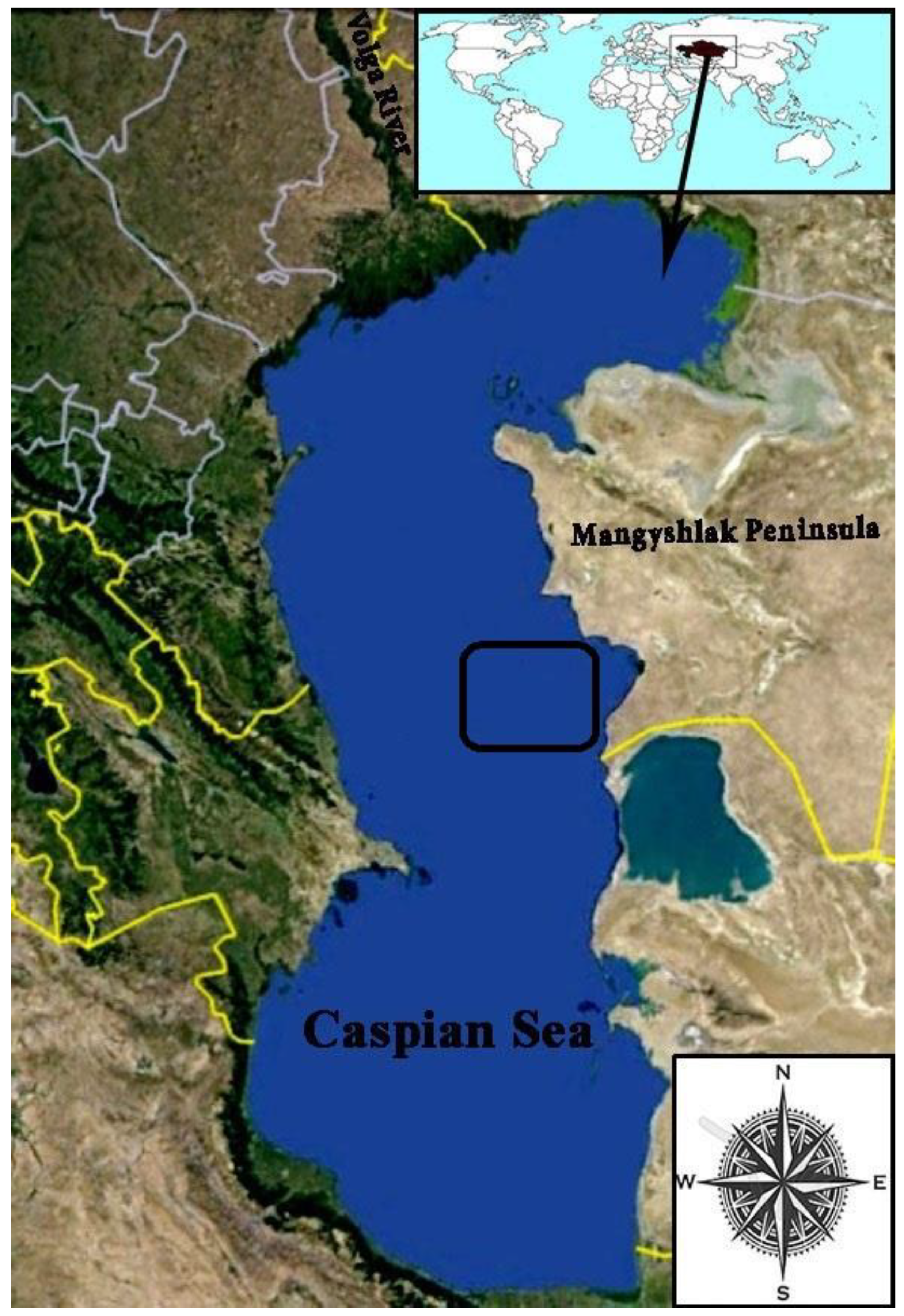
2.1. Description of Study Area
2.2. Field Sampling
2.3. Laboratory Processing
2.4. Statistical Analysis and Comparisons with Previous Studies
3. Results
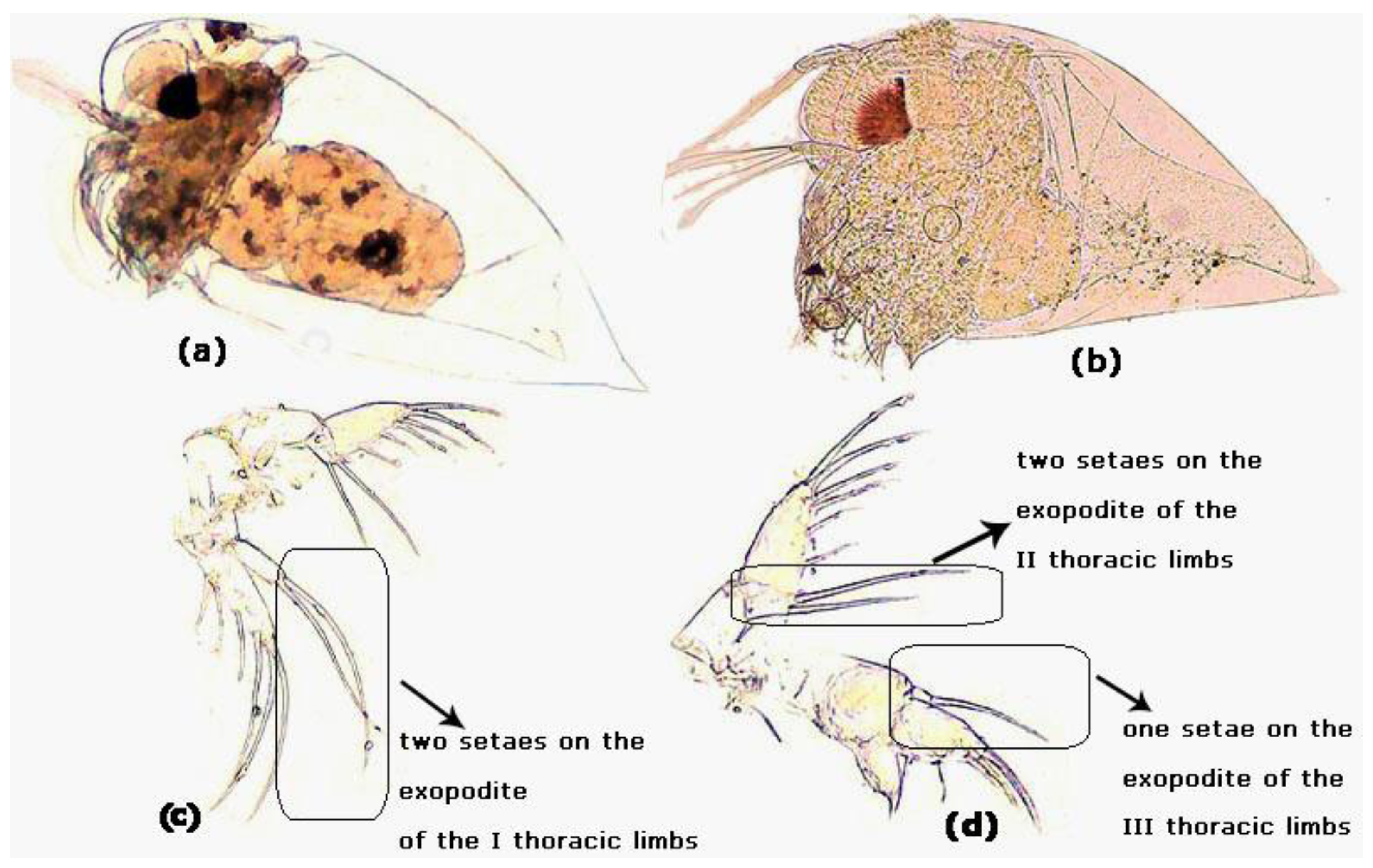
3.1. Morphological Characteristics of Evadne nordmanni
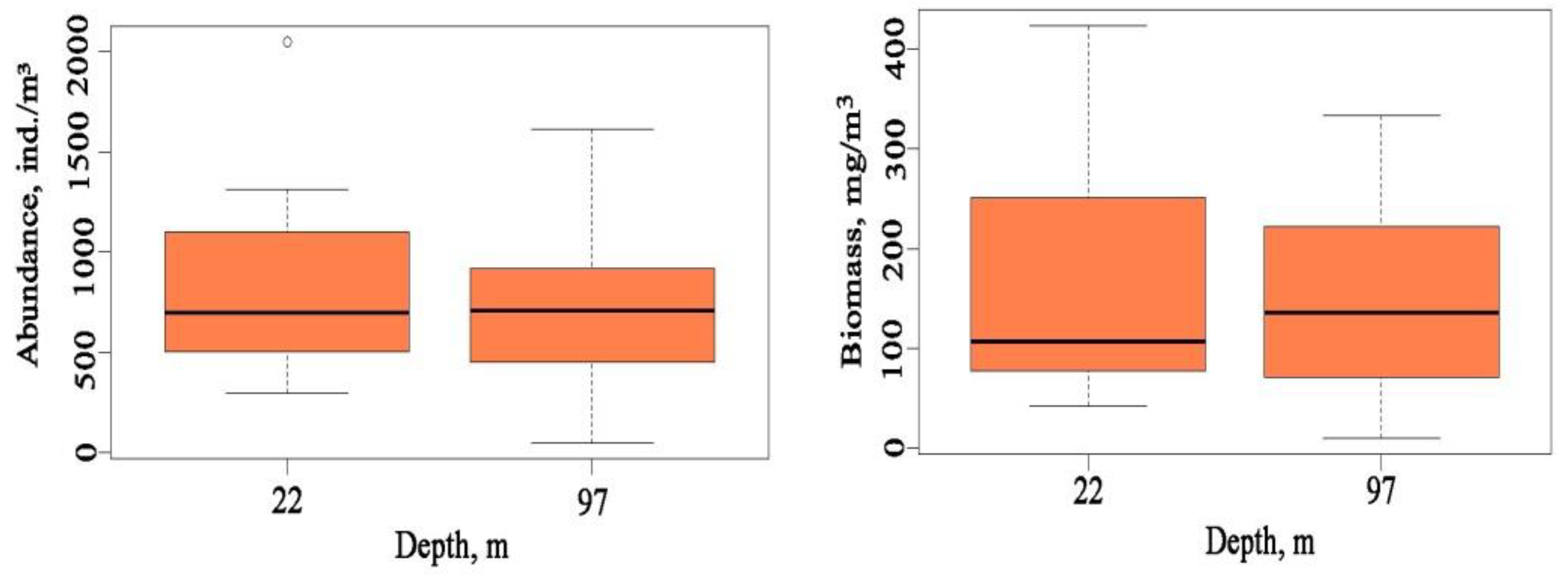
3.2. Abundance and Biomass of Evadne nordmanni
3.3. Coexistence of Evadne nordmanni with Other Planktonic Invertebrates in the Middle Caspian Sea
4. Discussion
4.1. Pathways of Introduction of Evadne nordmanni to the Caspian Sea
4.2. Coexistence of Evadne nordmanni with Other Planktonic Invertebrates in the Middle Caspian Sea and Potential Consequences on Biodiversity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mordukhai-Boltovskoi, F.D.; Riviere, I.K. Predatory cladocerans Podonidae, Polyphemidae, Cercopagidae and Leptodoridae of the fauna of the world. In Guide-Books on the Fauna of the USSR; Science: Leningrad, Russia, 1987; pp. 119–123. [Google Scholar]
- Mordukhai-Boltovskoi, P.D.; Rivier, I.K. A brief survey of the ecology and biology of the Caspian Polyphemoidea. Mar. Biol. 1971, 8, 160–169. [Google Scholar] [CrossRef]
- Rodionova, N.V.; Panov, V.E. Establishment of the Ponto-Caspian predatory cladoceran Evadne anonyx in the eastern Gulf of Finland, Baltic Sea. Aquat. Invasions 2006, 1, 7–12. [Google Scholar] [CrossRef]
- Põllupüü, M.; Simm, M.; Põllumäe, A. Distribution and population structure of the non-indigenous cladoceran Evadne anonoyx in comparison with the native Evadne nordmanni in the north-eastern Baltic Sea. In Proceedings of the 4th International Zooplankton Production Symposium, Hiroshima, Japan, 28 May–1 June 2007. [Google Scholar]
- Kalaus, M.; Ojaveer, H. Over one decade of invasion: The non-indigenous cladoceran Evadne anonyx G.O. Sars, 1897 in a low-salinity environment. Aquat. Invasions 2014, 9, 499–506. [Google Scholar] [CrossRef]
- Technical Project to Carry out Engineering and Geological Surveys at the Point of Laying Exploration Well V-1. Available online: https://www.gov.kz/api/v1/public/assets/2020/5/19/bf2ddd7e27bba574d938d5f92818963a_original.6770034.pdf (accessed on 10 June 2022).
- Birds.kz. Available online: https://www.birds.kz (accessed on 10 June 2022).
- Prusova, I.Y.; Gubanova, A.D.; Shadrin, N.V.; Kurashova, E.K.; Tinenkova, D.H. Acartia tonsa (Copepoda, Calanoida): A New Species in the Caspian and Azov Seas Zooplankton. Vestn. Zool. 2002, 36, 65–68. [Google Scholar]
- Krupa, E.; Aubakirova, M. Checklist and Distribution of Calanoida (Crustacea: Copepoda) in Kazakhstan (Central Asia). Water 2021, 13, 2015. [Google Scholar] [CrossRef]
- Kamakin, A.M.; Khodorevskaya, R.P.; Paritsky, Y.A. Influence of the new invastor CESTONE Mnemiopsis leidyi (A. Agassis, 1865) on the main parts of the Caspian sea ecosystem. Vestn. Astrakhan State Tech. Univ. 2018, 1, 35–48. [Google Scholar]
- Vostokov, S.V.; Gadgiev, A.A.; Vostokova, A.S.; Rabazanov, N.I.; The ctenophore Beroe, cf. ovata in the Caspian Sea. The beginning of a new stage in the evolution of the Caspian ecosystem? South Russ. Ecol. Dev. 2020, 15, 21–35. (In Russian) [Google Scholar] [CrossRef]
- Pires-Teixeira, L.M.; Neres-Lima, V.; Creed, J.C. How Do Biological and Functional Diversity Change in Invaded Tropical Marine Rocky Reef Communities? Diversity 2021, 13, 353. [Google Scholar] [CrossRef]
- Støttrup, J.G.; McEvoy, L.A. Production and Nutritional Value of Copepods, 2nd ed.; Støttrup, J.G., LMcEvoy, L.A., Eds.; John Wiley & Sons: New York, NY, USA, 2003; Volume 3, pp. 145–205. [Google Scholar]
- Benni, W.H. Advances using Copepods in Aquaculture. J. Plankton Res. 2017, 39, 972–974. [Google Scholar]
- Samchyshyna, L. Ecological Characteristic of Calanoids (copepoda, Calanoida) of the Inland Waters of Ukraine. Vestn. Zool. 2008, 42, 32–37. [Google Scholar] [CrossRef]
- Fao.org. Available online: https://www.fao.org/publications/sofia/2022/en/ (accessed on 11 July 2022).
- Kirichenko, O.I.; Zharkenov, D.K. Bleak is an alien species of fish in the reservoirs of the Irtysh basin and the problem of biological invasions. Selevinia 2009, 5, 155–158. [Google Scholar]
- Polianinova, A.A.; Tatarintseva, T.A.; Terletskaia, O.V.; Tinenkova, D.K.; Petrenko, E.L.; Kochneva, L.A. Hydrobiological Environment in the Middle of the Southern Part of the Caspian Sea in Conditions of Invasion of Ctenophoran Mnemiopsis leidyi. Results of Research Work; Izd-vo KaspNIRKh: Astrakhan, Russia, 2003; pp. 121–134. [Google Scholar]
- Wikipedia.org. Available online: https://en.wikipedia.org/wiki/Caspian_Sea (accessed on 19 July 2022).
- Abdybekova, A.; Assylbekova, S.; Abdibayeva, A.; Zhaksylykova, A.; Barbol, B.; Aubakirov, M.; Torgerson, P. Studies on the population biology of helminth parasites of fish species from the Caspian Sea drainage basin. J. Helminthol. 2021, 95, e12. [Google Scholar] [CrossRef]
- Winberg, G.G.; Lavrenteva, G.M. (Eds.) Zooplankton and Its Products. In Guidelines for the Collection and Processing of Materials in Hydrobiological Research in Freshwater Water Bodies; GosNIIORH: Leningrad, Russia, 1984; p. 34. (In Russian) [Google Scholar]
- Kutikova, L.A. Rotifers of the Fauna of the USSR; Science: Leningrad, Russia, 1964; p. 744. [Google Scholar]
- Krupa, E.G.; Dobrokhotova, O.V.; Stuge, T.S. Fauna of Calanoida (Crustacea: Copepoda) of Kazakhstan and Adjacent Territories; Etalon Print: Almaty, Kazakhstan, 2016; p. 208. [Google Scholar]
- Mordukhai-Boltovskoi, P.D. Polyphemidae of the Pontocaspian Basin. Hydrobiologia 1965, 25, 212–220. [Google Scholar] [CrossRef]
- Korovchinsky, N.M.; Kotov, A.A.; Sinyov, A.Y.; Neretina, A.N.; Garibyan, P.G. Cladocerans (Crustacea: Cladocera) of Northern Eurasia; KMK: Moscow, Russia, 2021; pp. 497–502. [Google Scholar]
- Balushkina, E.V.; Vinberg, G.G. The relationship between the length and body weight of planktonic crustaceans. In Experimental and Field Studies of the Biological Foundations of Lake Productivity; Vinberg, G.G., Ed.; Institute of Lake and River Fishery: Leningrad, Russia, 1979; pp. 58–79. [Google Scholar]
- Bakanov, A.I. Quantitative Assessment of Domination in Ecological Communities; Science: Tolyatti, Russia, 2005; pp. 37–67. [Google Scholar]
- Kabacoff, R. R in Action; University of Connecticut, Manning Publications Co.: New York, NY, USA, 2011; p. 771. [Google Scholar]
- Sharapova, L.I. The current state of zooplankton in deep biotopes eastern part of the Middle Caspian. In Proceedings of the Study of Aquatic and Terrestrial Ecosystems: History and Modernity, Sevastopol, Russia, 13–18 September 2021. [Google Scholar]
- Krupa, E. Assessment of Changes in the Structure of Zooplankton Communities to Infer Water Quality of the Caspian Sea. Diversity 2019, 11, 122. [Google Scholar] [CrossRef] [Green Version]
- Ibrasheva, S.I.; Smirnova, V.A. Cladocera of Kazakhstan; Mektep: Alma Ata, Kazakhstan, 1983; p. 135. (In Russian) [Google Scholar]
- Green, A.J.; Elmberg, J. Ecosystem services provided by waterbirds. Biol. Rev. 2013, 89, 105–122. [Google Scholar] [CrossRef]
- Streftaris, N.; Zenetos, A.; Papathanassiou, F. Globalisation in marine ecosystems: The story of non-indigenous marine species across European seas. Oceanogr. Mar. Biol. Annu. Rev. 2005, 43, 419–453. [Google Scholar]
- Battes, K.P.; Váncsa, É.; BarbuTudoran, L.; Cîmpean, M. A species on the rise in Europe: Sinodiaptomus sarsi (Rylov, 1923) (Copepoda, Calanoida), a new record for the Romanian crustacean fauna. BioInvasions Rec. 2020, 9, 320–332. [Google Scholar] [CrossRef]
- Reynolds, C.; Miranda, N.A.F.; Cumming, G.S. The role of waterbirds in the dispersal of aquatic alien and invasive species. Divers. Distrib. 2015, 21, 744–754. [Google Scholar] [CrossRef]
- Temreshev, I.I.; Esenbekova, P.A.; Kozhabaeva, G.E.; Isenova, G.Z.; Slivinsky, G.G. About the distribution the freshwater shrimps (Crustacea: Decapoda: Palaemonidae) in water bodies of South Kazakhstan and opportunities of their use as biogeoindicators of the condition of aquatic ecosystems. Bull. Natl. Acad. Sci. Repub. Kazakhstan 2017, 2, 215–223. [Google Scholar]
- Kirichenko, O.E.; Anuarbekov, S. The state of biodiversity of water bodies of the Irtysh basin and the impact of alien species on the ecosystem. Eurasian Union Sci. 2016, 4, 112–116. [Google Scholar]
- Assylbekova, S.Z.; Kulikov, E.V. Introduction of fish and aquatic invertebrates in water bodies of Kazakhstan: Results and perspectives. Vestn. Astrakhan State Tech. Univ. 2013, 3, 16–29. [Google Scholar]
- Bainbridge, V. Some Observations on Evadne nordmanni Lovén. J. Mar. Biol. Assoc. UK 1958, 37, 349–370. [Google Scholar] [CrossRef]
- Povazhny, V.V. Peculiarities of functioning of the zooplankton community of the Taganrog Bay of the Sea of Azov; Murmansk Marine Biological Institute: Murmansk, Russia, 2009; p. 129. [Google Scholar]
- Katechakis, A.; Stibor, H. Feeding selectivities of the marine cladocerans Penilia avirostris, Podon intermedius and Evadne nordmanni. Mar. Biol. 2004, 145, 529–539. [Google Scholar] [CrossRef]
- Sampey, A.; McKinnon, A.D.; Meekan, M.G.; McCormick, M.I. Glimpse into guts: Overview of the feeding of larvae of tropical shorefishes. Mar. Ecol. Prog. Ser. 2007, 339, 243–257. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Station | Depth, m | Coordinates | |
|---|---|---|---|
| Longitude | Latitude | ||
| 1 | 97 | 51°48.3535′ E | 42°07.2959′ N |
| 2 | 22 | ||
| 3 | 97 | 51°48.3535′ E | 42°07.5659′ N |
| 4 | 22 | ||
| 5 | 97 | 51°48.3535′ E | 42°07.836′ N |
| 6 | 22 | ||
| 7 | 97 | 51°47.6347′ E | 42°08.106′ N |
| 8 | 22 | ||
| 9 | 97 | 51°47.9941′ E | 42°08.106′ N |
| 10 | 22 | ||
| 11 | 97 | 51°44.7594′ E | 42°10.8065′ N |
| 12 | 22 | ||
| 13 | 97 | 51°51.9476′ E | 42°10.8065′ N |
| 14 | 22 | ||
| 15 | 97 | 51°51.9476′ E | 42°05.4055′ N |
| 16 | 22 | ||
| Taxa | Abundance | Contribution % | Biomass | Contribution % |
|---|---|---|---|---|
| Rotifera | ||||
| Synchaeta littoralis Rousselet, 1902 | 2.8 ± 2.7 | 0.06 | 0.01 ± 0.01 | 0.006 |
| Cladocera | ||||
| Evadne anonyx G.O. Sars, 1897 | 2.7 ± 0.9 | 0.06 | 0.2 ± 0.1 | 0.117 |
| Pleopis polyphemoides Leuckart, 1859 | 26.7 ± 14.9 | 0.61 | 2.6 ± 1.7 | 1.390 |
| Podon intermedius Lilljeborg, 1853 | 13.7 ± 11.3 | 0.31 | 1.5 ± 1.3 | 0.817 |
| Evadne nordmanni Lovén, 1836 | 799.4 ± 127.6 | 18.30 | 159.1 ± 28.8 | 84.891 |
| Copepoda | ||||
| Acartia tonsa Dana, 1849 | 1746.8 ± 351.7 | 40.0 | 13.6 ± 3.0 | 7.275 |
| Calanipeda aquaedulcis Krichagin, 1873 | 0.2 ± 0.1 | 0.004 | 0.0003 ± 0.0003 | 0.0002 |
| Halicyclops sarsi Akatova, 1935 | 0.8 ± 0.5 | 0.02 | 0.002 ± 0.001 | 0.001 |
| Others | ||||
| Bivalvia gen.sp. | 21.6 ± 4.3 | 0.49 | 0.09 ± 0.02 | 0.046 |
| Spionidae sp. | 0.7 ± 0.4 | 0.02 | 0.003 ± 0.003 | 0.002 |
| Cirripedia gen.sp. | 1457.3 ± 356.8 | 33.4 | 2.9 ± 0.7 | 1.573 |
| Ostracoda gen.sp. | 291.4 ± 58.7 | 6.7 | 7.3 ± 1.5 | 3.880 |
| Total | 4364.1 ± 751.1 | 100 | 187.4 ± 30.6 | 100 |
| Taxon Name | Years | ||
|---|---|---|---|
| 2008 [30] | 2020 [29] | 2021 | |
| Rotifera | |||
| Brachionus quadridentatus Hermann, 1783 | + | − | − |
| Synchaeta cecilia Rousselet, 1902 | + | − | − |
| Synchaeta littoralis Rousselet, 1902 | + | − | + |
| Synchaeta stylata Wierzejski, 1893 | + | − | − |
| Cladocera | |||
| Podonevadne camptonyx (G.O. Sars, 1897) | + | + | + |
| Podonevadne angusta (G.O. Sars, 1902) | + | + | − |
| Podonevadne trigona (G.O. Sars, 1897) | + | + | − |
| Pleopis polyphemoides (Leuckart, 1859) | + | + | + |
| Podon intermedius Lilljeborg, 1853 | + | + | + |
| Evadne anonyx Sars,1897 | + | + | + |
| Cornigerius maeoticus hircus (GO Sars, 1902) | + | − | − |
| Evadne nordmanni Lovén, 1836 | − | − | + |
| Copepoda | |||
| Acartia tonsa Dana, 1849 | + | + | + |
| Calanipeda aquedulcis Krichagin, 1873 | + | − | + |
| Halicyclops sarsi Akatova, 1935 | + | − | + |
| Idyaea furcata (Baird, 1837) | + | − | − |
| Ergasilidae gen.sp. | + | − | − |
| Calanoida gen.sp. | + | − | − |
| Others | |||
| Mnemiopsis leidyi A. Agassiz, 1865 | − | + | − |
| Hediste diversicolor (O.F. Müller, 1776) | + | − | − |
| Spionidae sp. | − | − | + |
| Bivalvia gen.sp. | + | + | + |
| Cirripedia gen.sp. | + | + | + |
| Nematoda gen.sp. | + | − | − |
| Total | 21 | 10 | 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aubakirova, M.; Krupa, E.; Magda, I.; Assylbekova, S.Z.; Abayev, A.; Abilov, B.; Tumenov, A.; Isbekov, K.B.; Mazhibayeva, Z. The First Record of Non-Indigenous Cladoceran Evadne nordmanni Lovén, 1836 (Cladocera, Podonidae) in the Middle Part of the Caspian Sea. Water 2022, 14, 2818. https://doi.org/10.3390/w14182818
Aubakirova M, Krupa E, Magda I, Assylbekova SZ, Abayev A, Abilov B, Tumenov A, Isbekov KB, Mazhibayeva Z. The First Record of Non-Indigenous Cladoceran Evadne nordmanni Lovén, 1836 (Cladocera, Podonidae) in the Middle Part of the Caspian Sea. Water. 2022; 14(18):2818. https://doi.org/10.3390/w14182818
Chicago/Turabian StyleAubakirova, Moldir, Elena Krupa, Igor Magda, Saule Zh. Assylbekova, Almat Abayev, Berdibek Abilov, Artur Tumenov, Kuanysh B. Isbekov, and Zhanara Mazhibayeva. 2022. "The First Record of Non-Indigenous Cladoceran Evadne nordmanni Lovén, 1836 (Cladocera, Podonidae) in the Middle Part of the Caspian Sea" Water 14, no. 18: 2818. https://doi.org/10.3390/w14182818