
Long-Term Exposure to Polystyrene Nanoplastics Impairs the Liver Health of Medaka
by
 , and
, and
Yinfeng Zhou
1,2,†, 
Li Zhao
1,2,†,
Haijing Xu
1,2,
Elvis Genbo Xu
3,
Mingyou Li
1,2,* and
Youji Wang
2,* 1
Key Laboratory of Integrated Rice-Fish Farming, Ministry of Agriculture and Rural Affairs, Shanghai Ocean University, Shanghai 201306, China
2
Key Laboratory of Exploration and Utilization of Aquatic Genetic Resources, Ministry of Education, Shanghai Ocean University, Shanghai 201306, China
3
Department of Biology, University of Southern Denmark, 5230 Odense, Denmark
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Water 2022, 14(17), 2767; https://doi.org/10.3390/w14172767
Submission received: 30 July 2022
/
Revised: 31 August 2022
/
Accepted: 1 September 2022
/
Published: 5 September 2022
(This article belongs to the Special Issue Effects of Micro- and Nano-Plastic Pollution on Marine Ecosystems and Human Health)
Abstract
:Highlights
What are the main findings?
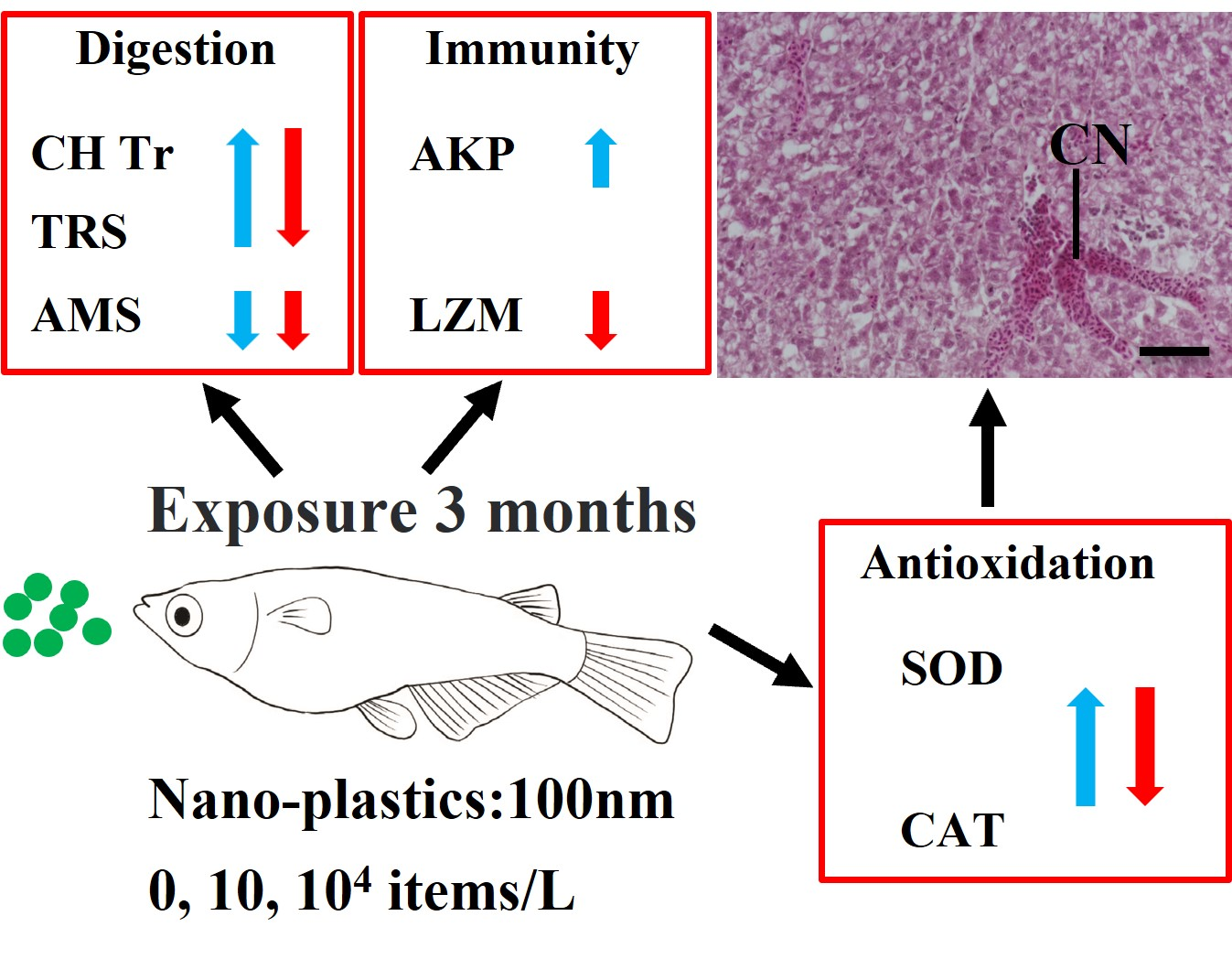
- The hepatotoxic effects of nanoplastics (NPs) are evaluated in medaka fish.
What is the implication of the main finding?
- Digestive, innate immune, and antioxidant enzyme activities are affected by NP exposure.
- Histopathology confirmed direct liver tissue damage by NP exposure.
- We conclude that NPs at as low as 10 items/L can be toxic to medaka in long term.
Abstract
Nanoplastic (NP) has become a global environmental concern due to its potentially wide distribution and unknown impacts on organisms. Many laboratory studies have reported the short-term toxicity of NPs but their long-term effects are unclear. Here, the chronic hepatoxic effects of NPs (diameter 100 nm; 0, 10, and 104 items/L) in medaka (Oryzias latipes) were evaluated after three-month exposure by investigating the alterations in enzymatic biomarkers (digestion, oxidative status, and immunity) and histopathology in the liver. The trypsin and chymotrypsin activities of digestive enzymes were induced at a low concentration of NPs (10 items/L) but inhibited at high concentration (104 items/L); only the amylase activity was significantly decreased in all NP-treated medaka. Oxidative stress was also induced by NP exposure, which was indicated by the responsive superoxide dismutase and catalase. Lysozyme was significantly reduced after exposure to a high concentration of NPs, while alkaline phosphatase was significantly induced at a low concentration. Exposure to NPs also caused liver damage (e.g., congestion and dilated sinusoids). Taken together, our findings show that chronic exposure to NPs at low particle concentrations can impair the liver health of medaka by affecting liver enzymatic functions and causing histological damage. This implies potentially long-term threats of NPs to aquatic organisms, which call for more long-term aquatic toxicological studies on various species using low environmental concentrations.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Plastics are applied around the world because of their low prices, endurance, and good insulation [1]. Hence, global plastic products have grown exponentially since the 1950s, reaching 348 million tons in 2018 [2]. Due to overuse and improper management of plastic waste, more and more plastic wastes end up in the environment, leading to serious environmental pollution [1,3]. Plastic fragments are known to be widely distributed in natural environments, including freshwater, marine, sediments, and terrestrial. Plastics can degrade into microscaled and nanoscaled fragments through weathering, biodegradation, and ultraviolet radiation under the natural environment [4]. Microplastics (MPs) are identified as having particle sizes under 5 mm, whereas particle sizes <1000 nm are defined as NPs [5]. NPs are considered more harmful to aquatic animals than MPs owing to their higher surface area, cell affinity, and adsorption ability, thereby allowing them to penetrate biological barriers and accumulate in different tissues more easily, including gut, liver, and kidney [6,7,8].
NPs have been reported to be associated with oxidative stress, reproduction, immunotoxicity, and the digestive system of organisms under laboratory conditions [9,10,11,12,13]. Specifically, oxidative stress can be manifested through changes in enzymatic activity [14]. Brandts et al. (2018) reported that NPs (45 nm) inhibited the activities of immune enzymes (e.g., alkaline phosphatase and esterase) in Dicentrarchus labrax within a 4-d exposure period [15]. The induction of systemic inflammation and oxidative stress was observed following exposure to 200-nm NPs (0.2 mg/L) in the zebrafish embryos within 96 h [16]. Moreover, a previous study demonstrated that all D. magna died after acute exposure to 52-nm particles at a concentration above 75 mg/L within 13 h [17]. Some other short-term toxic effects include impacts on immunity and digestive system, oxidation status, and even survival of organisms [18,19,20]. Lu et al. (2016) proved that histopathological changes (e.g., inflammation and oxidative stress) were found in the liver of zebrafish after being exposed to 70-nm NPs [7]. The digestive system is critical for the absorption of nutrients and the obtainment of energy with the help of related digestive enzymes, which are often used to manifest biotoxicity and digestibility [2]. Yet, data on the chronic effects of NPs on freshwater organisms remain limited, particularly under environmentally realistic scenarios, such as at low-particle-exposure concentrations [21].
To fill the knowledge gap, we investigated the long-term toxic effects of NPs on medaka fish (Oryzias latipes), focusing on the liver, which is the critical detoxification organ for foreign particulates. As a well-established model fish in ecotoxicology, medaka shows numerous advantages, including prolific egg laying, small size, rapid maturation, sexual dimorphism, and easy culture [22]. Although not well-demonstrated for nanoplastics, a number of earlier studies have shown that the liver is a key target organ of nanomaterials and is well-known for its critical biological roles in metabolism, digestion, immunity, and digestion function [23,24,25]. Therefore, we hypothesize that liver health and liver functions will be impaired after long-term exposure to NPs. To test this hypothesis, medaka were exposed to two relatively low particle concentrations of NPs for three months, simulating a more realistic environment scenario than many earlier acute tests that used high concentrations. Then, the effects of the NPs on the digestive system, antioxidant status, immune function, and histopathological changes in the liver of medaka were systematically quantified. These findings offer novel insights into the biological impacts of long-term exposure to NPs in fish, which is a prerequisite for ecological risk assessment of NPs.
2. Materials and Methods
2.1. Nanoplastics
One-hundred nanometer fluorescent polystyrene (PS) NPs (concentration: 10 mg/cm3; excitation: 488 nm; emission: 518 nm) were obtained from BaseLine Chromtech Research Centre (Tianjin, China). PS is one of the most commonly used plastic polymers worldwide [26] and has been successfully used as a model particle for studying the accumulation and biological effects of plastic particles in various organisms [27].
2.2. Experimental Fish
Medaka, fed with live Artemia sp. three times daily, was raised in a 14 h light/10 h dark cycle at 26 °C [28]. In addition, the dissolved oxygen (6 mg/L) and pH (6.5–7.0) were controlled in culture fresh water. The use of medaka fish was approved by the Committee for Laboratory Animal Research at Shanghai Ocean University.
2.3. Experimental Design
Before the experiment, 60 dph medaka were acclimated in a glass tank for two weeks and then randomly divided into glass tanks. For the toxicity test, the experimental fish were randomly divided into three different concentration groups. Each group comprised 3 replicate glass tanks and each tank contained 64 fish (32 females and 32 males). Low (10 items/L) and high (104 items/L) concentrations used in this study were selected, which is based on previously reported toxicological impacts of plastics on aquatic organisms [2]. Three concentration suspensions were prepared by diluting the NPs stock solution suspension, and ultrasonic treatment was required before preparation to disperse NPs in the suspension.
The NP suspension was renewed weekly during the exposure. All the medaka were exposed for three months and then collected and dissected for toxicological endpoint measurements. Before dissection, each fish was thoroughly rinsed with Milli-Q water to remove water-containing NPs on the skin surface. For liver histopathological analysis, livers of four fish were collected from each tank. For biochemical analysis, liver tissues from eight fish were pooled together as one replicate to ensure an adequate number of samples per group and to control for differences between individuals. The livers were stored at −80 °C before biochemical analysis.
2.4. Liver Histopathology
The livers from the NP-exposed and the control treatments were placed in 4% PFA. Then, the liver tissues were dehydrated with different ethanol, embedded in paraffin wax, sectioned at 4 μm-thickness, and then stained with hematoxylin & eosin (H&E). The liver tissue morphology was observed and then photographed using a Nikon Ds-Ri2 camera under an upright microscope (Nikon, Tokyo, Japan).
2.5. Biochemical Analysis
The liver tissue of medaka was weighed, homogenized in ice-cold PBS (phosphate-buffered saline), and centrifuged at 2500× g for 10 min (4 °C). The collected supernatant was used to measure enzyme activities (digestive, oxidative, and innate immune enzymes) using a commercial kit (Nanjing Jiancheng, Bioengineering Institute, Nanjing, China). Moreover, protein content was evaluated by the coomassie blue staining [29]. Each assay was measured in triplicate. Digestive enzymes included trypsin (TRS), lipase (LPS), chymotrypsin (CH Tr), and amylase (AMS); markers of oxidative stress included superoxide dismutase (SOD) and catalase (CAT); innate immune enzymes were lysozyme (LZM) and alkaline phosphatase (AKP).
2.6. Statistics
All data were statistically analyzed with SPSS 25 software (SPSS Inc., Chicago, IL, USA) and presented as means ± SD. All data were analyzed for distribution normality and variance homogeneity by using Shapiro–Wilk’s test and using Levene’s test, respectively. The effect of NP concentration on the enzyme activities of medaka was tested and considered significant (p < 0.05) by using one-way ANOVA followed by Tukey’s HSD tests. Origin 2018 was used for principal component analysis (PCA) on all enzymatic activities.
3. Results
3.1. Digestive Enzyme Activities
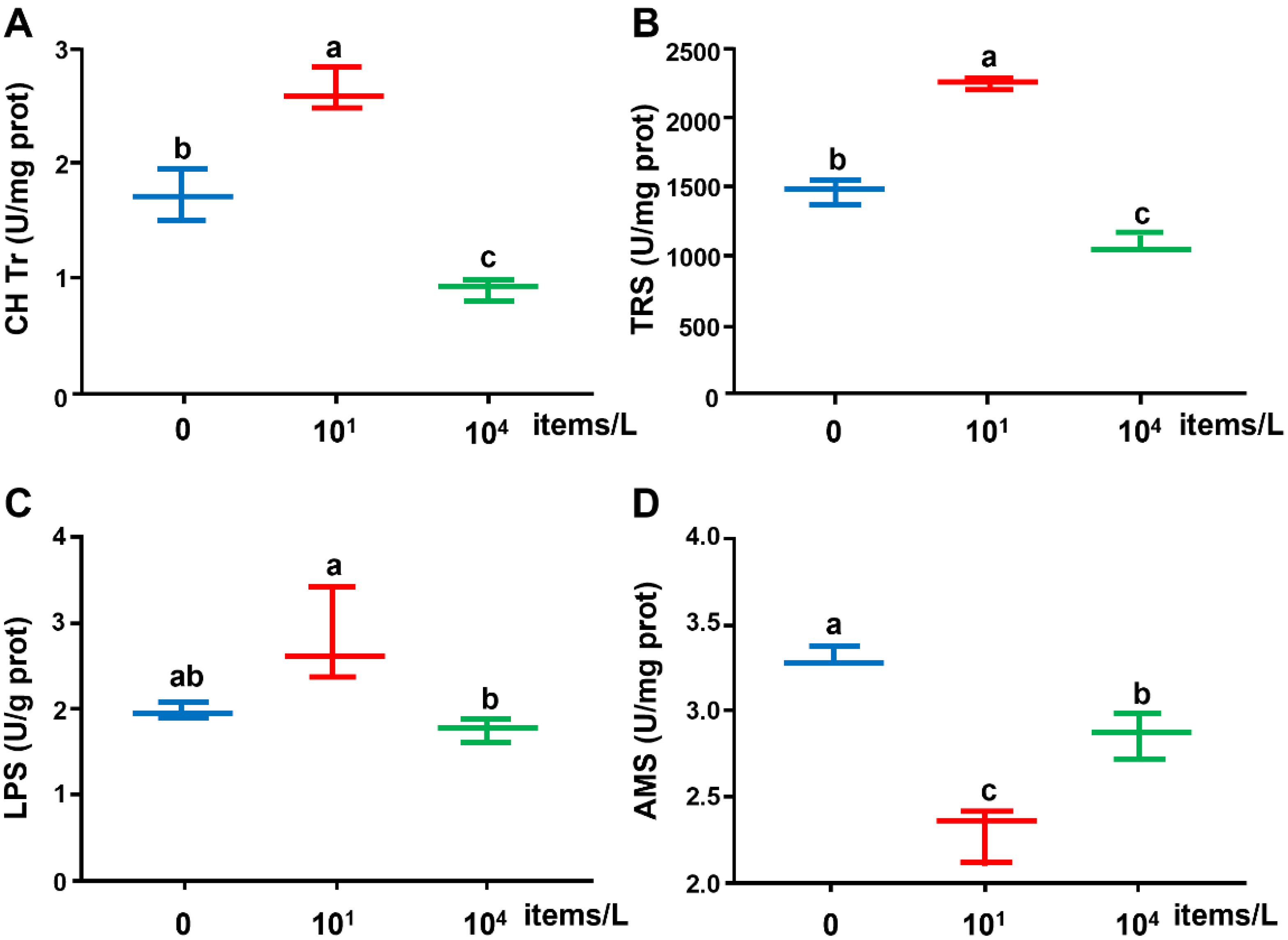
The CH Tr and TRS enzyme activities in the liver were induced by the low concentration (10 items/L) of NPs but inhibited by the high concentration (104 items/L) of NPs (Figure 1). Significant differences in all enzyme activities were detected between the low- and high-concentration groups. Significantly lower AMS enzyme activity was observed in NP-treated groups compared with the control (0 items/L) group.
3.2. Innate Immune Enzyme Activities
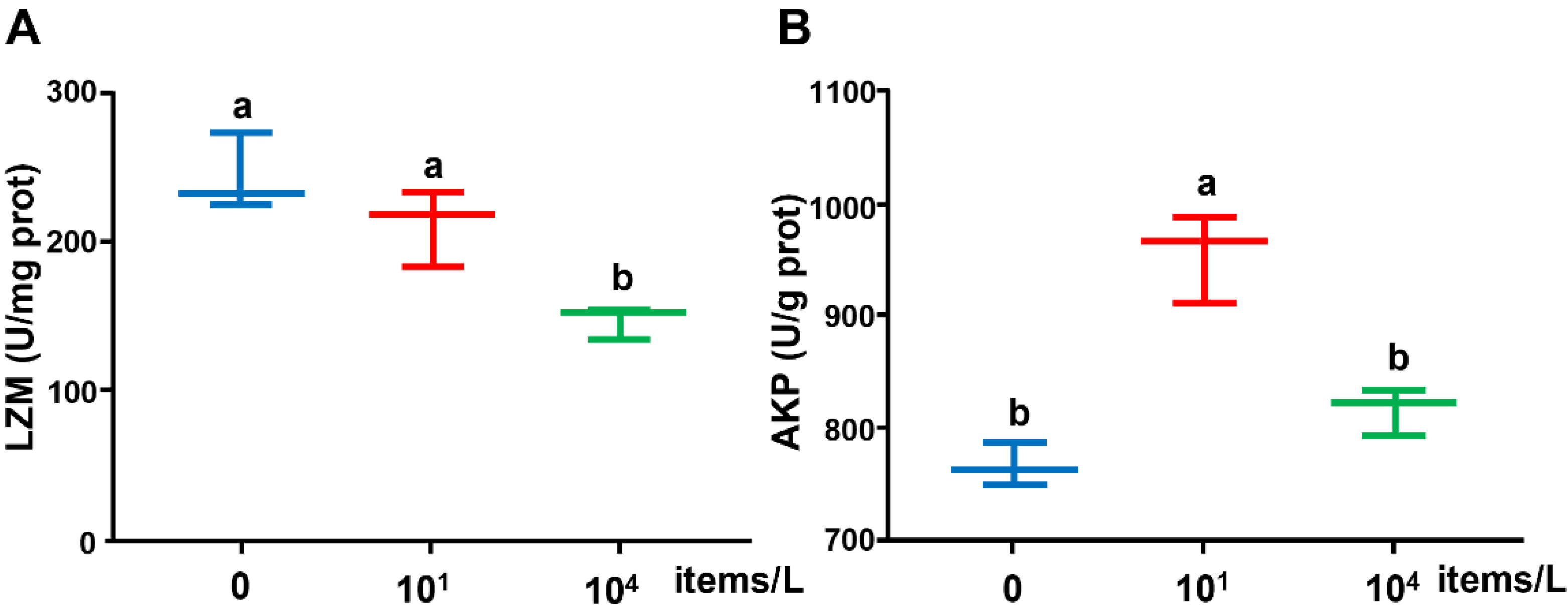
The LZM activity was significantly reduced at the high concentration of NPs (104 items/L) compared with the control (0 items/L) group. The enzyme activity of AKP was significantly promoted by low-concentration (10 items/L) NPs compared with the control (0 items/L), while no significant difference was found between the high-concentration (104 items/L) and control groups (Figure 2).
3.3. Antioxidant Enzyme Activities
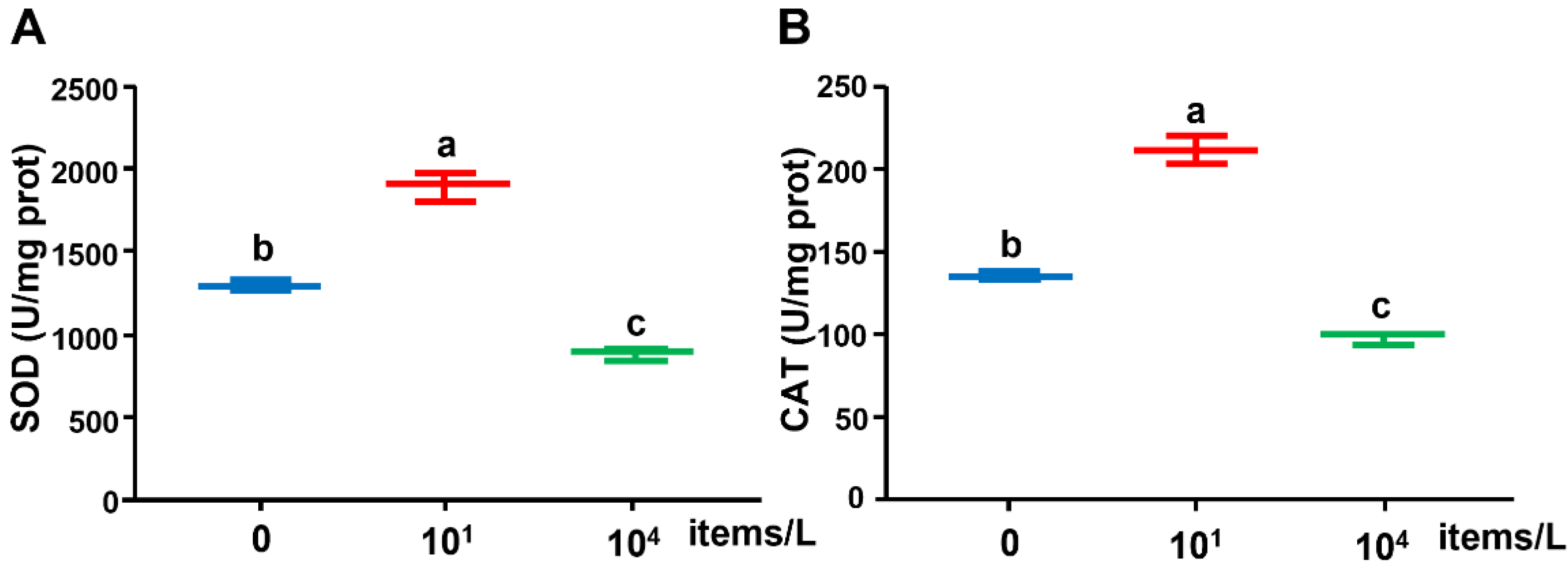
As shown in Figure 3, under the low (10 particles/L) NP treatment, the SOD activity was significantly promoted but inhibited with a high concentration of NPs (104 items/L) compared with the control. The trend of CAT enzyme activity was similar to that of SOD enzyme activity.
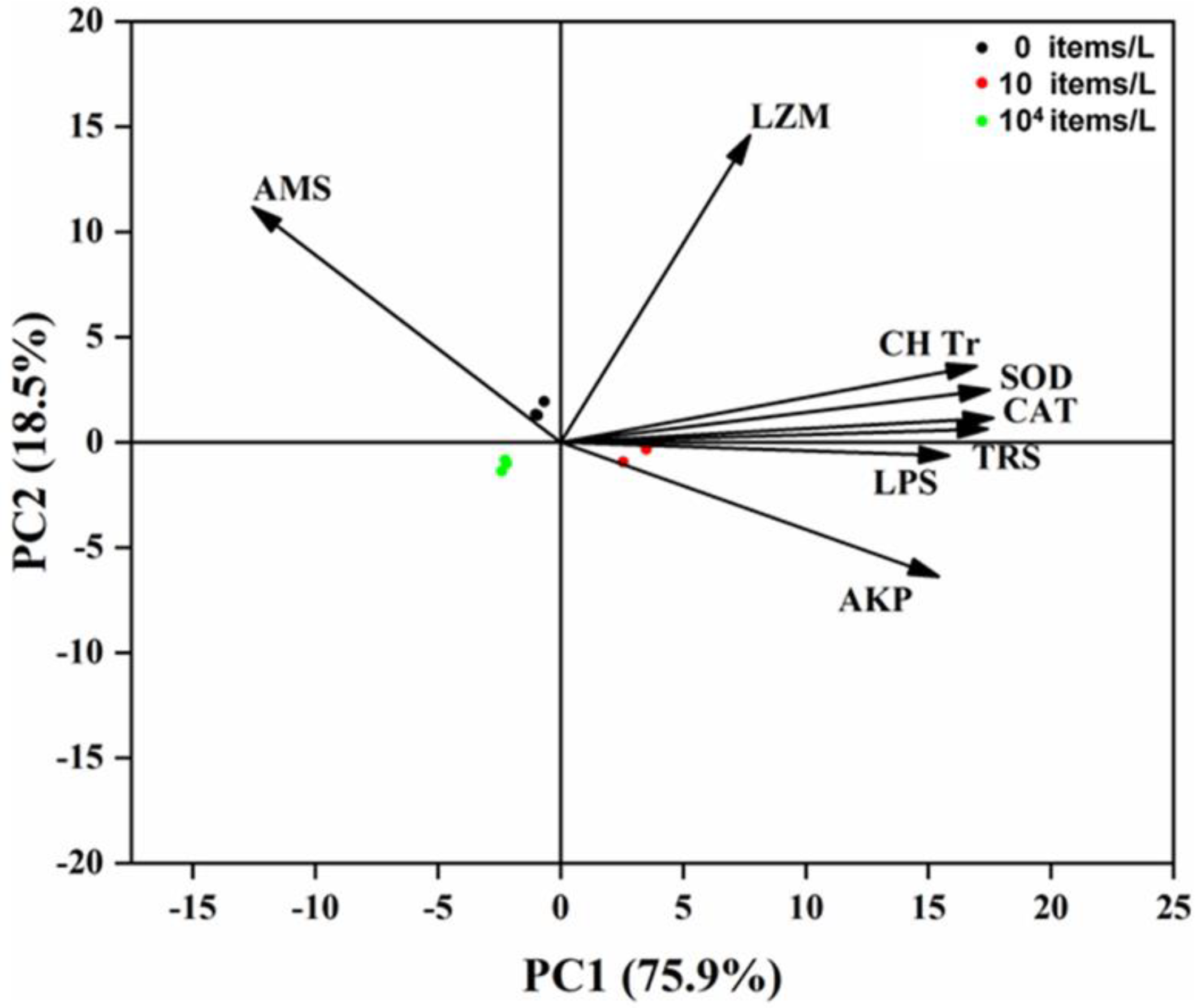
3.4. Principal Component Analysis
The PC1 accounted for 75.9% of all variables (Figure 4). Unexpectedly, the low-concentration (10 items/L) exposure treatment was separated from other treatments due to a majority of tested variables induced by the low concentration of NPs, including CH Tr, TRS, LPS, AKP, SOD, and CAT.
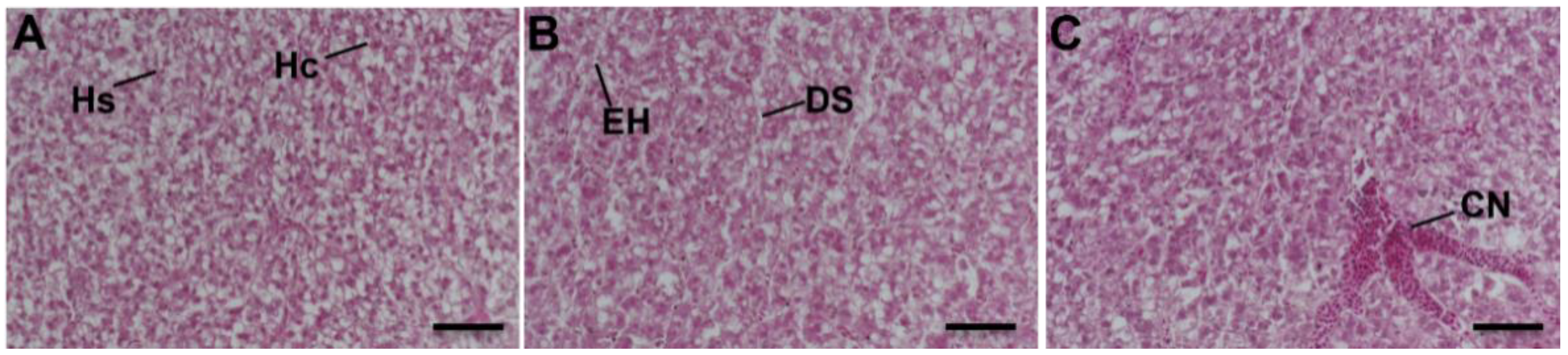
3.5. Liver Histopathology
The tissue structure of the liver was normal in the control group (Figure 5A). Hepatocytes are irregular polygons with large and round nuclei, generally located near one-third of the hepatic sinusoids. The hepatocytes were arranged in two rows in a plate shape and the hepatocyte cords were scattered around the central vein. Between adjacent hepatocytes, cords were irregular and interconnected hepatic sinusoids. Hepatocyte cords were bent, branched, and anastomotic. In the low-concentration NPs group, edema of hepatocytes (EH) and dilated sinusoids (DS) were observed (Figure 5B). The high-concentration NPs group presented congestion (Figure 5C).
4. Discussion
4.1. Effects of Long-Term Nanoplastics Exposure on Digestive Enzymes
Digestive enzyme activity is a sensitive indicator of digestion and absorption capacity in the liver [30]. Digestive enzymes contribute to hydrolyzing lipids, carbohydrates, and proteins for energy production [31]. In this study, the CH Tr and TRS enzyme activities in the low-concentration NPs group were significantly promoted compared with the control. This suggests that a low NP concentration induced the activity of digestive enzymes in medaka, attempting to enhance the digestion of NPs. Although not investigated in the present study, the enhanced digestive activities could lead to higher energy consumption because of the lack of nutritional value of plastics [11,32] and further result in an imbalance in energy metabolism and net energy loss [31,33]. Another possible explanation for the increase in digestive enzymes is that this is a compensatory secretory response to the low nutrient content of NPs and NP-induced starvation to improve digestion and absorption capacity [11]. A significant enhancement in the digestive enzymes (CH Tr and TRS) due to food restriction was observed in the sea bass Dicentrarchus labrax larvae [34]. Romano et al. (2018) found that the activities of TRS and CH Tr were significantly elevated in silver barb Barbodes gonionotus fry exposed to MPs [11].
Some digestive enzymes in the liver of medaka were inhibited after chronic NP exposure in this study. A significant decrease at high concentration (104 items/L) of NPs in CH Tr and TRS activity may be due to depletion of digestive enzymes, leading to digestive system failure [11]. Further, medaka may produce abnormal satiety due to the ingestion of high concentrations of NPs [35]. The activity of AMS showed a significant decrease in all NPs-treated groups, which implies that NPs inhibit carbohydrate metabolism [36]. Similarly, a reduction in digestive enzyme activities was found in different organisms upon NP exposure, e.g., the juvenile guppy (Poecilia reticulata) [33], juvenile large yellow croaker (Larimichthys crocea) [2], juvenile orange-spotted groupers (Epinephelus coioides), and thick-shell mussels (Mytilus coruscus) [37]. A decline in digestive enzymes due to exposure to plastics can reduce an organism’s energy reserves, as reduced digestibility limits the organism’s ability to obtain energy from food [31]. Kong et al. (2019) found that the decrease in digestive enzyme activity on marine mussels might be due to alternations in energy allocation and food intake [38].
4.2. Effects of Long-Term Nanoplastics Exposure on Innate Immune Enzymes
LZM and AKP have been used as effective indicators of immunity status in aquatic organisms [39,40]. LZM is considered to be a humoral defense factor in fish for combating external stressors [41]. In the immune system of fish, LZM is a lysosomal enzyme that fights against the invasion of foreign pathogens or viruses by lysing peptidoglycan in bacterial cell walls [1]. Our study showed that LZM in liver was significantly decreased in the medaka exposed to a high concentration of (104 items/L) NPs, which is consistent with previous studies conducted on blood clams Tegillarca granosa [42], juvenile large yellow croaker Larimichthys crocea [2], and juvenile Macrobrachium nipponense [1]. These results suggest that exposure to high levels of NPs may weaken lysosomal degradation ability and hamper the phagocytosis of pathogens. Based on the alterations in LZM activity, we speculated that the stress level had exceeded the LZM regulation capacity in the medaka’s immune defense system because of the prolonged exposure to NPs, resulting in damage to the immune system. AKP is the major phosphohydrolase in lysosomes [43] and has been applied as an indicator of immune response to environmental pollutants [44]. AKP can boost pathogen recognition and phagocytosis by altering the pathogen’s membrane structure and can also directly destroy pathogens by hydrolyzing phosphates present on their surface [45]. In the present study, after exposure to the low concentration of NPs (10 items/L), the AKP enzyme activity increased significantly in comparison with the control. This result suggests that a low concentration of NPs can activate the medaka’s immune system to fight against internalized NPs by accelerating the release of the AKP enzyme. Similar results in AKP activity were found in Mytilus, juvenile Macrobrachium nipponense, and Symphodon aequifasciatus [1,46,47]. The decrease in AKP activity in the high-concentration NP group may be attributed to the lysosomal membrane damage and permeability changes [48].
4.3. Effects of Long-Term Nanoplastics Exposure on Oxidase Enzymes
One of the plausible mechanisms of general toxicity of NPs is oxidative stress, which occurs when there is an imbalance between the production and elimination of reactive oxygen species (ROS) [23,49]. To maintain the balance, fish form a well-developed antioxidant defense system to scavenge excess ROS [50]. The antioxidant system mainly comprises both free radical scavengers and specific antioxidant enzymes (e.g., SOD, CAT, and GPX) [51]. Changes in antioxidant enzymes are often used as early indicators of oxidative stress [52]. Antioxidant enzymes can remove excess cellular ROS and other pro-oxidants [53]. SOD and CAT are key enzymes in the antioxidant defense system [54]. SOD can convert superoxide anion to H2O2, and then CAT catalyzes H2O2 into H2O and O2 [55]. The results in our study show that compared with control, an increase in the SOD and CAT enzyme activities was observed in the liver of medaka treated with a low concentration of (10 items/L) NPs, suggesting an activated antioxidant defense system. Similar results were observed in several other organisms such as Macrobrachium nipponense, Danio rerio, and Eriocheir sinensis [1,7,23,26]. At a higher concentration (104 items/L), chronic exposure to NPs inhibited the activities of SOD and CAT in the liver of medaka. This inhibition could be associated with the energy expenditure of the oxidative stress response to NPs exposure. Likewise, Li et al. (2020) stated that exposure to a high concentration of NPs (diameter 75 nm; 10, 20, 40 mg/L) reduced the activities of SOD and CAT in Macrobrachium nipponense, damaging the antioxidant defense system [1]. The decrease in antioxidant enzyme activity after prolonged exposure may be due to the gradual loss of resistance to ROS in medaka [56,57]. In summary, exposure to a low concentration of NPs enhances the antioxidant enzyme activities, attempting to maintain redox balance by eliminating excess ROS; exposure to a high concentration of NPs inhibits antioxidant enzyme activity and produces toxic effects due to its ROS exceeding the subscavenging threshold of the antioxidant defense system. PCA analysis showed that, in general, low-concentration NPs stimulated the enzyme activity and high-concentration NPs caused a significant inhibition in liver enzymatic activities.
4.4. Histopathological Effects of Long-Term Nanoplastics Exposure on Liver
Histopathological changes caused by environmental pollutants are direct biomarkers and physiological endpoints of toxicity in fish [58,59,60]. Fish liver histopathology is a sensitive tool used to indirectly detect the occurrence of toxic pollutants in the environment [61,62]. Many studies suggest that the liver tissue is likely the main target organ of nanomaterials [63,64]. In this study, changes in liver enzymatic activities and structural damage of the liver confirm that chronic NP exposure impairs liver functions in medaka. Nanomaterial exposure resulted in hepatic sinusoids, hepatic edema, and hyperemia in the liver of goldfish [7]. It has been reported that under the general stress of environmental pollutants, the liver sinusoids can respond and dilate, which increases the material exchange capacity of the liver tissue, thereby reducing the liver damage of the organism. In addition, due to the sinusoid dilation, the exchange of substances between hepatocytes and blood could change, resulting in liver tissue edema [61,65,66,67]. The induced oxidative species may be associated with tissue damage (e.g., vacuolation, infiltration, and necrosis), which has been demonstrated in Epinepehlus coioides following exposure to NPs [68].
5. Conclusions
This study showed that long-term exposure to NPs at two relatively low concentrations (10 items/L and 104 items/L) negatively affected the liver health of medaka fish. NP treatment led to significant alterations in digestive, innate immune, and antioxidant enzymes and liver tissue damage, suggesting that the hepatotoxic effects of NPs on medaka were likely associated with the impact of NPs on digestive, immune, and antioxidant defense systems. Despite the limitations of this study (e.g., single polymer type, the single time point of evaluation, no confirmative data on NP concentration and uptake), these new findings improve our understanding of the chronic effects of PS-NPs in fish and highlight the need to systematically assess the long-term impacts NPs on other aquatic organisms.
Author Contributions
Methodology, investigation, data curation, analysis, writing: Y.Z. and L.Z.; visualization: H.X.; funding, writing—review and editing: E.G.X., Y.W. and M.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Key R&D Program of China (2018YFD0901205) and National Natural Science Foundation of China (31672700). E.G.X. acknowledges the support of the Department of Biology, University of Southern Denmark, and Danmarks Frie Forskningsfond.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Shanghai Ocean University Animal Care and Use Committee with approval number SHOU-2021-118.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, Y.; Liu, Z.; Li, M.; Jiang, Q.; Wu, D.; Huang, Y.; Jiao, Y.; Zhang, M.; Zhao, Y. Effects of Nanoplastics on Antioxidant and Immune Enzyme Activities and Related Gene Expression in Juvenile Macrobrachium Nipponense. J. Hazard. Mater. 2020, 398, 122990. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Wang, S.; Wang, X.; Yu, X.; Hu, M.; Huang, W.; Wang, Y. Nanoplastics Impair the Intestinal Health of the Juvenile Large Yellow Croaker Larimichthys Crocea. J. Hazard. Mater. 2020, 397, 122773. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Ma, J.; Ji, R.; Pan, K.; Miao, A.J. Microplastics in Aquatic Environments: Occurrence, Accumulation, and Biological Effects. Sci. Total Environ. 2020, 703, 134699. [Google Scholar] [CrossRef] [PubMed]
- Shim, W.J.; Thomposon, R.C. Microplastics in the Ocean. Arch. Environ. Contam. Toxicol. 2015, 69, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Chen, Q.; Jiang, W.; Wang, L.; Xie, H.; Kalogerakis, N.; Ma, Y.; Ji, R. A Carbon-14 Radiotracer-Based Study on the Phototransformation of Polystyrene Nanoplastics in Water: Versus in Air. Environ. Sci. Nano 2019, 6, 2907–2917. [Google Scholar] [CrossRef]
- Zhang, B.; Chao, J.; Chen, L.; Liu, L.; Yang, X.; Wang, Q. Research Progress of Nanoplastics in Freshwater. Sci. Total Environ. 2021, 757, 143791. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, Y.; Deng, Y.; Jiang, W.; Zhao, Y.; Geng, J.; Ding, L.; Ren, H. Uptake and Accumulation of Polystyrene Microplastics in Zebrafish (Danio rerio) and Toxic Effects in Liver. Environ. Sci. Technol. 2016, 50, 4054–4060. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, B.; Bezirci, G.; Çağan, A.S.; Coppens, J.; Levi, E.E.; Oluz, Z.; Tuncel, E.; Duran, H.; Beklioğlu, M. Food Web Effects of Titanium Dioxide Nanoparticles in an Outdoor Freshwater Mesocosm Experiment. Nanotoxicology 2016, 10, 902–912. [Google Scholar] [CrossRef]
- Xie, S.; Zhou, A.; Wei, T.; Li, S.; Yang, B.; Xu, G.; Zou, J. Nanoplastics Induce More Serious Microbiota Dysbiosis and Inflammation in the Gut of Adult Zebrafish than Microplastics. Bull. Environ. Contam. Toxicol. 2021, 107, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, C.; Cuesta, A.; Esteban, M.Á. Effects of Dietary Polyvinylchloride Microparticles on General Health, Immune Status and Expression of Several Genes Related to Stress in Gilthead Seabream (Sparus aurata L.). Fish Shellfish Immunol. 2017, 68, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Romano, N.; Ashikin, M.; Teh, J.C.; Syukri, F.; Karami, A. Effects of Pristine Polyvinyl Chloride Fragments on Whole Body Histology and Protease Activity in Silver Barb Barbodes gonionotus Fry. Environ. Pollut. 2018, 237, 1106–1111. [Google Scholar] [CrossRef]
- Jin, Y.; Zhang, X.; Shu, L.; Chen, L.; Sun, L.; Qian, H.; Liu, W.; Fu, Z. Oxidative Stress Response and Gene Expression with Atrazine Exposure in Adult Female Zebrafish (Danio rerio). Chemosphere 2010, 78, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yan, L.; Junaid, M.; Chen, X.; Liao, H.; Gao, D.; Wang, Q.; Zhang, Y.; Wang, J. Impacts of Polystyrene Nanoplastics on the Oxidative Stress, Immune Responses, and Gut Microbiota to Grass Carp (Ctenopharyngodon idella). SSRN Electron. J. 2022, 1–27. [Google Scholar] [CrossRef]
- Li, Z.; Feng, C.; Wu, Y.; Guo, X. Impacts of Nanoplastics on Bivalve: Fluorescence Tracing of Organ Accumulation, Oxidative Stress and Damage. J. Hazard. Mater. 2020, 392, 122418. [Google Scholar] [CrossRef] [PubMed]
- Brandts, I.; Teles, M.; Tvarijonaviciute, A.; Pereira, M.L.; Martins, M.A.; Tort, L.; Oliveira, M. Effects of Polymethylmethacrylate Nanoplastics on Dicentrarchus Labrax. Genomics 2018, 110, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Ding, R.; Ma, Y.; Sun, Q.; Ren, X.; Sun, Z.; Duan, J. Cardiovascular Toxicity Assessment of Polyethylene Nanoplastics on Developing Zebrafish Embryos. Chemosphere 2021, 282, 131124. [Google Scholar] [CrossRef]
- Mattsson, K.; Johnson, E.V.; Malmendal, A.; Linse, S.; Hansson, L.A.; Cedervall, T. Brain Damage and Behavioural Disorders in Fish Induced by Plastic Nanoparticles Delivered through the Food Chain. Sci. Rep. 2017, 7, 11452. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Cai, M.; Wu, D.; Yu, P.; Jiao, Y.; Jiang, Q.; Zhao, Y. Effects of Nanoplastics at Predicted Environmental Concentration on Daphnia Pulex after Exposure through Multiple Generations. Environ. Pollut. 2020, 256, 113506. [Google Scholar] [CrossRef] [PubMed]
- Botterell, Z.L.R.; Beaumont, N.; Dorrington, T.; Steinke, M.; Thompson, R.C.; Lindeque, P.K. Bioavailability and Effects of Microplastics on Marine Zooplankton: A Review. Environ. Pollut. 2019, 245, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Rist, S.; Baun, A.; Hartmann, N.B. Ingestion of Micro- and Nanoplastics in Daphnia Magna—Quantification of Body Burdens and Assessment of Feeding Rates and Reproduction. Environ. Pollut. 2017, 228, 398–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comfort, K.K.; Braydich-Stolle, L.K.; Maurer, E.I.; Hussain, S.M. Less Is More: Long-Term in vitro Exposure to Low Levels of Silver Nanoparticles Provides New Insights for Nanomaterial Evaluation. ACS Nano 2014, 8, 3260–3271. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Iwasaki, N.; Chen, K.; Shimasaki, Y.; Oshima, Y. Tributyltin and Perfluorooctane Sulfonate Play a Synergistic Role in Promoting Excess Fat Accumulation in Japanese Medaka (Oryzias latipes) via in ovo Exposure. Chemosphere 2019, 220, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Gu, H.; Chang, X.; Huang, W.; Sokolova, I.M.; Wei, S.; Sun, L.; Li, S.; Wang, X.; Hu, M.; et al. Oxidative Stress Induced by Nanoplastics in the Liver of Juvenile Large Yellow Croaker Larimichthys crocea. Mar. Pollut. Bull. 2021, 170, 112661. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jia, X.; Zhu, H.; Zhang, Q.; He, Y.; Shen, Y.; Xu, X.; Li, J. The Effects of Exposure to Microplastics on Grass Carp (Ctenopharyngodon Idella) at the Physiological, Biochemical, and Transcriptomic Levels. Chemosphere 2022, 286, 131831. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Q.; Le Manach, S.; Sotton, B.; Huet, H.; Duvernois-Berthet, E.; Paris, A.; Duval, C.; Ponger, L.; Marie, A.; Blond, A.; et al. Deep Sexual Dimorphism in Adult Medaka Fish Liver Highlighted by Multi-Omic Approach. Sci. Rep. 2016, 6, 32459. [Google Scholar] [CrossRef] [PubMed]
- Mathalon, A.; Hill, P. Microplastic Fibers in the Intertidal Ecosystem Surrounding Halifax Harbor, Nova Scotia. Mar. Pollut. Bull. 2014, 81, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Liu, Z.; Wu, D.; Chen, M.; Lv, W.; Zhao, Y. Accumulation of Polystyrene Microplastics in Juvenile Eriocheir Sinensis and Oxidative Stress Effects in the Liver. Aquat. Toxicol. 2018, 200, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Gui, L.; Liu, R.; Hong, Y.; Li, M. Medaka Oct4 Is Essential for Gastrulation, Central Nervous System Development and Angiogenesis. Gene 2020, 733, 144270. [Google Scholar] [CrossRef]
- Candiano, G.; Bruschi, M.; Musante, L.; Santucci, L.; Ghiggeri, G.M.; Carnemolla, B.; Orecchia, P.; Zardi, L.; Righetti, P.G. Blue Silver: A Very Sensitive Colloidal Coomassie G-250 Staining for Proteome Analysis. Electrophoresis 2004, 25, 1327–1333. [Google Scholar] [CrossRef]
- Wang, Q.; Huang, F.; Liang, K.; Niu, W.; Duan, X.; Jia, X.; Wu, X.; Xu, P.; Zhou, L. Polystyrene Nanoplastics Affect Digestive Function and Growth in Juvenile Groupers. Sci. Total Environ. 2022, 808, 152098. [Google Scholar] [CrossRef]
- Trestrail, C.; Walpitagama, M.; Miranda, A.; Nugegoda, D.; Shimeta, J. Microplastics Alter Digestive Enzyme Activities in the Marine Bivalve, Mytilus Galloprovincialis. Sci. Total Environ. 2021, 779, 146418. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.L.; Rowe, D.; Thompson, R.C.; Galloway, T.S. Microplastic Ingestion Decreases Energy Reserves in Marine Worms. Curr. Biol. 2013, 23, R1031–R1033. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.N.; Wen, B.; Zhu, J.G.; Zhang, Y.S.; Gao, J.Z.; Chen, Z.Z. Exposure to Microplastics Impairs Digestive Performance, Stimulates Immune Response and Induces Microbiota Dysbiosis in the Gut of Juvenile Guppy (Poecilia Reticulata). Sci. Total Environ. 2020, 733, 138929. [Google Scholar] [CrossRef] [PubMed]
- Cara, B.; Moyano, F.J.; Zambonino, J.L.; Fauvel, C. Trypsin and Chymotrypsin as Indicators of Nutritional Status of Post-Weaned Sea Bass Larvae. J. Fish Biol. 2007, 70, 1798–1808. [Google Scholar] [CrossRef]
- Feng, S.; Zeng, Y.; Cai, Z.; Wu, J.; Chan, L.L.; Zhu, J.; Zhou, J. Polystyrene Microplastics Alter the Intestinal Microbiota Function and the Hepatic Metabolism Status in Marine Medaka (Oryzias melastigma). Sci. Total Environ. 2021, 759, 143558. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Gui, L.; Wei, W.; Xu, E.G.; Zhu, Y.; Zhou, W.; Sokolova, I.M.; Li, M.; Wang, Y. Surprisingly Low Particle Concentrations of Nanoplastics Impair the Gut Health of Medaka. Environ. Sci. Nano 2022. in peer review. [Google Scholar]
- Wang, X.; Huang, W.; Wei, S.; Shang, Y.; Gu, H.; Wu, F.; Lan, Z.; Hu, M.; Shi, H.; Wang, Y. Microplastics Impair Digestive Performance but Show Little Effects on Antioxidant Activity in Mussels under Low PH Conditions. Environ. Pollut. 2020, 258, 113691. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Wu, F.; Jiang, X.; Wang, T.; Hu, M.; Chen, J.; Huang, W.; Bao, Y.; Wang, Y. Nano-TiO2 Impairs Digestive Enzyme Activities of Marine Mussels under Ocean Acidification. Chemosphere 2019, 237, 124561. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Li, H.; Wang, X.; Li, X.; Liu, Y.; Sun, J.; Liu, C. A Review of the Immune Molecules in the Sea Cucumber. Fish Shellfish Immunol. 2015, 44, 1–11. [Google Scholar] [CrossRef]
- Jiang, J.; Zhou, Z.; Dong, Y.; Jiang, B.; Chen, Z.; Yang, A.; Wang, B.; Guan, X.; Gao, S.; Sun, H. The in vitro Effects of Divalent Metal Ions on the Activities of Immune-Related Enzymes in Coelomic Fluid from the Sea Cucumber Apostichopus japonicus. Aquac. Res. 2016, 47, 1269–1276. [Google Scholar] [CrossRef]
- Wang, G.; Wang, T.; Zhang, X.; Chen, J.; Feng, C.; Yun, S.; Cheng, Y.; Cheng, F.; Cao, J. Sex-Specific Effects of Fluoride and Lead Exposures on Histology, Antioxidant Physiology, and Immune System in the Liver of Zebrafish (Danio rerio). Ecotoxicology 2022, 31, 396–414. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Rong, J.; Guan, X.; Zha, S.; Shi, W.; Han, Y.; Du, X.; Wu, F.; Huang, W.; Liu, G. Immunotoxicity of Microplastics and Two Persistent Organic Pollutants Alone or in Combination to a Bivalve Species. Environ. Pollut. 2020, 258, 113845. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yu, P.; Cai, M.; Wu, D.; Zhang, M.; Chen, M.; Zhao, Y. Effects of Microplastics on the Innate Immunity and Intestinal Microflora of Juvenile Eriocheir Sinensis. Sci. Total Environ. 2019, 685, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Giri, S.S.; Jun, J.W.; Kim, H.J.; Kim, S.W.; Yun, S.; Park, S.C. Effects of Algal Toxin Okadaic Acid on the Non-Specific Immune and Antioxidant Response of Bay Scallop (Argopecten irradians). Fish Shellfish Immunol. 2017, 65, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Lin, T.; Zhan, W. Variations of Enzyme Activities in the Haemocytes of Scallop Chlamys Farreri after Infection with the Acute Virus Necrobiotic Virus (AVNV). Fish Shellfish Immunol. 2008, 25, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Canesi, L.; Ciacci, C.; Bergami, E.; Monopoli, M.P.; Dawson, K.A.; Papa, S.; Canonico, B.; Corsi, I. Evidence for Immunomodulation and Apoptotic Processes Induced by Cationic Polystyrene Nanoparticles in the Hemocytes of the Marine Bivalve Mytilus. Mar. Environ. Res. 2015, 111, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Jin, S.R.; Chen, Z.Z.; Gao, J.Z.; Liu, Y.N.; Liu, J.H.; Feng, X.S. Single and Combined Effects of Microplastics and Cadmium on the Cadmium Accumulation, Antioxidant Defence and Innate Immunity of the Discus Fish (Symphysodon Aequifasciatus). Environ. Pollut. 2018, 243, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.B.; Wang, G.; Chan, S.F. Effects of Artificial Infection of Litopenaeus Vannamei by Micrococcus Ysodeikticus and WSSV on the Activity of Immunity Related Enzymes. Fish Shellfish Immunol. 2015, 46, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Palić, D. Micro- and Nano-Plastics Activation of Oxidative and Inflammatory Adverse Outcome Pathways. Redox Biol. 2020, 37, 101620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shen, H.; Wang, X.; Wu, J.; Xue, Y. Effects of Chronic Exposure of 2,4-Dichlorophenol on the Antioxidant System in Liver of Freshwater Fish Carassius auratus. Chemosphere 2004, 55, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, D.R. Contaminant-Stimulated Reactive Oxygen Species Production and Oxidative Damage in Aquatic Organisms. Mar. Pollut. Bull. 2001, 42, 656–666. [Google Scholar] [CrossRef]
- Egea, J.; Fabregat, I.; Frapart, Y.M.; Ghezzi, P.; Görlach, A.; Kietzmann, T.; Kubaichuk, K.; Knaus, U.G.; Lopez, M.G.; Olaso-Gonzalez, G.; et al. European Contribution to the Study of ROS: A Summary of the Findings and Prospects for the Future from the COST Action BM1203 (EU-ROS). Redox Biol. 2017, 13, 94–162. [Google Scholar] [CrossRef] [PubMed]
- Ajima, M.N.O.; Pandey, P.K.; Kumar, K.; Poojary, N. Neurotoxic Effects, Molecular Responses and Oxidative Stress Biomarkers in Nile Tilapia, Oreochromis niloticus (Linnaeus, 1758) Exposed to Verapamil. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 196, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Dorantes-Aranda, J.J.; Seger, A.; Mardones, J.I.; Nichols, P.D.; Hallegraeff, G.M. Progress in Understanding Algal Bloom-Mediated Fish Kills: The Role of Superoxide Radicals, Phycotoxins and Fatty Acids. PLoS ONE 2015, 10, e0133549. [Google Scholar]
- de Andrade, L.L.; do Espirito Santo Pereira, A.; Fernandes Fraceto, L.; Bueno dos Reis Martinez, C. Can Atrazine Loaded Nanocapsules Reduce the Toxic Effects of This Herbicide on the Fish Prochilodus Lineatus? A Multibiomarker Approach. Sci. Total Environ. 2019, 663, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Yang, Y.; Tao, Y.; Jiang, Y.; Chen, B.; Zhu, X.; Cai, Z.; Li, B. A Mechanism Study on Toxicity of Graphene Oxide to Daphnia Magna: Direct Link between Bioaccumulation and Oxidative Stress. Environ. Pollut. 2018, 234, 953–959. [Google Scholar] [CrossRef]
- Company, R.; Serafim, A.; Cosson, R.P.; Fiala-Médioni, A.; Camus, L.; Colaço, A.; Serrão-Santos, R.; Bebianno, M.J. Antioxidant Biochemical Responses to Long-Term Copper Exposure in Bathymodiolus azoricus from Menez-Gwen Hydrothermal Vent. Sci. Total Environ. 2008, 389, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Ullah, S.; Li, Z.; Hasan, Z.; Khan, S.U.; Fahad, S. Malathion Induced Oxidative Stress Leads to Histopathological and Biochemical Toxicity in the Liver of Rohu (Labeo rohita, Hamilton) at Acute Concentration. Ecotoxicol. Environ. Saf. 2018, 161, 270–280. [Google Scholar] [CrossRef]
- Bucke, D.; Venthaak, D.; Lang, T.; Mellergaard, S. Times No. 19 Common Diseases and Parasites of Fish in the North Atlantic: Training Guide for Identification; International Council for the Exploration of the Sea (ICES): Copenhagen, Denmark, 1996. [Google Scholar]
- Ullah, S.; Begum, M.; Dhama, K.; Ahmad, S.; Hassan, S.; Alam, I. Malathion Induced DNA Damage in Freshwater Fish, Labeo Rohita (Hamilton, 1822) Using Alkaline Single Cell Gel Electrophoresis. Asian J. Anim. Vet. Adv. 2016, 11, 98–105. [Google Scholar] [CrossRef]
- Ahmed, M.K.; Habibullah-Al-Mamun, M.; Parvin, E.; Akter, M.S.; Khan, M.S. Arsenic Induced Toxicity and Histopathological Changes in Gill and Liver Tissue of Freshwater Fish, Tilapia (Oreochromis mossambicus). Exp. Toxicol. Pathol. 2013, 65, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Menard, A.; Drobne, D.; Jemec, A. Ecotoxicity of Nanosized TiO2. Review of in Vivo Data. Environ. Pollut. 2011, 159, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Anderson, D.; Dhawan, A. Zinc Oxide Nanoparticles Induce Oxidative DNA Damage and ROS-Triggered Mitochondria Mediated Apoptosis in Human Liver Cells (HepG2). Apoptosis 2012, 17, 852–870. [Google Scholar] [CrossRef] [PubMed]
- Decuzzi, P.; Godin, B.; Tanaka, T.; Lee, S.Y.; Chiappini, C.; Liu, X.; Ferrari, M. Size and Shape Effects in the Biodistribution of Intravascularly Injected Particles. J. Control. Release 2010, 141, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Devi, G.P.; Ahmed, K.B.A.; Varsha, M.K.N.S.; Shrijha, B.S.; Lal, K.K.S.; Anbazhagan, V.; Thiagarajan, R. Sulfidation of Silver Nanoparticle Reduces Its Toxicity in Zebrafish. Aquat. Toxicol. 2015, 158, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Rossbach, L.M.; Shaw, B.J.; Piegza, D.; Vevers, W.F.; Atfield, A.J.; Handy, R.D. Sub-Lethal Effects of Waterborne Exposure to Copper Nanoparticles Compared to Copper Sulphate on the Shore Crab (Carcinus maenas). Aquat. Toxicol. 2017, 191, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Ben Ameur, W.; de Lapuente, J.; El Megdiche, Y.; Barhoumi, B.; Trabelsi, S.; Camps, L.; Serret, J.; Ramos-López, D.; Gonzalez-Linares, J.; Driss, M.R.; et al. Oxidative Stress, Genotoxicity and Histopathology Biomarker Responses in Mullet (Mugil Cephalus) and Sea Bass (Dicentrarchus labrax) Liver from Bizerte Lagoon (Tunisia). Mar. Pollut. Bull. 2012, 64, 241–251. [Google Scholar] [CrossRef]
- Brandts, I.; Cánovas, M.; Tvarijonaviciute, A.; Llorca, M.; Vega, A.; Farré, M.; Pastor, J.; Roher, N.; Teles, M. Nanoplastics Are Bioaccumulated in Fish Liver and Muscle and Cause DNA Damage after a Chronic Exposure. Environ. Res. 2022, 212, 113433. [Google Scholar] [CrossRef]
Figure 1.
(A) CH Tr, (B) TRS, (C) LPS, and (D) AMS activities in the liver of medaka exposed to NPs (0, 10, 104 items/L) for three months (n = 8). Different lowercase letters indicate significant differences among treatments (p < 0.05).
Figure 1.
(A) CH Tr, (B) TRS, (C) LPS, and (D) AMS activities in the liver of medaka exposed to NPs (0, 10, 104 items/L) for three months (n = 8). Different lowercase letters indicate significant differences among treatments (p < 0.05).

Figure 2.
(A) LZM and (B) AKP activities in the liver of medaka exposed to NPs (0, 10, 104 items/L) for three months (n = 8). Different lowercase letters indicate significant differences among treatments (p < 0.05).
Figure 2.
(A) LZM and (B) AKP activities in the liver of medaka exposed to NPs (0, 10, 104 items/L) for three months (n = 8). Different lowercase letters indicate significant differences among treatments (p < 0.05).

Figure 3.
(A) SOD and (B) CAT activities in the liver of medaka exposed to NPs (0, 10, 104 items/L) for three months (n = 8). Different lowercase letters indicate significant differences among treatments (p < 0.05).
Figure 3.
(A) SOD and (B) CAT activities in the liver of medaka exposed to NPs (0, 10, 104 items/L) for three months (n = 8). Different lowercase letters indicate significant differences among treatments (p < 0.05).

Figure 4.
Biplot originating from principal component analysis (PCA) integrating biochemical indi−cators (CH Tr, TRS, LPS, AMS, LZM, AKP, SOD, CAT) and three different treatments.
Figure 4.
Biplot originating from principal component analysis (PCA) integrating biochemical indi−cators (CH Tr, TRS, LPS, AMS, LZM, AKP, SOD, CAT) and three different treatments.

Figure 5.
Liver histomorphology of medaka exposed to (A) 0 items/L (control), (B) 10 items/L, and (C) 104 items/L (H&E. Scale bar, 50 μm). Hc, Hepatocyte; Hs, Hepatic sinusoid; DS, Dilated sinusoids; EH, Edema of hepatocytes; CN, Congestion.
Figure 5.
Liver histomorphology of medaka exposed to (A) 0 items/L (control), (B) 10 items/L, and (C) 104 items/L (H&E. Scale bar, 50 μm). Hc, Hepatocyte; Hs, Hepatic sinusoid; DS, Dilated sinusoids; EH, Edema of hepatocytes; CN, Congestion.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhou, Y.; Zhao, L.; Xu, H.; Xu, E.G.; Li, M.; Wang, Y. Long-Term Exposure to Polystyrene Nanoplastics Impairs the Liver Health of Medaka. Water 2022, 14, 2767. https://doi.org/10.3390/w14172767
AMA Style
Zhou Y, Zhao L, Xu H, Xu EG, Li M, Wang Y. Long-Term Exposure to Polystyrene Nanoplastics Impairs the Liver Health of Medaka. Water. 2022; 14(17):2767. https://doi.org/10.3390/w14172767
Chicago/Turabian StyleZhou, Yinfeng, Li Zhao, Haijing Xu, Elvis Genbo Xu, Mingyou Li, and Youji Wang. 2022. "Long-Term Exposure to Polystyrene Nanoplastics Impairs the Liver Health of Medaka" Water 14, no. 17: 2767. https://doi.org/10.3390/w14172767
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.