
Fish Diversity and Abundance Patterns in Small Watercourses of the Central European Plain Ecoregion in Relation to Environmental Factors

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling
2.3. Application of Ecological Indices and Data Analyses
3. Results
3.1. Environmental Characteristics
3.2. Fish Diversity and Abundance Patterns
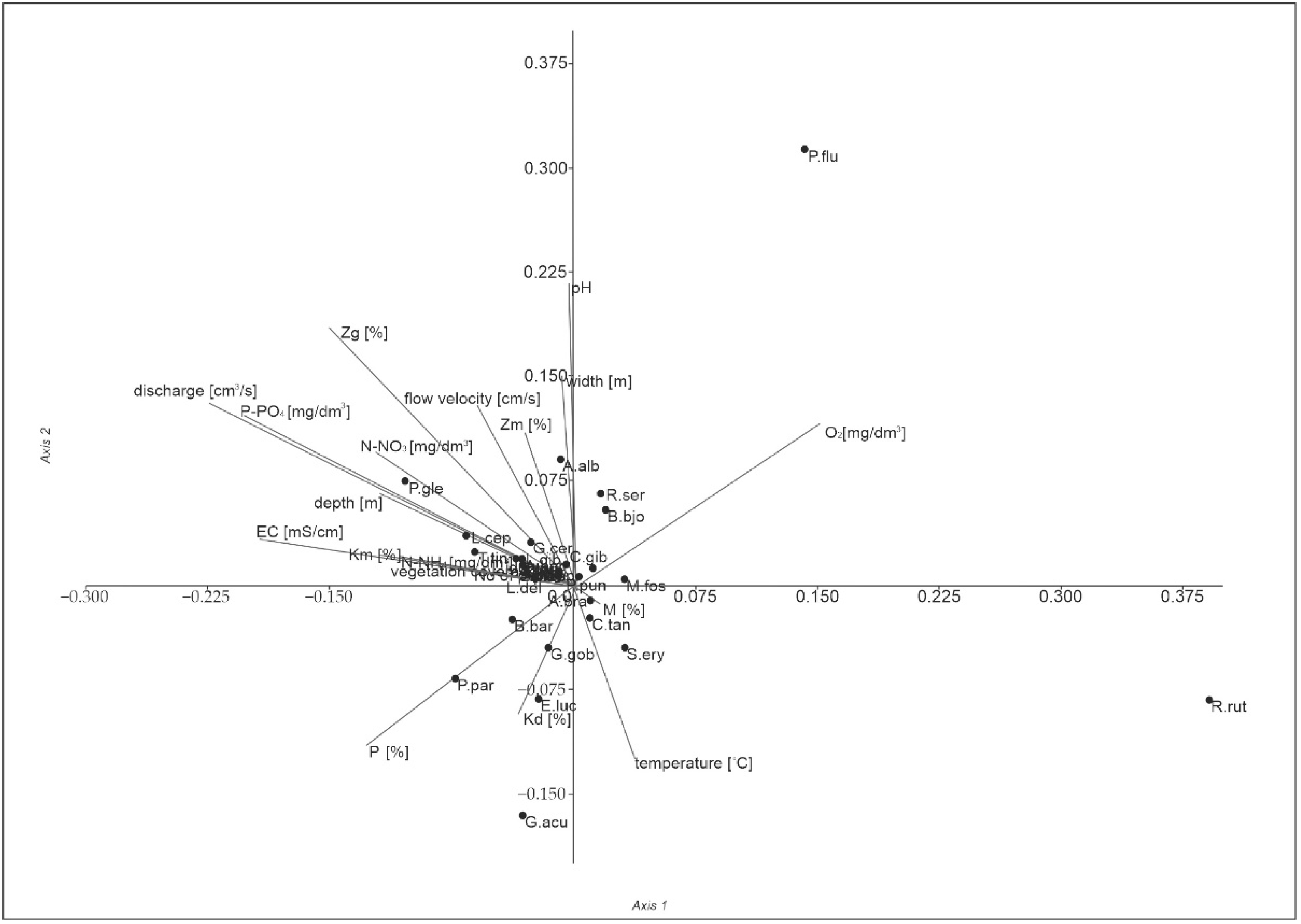
3.3. Effects of Environmental Gradients
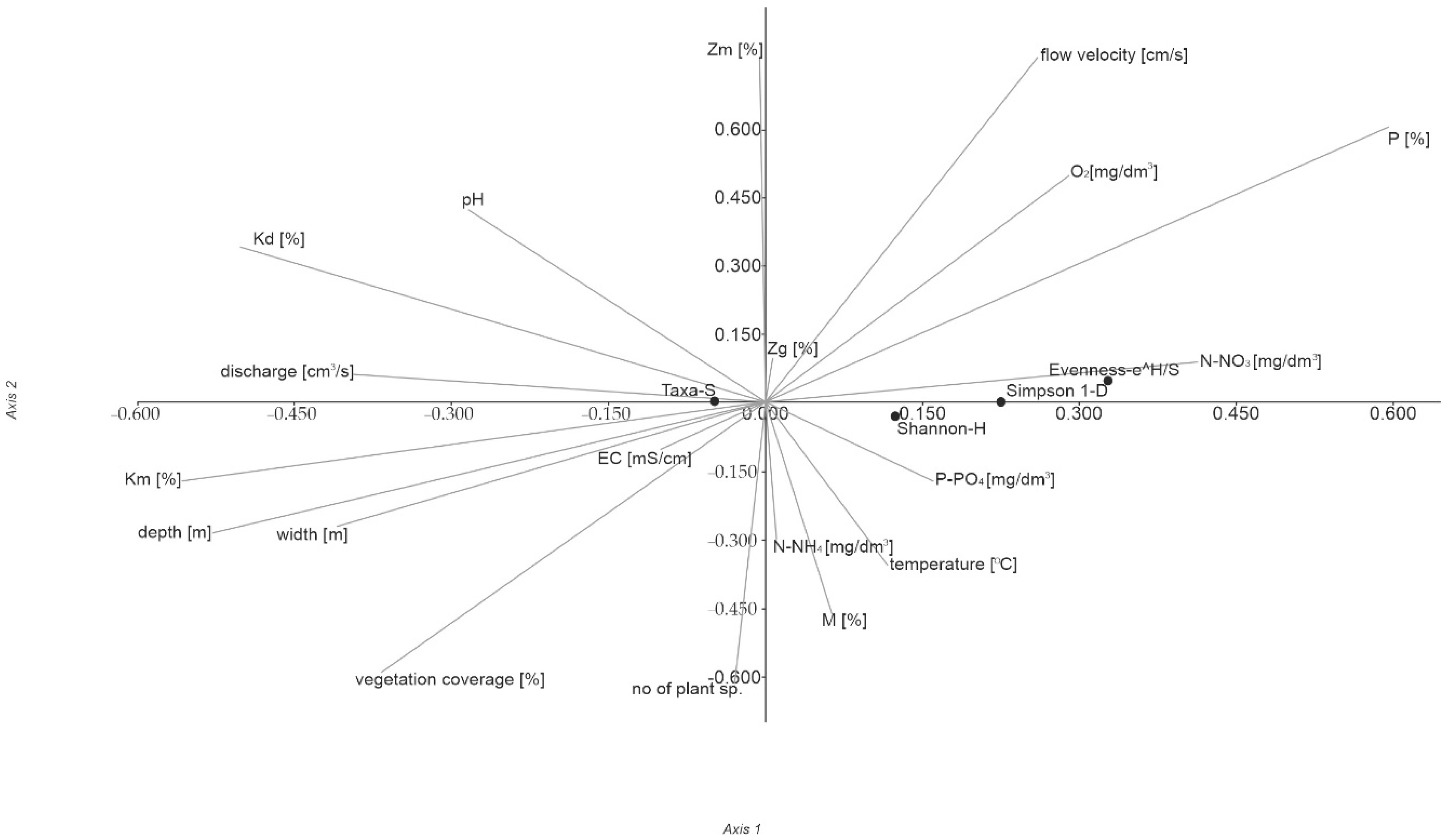
3.4. Biotic Indices
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oglęcki, P.; Ostrowski, P.S.; Utratna-Żukowska, M. Natural and Geomorphological Response of the Small Lowland River Valley for Anthropogenic Transformation. Resources 2021, 10, 97. [Google Scholar] [CrossRef]
- Gheoca, V. The River Valleys As Biodiversity Reservoirs For Land Snails In Highly Anthropic Areas—The Case Of Cisnădie River (Romania). Transylv. Rev. Syst. Ecol. Res. 2016, 18, 83–90. [Google Scholar] [CrossRef]
- Štefunková, Z.; Macura, V.; Doláková, G.; Majorošová, M. Evaluation of the hydro-ecological quality of the aquatic habitat of the Váh River. J. Water Land Dev. 2020, 46, 209–215. [Google Scholar]
- Kaletova, T.; Arifjanov, A.; Samiev, L.; Babajanov, F. Importance of river sediments in soil fertility. J. Water Land Dev. 2022, 52, 21–26. [Google Scholar]
- Kuriata-Potasznik, A.; Szymczyk, S.; Skwierawski, A. Influence of Cascading River–Lake Systems on the Dynamics of Nutrient Circulation in Catchment Areas. Water 2020, 12, 1144. [Google Scholar] [CrossRef]
- Abou Zaki, N.; Torabi Haghighi, A.; Rossi, P.M.; Tourian, M.J.; Bakhshaee, A.; Kløve, B. Evaluating Impacts of Irrigation and Drought on River, Groundwater and a Terminal Wetland in the Zayanderud Basin, Iran. Water 2020, 12, 1302. [Google Scholar] [CrossRef]
- Kelly-Quinn, M.; Bruen, M.; Carlsson, J.; Gurnell, A.; Jarvie, H.; Piggott, J. Managing the small stream network for improved water quality, biodiversity and ecosystem services protection (SSNet). Res. Ideas Outcomes 2019, 5, e33400. [Google Scholar] [CrossRef]
- Okafor, U.P.; Obeta, M.C.; Ayadiuno, R.U.; Onyekwelu, A.C.; Asuoha, G.C.; Eze, E.J.; Orji-Okafor, C.E.; Igboeli, E.E. Health implications of stream water contamination by industrial effluents in the Onitsha urban area of Southeastern Nigeria. J. Water Land Dev. 2021, 48, 105–114. [Google Scholar]
- Bănăduc, D.; Sas, A.; Cianfaglione, K.; Barinova, S.; Curtean-Bănăduc, A. The Role of Aquatic Refuge Habitats for Fish, and Threats in the Context of Climate Change and Human Impact, during Seasonal Hydrological Drought in the Saxon Villages Area (Transylvania, Romania). Atmosphere 2021, 12, 1209. [Google Scholar] [CrossRef]
- Malinowski, Ł.; Skoczko, I. Impacts of Climate Change on Hydrological Regime and Water Resources Management of the Narew River in Poland. J. Ecol. Eng. 2018, 19, 167–175. [Google Scholar] [CrossRef]
- Jakubčinová, K.; Harustiakova, D.; Števove, B.; Švolíková, K.; Makovinská, J.; Kováč, V. Distribution patterns and potential for further spread of three invasive fish species (Neogobius melanostomus, Lepomis gibbosus and Pseudorasbora parva) in Slovakia. Aquat. Invasions 2018, 13, 513–524. [Google Scholar] [CrossRef]
- Loucks, D.P.; van Beek, E. Water Resources Planning and Management: An Overview. In Water Resource Systems Planning and Management; Stedinger, J.R., Dijkman, J.P.M., Villars, M.T., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–49. [Google Scholar]
- Rechulicz, J.; Płaska, W.; Pęczuła, W.; Tarkowska-Kukuryk, M.; Mieczan, T. The structure of ichthyofauna and angling pressure on fish in upper section of Bystrzyca River. Teka Kom. Ochr. Kszt. Środ. Przyr.—OL PAN 2016, 13, 69–79. [Google Scholar]
- Fraker, M.E.; Keitzer, S.C.; Sinclair, J.S.; Aloysius, N.R.; Dippold, D.A.; Haw, Y.; Arnold, G.A.; Daggupati, P.; Johnson, M.-V.V.; Martin, J.F.; et al. Projecting the effects of agricultural conservation practices on stream fish communities in a changing climate. Sci. Total Environ. 2020, 747, 141112. [Google Scholar] [CrossRef] [PubMed]
- Vander Vorste, R.; McElmurray, P.; Bell, S.; Eliason, K.M.; Brown, B.L. Does Stream Size Really Explain Biodiversity Patterns in Lotic Systems? A Call for Mechanistic Explanations. Diversity 2017, 9, 26. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, F.A.; Reid, S.M.; Mandrak, N.E. A habitat-based framework to predict the effects of agricultural drain maintenance on imperilled fishes. J. Environ. Manag. 2018, 206, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Gomolka, Z.; Twarog, B.; Zeslawska, E. State Analysis of the Water Quality in Rivers in Consideration of Diffusion Phenomenon. Appl. Sci. 2022, 12, 1549. [Google Scholar] [CrossRef]
- Brysiewicz, A.; Czerniejewski, P. The Effect of Maintenance Works on Ichthyofauna in the Context of Hydrochemical Conditions of Small Watercourses of Central and North-Western Poland. J. Ecol. Eng. 2019, 20, 82–89. [Google Scholar] [CrossRef]
- Menke, K.; Smith, R.; Pirelli, L.; Van Hoesen, J. Mastering QGIS, 1st ed.; Pack Publishing: Birmingham, UK, 2015; p. 391. [Google Scholar]
- Corine Land Cover 2018. Available online: https://clc.gios.gov.pl/index.php/clc-2018/metadane (accessed on 15 July 2021).
- Wetzel, R.G.; Likens, G.E. Limnological Analysis, 3rd ed.; Springer: New York, NY, USA, 2000; p. 430. [Google Scholar]
- Blair, T.C.; McPherson, J.G. Grain-size and textural classification of coarse sedimentary particles. J. Sediment. Res. 1999, 69, 6–19. [Google Scholar] [CrossRef]
- Shtovba, S.; Petrychko, M. Jaccard index-Based Assessing the Similarity of Research Fields in Dimensions. In Proceedings of the 1st International Workshop on Digital Content & Smart Multimedia (DCSMart 2019), Lviv, Ukraine, 23–25 December 2019; Kryvinska, N., Izonin, I., Greguš, M., Poniszewska-Marańda, A., Dronyuk, I., Eds.; Lviv Polytechnic National University: Lviv, Ukraine, 2019. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9–18. [Google Scholar]
- Pawar, P.R.; Al-Tawaha, A.R.M.S. Diversity indices of macrobenthos for assessment of coastal pollution along Uran coast, Navi Mumbai. Adv. Environ. Biol. 2017, 11, 34–50. [Google Scholar]
- Dorić, S.; Čučuković, A. Community Structure and Diversity of Macrozoobenthos in Quarry Ribnica’s Creek as Indicator of Surface Water Management. Genet. Appl. 2018, 1, 29–35. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.gbif.org/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed on 15 October 2021).
- Crawley, M.J. The R Book, 2nd ed.; Wiley: Chichester, UK, 2013; p. 1051. [Google Scholar]
- Zar, J.D. Biostatistical Analysis, 5th ed.; Pearson: Hoboken, NJ, USA, 2010; p. 944. [Google Scholar]
- Fox, J. Applied Regression Analysis and Generalized Linear Models, 3rd ed.; Sage Publications, Inc.: Los Angeles, CA, USA, 2016; p. 791. [Google Scholar]
- Lander, J.P. R for Everyone. Advanced Analytics and Graphic, 2nd ed.; Pearson: Boston, MA, USA, 2017; p. 531. [Google Scholar]
- Ter Braak, C.J.F.; Verdonschot, P.F.M. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 256–289. [Google Scholar] [CrossRef]
- Bonanno, A.; Zgozi, S.; Basilone, G.; Hamza, M.; Barra, M.; Genovese, S.; Rumolo, P.; Nfate, A.; Elsger, M.; Goncharov, S.; et al. Acoustically detected pelagic fish community in relation to environmental conditions observed in the Central Mediterranean sea: A comparison of Libyan and Sicilian-Maltese coastal areas. Hydrobiologia 2015, 755, 209–224. [Google Scholar] [CrossRef]
- Allan, J.D. Influence of land use and landscape setting on the ecological status of rivers. Limnetica 2004, 23, 187–198. [Google Scholar] [CrossRef]
- Fu, L.; Jiang, Y.; Ding, J.; Liu, Q.; Peng, Q.Z.; Kang, M.Y. Impacts of land use and environmental factors on macroinvertebrate functional feeding groups in the Dongjiang River basin, southeast China. J. Freshw. Ecol. 2016, 31, 21–35. [Google Scholar] [CrossRef]
- Riley, R.H.; Townsend, C.R.; Niyogi, D.K.; Arbuckle, C.A.; Peacock, K.A. Headwater stream response to grassland agricultural development in New Zealand. N. Z. J. Mar. Freshw. Res. 2003, 37, 389–403. [Google Scholar] [CrossRef]
- Degerman, E.; Beier, U.; Breine, J.; Melcher, A.; Quataert, P.; Rogers, C.; Roset, N.; Simoens, I. Classification and Assessment of Degradation in European Running Waters. Fish. Manag. Ecol. 2007, 14, 417–426. [Google Scholar] [CrossRef]
- Murphy, P.N.C.; Mellander, P.E.; Melland, A.R.; Buckley, C.; Shore, M.; Shortle, G.; Wall, D.P.; Treacy, M.; Shine, O.; Mechan, S.; et al. Variable response to phosphorus mitigation measures across the nutrient transfer continuum in a dairy grassland catchment. Agric. Ecosyst. Environ. 2015, 207, 192–202. [Google Scholar] [CrossRef]
- Cavaille, P.; Dumont, B.; Van Looy, K.; Floury, M.; Tabacchi, E.; Evette, A. Influence of riverbank stabilization techniques on taxonomic and functional macrobenthic communities. Hydrobiologia 2018, 807, 19–35. [Google Scholar] [CrossRef]
- Merchan, D.; Casalı, J.; Del Valle de Lersundi, J.; Campo-Bescós, M.A.; Giménez, R.; Preciado, B.; Lafarga, A. Runoff, nutrients, sediment and salt yields in an irrigated watershed in southern Navarre (Spain). Agric. Water Manag. 2018, 195, 120–132. [Google Scholar] [CrossRef]
- Rasmussen, J.J.; McKnight, U.S.; Loinaz, M.C.; Thomsen, N.I.; Olsson, M.E.; Bjerg, P.L.; Binning, P.J.; Kronvang, B. A catchment scale evaluation of multiple stressor effects in headwater streams. Sci. Total Environ. 2013, 442, 420–431. [Google Scholar] [CrossRef]
- Effert-Fanta, E.L.; Fischer, R.U.; Wahl, D.H. Effects of riparian forest buffers and agricultural land use on macroinvertebrate and fish community structure. Hydrobiologia 2019, 841, 45–64. [Google Scholar] [CrossRef]
- Gerth, W.J.; Li, J.; Giannico, G.R. Agricultural land use and macroinvertebrate assemblages in lowland temporary streams of the Willamette Valley, Oregon, USA. Agric. Ecosyst. Environ. 2017, 236, 154–165. [Google Scholar] [CrossRef]
- Fierro, P.; Valdovinos, C.; Arismendi, I.; Díaz, G.; Jara-Flores, A.; Habit, E.; Vargas-Chacoff, L. Examining the influence of human stressors on benthic algae, macroinvertebrate, and fish assemblages in Mediterranean streams of Chile. Sci. Total Environ. 2019, 686, 26–37. [Google Scholar] [CrossRef]
- Tockner, K.; Robinson, C.T.; Uehlinger, U. Rivers of Europe, 1st ed.; Academic Press/Elsevier: London, UK, 2009; p. 728. [Google Scholar]
- Penczak, T.; Galicka, W.; Głowacki, Ł.; Koszaliński, H.; Kruk, A.; Zięba, G.; Grabowska, J.; Marszał, L. Fish assemblage changes relative to environmental factors and time in the Warta River, Poland, and its oxbow lakes. J. Fish Biol. 2003, 64, 483–501. [Google Scholar] [CrossRef]
- Penczak, T. Fish assemblages composition in a natural, then regulated, stream: A quantitative long-term study. Ecol. Modell. 2011, 222, 2103–2118. [Google Scholar] [CrossRef]
- Cheng, S.T.; Herricks, E.E.; Tsai, W.P.; Chang, F.J. Assessing the natural and anthropogenic influences on basin-wide fish species richness. Sci. Total Environ. 2016, 572, 825–836. [Google Scholar] [CrossRef]
- Sutela, T.; Vehanen, T.; Jounela, P. Longitudinal patterns of fish assemblages in European boreal streams. Hydrobiologia 2020, 847, 3277–3290. [Google Scholar] [CrossRef]
- Sutela, T.; Vehanen, T.; Jounela, P.; Aroviita, J. Species–environment relationships of fish and map-based variables in small boreal streams: Linkages with climate change and bioassessment. Ecol. Evol. 2021, 11, 10457–10467. [Google Scholar] [CrossRef]
- Matthews, W.J.; Marsh-Matthews, E. Effects of drought on fish across axes of space, time, and ecological complexity. Freshw. Biol. 2003, 48, 1232–1253. [Google Scholar] [CrossRef]
- Kruk, A. Long-term changes in fish assemblages of the Widawka and Grabia Rivers (Poland): Pattern recognition with a Kohonen artificial neural network. Ann. Limnol.—Int. J. Lim. 2007, 43, 253–269. [Google Scholar] [CrossRef]
- Kruk, A.; Lek, S.; Park, Y.-S.; Penczak, T. Fish assemblages in the large lowland Narew River system (Poland): Application of the self-organizing map algorithm. Ecol. Model. 2007, 203, 45–61. [Google Scholar] [CrossRef]
- Czerniejewski, P.; Czerniawski, R. Marine and Freshwater Fish of Poland, 1st ed.; FREL Scientific Publishing: Warsaw, Poland, 2016; p. 331. [Google Scholar]
- Wolter, C. Conservation of fish species diversity in navigable waterways. Landsc. Urban Plan. 2001, 53, 135–144. [Google Scholar] [CrossRef]
- Kruk, A.; Penczak, T. Impoundment impact on populations of facultative riverine fish. Ann. Limnol.—Int. J. Limnol. 2003, 39, 197–210. [Google Scholar] [CrossRef]
- Lonzarich, D.G.; Quinn, T.P. Experimental evidence for the effect of depth and structure on the distribution, growth, and survival of stream fishes. Can. J. Zool. 1995, 73, 2223–2230. [Google Scholar] [CrossRef]
- Penaluna, B.E.; Dunham, J.; Andersen, H.V. Nowhere to hide: The importance of in stream cover for stream-living Coastal Cutthroat Trout during seasonal low flow. Ecol. Freshw. Fish 2020, 30, 256–269. [Google Scholar] [CrossRef]
- Arthington, A.H.; Bunn, S.E.; Poff, N.L.; Naiman, R.J. The challenge of providing environmental flow rules to sustain river ecosystems. Ecol. Appl. 2016, 16, 1311–1318. [Google Scholar] [CrossRef]
- Bunn, S.E.; Arthington, A.H. Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ. Manag. 2002, 30, 492–507. [Google Scholar] [CrossRef]
- Dewson, Z.S.; James, A.B.W.; Death, R.G. A review of the consequences of decreased flow for instream habitat and macroinvertebrates. J. N. Am. Benthol. Soc. 2007, 26, 401–415. [Google Scholar] [CrossRef]
- Welcomme, R.L.; Winemiller, K.O.; Cowx, I.G. Fish environmental guilds as a tool for assessment of ecological condition of rivers. River Res. Appl. 2005, 22, 377–396. [Google Scholar] [CrossRef]
- Meixner, T.; Huth, A.K.; Brooks, P.D.; Conklin, M.H.; Grimm, N.B.; Bales, R.C.; Haas, P.A.; Petti, J.R. Influence of shifting flow paths on nitrogen concentrations during monsoon floods, San Pedro River, Arizona. J. Geophys. Res. Atmos. 2007, 112, G03S03. [Google Scholar]
- Evans, D.H.; Claiborne, J.B.; Currie, S. (Eds.) The Physiology of Fishes, 4th ed.; CRC Press: Boca Raton, FL, USA, 2014; p. 482. [Google Scholar]
- Roman, M.R.; Brandt, S.B.; Houde, E.D.; Pierson, J.J. Interactive Effects of Hypoxia and Temperature on Coastal Pelagic Zooplankton and Fish. Front. Mar. Sci. 2019, 6, 139. [Google Scholar] [CrossRef]
- Friedman, J.R.; Condon, N.E.; Drazen, J.C. Gill surface area and metabolic enzyme activities of demersal fishes associated with the oxygen minimum zone off California. Limnol. Oceanogr. 2012, 57, 1701–1710. [Google Scholar] [CrossRef]
- Eddy, F.B. Ammonia in estuaries and effects on fish. J. Fish Biol. 2005, 67, 1495–1513. [Google Scholar] [CrossRef]
- Lemarie, G.; Dosdat, A.; Coves, D.; Dutto, G.; Gasset, E.; Person-Le Ruyet, J. Effect of chronic ammonia exposure on growth of European seabass (Dicentrarchus labrax) juveniles. Aquaculture 2004, 229, 479–491. [Google Scholar] [CrossRef]
- Sinha, A.K.; Liew, H.J.; Diricx, M.; Blust, R.; De Boeck, G. The interactive effects of ammonia exposure, nutritional status and exercise on metabolic and physiological responses in gold fish (Carassius auratus L.). Aquat Toxicol. 2012, 109, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yu, N.; Qin, J.G.; Li, E.; Du, Z.; Chen, L. Effects of ammonia stress, dietary linseed oil and Edwardsiella ictaluri challenge on juvenile dark barbel catfish Pelteobagrus vachelli. Fish Shellfish Immunol. 2014, 38, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Camargo, J.A.; Alonso, A.; Salamanca, A. Nitrate toxicity to aquatic animals: A review with new data for freshwater invertebrates. Chemosphere 2005, 58, 1255–1267. [Google Scholar] [CrossRef]
- Cheng, S.-Y.; Tsai, S.-J.; Chen, J.-C. Accumulation of nitrate in the tissues of Penaeus monodon following elevated ambient nitrate exposure after different time periods. Aquat. Toxicol. 2002, 56, 133–146. [Google Scholar] [CrossRef]
- Camargo, J.A.; Alonso, A. Ecological and toxicological effects of inorganic nitrogen pollutionin aquatic ecosystems: A global assessment. Environ. Int. 2006, 32, 831–849. [Google Scholar] [CrossRef]
- Schindler, D.W.; Hecky, R.E.; Findlay, D.L.; Stainton, M.P.; Parker, B.R.; Paterson, M.J. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, D. The direct contribution of fish to lake phosphorus cycles. Ecol. Freshw. Fish 2006, 15, 86–95. [Google Scholar] [CrossRef]
- Gorman, M.W.; Zimmer, K.D.; Herwig, B.R.; Hanson, M.A.; Wright, R.G.; Vaughn, S.R.; Younk, J.A. Relative importance of phosphorus, fish biomass, and watershed land use as drivers of phytoplankton abundance in shallow lakes. Sci. Total Environ. 2014, 466–467, 849–855. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Watercourse Name | Region | Geographical Coordinates | Surface Area of Catchment (km2) | Land Use and Characteristics of Catchment 1 |
|---|---|---|---|---|
| Płonia (Plo) | NW PL | N 53.132223 E 15.133033 N 53.122824 E 15.159683 | 174.59 143.59 | A—54%, FO—30%, M—8%, U—5%, W—2%, MA—1% |
| Myśla (Mys) | NW PL | N 52.999399 E 14.977928 N 52.997431 E 15.043580 | 143.16 111.07 | A—68%, FO—22%, M—7%, U—1%, W—1%, MA—1% |
| Tywa (Tyw) | NW PL | N 53.226618 E 14.488537 N 53.230132 E 14.478140 | 270.23 274.25 | A—57%, FO—28%, M—7%, U—4%, W—3%, MA—1% |
| Rurzyca (Rur) | NW PL | N 52.976910 E 14.543279 N 52.966904 E 14.594320 | 83.03 68.41 | A—58%, FO—24%, M—12%, U—3%, MA—2%, W—1% |
| Wardynka (War) | NW PL | N 53.160318 E 15.621905 N 53.155871 E 15.619340 | 25.16 25.93 | FO—51%, A—36%, M—13% |
| Kanał Habdziński (KHb) | CE PL | N 52.109913 E 21.159885 N 52.079362 E 21.174069 | 21.57 11.03 | A—56%, M—16%, U—16%, FO—7%, MA—5% |
| Zielona (Zie) | CE PL | N 51.972162 E 21.044578 N 51.950092 E 21.014168 | 26.72 19.37 | A—59%, M—19%, FO—13%, U—7%, O—2% |
| Czarna Cedron (CCd) | CE PL | N 51.973815 E 21.217970 N 51.984218 E 21.222136 | 69.48 73.80 | O—34%, FO—32%, A—18%, U—13%, M—3% |
| Kraska (Kra) | CE PL | N 51.803904 E 20.879642 N 51.805846 E 20.886593 | 27.14 27.50 | O—44%, A—30%, FO—14%, M—8%, U—4% |
| Molnica (Mln) | CE PL | N 51.854814 E 20.810983 N 51.857631 E 20.816710 | 13.25 13.79 | O—68%, A—19%, FO—13% |
| Feature | CCd | KHb | Kra | Mln | Mys | Plo | Rur | Tyw | War | Zie | Average | SD | CV |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Flow velocity [cm/s] | 0.11 a | 0.07 a | 0.12 abc | 0.07 c | 0.16 abc | 0.37 b | 0.33 b | 0.39 b | 0.79 b | 0.14 abc | 0.24 | 0.27 | 112.7 |
| Discharge [cm3/s] | 0.18 ab | 0.21 a | 0.05 b | 0.01 b | 0.06 b | 1.82 b | 0.55 a | 0.93 a | 0.27 ab | 0.06 b | 0.47 | 0.89 | 186.7 |
| Width [m] | 4.81 a | 5.45 a | 2.25 b | 1.10 b | 1.71 b | 7.57 a | 3.88 abc | 3.92 abc | 2.55 ab | 2.49 ab | 3.89 | 2.20 | 56.4 |
| Depth [m] | 0.47 a | 0.41 ab | 0.21 b | 0.12 b | 0.15 b | 0.70 a | 0.38 ab | 0.46 a | 0.12 b | 0.16 b | 0.35 | 0.24 | 67.1 |
| Temp [°C] | 18.40 a | 17.16 a | 16.89 a | 20.39 a | 16.89 a | 16.54 a | 16.49 a | 16.47 a | 14.50 a | 17.97 a | 17.13 | 2.53 | 14.8 |
| pH | 7.37 a | 7.19 a | 7.70 b | 7.40 ab | 7.63 b | 7.74 b | 7.41 ab | 7.75 b | 7.71 b | 7.50 ab | 7.54 | 0.27 | 3.6 |
| EC [mS/cm] | 492 a | 569 a | 843 b | 691 b | 784 b | 605 ac | 705 bc | 665 bc | 577 abc | 565 ac | 650 | 142.4 | 21.9 |
| O2 [mg/dm3] | 5.47 abc | 4.72 abc | 6.40 abc | 7.93 b | 4.54 abc | 5.66 abc | 3.85 c | 6.18 abc | 6.41 abc | 6.46 abc | 5.62 | 2.06 | 36.5 |
| N-NO3 [mg/dm3] | 3.32 a | 3.88 a | 8.63 b | 13.08 c | 3.88 a | 3.16 a | 3.67 a | 3.95 a | 4.05 a | 7.19 b | 5.09 | 4.58 | 90.0 |
| N-NH4 [mg/dm3] | 0.14 abc | 0.32 abc | 0.17 abc | 0.14 abc | 0.43 abc | 0.38 abc | 1.43 b | 0.36 abc | 0.10 c | 0.12 abc | 0.38 | 0.56 | 149.8 |
| P-PO4 [mg/dm3] | 0.61 a | 0.73 ab | 0.84 ab | 1.13 ab | 0.94 ab | 1.28 b | 1.46 b | 0.84 ab | 0.74 ab | 0.86 ab | 0.94 | 0.54 | 57.8 |
| Variable | All Fish | Rheophilous Fish | Limnephilous Fish | Euryoecious Fish | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Stepwise Analysis | Corr Analysis | Stepwise Analysis | Corr Analysis | Stepwise Analysis | Corr Analysis | Stepwise Analysis | Corr Analysis | |||||||||||||
| t Value | Pr (>|t|) | Sign. Code | R | Sign. Code | t Value | Pr (>|t|) | Sign. Code | R | Sign. Code | t Value | Pr (>|t|) | Sign. Code | R | Sign. Code | t Value | Pr (>|t|) | Sign. Code | R | Sign. Code | |
| 99.54 | <0.00 | **** | 11.97 | <0.00 | **** | 24.97 | <0.00 | **** | 4.46 | <0.00 | **** | |||||||||
| Hydrometrical features of water | ||||||||||||||||||||
| Flow velocity [cm/s] | 0.00 | 0.00 | 0.00 | −0.09 | - | 2.10 | 0.04 | ** | 0.03 | - | 0.00 | 0.00 | 0.00 | −0.17 | - | 0.00 | 0.00 | 0.00 | 0.10 | - |
| Discharge [cm3/s] | 0.00 | 0.00 | 0.00 | −0.19 | * | 0.00 | 0.00 | 0.00 | −0.27 | ** | −1.84 | 0.07 | * | −0.27 | *** | −1.45 | 0.15 | - | −0.13 | - |
| Depth [m] | −5.71 | <0.00 | **** | −0.33 | **** | 0.00 | 0.00 | 0.00 | −0.49 | **** | −3.09 | <0.00 | *** | −0.37 | **** | 2.36 | 0.02 | ** | −0.23 | - |
| Width [m] | 0.00 | 0.00 | 0.00 | −0.33 | **** | 6.52 | <0.00 | **** | −0.51 | **** | 0.00 | 0.00 | 0.00 | −0.36 | **** | 0.00 | 0.00 | 0.00 | −0.23 | - |
| Chemical and physical features of water | ||||||||||||||||||||
| Temperature [°C] | 0.00 | 0.00 | 0.00 | −0.03 | - | 0.00 | 0.00 | 0.00 | 0.16 | - | 0.00 | 0.00 | 0.00 | −0.03 | - | 0.00 | 0.00 | 0.00 | 0.20 | - |
| pH | 0.00 | 0.00 | 0.00 | −0.01 | - | −1.90 | 0.06 | * | −0.13 | - | 0.00 | 0.00 | 0.00 | −0.01 | - | −2.38 | 0.02 | ** | −0.07 | - |
| EC [mS/cm] | 0.00 | 0.00 | 0.00 | 0.23 | ** | 0.00 | 0.00 | 0.00 | 0.20 | - | 1.61 | 0.11 | - | 0.15 | - | 2.82 | 0.01 | *** | 0.03 | - |
| O2 [mg/dm3] | 1.52 | 0.13 | - | −0.06 | - | −2.78 | 0.01 | *** | 0.04 | - | 0.00 | 0.00 | 0.00 | 0.12 | - | −3.46 | <0.00 | *** | 0.44 | *** |
| N-NO3 [mg/dm3] | 2.21 | 0.03 | ** | 0.01 | - | 0.00 | 0.00 | 0.00 | 0.01 | - | 0.00 | 0.00 | 0.00 | −0.09 | - | −1.92 | 0.06 | * | −0.09 | - |
| P-PO4 [mg/dm3] | −3.08 | 0.003 | *** | −0.13 | - | 0.00 | 0.00 | 0.00 | −0.04 | - | −2.19 | 0.03 | ** | −0.20 | * | 1.61 | 0.12 | - | 0.02 | - |
| N-NH4 [mg/dm3] | 0.00 | 0.00 | 0.00 | 0.08 | - | 0.00 | 0.00 | 0.00 | 0.00 | - | −1.69 | 0.10 | * | −0.14 | - | 0.00 | 0.00 | 0.00 | −0.04 | - |
| Streambed substrate composition | ||||||||||||||||||||
| Kd | 0.00 | 0.00 | 0.00 | 0.06 | - | 0.00 | 0.00 | 0.00 | 0.29 | - | 0.00 | 0.00 | 0.00 | −0.13 | - | 0.00 | 0.00 | 0.00 | −0.02 | - |
| Km | 0.00 | 0.00 | 0.00 | −0.07 | - | 0.00 | 0.00 | 0.00 | 0.27 | - | 0.00 | 0.00 | 0.00 | −0.23 | - | 0.00 | 0.00 | 0.00 | −0.17 | - |
| Zd | 0.00 | 0.00 | 0.00 | −0.26 | - | 0.00 | 0.00 | 0.00 | −0.24 | - | 0.00 | 0.00 | 0.00 | −0.34 | ** | 0.00 | 0.00 | 0.00 | 0.34 | - |
| Zm | 0.00 | 0.00 | 0.00 | −0.04 | - | 0.00 | 0.00 | 0.00 | −0.03 | - | 0.00 | 0.00 | 0.00 | −0.11 | - | 0.00 | 0.00 | 0.00 | 0.04 | - |
| P | 0.00 | 0.00 | 0.00 | 0.24 | ** | 0.00 | 0.00 | 0.00 | 0.32 | ** | 0.00 | 0.00 | 0.00 | 0.23 | ** | 0.00 | 0.00 | 0.00 | 0.18 | - |
| M | 0.00 | 0.00 | 0.00 | −0.08 | - | 0.00 | 0.00 | 0.00 | 0.16 | - | 0.00 | 0.00 | 0.00 | −0.04 | - | 0.00 | 0.00 | 0.00 | −0.20 | - |
| Macrophytes | ||||||||||||||||||||
| No of plant sp. | 0.00 | 0.00 | 0.00 | 0.10 | - | 0.00 | 0.00 | 0.00 | −0.05 | - | 0.00 | 0.00 | 0.00 | 0.29 | *** | 0.00 | 0.00 | 0.00 | −0.08 | - |
| Vegetation coverage (%) | 0.00 | 0.00 | 0.00 | 0.15 | - | 0.00 | 0.00 | 0.00 | 0.24 | * | 0.00 | 0.00 | 0.00 | 0.26 | ** | 0.00 | 0.00 | 0.00 | −0.28 | * |
| Axis | Eigenvalue | % | p |
|---|---|---|---|
| 1 | 0.061 | 46.26 | 0.097 |
| 2 | 0.050 | 37.81 | 0.022 |
| 3 | 0.021 | 15.93 | 0.058 |
| Axis | Eigenvalue | % | p |
|---|---|---|---|
| 1 | 0.012 | 30.15 | 0.805 |
| 2 | 0.008 | 19.72 | 0.860 |
| 3 | 0.006 | 15.52 | 0.707 |
| 4 | 0.004 | 10.46 | 0.829 |
| KHb | Kra | Mln | Mys | Pln | Rur | Tyw | War | Zie | |
|---|---|---|---|---|---|---|---|---|---|
| CCd | 0.389 | 0.350 | 0.250 | 0.400 | 0.526 | 0.353 | 0.321 | 0.357 | 0.316 |
| KHb | 0.529 | 0.462 | 0.421 | 0.333 | 0.294 | 0.286 | 0.385 | 0.500 | |
| Kra | 0.500 | 0.611 | 0.429 | 0.500 | 0.462 | 0.333 | 0.625 | ||
| Mln | 0.294 | 0.278 | 0.308 | 0.192 | 0.444 | 0.462 | |||
| Mys | 0.476 | 0.316 | 0.393 | 0.235 | 0.350 | ||||
| Pln | 0.368 | 0.379 | 0.222 | 0.400 | |||||
| Rur | 0.259 | 0.455 | 0.571 | ||||||
| Tyw | 0.154 | 0.333 |
| Code | Taxa_S | Simpson_1-D | Shannon_H | Evenness_e^H/S |
|---|---|---|---|---|
| CCd | 13 | 0.837 | 2.114 | 0.637 |
| KHb * | 12 | 0.761 | 1.844 | 0.527 |
| Kra | 14 | 0.850 | 2.135 | 0.604 |
| Mln * | 7 | 0.715 | 1.437 | 0.601 |
| Mys * | 15 | 0.831 | 2.229 | 0.619 |
| Pln | 16 | 0.873 | 2.302 | 0.625 |
| Rur * | 10 | 0.794 | 1.856 | 0.639 |
| Tyw * | 24 | 0.906 | 2.619 | 0.572 |
| War | 6 | 0.600 | 1.215 | 0.562 |
| Zie * | 12 | 0.791 | 1.853 | 0.531 |
| Axis | Eigenvalue | % | p |
|---|---|---|---|
| 1 | 0.010 | 97.95 | 0.030 |
| 2 | 0.000 | 1.63 | 0.134 |
| 3 | 4.310 | 0.421 | 0.029 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brysiewicz, A.; Czerniejewski, P.; Dąbrowski, J.; Formicki, K.; Więcaszek, B. Fish Diversity and Abundance Patterns in Small Watercourses of the Central European Plain Ecoregion in Relation to Environmental Factors. Water 2022, 14, 2697. https://doi.org/10.3390/w14172697
Brysiewicz A, Czerniejewski P, Dąbrowski J, Formicki K, Więcaszek B. Fish Diversity and Abundance Patterns in Small Watercourses of the Central European Plain Ecoregion in Relation to Environmental Factors. Water. 2022; 14(17):2697. https://doi.org/10.3390/w14172697
Chicago/Turabian StyleBrysiewicz, Adam, Przemysław Czerniejewski, Jarosław Dąbrowski, Krzysztof Formicki, and Beata Więcaszek. 2022. "Fish Diversity and Abundance Patterns in Small Watercourses of the Central European Plain Ecoregion in Relation to Environmental Factors" Water 14, no. 17: 2697. https://doi.org/10.3390/w14172697