
Inorganic Nitrogen Uptake Characteristics of Three Typical Bloom-Forming Algae in the East China Sea
by
 ,
,
Guangmao Ding
1,†, 
Huorong Chen
1,†,
Haifeng Gu
2,
Youquan Zhang
1,
Rongmao Li
1 and
Shufeng Zhang
3,* 1
Fishery Resources Monitoring Center of Fujian Province, Fuzhou 350003, China
2
Third Institute of Oceanography, Ministry of Natural Resources, Xiamen 361005, China
3
State Key Laboratory of Marine Environmental Science, College of Ocean and Earth Sciences, Xiamen University, Xiamen 361005, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Water 2022, 14(16), 2580; https://doi.org/10.3390/w14162580
Submission received: 29 July 2022
/
Revised: 11 August 2022
/
Accepted: 15 August 2022
/
Published: 21 August 2022
(This article belongs to the Special Issue Harmful Algal Blooms and the Mechanism of Hypoxia in Coastal Waters)
Abstract
:Inorganic nitrogen (N) is an important element for eutrophication and harmful algal bloom (HAB) formation. However, the roles of inorganic N in HAB outbreaks are still unclear. Here, we compared the affinities and abilities for inorganic N uptake and assimilation among three typical bloom-forming algae in the East China Sea (ECS), Skeletonema costatum, Prorocentrum donghaiense and Alexandrium pacificum by investigating the uptake and enzymatic (nitrate reductase (NR) and glutamine synthetase (GS) kinetics for nitrate and ammonia. The Ks of nitrate and ammonium in S. costatum was lower than those in P. donghaiense and A. pacificum. The NR activity of S. costatum and P. donghaiense exhibited a positive relationship with the nitrate concentration, and NR activity of S. costatum was nearly 4-fold higher than that of P. donghaiense at high nitrate concentration. However, the NR activity of A. pacificum could not be detected. The GS activity of three species decreased with the increase of ammonium concentrations, and the highest GS activity was detected in A. pacificum. S. costatum presented the highest affinity for nitrate and ammonium, followed by P. donghaiense and A. pacificum. Moreover, P. donghaiense exhibited the highest affinity for intracellular ammonium. Our results characterized the differences in inorganic nitrogen uptake among the three typical bloom-forming algae, which may contribute to the formation of blooms in the coastal waters of the ECS.
1. Introduction
Inorganic nitrogen (N) is a category of N nutrient for phytoplankton growth, which drives the eutrophication and the formation of harmful algal blooms (HABs) in coastal waters [1,2]. Competition for inorganic N among different phytoplankton species has been demonstrated to be important in the formation and succession of HABs [1,2,3]. Hence, the relationship between inorganic N and phytoplankton has become the focus of attention [1,2,3]. Inorganic N assimilated by phytoplankton is an important part of the N cycle in nature [1,2]. Differences in the uptake kinetics for nitrate and ammonium among algal species have been reported and may determine the relative growth rates of algae and therefore influence competitive relationships [4,5,6,7,8]. Ammonium is the preferred inorganic N source for Karenia mikimotoi [7], while Gymnodinium catenatum has the maximum uptake rate of nitrate than that of ammonium [8]. Phytoplankton with low half-saturation constant (Ks) may be favored under inorganic N-stressed conditions as they can efficiently take up inorganic N, while species with a high maximum uptake rate (Vmax) may gain an advantage under abundant inorganic N conditions [8,9]. Likewise, different affinities for ammonium versus nitrate as a N source may favor some species [7,8,9,10]. Some taxonomic groups, such as K. mikimotoi [7,10], may also be affected by the nature of the inorganic N. For instance, nitrate impulses usually result in diatom blooms in the coastal upwelling system [10,11]. Phytoplankton preferentially use ammonium, but its utilization can also inhibit the assimilation of nitrate [12,13]. However, the utilization mechanism of phytoplankton for nitrate and ammonium are still unclear.
There are three fundamental stages to assimilating nitrate into phytoplankton cells: (a) transportation across the cytoplasm membrane using a special carrier; (b) reduction of nitrate to ammonium catalyzed by the sequential action of nitrate reductase (NR) and nitrite reductase (NiR); and (c) incorporation of ammonium into glutamate to produce glutamine via one of two pathways: glutamate dehydrogenase (GDH) catalyzes the uptake of NH4+ in glutamate by 2-oxoglutarate (2-OG), or alternatively, in the GS/GOGAT pathway, NH4+ is incorporated in glutamine by glutamine synthetase (GS) and converted with 2-OG into glutamate via glutamate synthase (GOGAT) [14]. Research on assimilatory NR has been carried out for many years, three eukaryotic assimilatory NR forms have been identified and characterized, two of which occur in eukaryotic algae and higher plants (EC 1.6.6.1, specific for NADH and EC 1.6.6.2, using NADH or NADPH) and one of which is found only in fungi (EC 1.6.6.3, specific for NADPH). NR contains three groups of prostheses, FAD, heme-iron and a cofactor of molybdopterin which participate sequentially in reducing nitrate to nitrite [15]. NR’s physiological role in reducing nitrates is fundamental to algae, vascular plants, fungi and various bacteria [15,16]. NR meets multiple criteria for a rate-limiting enzyme [17]; however, appropriate experiments to test this are definitely lacking. Some studies have suggested that nitrate incorporation is restricted by nitrate uptake [18] or by enzymatic steps downstream of NR, such as NiR [17]. While GS might be another checkpoint [19]. The GS enzyme in the GS/GOSAT pathway of ammonium assimilation has been reported in various algal and cyanobacteria as well as in animal cells [19,20]. Almost without exception, GS activity shows an extremely low Ks for ammonium [21].
Skeletonema costatum, Prorocentrum donghaiense and Alexandrium pacificum are three key bloom species in the coast of the East China Sea (ECS), which have caused extensive and recurrent blooms in the past decades [22]. In situ investigations have shown that blooms in the coast of ECS exhibited the succession pattern, diatom blooms formed by S. costatum or other diatom species usually occurred in the early spring (March) of each year, then followed by dinoflagellate blooms formed by P. donghaiense and/or A. pacifium (as A. catenella) [22,23]. Considerable data from the field investigations and mesocosm experiments have demonstrated that eutrophication, especially the increasing input of N from the Changjiang River is the major reason resulting in the occurrence of blooms in ECS [24,25,26]. In early spring, the concentrations of NO3-N in the coastal waters of ECS were high due to the supplements from the Changjiang River together with a strongly vertical mixture of seawater in winter, which provided sufficient nutrients for phytoplankton species, and results in the change of N:P ratio [27]. It has been postulated that “excess nitrogen” resulted in a high N:P ratio and phosphate limitation in ECS, which promotes the formation of large-scale dinoflagellate blooms in spring, and the shift of major causative species from diatoms to dinoflagellates [27]. However, little is known about their inorganic N requirements and preferences leading to large-scale blooms.
The present study investigated the uptake and enzymatic kinetics of nitrate and ammonia among three typical bloom-forming algae in the ECS, S. costatum, P. donghaiense and A. pacificum, and compared the affinity for inorganic N and abilities of inorganic N uptake and assimilation. The goal of this study is to reveal the inorganic N uptake characteristics of the three typical bloom species in the coastal waters of the ECS.
2. Materials and Methods
2.1. Algal Strains and Culture Conditions
S. costatum, P. donghaiense and A. pacificum were isolated from the coastal water of ECS, China in May 2002. Fifty milliliters surface bloom waters (0.5 m depth) were collected, and single cells of each strains were picked up under a microscope (Axio Imager A2, Carl Zeiss, Germany) by serial dilution. The identified algal strains were maintained in the Culture Collection Center of Marine Algae, Xiamen University, China. Unialgal isolates are routinely maintained in f/2 medium with (for S. costatum) or without SiO32− (for P. donghaiense and A. pacificum) with sterilized seawater collected from the Taiwan Strait (low N; station: 24.154095 N, 118.605045 E; salinity 33‰) at 20 °C under a 12:12 h light:dark photoperiod at a light intensity of approximately 100 μmol m−2 s−1 provided by fluorescent lamps. All glassware used in this study were washed with 30% HCl to remove ammonium and trace metals, and thoroughly rinsed with Milli Q water before the experiment [8].
The exponential growing cells of the three algae species according to the results of the cell count under a microscope (Axio Imager A2, Carl Zeiss, Jena, Germany; Objective lense 40×) were inoculated into new f/2 medium with low N, respectively, and 3.0 µM of NH4Cl or NaNO3 was added every day to acclimate the algae species for about 5 generations. When the inorganic N in the culture medium was exhausted, cells were cultured for 2 days to deplete the intracellular N and used for experiments.
2.2. Time-Course Experiments
To determine the optimal time for nutrient uptake kinetic experiments, the time course experiments were conducted [28,29]. At the exponential phase, A. pacificum and P. donghaiense cells were harvested by centrifugation at 8000× g for 15 min and S. costatum at 10,000× g for 20 min at 20 °C, cell pellets were washed with sterilized seawater collected from the Taiwan Strait (low N). The cell pellets were resuspended in 2 L seawater (for S. costatum) or without SiO32− (for P. donghaiense and A. pacificum) with f/2 medium containing no N source, either 8.0 µM nitrate (NaNO3) or 4.0 µM ammonium (NH4Cl) was added. The concentrations of nitrate and ammonium were analyzed at 0, 10, 20, 30, 40, 60, 90 and 120 min. The concentrations of nitrate and ammonium were determined after filtering through a membrane filter (0.45 µm) and measured using continuous flow analysis (CFA-SAN Plus; Skalar Analytik, Erkelenz, Germany) by the method of Wood et al. [30] and Solórzano [31], respectively. Three 1-mL culture media samples were collected daily at 10:00 am for cell counting, and cell number was manually counted with a light microscope, the concentrations of nitrate and ammonium, and the enzyme activity of nitrate reductase (NR) and glutamine synthetase (GS) were also analyzed daily.
2.3. Nutrient Uptake Experiments
To obtain nitrate uptake rates, the acclimated S. costatum, P. donghaiense and A. pacificum were inoculated into f/2 medium without N, NaNO3 was added to the final concentration of 1.0, 2.0, 4.0, 6.0, 8.0 and 10.0 µM with the initial cell density about 1 × 104 cell mL−1, respectively. For NH4+ uptake rates, NH4Cl was added to the final concentrations of 0.25, 0.5, 1.0, 1.5, 2.0 and 3.0 µM, respectively. Concentrations of nitrate and ammonium were analyzed at the end of the incubation time which has been determined in the time-course experiments. All experiments were conducted between 10:00 and 12:00 am because no cell division occurred according to the results of the cell count under a microscope.
2.4. Extraction of NR and GS
About 200 mL of A. pacificum and P. donghaiense cultures were harvested at the exponential growth phase with the cell density about 1 × 105 cell mL−1 by centrifugation at 8000× g for 15 min at 4 °C, and about 400 mL of S. costatum culture was harvested at the exponential growth phase with cell density of about 4 × 105 cell mL−1 by filtering onto a polycarbonate membrane (3.0 μm, Millipore). The enriched cells of A. pacificum, P. donghaiense and S. costatum from each culture were washed twice with sterilized low-N seawater. Then, cells were centrifuged at 12,000 rpm for 2 min at 4 °C to remove the supernatant, and cells were immediately frozen at −80 °C for preparation of crude enzyme.
The crude NR enzyme was prepared with 200.0 mM phosphate buffer (pH 7.9) containing 0.03% (w/v) dithiothreitol (DTT), 0.3% (w/v) polyvinyl pyrrolidone (PVP), 0.1% (v/v) Triton X-100, 5 mM ethylene-diaminetetraacetic acid (EDTA) and 3% (w/v) bovine serum albumin (BSA), with the help of a sonicator (Fisher, Waltham, MA, USA) under 280 W for 2 s, 2 s interval, and 40 cycles. The cell debris were removed by centrifugation at 14,000× g for 30 min and the suspension was collected for analysis of enzyme activity. The crude GS enzyme were extracted using 50.0 mM Tris-HCl (pH 7.5) containing 2.0 mM DTE, 1 mM EDTA and 2.5 mM MgCl2 with the help of a sonicator (Fisher, Waltham, MA, USA) under 280 W for 2 s, 2 s interval, and 40 cycles. The cell debris were removed by centrifugation at 14,000× g for 30 min. To remove pigments, a solution of 100.0 mM streptomycin sulphate at pH 7.0 was added to the supernatant (0.1 mL per 1 mL). After stirring for 15 min at 4 °C, the suspension was centrifuged at 14,000× g for 30 min.
2.5. Analysis of NR and GS Activities
NR activity was determined using the method of Lomas and Gelibert [11]. Briefly, The crude enzyme extract (100 μL), flavin adenine dinucleotide (100 μL; FAD, 2.0 mM), nicotinamide adenine dinucleotide (100 μL; NADH, 0.5 mM) and 200 mM phosphate buffer (350 μL; pH 7.9) were added to a tube, and the reaction was initiated by adding 250 μL 200 mM KNO3. A second tube, the same buffer was added as control, and the third without crude enzyme extract as a reagent blank. All tubes were incubated at 23 °C for 45 min, then 2.0 mL of 550 mM zinc acetate was added to the first and the third tube to stop the reaction. Zinc acetate was added to the second tube immediately after the reaction started. The tubes were centrifuged and excess NADH was oxidized with the addition of 20 μL 125.0 µM phenazine methosulphate (PMS). Nitrite produced was measured by colorimetry with sulfanilamide and N-(1-napthyl)-ethylenediamine solutions. A unit of NR activity was calculated as the activity of nitrite production per minute.
GS activity was determined using the methods of Takabayashi et al. [32]. Briefly, to one tube, the crude enzyme extract (100 μL), 1.0 M imidazole-HCl buffer (960 μL), 0.1 mM glutamine (600 μL; pH 7.3), 0.01 mM MnCl2 (60 μL), 0.01 mM ADP (80 μL; pH 7.3), 1.0 M K-arsenate (40 μL) and 2.0 M hydroxylamine (60 μL) were added. To the second tube, the same solution was added as control. The third tube contained no crude enzyme extract as a reagent blank. The reactions were conducted at 37 °C for 30 min and stopped by the addition of 2.0 mL mixture (4.0 mL 10% FeCl3, 1.0 mL 24% trichloroacetic acid, 0.5 mL 6.0 M HCl and 6.5 mL Milli Q water). In the second tube, the reaction was stopped immediately when the solution was added. After stopping the reaction, the absorbance was measured at 540 nm. One unit of GS activity was defined as the activity inorganic N produced per minute.
2.6. Data Analyzing
Curve fitting was analyzed using a computerized, iterative non-linear least-squares technique (Kaleidograph) which utilizes the Levenberg-Marquardt algorithm using the software GraphPad [4]. Data were initially made linear and plotted using a double reciprocal Hanes-Woolfe method [29]. The results were directly fitted to the Michaelis-Menten formulation:
where V is the maximum uptake rate (pmol cell−1 h−1 or fmol cell−1 h−1), Ks is the half-saturation constant (µM) for the N substrate and S is the ambient N concentration (µM). All data processing and statistical analyses were conducted using Sigmaplot Statistical Software (version 8.0, SPSS Inc.).
V = Vmax S/(Ks + S)
3. Results
3.1. Reaction Time in N Uptake Kinetics Experiment
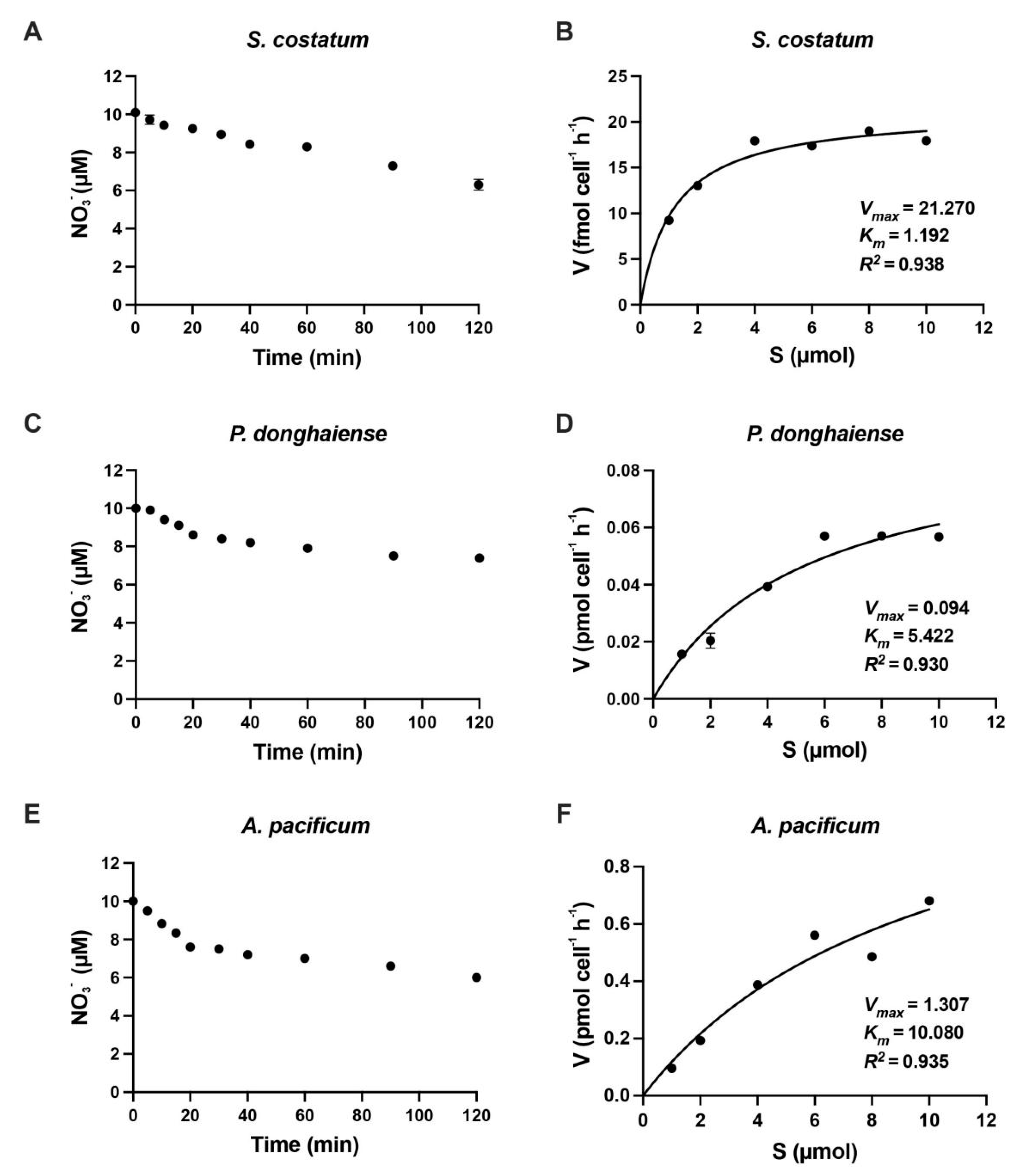
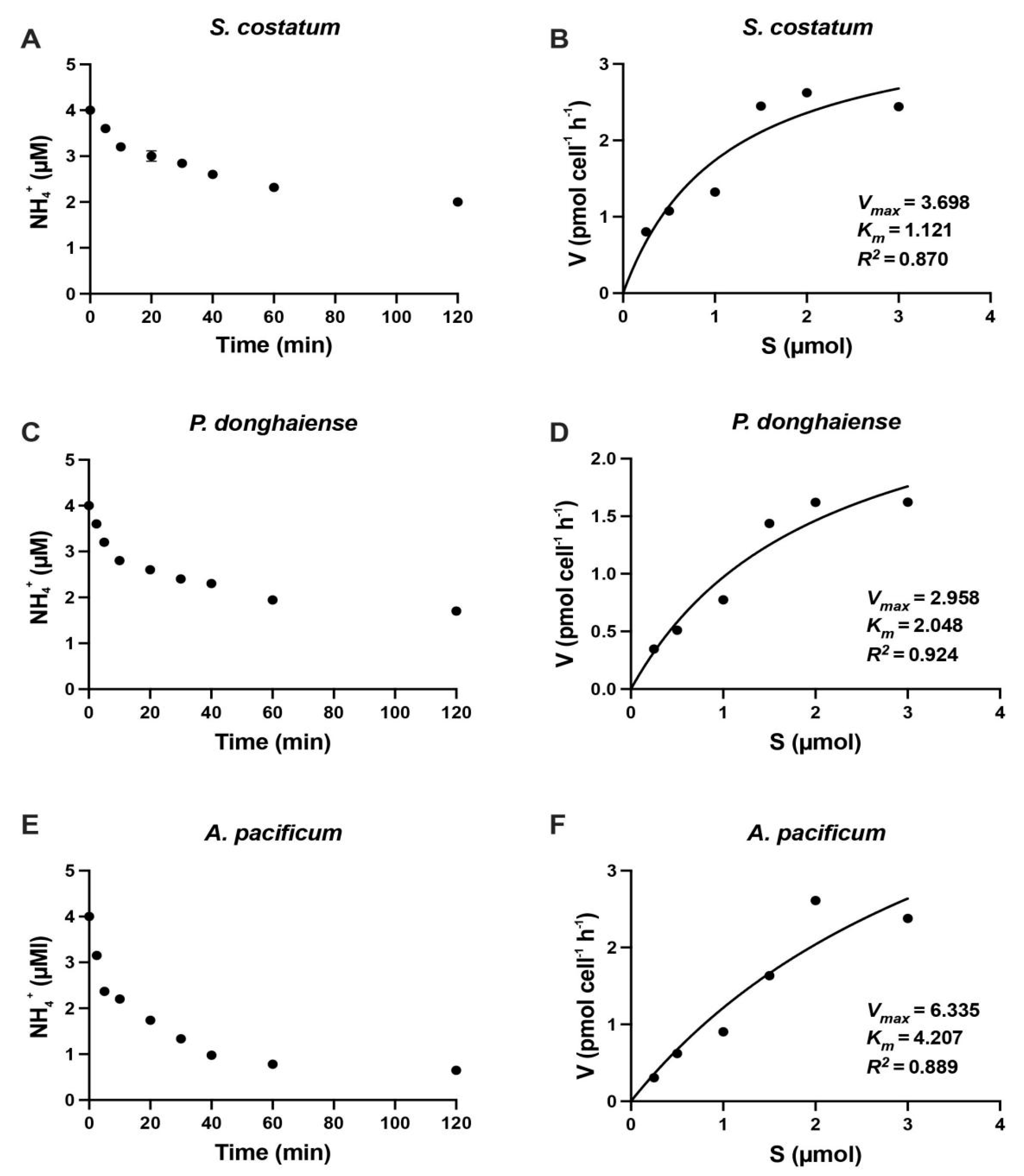
The nitrate and ammonium uptake rates of S. costatum, P. donghaiense and A. pacificum as the functions of the ambient N concentrations are shown in Figure 1 and Figure 2. The uptake rates of nitrate were relatively constant for the first 20 min in all three species: 0.04–0.1 µmol min−1 for S. costatum, 0.06–0.07 µmol min−1 for P. donghaiense and 0.1–0.14 µmol min−1 for A. pacificum, then decreased to 0.02–0.04 µmol min−1, 0.0033–0.004 µM min−1 and 0.02 - 0.03 µmol min−1, respectively (Figure 1 and Figure 2). Similarly, for ammonium, the first 10 min was selected because the uptake rate presented linear relationship with the time.
3.2. Kinetics of Inorganic N Uptake by S. costatum, P. donghaiense and A. pacificum
The data from the short-term uptake experiment fitted well with a Michaelis- Menten curve using N-deficient cells of S. costatum (r2 = 0.97 for nitrate and r2 = 0.93 for ammonium), P. donghaiense (r2 = 0.95 for nitrate and r2 = 0.96 for ammonium) and A. pacificum (r2 = 0.97 for nitrate and r2 = 0.98 for ammonium). The Ks and Vmax obtained from the short-term uptake experiments of nitrate and ammonium are shown in Table 1 and Table 2. The Vmax for nitrate were 0.02, 0.098 and 1.3 pmol cell−1 h−1, and Ks were 1.19, 5.98 and 10.02 µM for S. costatum, P. donghaiense and A. pacificum, respectively, while the Vmax for ammonium were 0.037, 0.296 and 4.21 pmol cell−1 h−1 and the Ks were 1.12, 2.04 and 4.27 µM, respectively for three species.
3.3. NR Kinetics of S. costatum and P. donghaiense
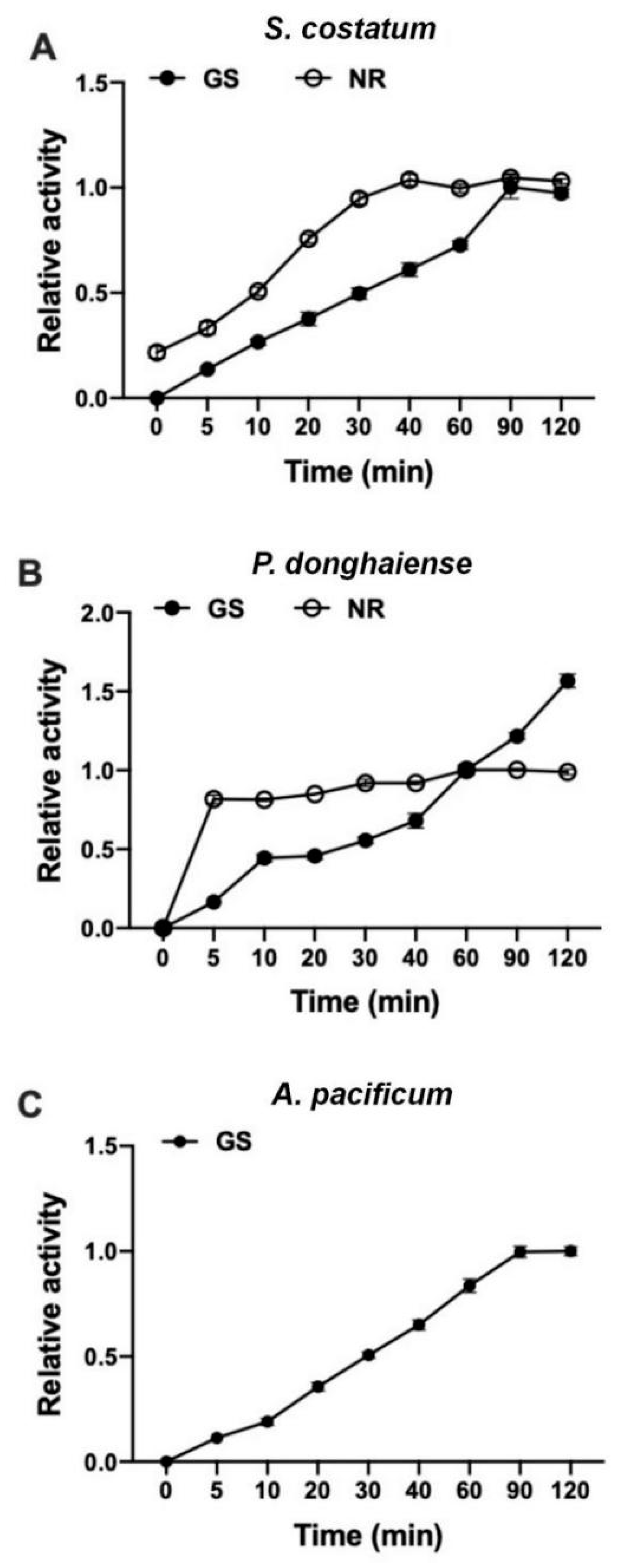
The NR activity increased linearly with the incubation time within the first 40 min for S. costatum but only 10 min for P. donghaiense (Figure 3). Therefore, the first 40 min was selected as the reaction time for S. costatum and 10 min for P. donghaiense. However, NR activity of A. pacificum was not detected.
The kinetics of NR activity were examined at two ranges of NO3− concentrations, 0 to 200 µM and 0 to 60 mM. Ks and Vmax for S. costatum and P. donghaiense at two nitrate concentration ranges are shown in Figure 4 and Table 3. For S. costatum, Ks and V max varied slightly at different nitrate concentration ranges; however, P. donghaiense exhibited one Ks and Vmax value. Ks of P. donghaiense was 2-fold higher than that of S. costatum.
3.4. GS Kinetics of S. costatum, P. donghaiense and A. pacificum
The GS activity increased linearly with the incubation time within the first 40 min for both S. costatum and A. pacificum at the exponential phase. However, for P. donghaiense, the GS activity increased linearly with the incubation time within the first 120 min (Figure 3). Therefore, the first 40 min was selected as the reaction time for S. costatum and A. pacificum and 120 min for P. donghaiense.
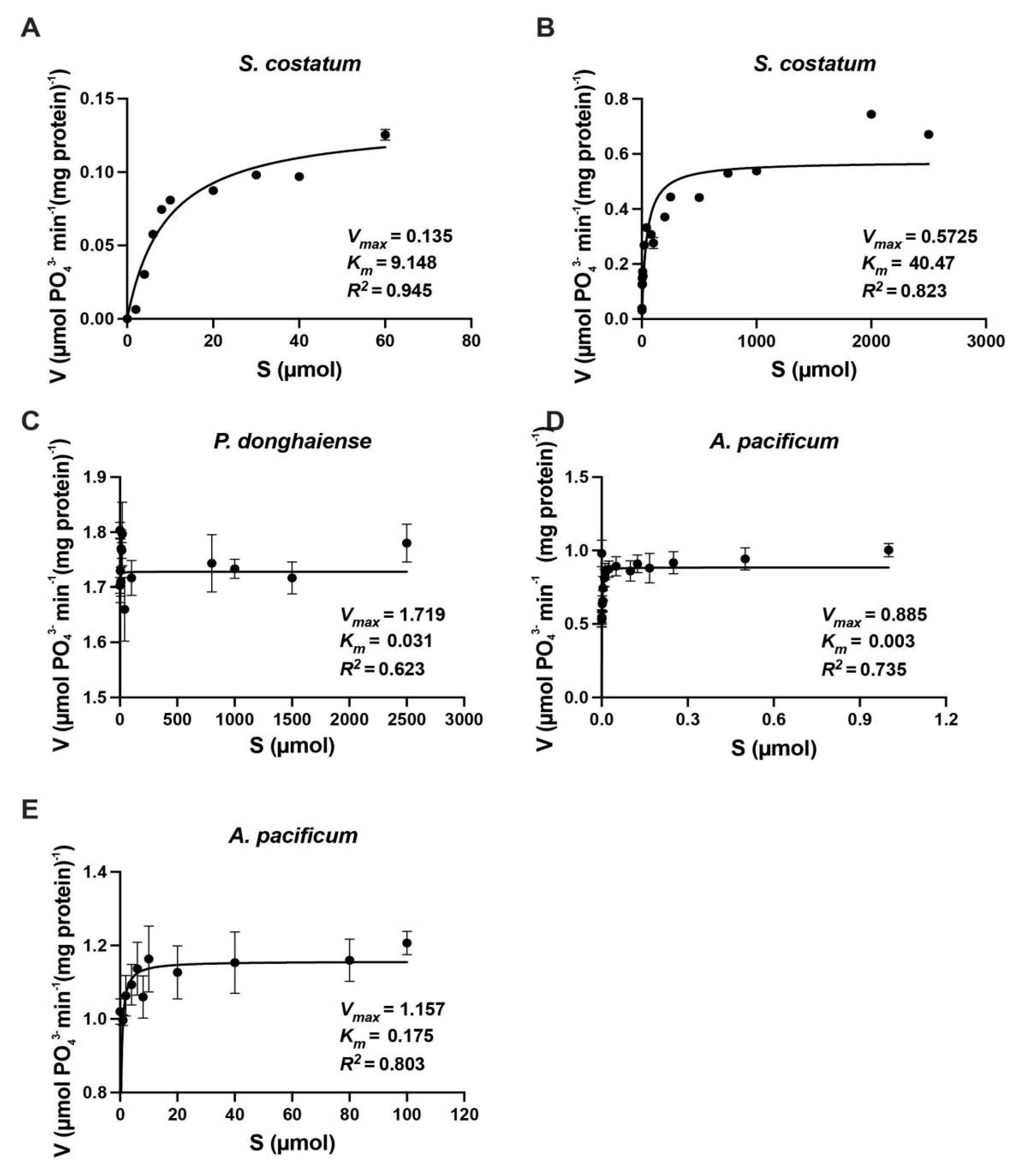
Ks and Vmax of S. costatum, P. donghaiense and A. pacificum at low and high ammonium concentration ranges are shown in Figure 5 and Table 4. Ks and Vmax of S. costatum and A. pacificum at low NH4+ concentration range (0 to 200 µM) were lower than those at high NH4+ concentration range (0 to 3 mM). However, for P. donghaiense, Ks and Vmax were the same at two NH4+ concentration ranges, 0.043 µmol and 0.85 µmol PO43− min−1 (mg protein)−1, respectively.
4. Discussion
4.1. Uptake Kinetics of Inorganic N by S. costatum, P. donghaiense and A. pacificum
The kinetic parameters of N uptake can be used to evaluate the preference for different N substrate at low and high ambient N concentration representative of oligotrophic and eutrophic areas. Ks and Vmax have been used as indicators of marine phytoplankton ability to take up nutrients at low and/or high concentrations, respectively. Ks is regarded as an index of cell affinity to nutrients and ecologically significant in respect of interspecific competitive interactions [39]. While Vmax is usually obtained using nutrient concentrations far higher than those in the natural environment, which will be useful in predicting species response to episodic high nutrient pulses, such as those following an upwelling or discharge event [40]. Previous studies have shown that diatoms exhibit significantly lower Ks values and higher NO3− uptake rates than dinoflagellates [11,39,41]. In the present study, the Ks of nitrate and ammonium in S. costatum was lower than those of P. donghaiense and A. pacificum, indicating that S. costatum possessed higher affinity to inorganic N than P. donghaiense and A. pacificum in the environment with low inorganic N concentrations. It is well known that cell size and shape determine species-specific differences in Ks values to some extent suggesting that the Ks presented a positive relationship with cell size. Cell size of A. pacificum is 17,724.29 × 4358.20 µm3, much larger than P. donghaiense (1798.87 × 511.99.20 µm3) and S. costatum (113.29 × 52.43 µm3) (Table 1), respectively [42]. Cell size not only influences Ks, but also causes an impact on maximum specific nitrogen uptake rates [43]. The cell volume can be addressed by normalizing uptake rates to either biomass or cellular surface area. Among the three bloom species, A. pacificum has higher surface area normalized NO3− and NH4+ uptake rates than P. donghaiense and S. costatum (Table 1 and Table 2). This is consistent with the previous conclusion that Vmax is functionally related to and positively correlated with K s [39,40,43,44].
It has been demonstrated that different affinities for ammonium versus nitrate as a nitrogen source may favor some species [7,8,9], some phytoplankton species can utilize low concentrations of NH4+ more efficiently than equivalent concentrations of NO3− because less energy is required to incorporate ammonia instead of nitrate which requires reduction with the help of the NR [1,3]. In the present study, the Vmax-NH4+ for N-starved cells of three algal species was higher than the Vmax-NO3−, while Ks-NH4+ for N-starved cells of the three species was lower than Ks-NO3−, indicating that three species have higher affinity for NH4+ than for NO3−. Based on our experiments, the preference for inorganic N among the three species followed the order: NH4+ > NO3− when high ambient N conditions were added into N-starved cultures.
With the kinetic data taken as a whole, the patterns of nitrate uptake for the three species are consistent with what would be expected for a two-component nitrate uptake system: S. costatum belongs to high-affinity and low-capacity constitutive component, and A. pacificum and P. donghaiense belong to low-affinity and high-capacity inducible uptake component. This different nutrient utilization strategy among three species may have determined their competitive ability to nutrients in ambient nutrient concentration, i.e., S. costatum present a high competitive ability to nitrogen nutrients compared to other two species.
4.2. Enzymatic Kinetics of Inorganic N by S. costatum, P. donghaiense and A. pacificum
Enzymes involved in the N metabolism have been investigated extensively. Many phytoplankton species were found to have a diel periodicity in cell division, nitrate, ammonium uptake and NR activity [16,45,46], and consequently, species-specific differences in patterns of diel periodicity can lead to different responses. The difference between diatoms and dinoflagellates appears to be in the phasing of peak NR activity. Diatoms consistently have a maximum NR activity after 3 h of illumination [47]. However, dinoflagellates have a much more varied phasing of maximum NR activity ranging from 3 to 9 h after the beginning of the photoperiod [48,49]. The data presented here were collected at 9:00 to 10:00 am, 3 h after the light is turned on. The Ks values for the diatom, S. costatum ranged from 0.082 to 0.29 mM, fall in a wide range of previous studies [38,50,51,52]. The Ks values for P. donghaiense were 2-fold higher of those for S. costatum, and the Vmax-NR rates for S. costatum are nearly 10-fold of those for P. donghaiense, which suggested that both the affinity for intracellular nitrate and the reduction rate of nitrate to nitrite in S. costatum was higher than that in P. donghaiense. However, NR activity of A. pacificum was not detected, which might be due to the low expression or activity of NR enzyme in A. pacificum.
Since the discovery of the enzyme GOGAT, many studies have been devoted to GS activity in different algal species. GS activity is also known to have a diel periodicity by exhibiting the maximum value during the dark period and the minimum during the light period [53,54]. In the present study, samples were collected at 9:00 to 10:00 am after a 3-h photoperiod. The GS exhibited a very low Ks for ammonium compared to the previous studies. Bressler and Ahmed [38] investigated GS activities of 15 marine phytoplankton species and found Km-NH4+ values were quite low, from 1.8 to 8.2 µM. The value of the apparent Michaelis constant for the physiological activity of the purified enzyme for ammonium was 0.05 mM in a green algae, Monovaphidium braunii [53]. In a marine environment, nutrients are always limited for phytoplankton species, and the ability to store nutrients and efficiently utilize N is important. Phytoplankton species have evolved different mechanisms to adapt to the critical conditions limiting their growth. Some species have the ability to absorb ammonium from the environment at an accelerated rate, while other species can utilize and assimilate ammonium through a highly efficient enzyme system which presents a high affinity for this ion [1,40]. In the present study, Ks-GS for NH4+ was investigated at low and high NH4+ levels, Ks-GS of S. costatum and A. pacificum showed a direct increase with the increasing ammonium concentrations, while Ks of P. donghaiense was the lowest and presented only one value at low and high ammonium concentration ranges, indicating P. donghaiense possesses the highest affinity for intracellular ammonium compared to other two species.
In early spring, the coastal water of ECS with strongly vertical mixture in winter provide sufficient N for phytoplankton, S. costatum with low Ks and high Vmax recover rapidly from N-starvation conditions, as well as its high growth rate comparing to dinoflagellates, S. costatum becomes the dominant species. At the end of the bloom of S. costatum, nutrients were consumed, which inhibited cell division and growth of S. costatum. Meanwhile, dinoflagellates began to bloom using their storage nutrients as well as their high affinity to NH4+, high ammonium assimilation rate.
5. Conclusions
Our results characterized the affinities and abilities for inorganic N uptake and assimilation among three typical bloom-forming algae in the ECS, S. costatum, P. donghaiense and A. pacificum by investigating the uptake and enzymatic (NR and GS) kinetics for nitrate and ammonia. S. costatum with low Ks of nitrate and ammonium possessed high affinity to inorganic N in competition with P. donghaiense and A. pacificum under low inorganic N conditions. While A. pacificum possessed higher NO3− uptake rates than P. donghaiense and S. costatum during the starvation phase. A. pacificum has higher surface area normalized NO3− and NH4+ uptake rates than P. donghaiense and S. costatum. The differences in uptake and assimilation strategy for inorganic N among the three species might contribute to the formation of blooms in the coastal of ECS.
Author Contributions
Conceptualization, G.D. and S.Z.; methodology, G.D., H.G., Y.Z., R.L. and H.C.; software, G.D.; validation, G.D.; formal analysis, G.D., H.C., Y.Z., R.L. and H.C.; investigation, G.D., H.G., R.L., H.C. and S.Z.; resources, G.D., R.L. and H.C.; data curation, G.D. and S.Z.; writing—original draft preparation, G.D., H.G. and S.Z.; writing—review and editing, G.D. and S.Z.; visualization, G.D. and S.Z.; supervision, G.D. and S.Z.; project administration, G.D. and S.Z.; funding acquisition, G.D., R.L. and S.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Fujian Marine Economic Development Special Fund Project (FJHJF-L-2022-12), the National Key R&D Program of China (2017YFC1404803) and the Open Fund of CAS Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences (KLMEES201806).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Glibert, P.M.; Berdalet, E.; Burford, M.A.; Pitcher, G.; Zhou, M.J. Global Ecology and Oceanography of Harmful Algal Blooms; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Heisler, J.; Glibert, P.M.; Burkholder, J.M.; Anderson, D.M.; Cochlan, W.; Dennison, W.C.; Dortch, Q.; Gobler, C.J.; Heil, C.A.; Lewitus, A.; et al. Eutrophication and harmful algal blooms: A scientifific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Glibert, P.M.; Burkholder, J.M. Harmful algal blooms and eutrophication: Nutrient sources, composition, and consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- Killberg-Thoreson, L.; Mulholland, M.R.; Heil, C.A.; Sanderson, M.P.; O’Neil, J.M.; Bronk, D.A. Nitrogen uptake kinetics in field populations and cultured strains of Karenia brevis. Harmful Algae 2014, 38, 73–85. [Google Scholar] [CrossRef]
- Cochlan, W.P.; Bronk, D.A. Nitrogen uptake kinetics in the Ross Sea, Antarctica. Deep Sea Res. II 2001, 48, 4127–4153. [Google Scholar] [CrossRef]
- Shi, F.; Wei, X.X.; Feng, J.F.; Zhu, L. Nitrogen uptake kinetics of a diatom and model prediction analysis in different inorganic nitrogen conditions. J. Agro Environ. Sci. 2018, 37, 1833–1841. [Google Scholar]
- Lecorre, P.; Lhelguen, S.; Wafar, M. Nitrogen-source for uptake by Gyrodinium cf. aureolum in a tidal front. Limnol. Oceanogr. 1993, 38, 446–451. [Google Scholar]
- Yamamoto, T.; Oh, S.J.; Kataoka, Y. Growth and uptake kinetics for nitrate, ammonium and phosphate by the toxic dinoflagellate Gymnodinium catenatum isolated from Hiroshima Bay, Japan. Fish. Sci. 2011, 70, 108–115. [Google Scholar] [CrossRef]
- Maguer, J.F.; Helguen, S.L.; Madec, C.; Labry, C.; Corre, P.L. Nitrogen uptake and assimilation kinetics in Alexandrium minutum (Dinophyceae): Effect of N-limited growth rate on nitrate and ammonium interactions. J. Phycol. 2007, 43, 295–303. [Google Scholar] [CrossRef]
- Closset, I.; McNair, H.M.; Brzezinski, M.A.; Krause, J.W.; Thamatrakoln, K.K.; Jones, J.L. Diatom response to alterations in upwelling and nutrient dynamics associated with climate forcing in the California Current System. Limnol. Oceanogr. 2021, 66, 1578–1593. [Google Scholar] [CrossRef]
- Lomas, M.W.; Glibert, P.M. Comparisons of nitrate uptake, storage, and reduction in marine diatoms and flagellates. J. Phycol. 2000, 36, 903–913. [Google Scholar] [CrossRef]
- Dortch, Q. The interaction between ammonium and nitrate uptake in phytoplankton. Mar. Ecol. Prog. 1990, 61, 183–201. [Google Scholar] [CrossRef]
- Glibert, P.M.; Wilkerson, F.P.; Dugdale, R.C.; Raven, J.A.; Dupont, C.L.; Leavitt, P.R.; Parker, A.E.; Burkholder, J.M.; Kana, T.M. Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions. Limnol. Oceanogr. 2016, 61, 165–197. [Google Scholar] [CrossRef]
- Capone, D.G. The marine microbial nitrogen cycle. In Microbial Ecology of the Oceans; David, L.K., Ed.; John Wiley and Sons INC.: New York, NY, USA, 2008; pp. 1–6. [Google Scholar]
- Crawford, N.M. Nitrate: Nutrient and signal for plant growth. Plant Cell 1995, 7, 859–868. [Google Scholar]
- Granbom, M.; Chow, F.; Lopes, P.F.; de Oliveira, M.C.; Colepicolo, P.; de Paula, E.J.; Pedersén, M. Characterisation of nitrate reductase in the marine macroalga Kappaphycus alvarezii (Rhodophyta). Aquat. Bot. 2004, 78, 295–305. [Google Scholar] [CrossRef]
- Bender, D.; Schwarz, G. Nitrite-dependent nitric oxide synthesis by molybdenum enzymes. FEBS Lett. 2018, 592, 797. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Yan, C.; Zhang, X.Y.; Peng, W.; Geng, D.G.; Zhao, S.M.; Zhang, L.M.; Sun, Y.R. Isolation and characterization of a nitrate reductase deficient mutant of Chlorella ellipsoidea (Chlorophyta). J. Appl. Phycol. 2005, 17, 281–286. [Google Scholar] [CrossRef]
- Castro-Rodríguez, V.; García-Gutiérrez, A.; Canales, J.; Avila, C.; Kirby, E.G.; Cánovas, F.M. The glutamine synthetase gene family in populus. BMC Plant Biol. 2011, 11, 119. [Google Scholar] [CrossRef]
- Alaoui, S.; Diez, J.; Toribio, R.; Gómez-Baena, G.; Dufresne, A.; García-Fernández, J.M. Glutamine synthetase from the marine cyanobacteria Prochlorococcus spp.: Characterization, phylogeny and response to nutrient limitation. Environ. Microbiol. 2003, 5, 412–423. [Google Scholar] [CrossRef]
- Mulholland, M.R.; Lomas, M.W. Nitrogen uptake and assimilation-sciencedirect. In Nitrogen in the Marine Environment, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 303–384. [Google Scholar]
- Zhou, M.J.; Zhu, M.Y. Progress of the project “Ecology and Oceanography of Harmful Algal Blooms in China”. Adv. Earth Sci. 2006, 21, 673–679. [Google Scholar]
- Yan, T.; Zhou, M.J.; Qian, P.Y. Competition among dinoflagellate Alexandrium tamarense, raphidophyte Heterosigma carterae and diatom Skeletonema costatum under combinations of two temperatures and five salinities. J. Oceanol. Limnol. 2003, 21, 245–250. [Google Scholar]
- Li, R.X.; Zhu, M.Y.; Wang, Z.L.; Shi, X.Y.; Chen, B.Z. Mesocosm experiment on competition between two HAB species in East China Sea. Chin. Appl. Ecol. 2003, 14, 1049–1054. [Google Scholar]
- Chen, H.L.; Lu, S.H.; Zhang, C.S.; Zhu, D.D. A Survey on the red tide of Prorocentrum donghaiense in East China Sea, 2004. Ecol. Sci. 2006, 25, 226–230. [Google Scholar]
- Zhang, C.S.; Wang, J.T.; Zhu, D.D.; Shi, X.Y.; Wang, X.L. The preliminary analysis of nutrients in harmful algal blooms in the East China Sea in the spring and summer of 2005. Acta Oceanol. Sin. 2008, 30, 153–159. [Google Scholar]
- Zhou, M.J.; Shen, Z.L.; Yu, R.C. Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang (Yangtze) River. Cont. Shelf Res. 2008, 28, 1483–1489. [Google Scholar] [CrossRef]
- Harrison, P.; Parslow, J.S.; Conway, H.L. Determination of nutrient uptake kinetic parameters: A comparison of methods. Mar. Ecol. Prog. Ser. 1989, 52, 301–312. [Google Scholar] [CrossRef]
- Nishikawa, T.; Tarutani, R.; Yamamoto, R. Nitrate and phosphate uptake kinetics of the harmful diatom Coscinodiscus wailesii, a causative organism in the bleaching of aquacultured Porphyra thalli. Harmful Algae 2010, 9, 563–567. [Google Scholar] [CrossRef]
- Wood, E.D.; Armstrong, F.A.J.; Richards, F.A. Determination of nitrate in sea water by cadmium-copper reduction to nitrate. J. Mar. Biol. Assoc. UK 1967, 47, 23–31. [Google Scholar] [CrossRef]
- Solórzano, L. Determination of ammonium in natural waters by the phenolhypochlorite method. Limnol. Occanogr. 1969, 14, 799–801. [Google Scholar] [CrossRef]
- Takabayashi, M.; Wilkerson, F.P.; Robertson, D. Response of glutamine synthetase gene transcription and enzyme activity to external nitrogen sources in the diatom Skeletonema costatum (bacillariophyceae)1. J. Phycol. 2005, 41, 84–94. [Google Scholar] [CrossRef]
- Matsuda, A.; Nishijima, T.; Fukami, K. Effects of nitrogenous and phosphorus nutrients on the growth of toxic dinoflagellate Alexandrium catenella. Nippon Suisan Gakk. 1999, 65, 847–855. [Google Scholar] [CrossRef]
- Collos, Y.; Gagne, C.; Laabir, M.; Vaquer, A. Nitrogenous nutrition of Alexandrium catenella (Dinophyceae) in cultures and in Thau lagoon, southern France. J. Phycol. 2004, 40, 96–103. [Google Scholar] [CrossRef]
- MacIsaac, J.J.; Grunseich, G.S.; Glover, H.E.; Clarice, M.Y. Light and nutrient limitation in Gonyaulax excavate: Nitrogen and carbon trace results. In Toxic Dinoflagellate Blooms; Elsevier: New York, NY, USA, 1979; pp. 107–110. [Google Scholar]
- Eppley, R.W.; Coatsworth, J.L.; Solorzano, L. Studies of nitrate reductase in marine phytoplankton. Limnol. Oceanogr. 1969, 14, 194–205. [Google Scholar] [CrossRef]
- Nakamura, Y. Kinetics of nitrogen- or phosphorus-limited growth and effects of growth conditions on nutrient uptake in Chattonella antiqua. J. Oceanogr. Soc. Japan 1985, 41, 381–387. [Google Scholar] [CrossRef]
- Bressler, S.L.; Ahmed, S.I. Detection of glutamine synthetase activity in marine phytoplankton: Optimization of the biosynthetic assay. Mar. Ecol. Prog. Ser. 1984, 14, 207–217. [Google Scholar] [CrossRef]
- Eppley, R.W.; Thomas, W.H. Comparison of half-saturation constants for growth and nitrate uptake of marine phytoplankton. J. Phycol. 2010, 5, 375–379. [Google Scholar] [CrossRef]
- Bonachela, J.A.; Levin, R. Dynamic model of flexible phytoplankton nutrient uptake. Proc. Natl. Acad. Sci. USA 2011, 108, 20633–20638. [Google Scholar] [CrossRef]
- Huang, K.; Feng, Q.; Zhang, Y.; Ou, L.; Qi, Y. Comparative uptake and assimilation of nitrate, ammonium, and urea by dinoflagellate Karenia mikimotoi and diatom Skeletonema costatum s.l. in the coastal waters of the East China Sea. Mar. Pollut. Bull. 2020, 155, 111200. [Google Scholar] [CrossRef]
- Ou, L.J. Ecophysiological Responses of Typical Harmful Algal Bloom Species to Phosphorus; Xiamen University: Xiamen, China, 2008. [Google Scholar]
- Liao, T.C.; Yu, H.G.; Dai, C.J.; Zhao, M. Impact of cell size effect on nutrient-phytoplankton dynamics. Complexity 2019, 2019, 8205696. [Google Scholar] [CrossRef]
- Button, D.K. Biochemical basis for whole-cell uptake kinetics: Specific affinity, oligotrophic capacity, and the meaning of the Michaelis constant. Appl. Environ. Microbiol. 1991, 57, 2033–2038. [Google Scholar] [CrossRef]
- Joseph, L.; Villareal, T.A. Nitrate reductase activity as a measure of nitrogen incorporation in Rhizosolenia formasa (H. Peragallo): Internalnitrate and diel effects. J. Exp. Mar. Biol. Ecol. 1998, 229, 159–176. [Google Scholar] [CrossRef]
- Lopes, P.F.; de Oliveira, M.C.; Colepicolo, P. Characterization and daily variation of nitrate reductase in Gracilaria tenuistipitata (Rhodophyta). Biochem. Biophys. Res. Commun. 2002, 295, 50–54. [Google Scholar] [CrossRef]
- Kim, J.; Brown, C.M.; Min, K.K.; Burrows, E.H.; Falkowski, P.G. Effect of cell cycle arrest on intermediate metabolism in the marine diatom Phaeodactylum tricornutum. Proc. Natl. Acad. Sci. USA 2017, 114, E8007–E8016. [Google Scholar] [CrossRef]
- Ramalho, C.B.; Hastings, J.W.; Colepicolo, P. Circadian oscillation of nitrate reductase activity in Gonyaulax polyedra is due to changes in cellular protein levels. Plant Physiol. 1995, 107, 225–231. [Google Scholar] [CrossRef]
- Varela, D.E.; Harrison, P.J. Effect of ammonium on nitrate utilization by Emiliania huxleyi, a coccolithophore from the oceanic northeastern Pacific. Mar. Ecol. Prog. Ser. 1999, 186, 67–74. [Google Scholar] [CrossRef]
- Serra, J.L.; Llama, M.J.; Cadenas, E. Characterization of the nitrate reductase enzyme activity in the diatom Skeletonema costatum. Plant Sci. Lett. 1978, 13, 41–48. [Google Scholar] [CrossRef]
- Gao, Y.; Smith, G.J.; Alberte, R.S. Nitrate reductase from the marine diatom Skeletonema costatum biochemical and immunological characterization. Plant Physiol. 1993, 103, 1437–1445. [Google Scholar] [CrossRef]
- Berges, J.A.; Harrison, P.J. Nitrate reductase activity quantitatively predicts the rate of nitrate incorporation under steady state light limitation: A revised assay and characterization of the enzyme in three species of marine phytoplankton. Limnol. Oceanogr. 1995, 40, 82–93. [Google Scholar] [CrossRef]
- García-Fernández, J.M.A.; López-Ruiz, J.; Alhama, J.D. Light regulation of glutamine synthetase in the green alga Monoraphidium braunii. J. Plant Physiol. 1995, 146, 577–583. [Google Scholar] [CrossRef]
- Maurin-Defossez, C.; le Gal, Y. Diel periodicity of glutamine synthetase activity during the cell cycle of Emiliania huxleyi. Plant Physiol. Biochem. 1998, 36, 233–236. [Google Scholar] [CrossRef]
Figure 1.
The nitrate concentrations of N-starved cultures of Skeletonema costatum, Prorocentrum donghaiense and Alexandrium pacificum after addition of nitrate (A,C,E) and the nitrate uptake rate of S. costatum, P. donghaiense and A. pacificum as a function of the ambient nitrate concentration, respectively (B,D,F). The curve was adjusted to the observed values using a method of the lowest nonlinear square. Data are described as mean ± standard deviation (n = 3).
Figure 1.
The nitrate concentrations of N-starved cultures of Skeletonema costatum, Prorocentrum donghaiense and Alexandrium pacificum after addition of nitrate (A,C,E) and the nitrate uptake rate of S. costatum, P. donghaiense and A. pacificum as a function of the ambient nitrate concentration, respectively (B,D,F). The curve was adjusted to the observed values using a method of the lowest nonlinear square. Data are described as mean ± standard deviation (n = 3).

Figure 2.
Changes in ammonium concentrations of N-starved cultures of Skeletonema costatum, Prorocentrum donghaiense and Alexandrium pacificum after addition of ammonium (A,C,E) and ammonium uptake rate of S. costatum, P. donghaiense and A. pacificum as a function of the ambient ammonium concentrations, respectively (B,D,F). The curve was adjusted to the observed values using a method of the lowest nonlinear square. Data are described as mean ± standard deviation (n = 3).
Figure 2.
Changes in ammonium concentrations of N-starved cultures of Skeletonema costatum, Prorocentrum donghaiense and Alexandrium pacificum after addition of ammonium (A,C,E) and ammonium uptake rate of S. costatum, P. donghaiense and A. pacificum as a function of the ambient ammonium concentrations, respectively (B,D,F). The curve was adjusted to the observed values using a method of the lowest nonlinear square. Data are described as mean ± standard deviation (n = 3).

Figure 3.
Change in relative (%) GS and NR activity with respect to reaction time. Data are described as mean ± standard deviation (n = 3).
Figure 3.
Change in relative (%) GS and NR activity with respect to reaction time. Data are described as mean ± standard deviation (n = 3).

Figure 4.
Kinetic curves of nitrate reductase for Skeletonema costatum (A) and Prorocentrum donghaiense (B). The solid line in each panel is the Michaelis-Menten equation fit to the data. Data are described as mean ± standard deviation (n = 3).
Figure 4.
Kinetic curves of nitrate reductase for Skeletonema costatum (A) and Prorocentrum donghaiense (B). The solid line in each panel is the Michaelis-Menten equation fit to the data. Data are described as mean ± standard deviation (n = 3).

Figure 5.
Representative GS kinetic curves for Skeletonema costatum (A,B), Prorocentrum donghaiense (C) and Alexandrium pacificum (D,E). The solid line in each panel is the Michaelis-Menten equation fit to the data. Data are described as mean ± standard deviation (n = 3).
Figure 5.
Representative GS kinetic curves for Skeletonema costatum (A,B), Prorocentrum donghaiense (C) and Alexandrium pacificum (D,E). The solid line in each panel is the Michaelis-Menten equation fit to the data. Data are described as mean ± standard deviation (n = 3).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Kinetics parameters for uptake of nitrate by Skeletonema costatum, Prorocentrum donghaiense and Alexandrium pacificum.
Table 1.
Kinetics parameters for uptake of nitrate by Skeletonema costatum, Prorocentrum donghaiense and Alexandrium pacificum.
| Species | KS (µM) | Vmax (pmol cell−1 h−1) | Reference |
|---|---|---|---|
| Alexandrium catenella | 7.7 | — | [33] |
| Alexandrium pacificum | 10.02 | 1.30 | Present study |
| Alexandrium catenella | 0.6–28.1 | — | [34] |
| Alexandrium tamarense | 2.84 | — | [35] |
| Alexandrium minutum | 0.22–0.28 | 0.29–0.40 | [9] |
| Gymnodinium catenatum | 7.59 | 6.48 | [8] |
| Prorocentrum donghaiense | 5.98 | 0.098 | Present study |
| Skeletonema costatum | 1.19 | 0.021 | Present study |
| Skeletonema costatum | 0.4–0.5 | 0.063 | [36] |
| Prorocentrum minimum | 5.0 | 0.102 | [11] |
| Thalassiosira weissflogii | 2.8 | 0.310 | [11] |
| Chaetoceros sp. | 3.1 | 0.024 | [11] |
| Pavlova lutheri | 22.7 | 0.021 | [11] |
Note(s): Data are described as mean ± standard deviation (n = 3). “—” indicates not detected.
Table 2.
Kinetics parameters for uptake of ammonium by Skeletonema costatum, Prorocentrum donghaiense and Alexandrium pacificum.
Table 2.
Kinetics parameters for uptake of ammonium by Skeletonema costatum, Prorocentrum donghaiense and Alexandrium pacificum.
| Species | KS (µM) | Vmax (pmol cell−1 h−1) | Reference |
|---|---|---|---|
| Alexandrium catenella | 3.3 | — | [33] |
| Alexandrium pacificum | 4.27 | 4.21 | Present study |
| Alexandrium catenella | 0.6–28.1 | — | [34] |
| Alexandrium minutum | 0.25–0.33 | 0.65–0.82 | [9] |
| Alexandrium tamarense | 1.49 | — | [35] |
| Gymnodinium catenatum | 33.6 | 3.37 | [8] |
| Prorocentrum donghaiense | 2.04 | 0.296 | Present study |
| Skeletonema costatum | 1.12 | 0.037 | Present study |
| Skeletonema costatum | 0.8–3.6 | — | [36] |
| Chattonella antiqua | 2.19 | 2.02 | [37] |
Note(s): Data are described as mean ± standard deviation (n = 3). “—” indicates not detected.
Table 3.
Ks and Vmax values for the different levels of substrate NO3− for Skeletonema costatum and Prorocentrum donghaiense.
Table 3.
Ks and Vmax values for the different levels of substrate NO3− for Skeletonema costatum and Prorocentrum donghaiense.
| Species | Low Concentration | High Concentration | Reference | ||
|---|---|---|---|---|---|
| KS (µM) | Vmax (µmol NO2− min−1 (mg protein)−1) | KS (µM) | Vmax (µmol NO2− min−1 (mg protein)−1) | ||
| Skeletonemacostatum | 82.69 | 2.26 | 91.29 | 2.4 | Present study |
| Prorocentrumdonghaiense | 168.48 | 0.29 | 168.48 | 0.29 | Present study |
| Skeletonemacostatum | 290 | — | — | — | [11] |
Note(s): Data are described as mean ± standard deviation (n = 3). “—” indicates not detected.
Table 4.
Km and Vmax values for the different levels of substrate NH4+ for Skeletonema costatum, Alexandrium pacificum and Prorocentrum donghaiense.
Table 4.
Km and Vmax values for the different levels of substrate NH4+ for Skeletonema costatum, Alexandrium pacificum and Prorocentrum donghaiense.
| Species | Low Concentration | High Concentration | Reference | ||
|---|---|---|---|---|---|
| KS (µM) | Vmax (µmol PO43− min−1 (mg protein)−1) | KS (µM) | Vmax (µmol PO43− min−1 (mg protein)−1) | ||
| Skeletonemacostatum | 9.30 | 0.34 | 41.1 | 0.57 | Present study |
| Prorocentrumdonghaiense | 0.04 | 0.85 | 0.04 | 0.85 | Present study |
| Alexandriumpacificum | 0.16 | 1.15 | 10.35 | 1.66 | Present study |
| Skeletonema costatum | 8.20 | 0.32 | — | — | [38] |
| Isochrysisgalbana | 8.20 | 0.22 | — | — | [38] |
| Pavlovalutheri | 1.80 | 0.39 | — | — | [38] |
Note(s): Data are described as mean ± standard deviation (n = 3). “—” indicates not detected.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ding, G.; Chen, H.; Gu, H.; Zhang, Y.; Li, R.; Zhang, S. Inorganic Nitrogen Uptake Characteristics of Three Typical Bloom-Forming Algae in the East China Sea. Water 2022, 14, 2580. https://doi.org/10.3390/w14162580
AMA Style
Ding G, Chen H, Gu H, Zhang Y, Li R, Zhang S. Inorganic Nitrogen Uptake Characteristics of Three Typical Bloom-Forming Algae in the East China Sea. Water. 2022; 14(16):2580. https://doi.org/10.3390/w14162580
Chicago/Turabian StyleDing, Guangmao, Huorong Chen, Haifeng Gu, Youquan Zhang, Rongmao Li, and Shufeng Zhang. 2022. "Inorganic Nitrogen Uptake Characteristics of Three Typical Bloom-Forming Algae in the East China Sea" Water 14, no. 16: 2580. https://doi.org/10.3390/w14162580
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.