
Gastropod Assemblages Associated with Habitat Heterogeneity and Hydrological Shifts in Two Shallow Waterbodies

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
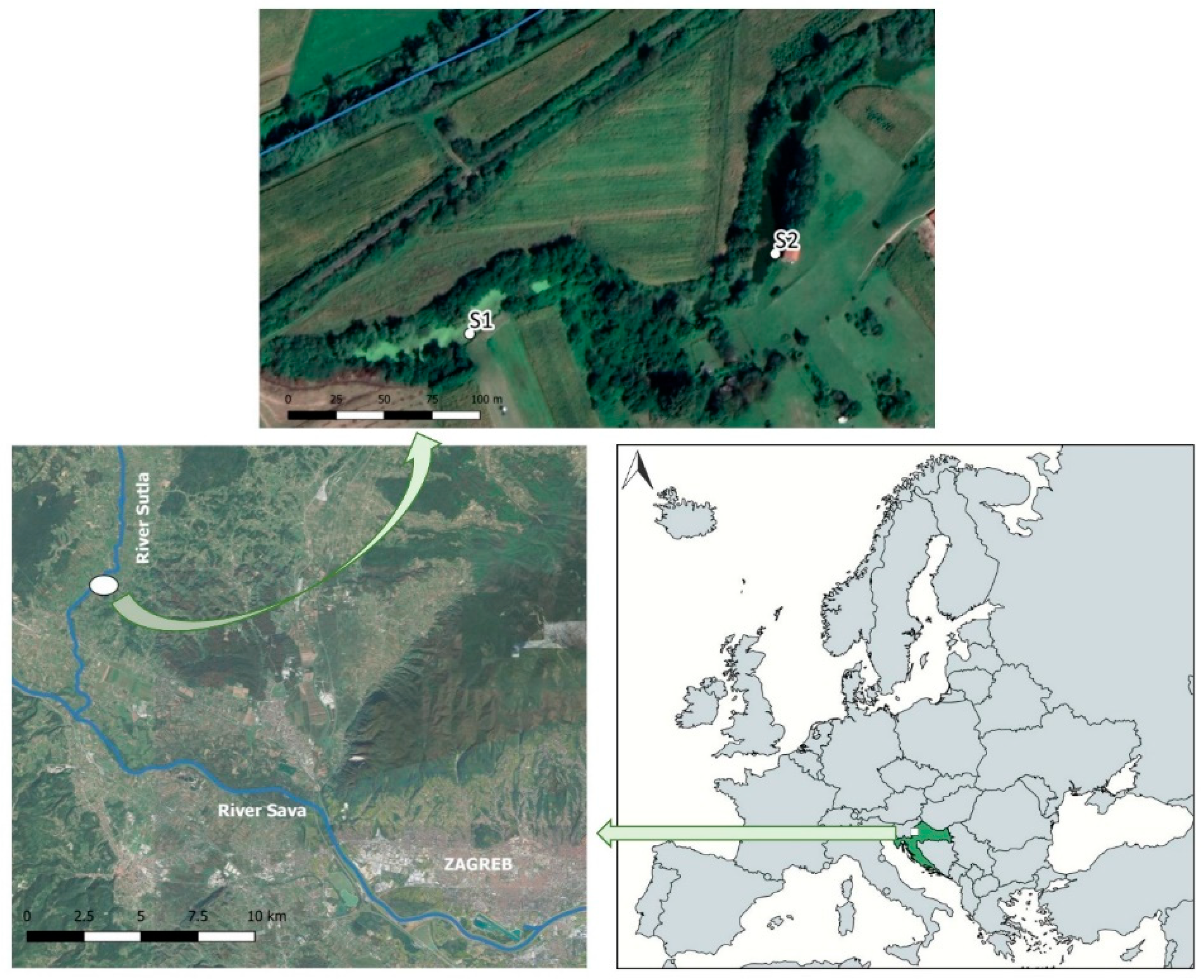
2.1. Study Area
2.2. Macroinvertebrates and Epiphyton Sampling and Biocoenotical Analyses
2.3. Environmental Parameters
2.4. Data Analysis
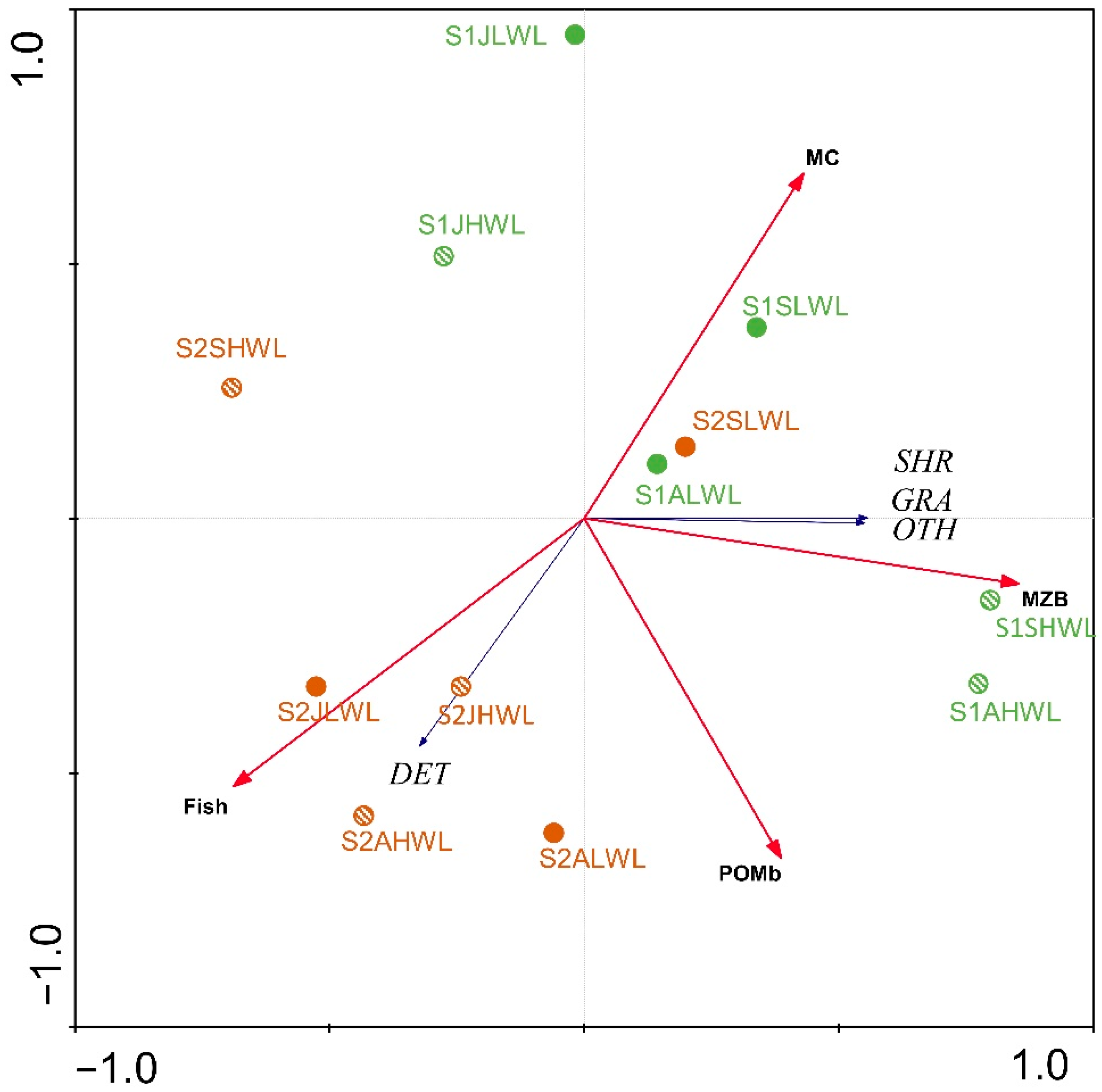
3. Results
3.1. Environmental Variables
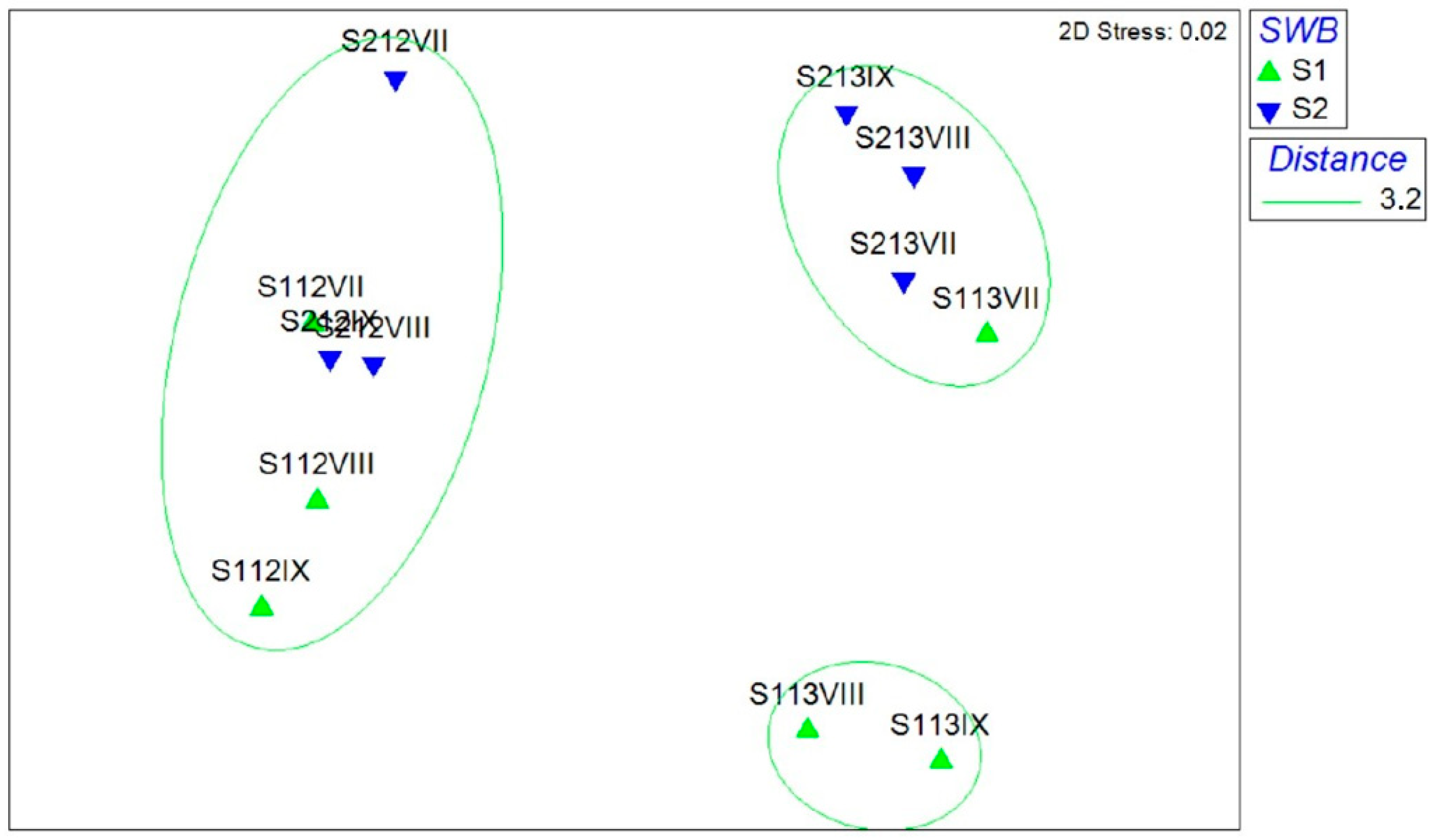
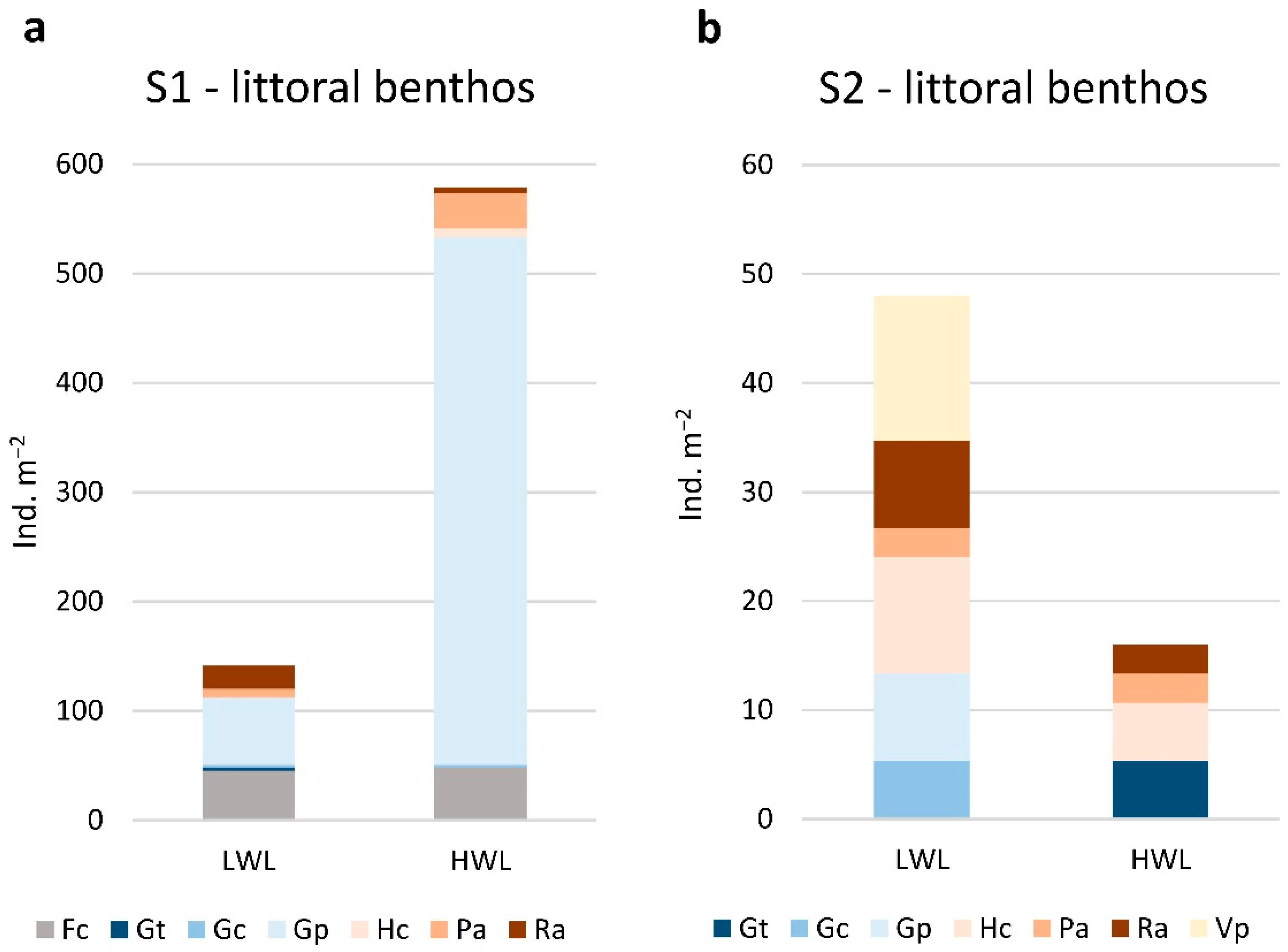
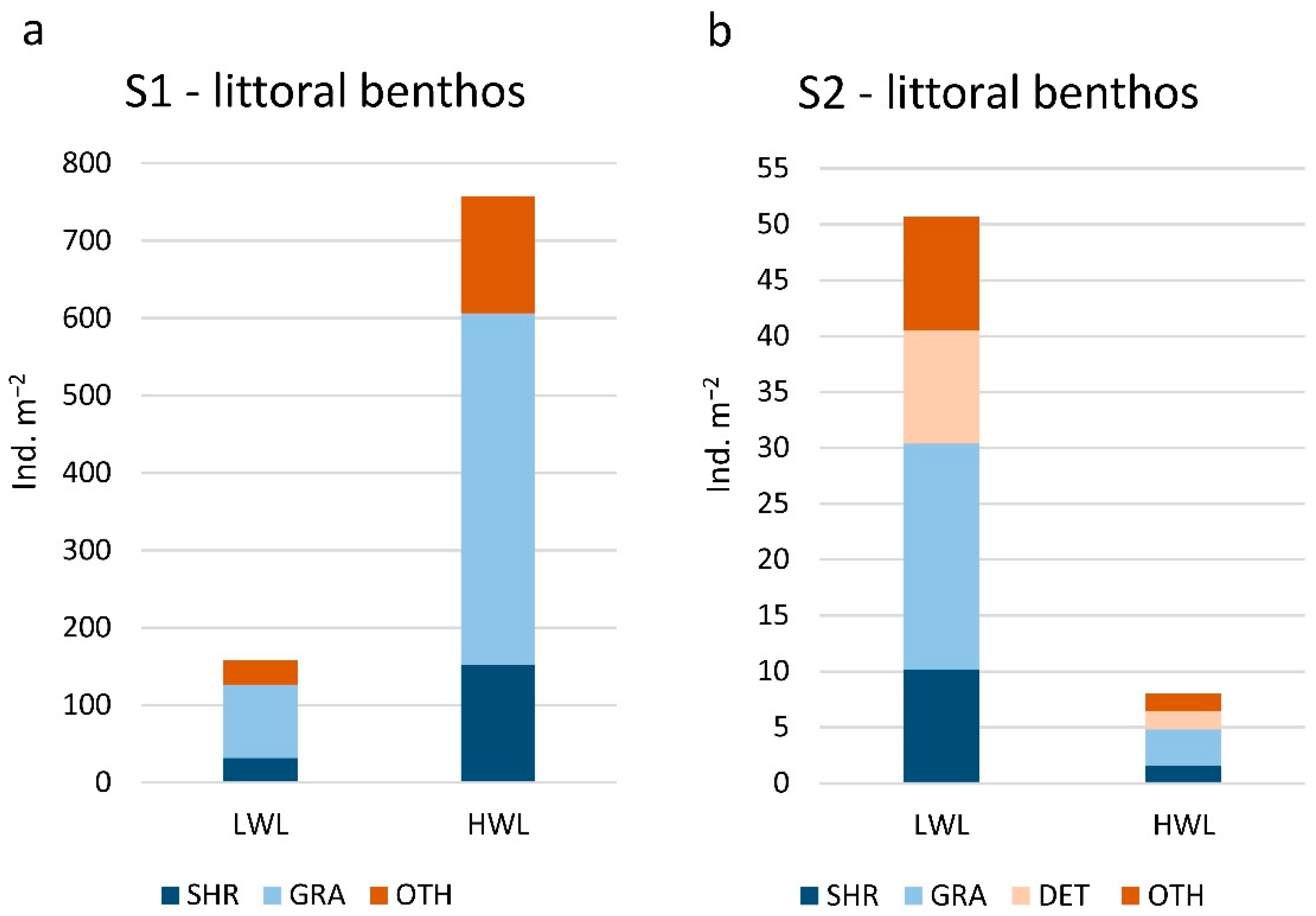
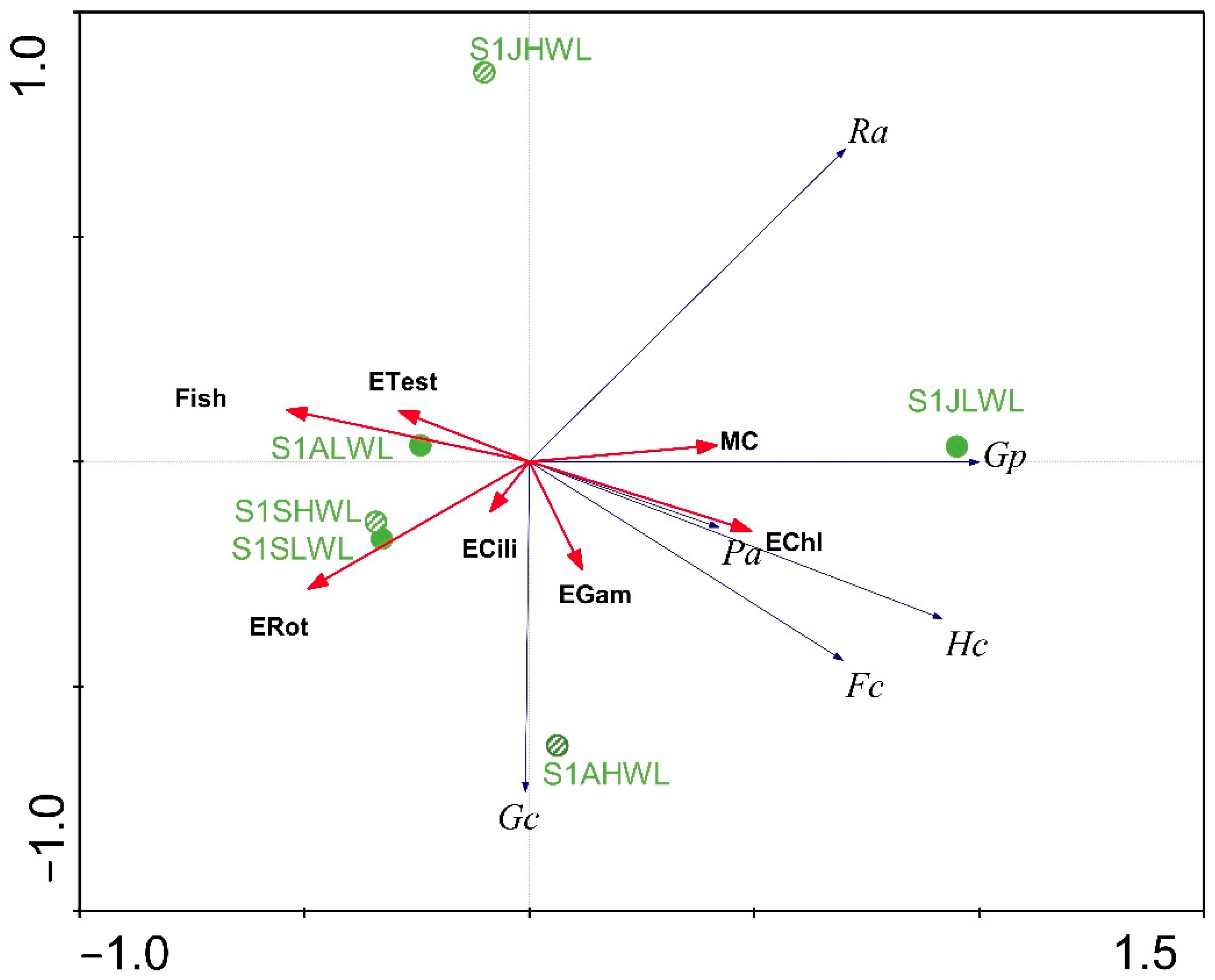
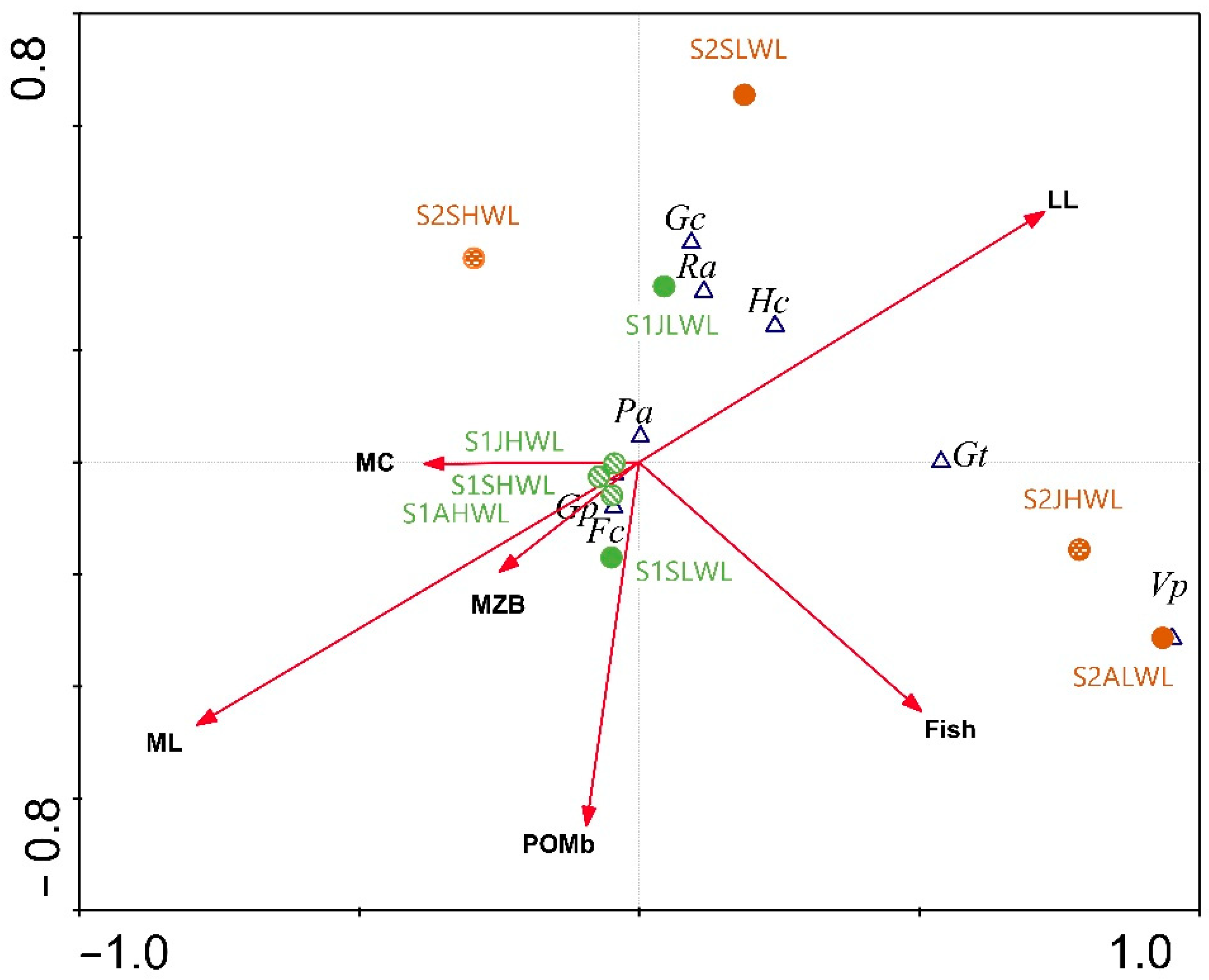
3.2. Gastropod Assemblage
3.3. Gastropod Functional Feeding Groups
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castro, B.B.; Antunes, S.C.; Pereira, R.; Soares, A.M.V.M.; Gonçalves, F. Rotifer community structure in three shallow lakes: Seasonal fluctuations and explanatory factors. Hydrobiologia 2005, 543, 221–232. [Google Scholar] [CrossRef]
- Špoljar, M.; Dražina, T.; Lajtner, J.; Duić Sertić, M.; Radanović, I.; Wallace, R.L.; Matulić, D.; Tomljanović, T. Zooplankton assemblage in four temperate shallow waterbodies in association with habitat heterogeneity and alternative states. Limnologica 2018, 71, 51–61. [Google Scholar] [CrossRef]
- Kozak, A.; Celewicz-Gołdyn, S.; Kuczyńska-Kippen, N. Cyanobacteria in small water bodies: The effect of habitat and catchment area conditions. Sci. Total Environ. 2019, 646, 1578–1587. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N.; Špoljar, M.; Mleczek, M.; Zhang, C. Elodeids, but not helophytes, increase community diversity and reduce trophic state: Case study with rotifer indices in field ponds. Ecol. Indic. 2021, 128, 107829. [Google Scholar] [CrossRef]
- Cremona, F.; Planas, D.; Lucotte, M. Biomass and composition of macroinvertebrate communities associated with different types of macrophyte architectures and habitats in a large fluvial lake. Fund. Appl. Limnol. 2008, 171, 119–130. [Google Scholar] [CrossRef]
- Fan, Z.; Wang, Z.; Li, Y.; Wang, W.; Tang, C.; Zeng, F. Water Level Fluctuation under the Impact of Lake Regulation and Ecological Implication in Huayang Lakes, China. Water 2020, 12, 702. [Google Scholar] [CrossRef] [Green Version]
- Špoljar, M.; Sertić Perić, M.; Wang, H.; Zhang, C.; Kuczyńska-Kippen, N.; Fressl, J.; Ercegovac, Z. Does the size structure of the littoral community reflect water level fluctuations in shallow waterbodies? Ecol. Indic. 2021, 132, 108330. [Google Scholar] [CrossRef]
- Blindow, I. Long- and short-term dynamics of submerged macrophytes in two shallow eutrophic lakes. Freshw. Biol. 1992, 28, 15–27. [Google Scholar] [CrossRef]
- Van Geest, G.J.; Coops, H.; Scheffer, M.; van Nes, E.H. Long transients near the ghost of a stable state in eutrophic shallow lakes with fluctuating water levels. Ecosystems 2007, 10, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Mjelde, M.; Hellsten, S.; Ecke, F. A water level drawdown index for aquatic macrophytes in Nordic lakes. Hydrobiologia 2013, 704, 141–151. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N.; Nagengast, B.; Celewicz-Gołdyn, S.; Klimko, M. Zooplankton community structure within various macrophyte stands of a small water body in relation to seasonal changes in water level. Oceanol. Hydrobiol. Stud. 2009, 38, 125–133. [Google Scholar] [CrossRef]
- Vadeboncoeur, Y.; Steinman, A.D. Periphyton Function in Lake Ecosystems. Sci. World J. 2002, 2, 1449–1468. Available online: https://academic.oup.com/bioscience/article/52/1/44/291662?login=true (accessed on 7 June 2022). [CrossRef] [Green Version]
- Carmignani, J.R.; Roy, A.H. Ecological impacts of winter water level drawdowns on lake littoral zones. Aquat. Sci. 2017, 79, 803–824. [Google Scholar] [CrossRef] [Green Version]
- Bolpagni, R.; Poikane, S.; Laini, A.; Bagella, S.; Bartoli, M.; Cantonati, M. Ecological and Conservation Value of Small Standing-Water Ecosystems: A Systematic Review of Current Knowledge and Future Challenges. Water 2019, 11, 402. [Google Scholar] [CrossRef] [Green Version]
- Water Resources in Europe in the Context of Vulnerability: EEA 2012 State of Water Assessment; EEA Report no. 11/2012; European Environment Agency: Copenhagen, Denmark, 2012; p. 92.
- Evtimova, V.V.; Donohue, I. Water-level fluctuations regulate the structure and functioning of natural lakes. Freshw. Biol. 2016, 61, 251–264. [Google Scholar] [CrossRef]
- Bill, A.R. Effects of Lake Water Level Fluctuations on Macrophytes and Littoral Macroinvertebrates. Doctoral Thesis, University of Stirling, Scotland, UK, 2020. Available online: https://dspace.stir.ac.uk/handle/1893/31954#.Ym0sPNpBxPY (accessed on 25 March 2022).
- Strayer, D.L.; Findlay, S.E.G. Ecology of freshwater shore zones. Aquat. Sci. 2010, 72, 127–163. [Google Scholar] [CrossRef] [Green Version]
- O’Leary, S.J.; Wantzen, K.M. Flood pulse effects on benthic invertebrate assemblages in the hypolacustric interstitial zone of Lake Constance. Int. J. Limnol. 2012, 48, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Gownaris, N.J.; Rountos, K.J.; Kaufman, L.; Kolding, J.; Lwiza, K.M.M.; Pikitch, E.K. Water level fluctuations and the ecosystem functioning of lakes. J. Great Lakes Res. 2018, 44, 1154–1163. [Google Scholar] [CrossRef]
- Briers, R.A.; Biggs, J. Indicator taxa for the conservation of pond invertebrate diversity. Aquat. Conserv. Mar. Freshw. Ecosyst. 2003, 13, 323–330. [Google Scholar] [CrossRef]
- Ruggiero, A.; Céréghino, R.; Figuerola, J.; Marty, P.; Angélibert, S. Farm ponds make a contribution to the biodiversity of aquatic insects in a French agricultural landscape. C. R. Biol. 2008, 331, 298–308. [Google Scholar] [CrossRef]
- Vilenica, M.; Pozojević, I.; Vučković, N.; Mihaljević, Z. How suitable are man-made water bodies as habitats for Odonata? Knowl. Manag. Aquat. Ecosyst. 2020, 421, 13. [Google Scholar] [CrossRef]
- de los Rios-Escalante, P.; Roa, G. Species assemblages of zooplanktonic crustaceans in mountain shallow ponds of Chile (Parque Cañi). Zoologia 2010, 27, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Barbiero, R.P.; Rudstam, L.G.; Watkins, J.M.; Lesht, B.M. A cross-lake comparison of crustacean zooplankton communities in the Laurentian Great Lakes, 1997–2016. J. Great Lakes Res. 2019, 45, 672–690. [Google Scholar] [CrossRef]
- Lombardo, P.; Cooke, G.D. Resource use and partitioning by two co-occuring freshwater gastropod species. Arch. Hydrobiol. 2004, 159, 229–251. [Google Scholar] [CrossRef]
- Kołodziejczyk, A. Occurrence of gastropoda in the lake littoral and their role in the production and transformation of detritus. II. Ecological activity of snails. Ekol. Pol. 1984, 32, 469–492. [Google Scholar]
- Lodge, D. Selective grazing on periphyton: A determinant of freshwater gastropod microdistributions. Freshw. Biol. 1986, 16, 831–841. [Google Scholar] [CrossRef]
- Stickle, W.B.; Carrington, E.; Hayford, H. Seasonal changes in the thermal regime and gastropod tolerance to temperature and desiccation stress in the rocky intertidal zone. J. Exp. Mar. Biol. 2017, 488, 83–91. [Google Scholar] [CrossRef]
- Brönmark, C. Interactions between Epiphytes, Macrophytes and Freshwater Snails. J. Moll. Stud. 1989, 55, 299–311. Available online: https://www.jstor.org/stable/4219655 (accessed on 16 February 2022). [CrossRef]
- Cardoso, L.D.S.; Marques, D.D.M. Structure of the zooplankton community in a subtropical shallow lake (Itapeva Lake-South of Brazil) and its relationship to hydrodynamic aspects. Hydrobiologia 2004, 518, 123–134. [Google Scholar] [CrossRef]
- Špoljar, M.; Dražina, T.; Ostojić, A.; Miliša, M.; Gligora Udovič, M.; Štafa, D. Bryophyte communities and seston in a karst stream (Jankovac Stream, Papuk Nature Park, Croatia). Ann. Limnol. 2012, 481, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Dražina, T.; Korša, A.; Špoljar, M.; Maguire, I.; Klobučar, G. Epifauna of native and alien freshwater crayfish species (Crustacea: Decapoda): A host-specific community? Freshw. Sci. 2018, 37, 593–604. [Google Scholar] [CrossRef]
- Zhang, C.; Huang, Y.; Špoljar, M.; Zhang, W.; Kuczyńska-Kippen, N. Epiphyton dependency of macrophyte biomass in shallow reservoirs and implications for water transparency. Aquat. Bot. 2018, 150, 46–52. [Google Scholar] [CrossRef]
- Lombardo, P.; Cooke, G.D. Consumption and preference of selected food types by two freshwater gastropod species. Arch. Hydrobiol. 2002, 155, 667–685. [Google Scholar] [CrossRef]
- Messyasz, B.; Kuczyńska-Kippen, N.; Nagengast, B. The epiphytic communities of various ecological types of aquatic vegetation of five pastoral ponds. Biologia 2009, 64, 88–96. [Google Scholar] [CrossRef]
- Walker, P.D.; Wijnhoven, S.; van der Velde, G. Macrophyte presence and growth form influence macroinvertebrate community structure. Aquat. Bot. 2013, 104, 80–87. [Google Scholar] [CrossRef]
- Lv, T.; Guan, X.; Fan, S.; Hanm, C.; Gao, Z.; Liu, C. Snail communities increase submerged macrophyte growth by grazing epiphytic algae and phytoplankton in a mesocosm experiment. Ecol. Evol. 2022, 12, e8615. [Google Scholar] [CrossRef]
- Kloskowski, J.; Nieoczym, M.; Stryjecki, R. Between-habitat distributions of pond tadpoles and their insect predators in response to fish presence. Hydrobiologia 2020, 847, 1343–1356. [Google Scholar] [CrossRef] [Green Version]
- Martin, T.H.; Crowder, L.B.; Dumas, C.F.; Burkholder, J.M. Indirect effects of fish on macrophytes in Bays Mountain Lake: Evidence for a littoral trophic cascade. Oecologia 1992, 89, 476–481. [Google Scholar] [CrossRef]
- Zębek, E.; Szymańska, U. Gastropods and perophytic algae relationships in the vicinity of a small hydroelectric plant on the Pasleka River in norteast Poland. Arch. Pol. Fish. 2014, 22, 69–80. [Google Scholar] [CrossRef]
- Bae, M.-J.; Park, Y.-S. Key Determinants of Freshwater Gastropod Diversity and Distribution: The Implications for Conservation and Management. Water 2020, 12, 1908. [Google Scholar] [CrossRef]
- Moisez, E.; Spilmont, N.; Seuront, L. Microhabitats choice in intertidal gastropods is species-, temperature- and habitat-specific. J. Therm. Biol. 2020, 94, 102785. [Google Scholar] [CrossRef] [PubMed]
- Evtimova, V.V.; Donohue, I. Quantifying ecological responses to amplified water level fluctuations in standing waters: An experimental approach. J. Appl. Ecol. 2014, 51, 1282–1291. [Google Scholar] [CrossRef]
- Glöer, P. The Freshwater Gastropods of the West-Palaearctis: Identification Key, Anatomy, Ecology, Distribution; Biodiversity Research Lab: Hetlingen, Germany, 2019. [Google Scholar]
- Moog, O. Fauna Aquatica Austriaca, Edition 2002. In Wasserwirtschaftskataster; Moog, O., Ed.; Bundesministerium für Land- und Forstwirtschaft, Umwelt und Wasserwirtschaft: Vienna, Austria, 2002. [Google Scholar]
- APHA Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998.
- Špoljar, M.; Dražina, T.; Habdija, I.; Meseljević, M.; Grčić, Z. Contrasting zooplankton assemblages in two oxbow lakes with low transparencies and narrow emergent macrophyte belts (Krapina River, Croatia). Int. Rev. Hydrobiol. 2011, 96, 175–190. [Google Scholar] [CrossRef]
- Nusch, E.A. Comparison of different methods for chlorophyll and phaeopigment determination. Arch. Für Hydrobiol. 1980, 14, 14–36. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed.; W.H. Freeman: New York, NY, USA, 1995. [Google Scholar]
- ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Wantzen, K.M.; Junk, W.J.; Rothhaupt, K.-O. An extension of the floodpulse concept (FPC) for lakes. Hydrobiologia 2008, 613, 151–170. [Google Scholar] [CrossRef] [Green Version]
- Gownaris, N.J.; Pikitch, E.K.; Aller, J.Y.; Kaufman, L.S.; Kolding, J.; Lwiza, K.M.M.; Obiero, K.O.; Ojwang, W.O.; Malala, J.O.; Rountos, K.J. Fisheries and water level fluctuations in the world’s largest desert lake. Ecohydrology 2017, 10, e1769. [Google Scholar] [CrossRef]
- Mastrantuono, L.; Solimini, A.G.; Nõges, P.; Bazzanti, M. Plant-associated invertebrates and hydrological balance in the large volcanic Lake Bracciano (Central Italy) during two years with different water levels. Hydrobiologia 2008, 599, 143–152. [Google Scholar] [CrossRef]
- Jezidžić, L. Freshwater Snails (Gastropoda) as Indicators of Čonakut Channel Trophic Level in Kopački Rit. Master Thesis, Department of Biology, University J. J. Strossmayer in Osijek, Osijek, Croatia, 2010; p. 89. Available online: https://www.bib.irb.hr/544611 (accessed on 26 April 2022). (In Croatian).
- Pip, E. The ecology of freshwater gastropods in the central Canada region. Nautilus 1986, 100, 56–66. [Google Scholar]
- Cattaneo, A.; Galanti, G.; Gentinetta, S.; Romo, S. Epiphytic algae and macroinvertebrates on submerged and floating-leaved macrophytes in an Italian lake. Freshw. Biol. 1998, 39, 725–740. [Google Scholar] [CrossRef]
- Lorencová, E.; Beran, L.; Nováková, M.; Horsáková, V.; Rowson, B.; Hlaváč, J.Č.; Nekola, J.C.; Horsák, M. Invasion at the population level: A story of the freshwater snails Gyraulus parvus and G. laevis. Hydrobiologia 2021, 848, 4661–4671. [Google Scholar] [CrossRef]
- Ville, S.P. Shipping in the Port of Newcastle, 1780–1800. J. Transp. Hist. 1988, 9, 60–77. [Google Scholar] [CrossRef]
- Van Leeuwen, C.H.A.; Van Der Velde, G.; Van Lith, B.; Klaassen, M. Experimental quantification of long distance dispersal potential of aquatic snails in the gut of migratory birds. PLoS ONE 2012, 7, e32292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beran, L.; Horsák, M. Gyraulus parvus (Mollusca: Gastropoda) in the Czech Republic. Acta Soc. Zool. Bohem. 2002, 66, 81–84. [Google Scholar]
- Quijón, P.A.; Contreras, H.; Jaramillo, E. Population Biology of the Intertidal Snail Chilina ovalis Sowerby (Pulmonata) in the Queule River Estuary, South-Central Chile. Estuaries Coasts 2001, 24, 69–77. [Google Scholar] [CrossRef]
- Walther, A.C.; Burch, J.B.; Foighil, D.Ó. Molecular phylogenetic revision of the freshwater limpet genus Ferrissia (Planorbidae: Ancylinae) in North America yields two species: Ferrissia (Ferrissia) rivularis and Ferrissia (Kincaidilla) fragilis. Malacologia 2010, 53, 25–45. [Google Scholar] [CrossRef]
- Marrone, F.; Lo Bruto, S.; Arculeo, M. Cryptic invasion in Southern Europe: The case of Ferrissia fragilis (Pulmonata: Ancylidae) Mediterranean populations. Biologia 2011, 663, 484–490. [Google Scholar] [CrossRef] [Green Version]
- Vecchioni, L.; Marrone, F.; Arculeo, M.; Arizza, V. Are there autochthonous Ferrissia (Mollusca, Planorbidae) in the Palaearctic? Molecular evidence of a widespread North American invasion of the Old World. Eur. Zool. J. 2017, 84, 411–419. [Google Scholar] [CrossRef]
- Vinarski, M.V. The history of an invasion: Phases of the explosive spread of the physid snail Physella acuta through Europe, Transcaucasia and Central Asia. Biol. Invasions 2017, 19, 1299–1314. [Google Scholar] [CrossRef]
- Spyra, A.; Cieplok, A.; Strzelec, M.; Babczyńska, A. Freshwater alien species Physella acuta (Draparnaud, 1805)-A possible model for bioaccumulation of heavy metals. Ecotoxicol. Environ. Saf. 2019, 185, 109703. [Google Scholar] [CrossRef]
- Cowie, R. Invertebrate invasions on Pacific Islands and the replacement of unique native faunas: A synthesis of the land and freshwater snails. Biol. Invasions 2001, 3, 119–136. [Google Scholar] [CrossRef]
- Strayer, D.L. Alien species in fresh waters: Ecological effects, interactions with other stressors, and prospects for the future. Freshw. Biol. 2010, 55, 152–174. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N.; Pronin, M. Diversity and zooplankton species associated with certain hydroperiods and fish state in field ponds. Ecol. Indic. 2018, 90, 171–178. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
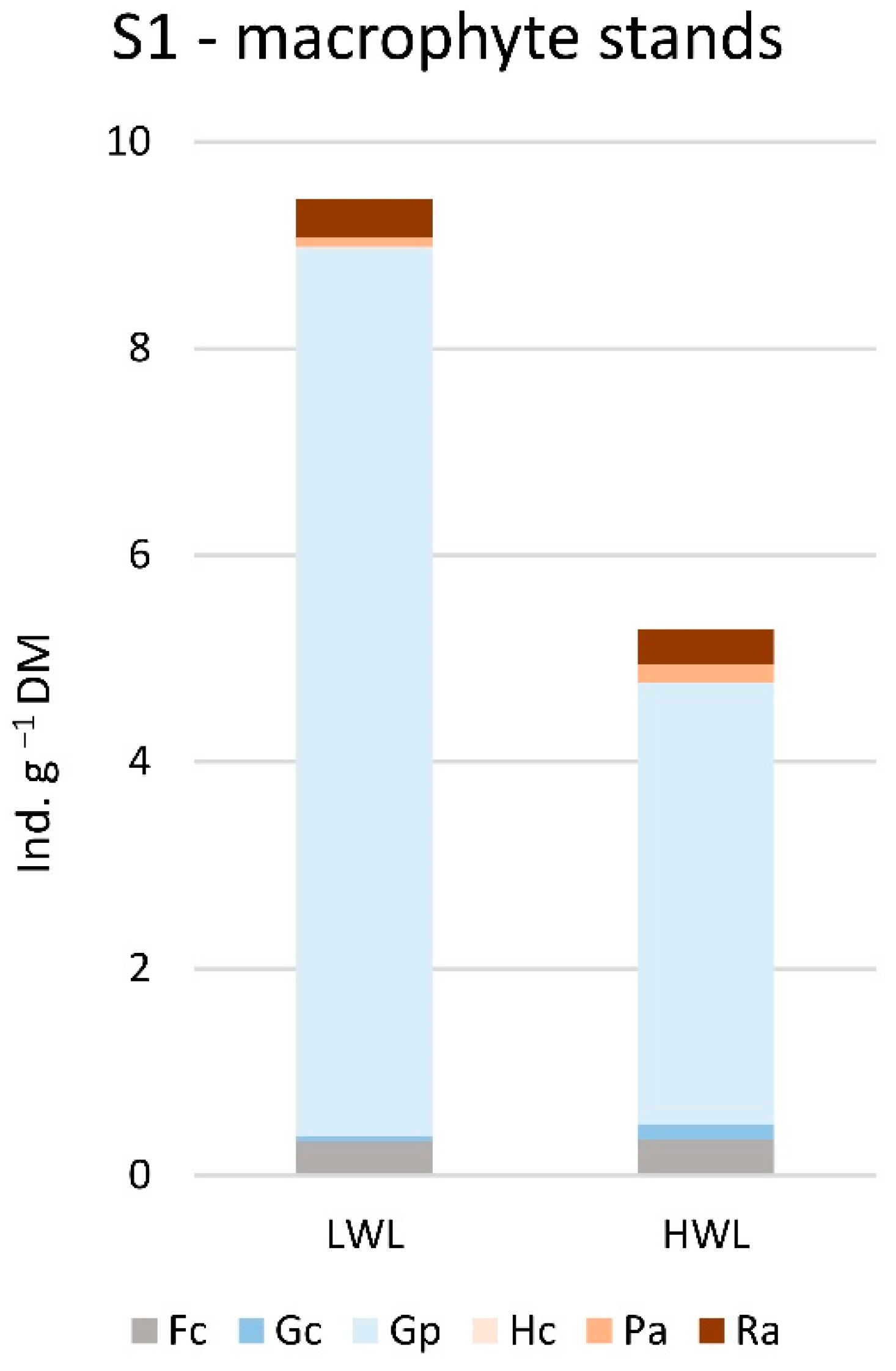{kind=link}
{kind=link}
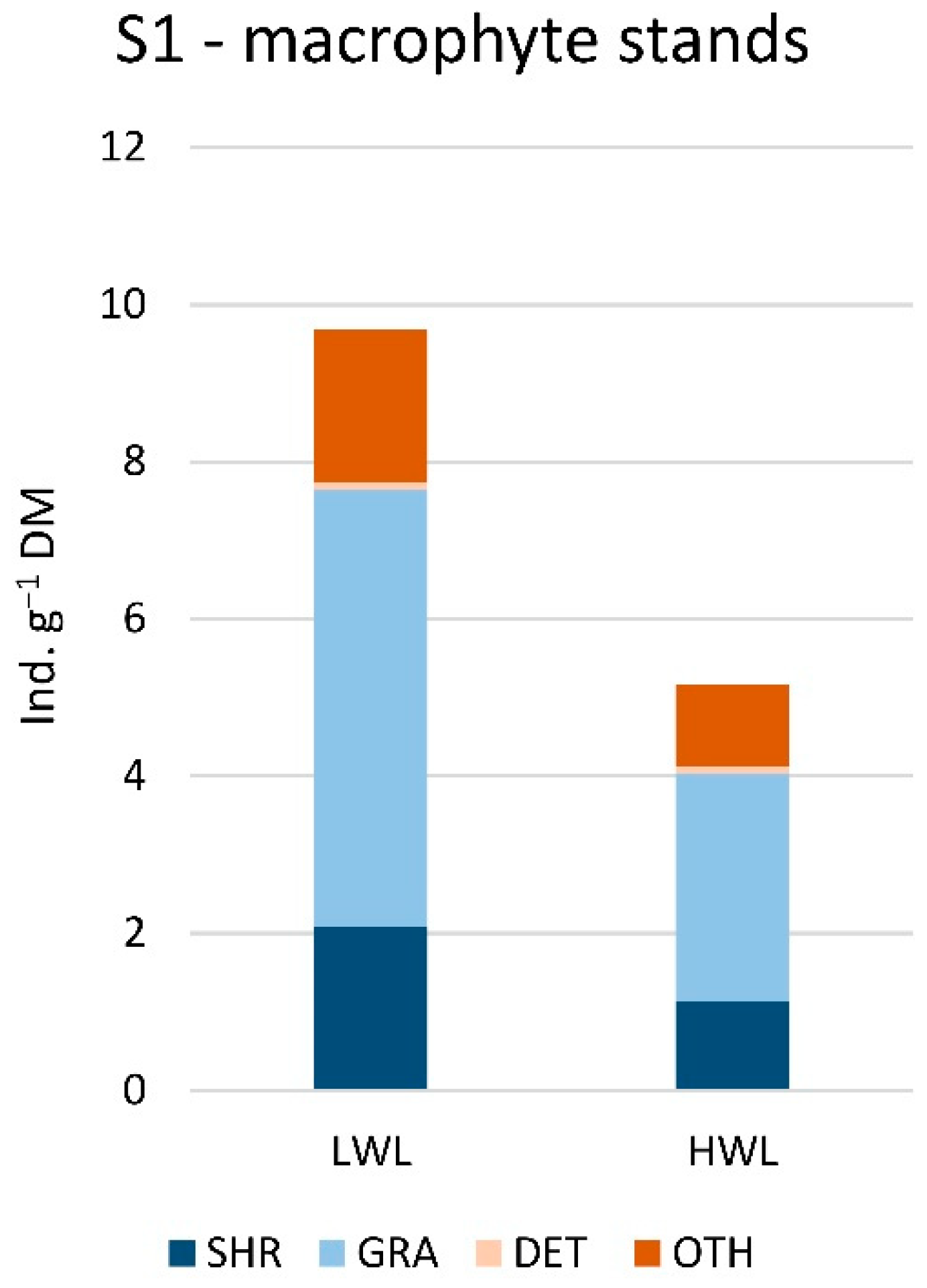{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean ± SD | ||
|---|---|---|
| Variable | LWL | HWL |
| Temperature (°C) | 23.2 ± 4.8 | 19.7 ± 3.0 |
| DO (mg O2 L−1) | 6.8 ± 2.7 | 5.9 ± 1.3 |
| pH | 8.33 ± 0.70 | 7.95 ± 0.35 |
| EC (µS cm−1) | 485.8 ± 159.5 | 430.5 ± 124.3 |
| DOM (mg O2(Mn) L−1) | 17.0 ± 3.8 | 21.5 ± 5.6 |
| POM (µg L−1 (×10−3)) | 6.395 ± 6.215 | 13.503 ± 18.404 |
| TN (mg N L −1) | 0.896 ± 0,327 | 0.399 ± 0.113 |
| TP (mg P L−1) | 0.362 ± 0.086 | 0.318 ± 0.026 |
| Chl a (µg L−1) | 27.528 ± 18.549 | 26.668 ± 34.119 |
| Species | Av. Ab. | Av. Sim. | Contrib. (%) |
|---|---|---|---|
| LWL | |||
| Gyraulus parvus | 2.04 | 5.72 | 52.8 |
| Radix auricularia | 1.23 | 1.45 | 13.35 |
| Ferrissia californica | 1.27 | 1.42 | 13.12 |
| Physella acuta | 0.9 | 1.26 | 11.62 |
| HWL | |||
| Gyraulus parvus | 3.02 | 7.16 | 32.21 |
| Physella acuta | 1.64 | 5.17 | 23.24 |
| Hippeutis complanatus | 1.31 | 3.5 | 15.75 |
| Ferrissia californica | 1.69 | 3.22 | 14.47 |
| Radix auricularia | 1.1 | 3.18 | 14.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lajtner, J.; Kozak, A.; Špoljar, M.; Kuczyńska-Kippen, N.; Dražina, T.; Sertić Perić, M.; Tkalčec, I.; Gottstein, S.; Zrinščak, I. Gastropod Assemblages Associated with Habitat Heterogeneity and Hydrological Shifts in Two Shallow Waterbodies. Water 2022, 14, 2290. https://doi.org/10.3390/w14152290
Lajtner J, Kozak A, Špoljar M, Kuczyńska-Kippen N, Dražina T, Sertić Perić M, Tkalčec I, Gottstein S, Zrinščak I. Gastropod Assemblages Associated with Habitat Heterogeneity and Hydrological Shifts in Two Shallow Waterbodies. Water. 2022; 14(15):2290. https://doi.org/10.3390/w14152290
Chicago/Turabian StyleLajtner, Jasna, Anna Kozak, Maria Špoljar, Natalia Kuczyńska-Kippen, Tvrtko Dražina, Mirela Sertić Perić, Ines Tkalčec, Sanja Gottstein, and Ivana Zrinščak. 2022. "Gastropod Assemblages Associated with Habitat Heterogeneity and Hydrological Shifts in Two Shallow Waterbodies" Water 14, no. 15: 2290. https://doi.org/10.3390/w14152290