
Species Diversity, Distribution, and Abundance of Freshwater Snails in KwaZulu-Natal, South Africa
by
 , ,
, ,
Onyekachi Esther Nwoko
1,* ,
,
Chester Kalinda
1,2,3,
Tawanda Manyangadze
1,4 and
Moses John Chimbari
1,5 1
Discipline of Public Health Medicine, College of Health Sciences, University of KwaZulu-Natal, Durban 4000, South Africa
2
Bill and Joyce Cummings Institute of Global Health, University of Global Health Equity (UGHE), P.O. Box 6955, Kigali 20093, Rwanda
3
Institute of Global Health Equity Research (IGHER), University of Global Health Equity (UGHE), P.O. Box 6955, Kigali 20093, Rwanda
4
Department of Geosciences, School of Geosciences, Disasters and Development, Faculty of Science and Engineering, Bindura University of Science Education, Bindura P.O. Box 1020, Zimbabwe
5
Department of Behavioural Science, Medical and Health Sciences, Great Zimbabwe University, Masvingo P.O. Box 1235, Zimbabwe
*
Author to whom correspondence should be addressed.
Water 2022, 14(14), 2267; https://doi.org/10.3390/w14142267
Submission received: 9 May 2022
/
Revised: 27 June 2022
/
Accepted: 7 July 2022
/
Published: 21 July 2022
(This article belongs to the Section Biodiversity and Functionality of Aquatic Ecosystems)
Abstract
:Disease distribution is correlated to the distribution of the freshwater snails which in turn is influenced by the physicochemical status of the habitats. This study aimed to evaluate freshwater snail species diversity, abundance, and distribution in KwaZulu-Natal (KZN) province, South Africa, between December 2020 to February 2021. A total of 4576 freshwater snails consisting of 8 species were collected from 127 sites in 11 districts. Tarebia granifera snails were the most abundant (n = 2201), while bivalves (n = 95) were the least abundant. The highest and least Shannon–Weiner Simpson’s diversity indices were recorded in Ugu and iLembe districts, respectively. A negative relationship was observed between rainfall, Bulinus tropicus, Lymnaea natalensis, bivalves, and Physa acuta, while temperature had a positive relationship with B. globosus, B. pfeifferi, and T. granifera. A positive relationship was observed between B. globosus and B. pfeifferi (r = 0.713, p < 0.05). Snail presence constitutes potential health and economic risks to humans and animals in contact with the waterbody. Hence, our study described the current distribution, abundance, and species diversity of freshwater snails in the KZN province with insights into the possibilities of snail-based biological control for schistosomes intermediate host snails.
1. Introduction
Biodiversity is a key indicator of the health of any ecosystem as it describes the variation attainable in the ecosystem in terms of resources and species. Five factors have been identified as threats to global freshwater biodiversity: overexploitation, water pollution, flow modification, destruction or degradation of habitat, and invasion by exotic species with environmental changes superimposed on these factors [1]. A more diverse ecosystem can recover from ecological threats [2]. Species diversity refers to the number of different species present in an ecosystem and the relative abundance of each species [3]. An ecosystem with poor species diversity may not function properly or efficiently. There are two indices that measure species diversity: species richness and species evenness. Species richness is the number of different species in an ecosystem, while species evenness is the variation in the abundance of individuals per species [4]. An ecosystem has low species evenness when there are large disparities in the number of individuals within each species, whereas a high species evenness is when the number of individuals within a species is constant. The more even the number of animals per species within an ecosystem, the greater the species diversity. It is important to consider species richness and diversity because each species plays an important role and boosts ecosystem productivity.
Freshwater snails belong to the class Gastropoda, which is found in every continent excluding the Antarctic and in nearly all aquatic habitats, including rivers, lakes, streams, swamps, underground aquifers and springs, ponds, drainages, ditches, and other ephemeral and seasonal waterbodies [5]. Freshwater gastropods are classified into three major subclasses: Prosobranchia (with gills in front of the heart), Opisthobranchia (with gills to the right and behind the heart), and Pulmonata (with a lung instead of gills) [6]. There are approximately 4000 freshwater snail species and 57 extinct species globally. In Africa, there are 400 species of freshwater snails with 51 gastropod species: 43 indigenous and 8 exotic species from Southern Africa [7]. Freshwater snails play a significant role in public and veterinary health as some serve as intermediate hosts of blood fluke trematodes and nematodes [8].
Studies in Zimbabwe have investigated Bulinus tropicus as a potential competitor snail for Bulinus globosus [9,10]. However, Ndlela and Madsen [11] showed that B. tropicus did not have a significant effect on the density of B. globosus, suggesting little or no competition between the two species. Perissinotto et al. [12] described Tarebia granifera as a non-native invasive species with a high abundance that poses threats to indigenous gastropods species abundance and richness such as Bulinus spp. Although the distribution and abundance of some freshwater snails like B. globosus [13], Biomphalaria pfeifferi [14], Lymnaea natalensis [15], B. tropicus [16,17], Bulinus forskalii [18], Physa acuta [19], and T. granifera [12] have been studied in South Africa, the knowledge of total diversity of freshwater snails is still incomplete in KZN. This is because most of the studies were conducted in a specific district and did not extend to the entire province. In addition, interests of previous researchers was restricted to specific snail species.
The mechanism and outcome of the interaction between non-schistosome intermediate host snails and schistosome intermediate host snails in KZN are not well understood. Hence, we conducted a comprehensive survey of freshwater snails in KwaZulu-Natal (KZN) province, South Africa, to (1) assess the distribution, abundance, and species diversity of freshwater snails (2) examine the effects of physicochemical parameters and climatic factors on the distribution, abundance, and species diversity of freshwater snails; and (3) determine the interactions between schistosome and non-schistosome intermediate host snails. The following research hypothesis was formulated: there is a relationship between snail species diversity, distribution, abundance, and physicochemical parameters in water bodies. The findings from this study are of interest as they will form the basis for developing biological control strategies against schistosomiasis intermediate host snails.
2. Materials and Methods
2.1. Study Area
The study was conducted in KwaZulu-Natal (KZN) province. KZN (geographical coordinates 2, ) is one of the nine provinces in South Africa and has a surface area of . It is located on the eastern seaboard of South Africa. The province is divided into one metropolitan municipality (eThekwini) and 10 district municipalities. KZN has an estimated total population of 10.3 million [20]. The province experiences four seasons: rainy (December–February), post-rainy (March–May), cold/dry (June–August), and hot/dry (September–November) [13]. The topography in the province varies from 3448 m above mean sea level in the interior and drops to sea level at the coast. The mean temperatures rise above 25 °C from September to February and drop to 20 °C from March to August. Annual mean precipitation ranges from about 964–1382 mm/year around the coastal areas of Durban, Richards Bay, and Margate to 813–847 mm/year around midland areas of Pietermaritzburg and Newcastle [21] (Figure 1).
2.2. Snail Sampling
A cross-sectional snail sampling survey was conducted in all the districts of KZN province between December 2020 and February 2021. Each site was sampled once within three months. The sampling sites per district are presented in Table 1. Snail sampling was carried out by 2 trained research assistants using a metal scoop complemented by hand picking for 15 min per collection site [22]. The collected snails were put in vials containing water from the sites and transported to the place of processing. The freshwater snails were rinsed off and identified morphologically to the species level using the identification key developed by Brown and Kristensen [23].
2.3. Physicochemical Parameters and Climatic Factors
Physicochemical parameters of the water were measured at all the sites simultaneously with snail samplings. The parameters measured included pH, salinity (psu), pressure (psi), electrical conductivity (EC) (, and dissolved oxygen (DO) (ppm) using a portable multiparameter meter (Hanna Instruments, HI9829, Woonsocket, Rhode Island, USA). These physicochemical parameters were chosen because they are known to affect snail abundance, distribution, and diversity [24]. Climatic data were obtained through remote sensing. A 3-month average (December 2020–February 2021) for maximum land surface temperature (LST), minimum land surface temperature, Normalized Difference Vegetation Index (NDVI), Normalized Difference Water Index (NDWI), Enhanced Vegetation Index (EVI), and precipitation were used [13,25]. All the datasets excluding LST were accessed between 10 and 20 July 2021 through the International Research Institute for Climate and Society (IRI) data library portal (http://iridl.ldeo.columbia.edu/SOURCES/). LST data set was accessed between 27 and 30 July 2021 from Climate Engine: Cloud Computing of Climate and Remote Sensing Data (http://climateengine.org).
2.4. Data Analysis
All collected data were entered into an Excel spreadsheet and statistical analyses were carried out using R software (version 4.1.2; R studio Team, 2021). The distribution and abundance of snails according to snail species and districts in KZN province were presented using descriptive analysis with proportions.
Five biodiversity indices were used to evaluate species diversity, richness, and evenness. Margalef’s index (d) was used to evaluate species richness [26]. The Shannon–Wiener diversity [26] and Simpson’s [27] indices were used to assess the species diversity of freshwater snails. Pielou’s evenness index ) was used to evaluate species evenness. Its values are between 0 and 1; values closer to 0 represented uneven populations that were dominated by one species, while values closer to 1 represented even populations comprising several species with similar abundances [26]. An effective number of species (ENS) is the number of equally abundant species necessary to produce the observed value of diversity. The value of ENS is equal to species richness in an even community but lesser than species richness in an uneven community [28].
Spearman’s correlation was used to determine the effects of physicochemical factors and climatic factors on the distribution, abundance, and species diversity of freshwater snails. In addition, interactions between snail species were accessed using Spearman’s correlation [29].
3. Results
3.1. Snail Distribution and Abundance
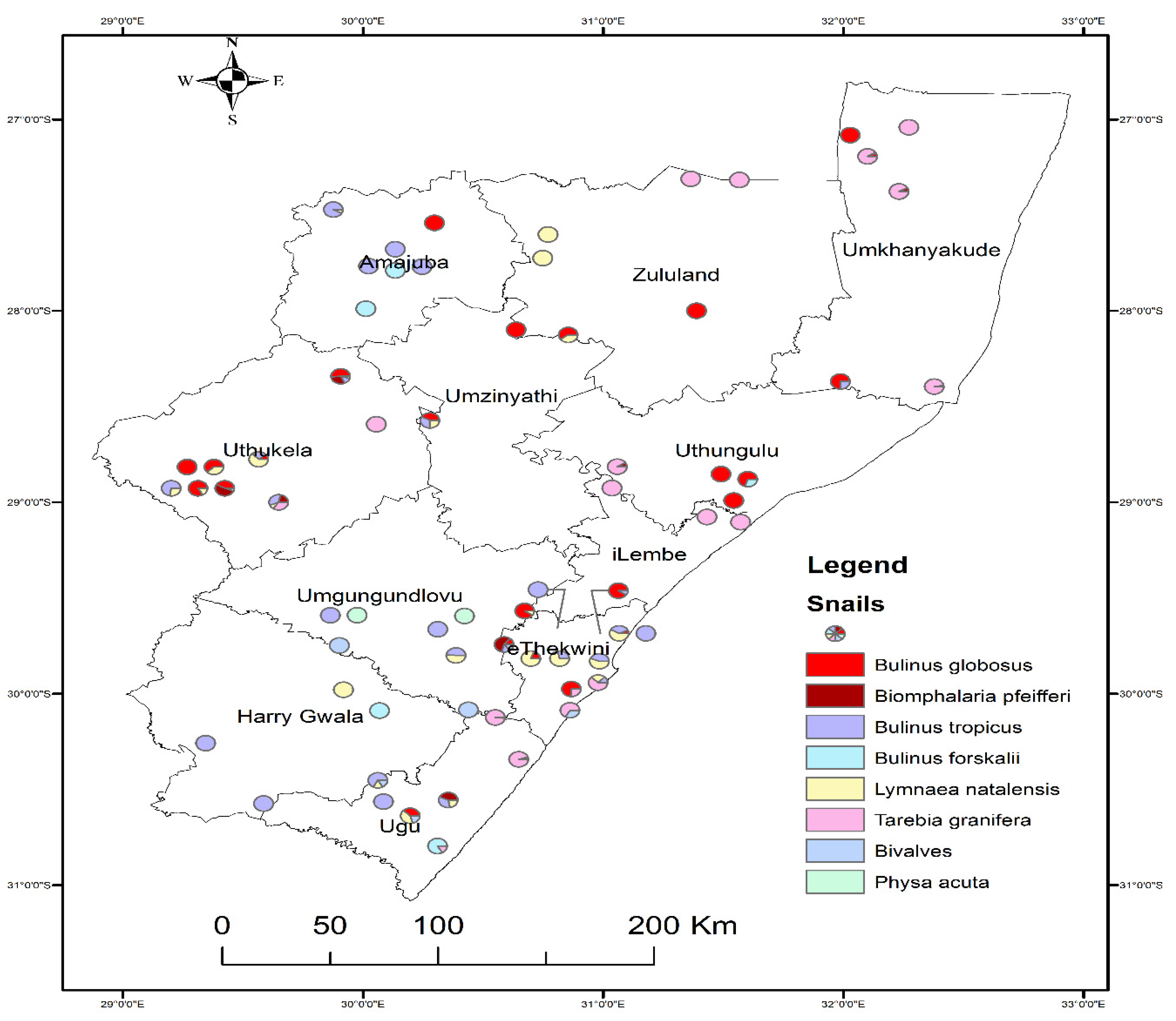
A total of 4576 freshwater snails were collected from 127 different sampling sites in 11 districts. Based on morphological identification, the eight freshwater snail species collected are Biomphalaria pfeifferi, Bulinus globosus, Lymnaea natalensis, Tarebia granifera, Bulinus tropicus, Bulinus forskalii, bivalves, and Physa acuta (Table 1 and Figure S1). T. granifera was the most abundant (n = 2201, 48.1%) and bivalves was least abundant (n = 95, 2.1%). B. globosus (n = 488, 10.7%) was more abundant compared to B. pfeifferi (n = 246, 5.4%). B. globosus snails were found in 9 districts in the province, while B. tropicus and L. natalensis were found in 8 districts in the province. Furthermore, T. granifera was found in 7 districts, B. pfeifferi in 5 districts, B. forskalii and bivalves in 3 districts, while P. acuta was found in two districts in the province (Table 1).
3.2. Snail Diversity, Richness, and Evenness
The Shannon–Weiner diversity index (H′) of freshwater snails in KZN ranged from 0.10 to 1.63. Ugu district recorded the highest species diversity (H′ = 1.63 and D = 0.74) while the lowest species diversity was recorded in iLembe district (H′ = 0.10 and D = 0.04). Snail abundances were found to be less even among the different snail species in iLembe district (J′ = 0.14), while there were similar abundances amongst snail species in uThukela district (J′ = 0.88). The value for ENS in all the districts is lower than the corresponding number of species found in each district which shows species unevenness (Table 2). Rainfall and temperature were identified as climatic factors that affect snail diversity negatively from correlation analysis (Table S1).
3.3. Interactions between Snail Species
Table 3 presents the interaction between different snail species. A positive correlation was recorded between B. globosus and B. pfeifferi (r = 0.713). In addition, B. tropicus was positively correlated with L. natalensis (r = 0.840), bivalves (r = 0.73) and P. acuta (r = 0.718). Furthermore, L. natalensis correlated positively with bivalves (r = 0.6794) and P. acuta (r = 0.784). Finally, bivalves had a positive correction with P. acuta (r = 0.593).
3.4. Relationship between Snail Abundance, Physicochemical Parameters, and Environmental Factors
The values of the physicochemical parameters measured during the study are summarized in Table 4. The mean (±SD) water pH values varied from 6.42 ± 1.04 to 7.98 ± 0.55. DO varied across the different districts over the sampling period, with the lowest DO value recorded in the Zululand district (4.33 ± 3.64) ppm, and the highest value was recorded Ugu district (9.40 ± 10.27).
The relationship between physicochemical parameters, environmental factors, and snail abundance are presented in Table 5. A positive correlation was observed between pH and abundance of B. globosus (r = 0.304) and T. granifera (r = 0.356), while a negative relationship was observed between pH and B. tropicus, B. forskalii, L. natalensis, bivalves, and P. acuta. DO showed a positive correlation with B. tropicus (r = 0.553), B. forskalii (r = 0.437), L. natalensis (r = 0.3415), bivalves (r = 0.252) and P. acuta (r = 0.589) but a negative correlation with B. globosus (r = −0.249). Salinity was positively correlated with B. globosus (r = 0.347), B. pfeifferi (r = 0.3564), B. forskalii (r = 0.587) and T. granifera (r = 0.338). Rainfall was negative correlated with B. tropicus (r = −0.7615), L. natalensis (r = −0.8047), bivalves (r = −0.8476) and P. acuta (r = −0.6706). Maximum and minimum land surface temperatures (LST) were positively correlated with B. globosus, B. pfeifferi, T. granifera, and negatively correlated with B. tropicus, B. forskalii, L. natalensis, and P. acuta (Table 5).
4. Discussion
Our study presents recent species diversity, distribution, and abundance of freshwater snails in 11 districts in KZN province, South Africa. A total of 4576 freshwater snails were collected from 127 sites, of which 8 species were identified morphologically. They were B. globosus, B. pfeifferi, B. tropicus, B. forskalii, L. natalensis, T. granifera, bivalves, and P. acuta. Perissinotto, Miranda, Raw and Peer [12] recorded 12 snail species in the biodiversity census performed in Lake St. Lucia, iSimangaliso Wetland Park in 2012 and 2013, which is more than the snail species identified in our study. Possible exclusion of some snail species could be due to competition with alien invasive snails, different sampling methods (the sites included in our study were selected randomly while Perissinotto, Miranda, Raw and Peer [12] sampled on an opportunistic basis), and snail sampling season (our data was collected in the rainy season; hence, some snails might have been washed off due to high velocities of flowing water and the foci nature of freshwater snails) may be some reasons for the differences in the snail species recorded [14,30].
Bivalves had the lowest distribution and abundance (n = 95) in the KZN province. This finding is consistent with the study by Sharma et al. [31] in which gastropods were widely distributed and more abundant than bivalves. This could be attributed to the ability of gastropods to tolerate and adapt to various environmental conditions compared to bivalves [32]. B. globosus was more abundant (n = 488) and recorded in 9 districts compared to B. pfeifferi (n = 246) recorded in 5 districts. This finding contradicts that of Manyangadze, Chimbari, Rubaba, Soko and Mukaratirwa [14] carried out in uMkhanyakude, one of the districts in KZN province where the abundance of B. pfeifferi was more than B. globosus. The difference could be due to the different scales or coverage of the study areas. Manyangadze, Chimbari, Rubaba, Soko and Mukaratirwa [14] study was carried out on a micro-scale, not even at the district level. Furthermore, the current study was cross-sectional in the rainy season only, while the former was a longitudinal study for one year with data for all the months and seasons.
The highest Shannon–Weiner diversity index (H′), Simpson’s index (D), and Margalef’s index (d) were recorded in the Ugu district, where eight freshwater snail species were identified, and iLembe district recorded the least, where two freshwater snails were identified. The difference in the number of freshwater snails recorded could be because the physicochemical parameters and climatic factors are within the range that is favorable for freshwater snails to inhabit in Ugu district compared to the iLembe district. Although the Ugu district recorded the highest species diversity, Pieolu’s evenness index recorded similar abundances amongst snail species in the uThukela district. This could be because of T. granifera’s ability to reach high densities, making the ecosystem uneven and unstable. The entire freshwater snail community was uneven and is evident in T. granifera, accounting for the largest proportion (48.1%) of snails collected and that could be explained by the high level of tolerance possessed by non-native invasive snails. Rainfall and temperature were identified as key climatic factors affecting snail diversity negatively.
During the 1990s aquarium trade, T. granifera may have been introduced into KwaZulu-Natal but was discovered in the St. Lucia estuary in 2005 for the first time [33]. However, this has changed as T. granifera has invaded different water bodies in the northeastern areas of South Africa [33]. In our study, T. granifera was found to be the snail with the highest abundance (n = 2201). Our finding is consistent with those of Miranda and Perissinotto [34]; and Perissinotto, Miranda, Raw and Peer [12] who showed that T. granifera is known to attain high densities and is capable of causing an ecological disturbance and displacing native gastropods like Bulinus spp. Sisonke district (0.06 ± 0.03 psu) recorded the lowest salinity, while the highest salinity level was found in the Ugu district (13.12 ± 38.82 psu). Both values are suitable for T. granifera’s growth, reproduction, survival, and distribution patterns. According to Miranda et al. [35], T. granifera survives and reproduces at salinities up to 20 psu, but its activity declines with salinities from 30 psu to 40 psu and is adversely affected with salinities above 40 psu. A positive correlation was observed between salinity and B. globosus, B. pfeifferi, B. forskalii, and T. granifera.
Rainfall was negatively correlated with B. pfeifferi, B. tropicus, L. natalensis, bivalves, and P. acuta. This could be because there were heavy rains prior to sampling and hence floods or fast flowing water may have swept away many freshwater snails and food causing major fluctuations in the population density [36]. In addition, snail abundance and diversity reduce with high rainfall as snails cannot attach themselves to rocks or water vegetation and are washed away by the high velocities (>0.3 ms−1) of flowing water [37,38]. On the other hand, B. globosus, B. forskalii, and T. granifera showed a positive correlation, and this could be because of snail abundance and diversity increase with low rainfall or at the beginning of the rainy season [37,38]. This is consistent with literature showing that B. globosus is positively correlated with rainfall [39].
Freshwater snails prefer habitats with pH levels ranging from 6.5–8.5 as extreme pH levels increase the solubility of elements and compounds making toxic chemicals more mobile and increasing the risk of absorption by aquatic life and leading to death. In this study, snails were found in sites with pH levels ranging from 6.42–7.98, which falls within the favorable range for surface water [38]. A significant positive correlation was recorded between pH, B. globosus, and T. granifera, while a negative correlation was recorded with B. tropicus, B. forskalii, L. natalensis, bivalves, and P. acuta. These findings are consistent with other studies where some freshwater snails were found to be positively correlated with snail abundance while others were negatively correlated [32]. However, Ofoezie [39] and Cañete et al. [40] opined that pH might not be an influencing factor in snail abundance.
Temperature has been identified as an important factor that has direct effect on freshwater snail distribution and abundance [41]. Air temperature range of 20 °C–25 °C was recorded during the study period in Amajuba and uMgungundlovu districts, respectively. Studies have shown that extreme temperatures may be unsuitable for the development of snails. Kalinda et al. [42] reported that the growth rate of snails is slowed down at 15.5 °C and 31.0 °C, while Woolhouse and Chandiwana [43] observed that a temperature above 24 °C results in increased snail mortality. Maximum and minimum LST had positive correlations with B. globosus, B. pfeifferi, T. granifera, and negative correlations with B. tropicus, B. forskalii, L. natalensis, and P. acuta. However, minimum LST showed a negative effect on T. granifera, B. tropicus, and L. natalensis.
Snail distribution and abundance may be limited by low dissolved oxygen [36]. In our study, DO varied between the districts. Zululand district had the smallest mean DO value (4.33 ± 3.64 ppm), while the Ugu district recorded the highest mean DO value (9.40 ± 10.27 ppm). These values fall within the desirable range (0.4 ppm–16.0 ppm) of DO for freshwater snails [44]. Unfavorable conditions such as pollution and densely packaged floating vegetation are known to lead to a shortage of DO that could lead to the death of freshwater snails [36]. The low DO level recorded in the Zululand district could be ascribed to low water quality resulting from probable increased organic decomposition activities due to the death and decay of aquatic macrophytes [44,45]. In this study, a positive relationship was recorded between dissolved oxygen and some freshwater snails and that corroborates the reported findings [45,46]. However, a negative relationship was observed between DO and B. globosus. This result contradicts that of Brown [36] where B. globosus showed a preference for high oxygen concentration as its mobility is reduced in an environment with low oxygen concentration leading to impaired feeding and reproduction.
Interactions between different freshwater snails differed, with 5 districts out of 11 districts having both B. globosus and B. pfeifferi. B. globosus had a positive correlation with B. pfeifferi. This agrees with findings from Manyangadze, Chimbari, Gebreslasie, Ceccato and Mukaratirwa [13] and shows evidence of cohabitation between B. globosus and B. pfeifferi. T. granifera had a negative correlation with all snail species, excluding B. pfeifferi and B. globosus. These results support the work by Miranda and Perissinotto [34] and Perissinotto, Miranda, Raw and Peer [12] where the increasing abundance of T. granifera might affect indigenous species like Bulinus spp. In contrast, the positive correlation between B. pfeifferi, B. globosus, and T. granifera could be because the data collected is a snapshot of snail distribution, and this could change if data from different seasons were considered. Further experimental and field studies should be carried out to verify the use of T. granifera as a biological control agent for schistosomiasis in South Africa. In the Caribbean, T. granifera was introduced into waterbodies to compete and displace the native intermediate host snail Biomphalaria glabrata, responsible for Schistosoma mansoni [47,48,49]. A negative correlation was observed between B. tropicus and B. globosus. This result supports the findings of Ndlela and Madsen [11] and Ndlela, Chimbari and Madsen [10] and suggests no competition between both species. A non-significant negative correlation was observed between B. globosus and the invasive P. acuta. This finding stirs up some curiosity as research has shown that P. acuta has higher fecundity, shorter hatching time, higher salinity tolerance, higher temperature tolerance, and tolerates fast current velocities up to 0.6 ms−1 compared to other pulmonates that are not found in water exceeding the velocity of 0.3 ms−1 [50]. P. acuta responds to disturbances to its habitats, like floods, flow regulation, and pollution, by reproducing very fast and exploiting new environments better than the native B. tropicus [19]. However, our results showed a positive correlation between B. tropicus and P. acuta. In South Australia and New Zealand, P. acuta was found to outcompete the native species Glyptophysa gibbosa and Physastra variabilis (syn. Glyptophysa variabilis), respectively [51].
Although snail sampling was carried out over a large geographical range across KZN, snail sampling in the sites was performed once over three months (December 2020–February 2021). More sustained sampling to account for the four seasons experienced in KZN should be considered in future studies to have a better understanding of freshwater snail diversity, distribution, and abundance and how it is affected by the different seasons. Experimental studies are also needed to investigate further the relationship between schistosome intermediate host snails and non-schistosome intermediate host snails, as there is potential to use non-schistosome intermediate host snails as a biological control for schistosomiasis.
5. Conclusions
The study revealed the species unevenness in the molluscan community in KZN, with T. granifera accounting for the largest proportion (48.1%) of snails collected out of the eight snail species recorded. The presence of freshwater snails, known to be the intermediate host of trematode flukes such as schistosomiasis and fascioliasis, amongst others, constitute potential economic and health risks to humans and animals in contact with the water bodies. The status of physicochemical parameters and climatic factors in the water body is favorable for the species diversity, distribution, and abundance, which may support an increase in their population. Salinity, dissolved oxygen temperature, rainfall, and pH were identified as physicochemical parameters that influence snail distribution, abundance, and diversity. Interactions between snails also reveal the potential of using T. granifera and P. acuta as a biological control against potential intermediate hosts of schistosomes.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w14142267/s1, Table S1: Correlation coefficient between physicochemical parameters and species diversity indices; Table S2: Correlation coefficient between snail species and species diversity indices. Figure S1: Snails species collected and identified from different waterbodies in KwaZulu-Natal, South Africa.
Author Contributions
Conceptualization, O.E.N., T.M., C.K., and M.J.C.; methodology, O.E.N., T.M., C.K., and M.J.C.; formal analysis, O.E.N., T.M., C.K., and M.J.C.; data curation, O.E.N., T.M., and C.K.; writing—original draft preparation, O.E.N.; writing—review and editing, O.E.N., T.M., C.K., and M.J.C.; supervision, M.J.C.; funding acquisition, M.J.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Tackling Infections to Benefit Africa (TIBA) through grant UoERef:CT-4987 (d) and the University of KwaZulu-Natal through a Ph.D. studentship bursary awarded to O.E.N by the College of Health Sciences. The views expressed in this publication are those of the author(s) and not necessarily those of the TIBA.
Institutional Review Board Statement
The University of KwaZulu Natal biomedical research ethics committee (BREC) issued the ethical approval (Ref No: BREC/00001305/2020). This review is part of the approved thesis protocol.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
IREX funding through the UASP alumni small grants program is acknowledge for providing a platform for the development of this publication. The authors appreciate the efforts of the TIBA research team and community research assistants that assisted in the data collection process. Thanks to Nokwanda Majola and Ntombela Sambulo for the administrative and technical support provided.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study, in the collection, analyses, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef]
- Revenga, C.; Campbell, I.; Abell, R.; De Villiers, P.; Bryer, M. Prospects for monitoring freshwater ecosystems towards the 2010 targets. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 397–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiarucci, A.; Bacaro, G.; Scheiner, S.M. Old and new challenges in using species diversity for assessing biodiversity. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2426–2437. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, R.J.; Willis, K.J.; Field, R. Scale and species richness: Towards a general, hierarchical theory of species diversity. J. Biogeogr. 2001, 28, 453–470. [Google Scholar] [CrossRef] [Green Version]
- Strong, E.E.; Gargominy, O.; Ponder, W.F.; Bouchet, P. Global diversity of gastropods (Gastropoda; Mollusca) in freshwater. In Freshwater Animal Diversity Assessment; Springer: Berlin/Heidelberg, Germany, 2007; pp. 149–166. [Google Scholar]
- Hamli, H.; Azmai, S.S.; Hamed, S.A. Diversity and habitat characteristics of local freshwater Gastropoda (Caenogastropoda) from Sarawak, Malaysia. Singap. J. Sci. Res. 2020, 10, 23–27. [Google Scholar] [CrossRef]
- Pyron, M.; Brown, K.M. Introduction to mollusca and the class Gastropoda. In Thorp and Covich’s Freshwater Invertebrates; Elsevier: Amsterdam, The Netherlands, 2015; pp. 383–421. [Google Scholar]
- Madsen, H.; Hung, N. An overview of freshwater snails in Asia with main focus on Vietnam. Acta Trop. 2015, 141, 372–384. [Google Scholar] [CrossRef]
- Chimbari, M.J. Enhancing schistosomiasis control strategy for Zimbabwe: Building on past experiences. J. Parasitol. Res. 2012, 2012, 353768. [Google Scholar] [CrossRef]
- Ndlela, B.; Chimbari, M.J.; Madsen, H. Interactions between Bulinus globosus and B. tropicus (Gastropoda: Planorbidae) in a pond experiment in Zimbabwe. Afr. J. Aquat. Sci. 2007, 32, 13–16. [Google Scholar] [CrossRef]
- Ndlela, B.; Madsen, H. Laboratory and quasi-field studies on interspecific competition between Bulinus globosus and B. tropicus (Gastropoda: Planorbidae). South. Afr. J. Aquat. Sci. 2001, 26, 17–21. [Google Scholar] [CrossRef]
- Perissinotto, R.; Miranda, N.A.; Raw, J.L.; Peer, N. Biodiversity census of lake St Lucia, iSimangaliso wetland park (South Africa): Gastropod molluscs. ZooKeys 2014, 440, 1–43. [Google Scholar] [CrossRef]
- Manyangadze, T.; Chimbari, M.J.; Gebreslasie, M.; Ceccato, P.; Mukaratirwa, S. Modelling the spatial and seasonal distribution of suitable habitats of schistosomiasis intermediate host snails using Maxent in Ndumo area, KwaZulu-Natal Province, South Africa. Parasites Vectors 2016, 9, 572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manyangadze, T.; Chimbari, M.J.; Rubaba, O.; Soko, W.; Mukaratirwa, S. Spatial and seasonal distribution of Bulinus globosus and Biomphalaria pfeifferi in Ingwavuma, uMkhanyakude district, KwaZulu-Natal, South Africa: Implications for schistosomiasis transmission at micro-geographical scale. Parasites Vectors 2021, 14, 222. [Google Scholar] [CrossRef] [PubMed]
- Moema, E.B.E.; King, P.H.; Baker, C. Cercariae developing in Lymnaea natalensis Krauss, 1848 collected in the vicinity of Pretoria, Gauteng Province, South Africa. Onderstepoort J. Vet. Res. 2008, 75, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, P.H.; Van As, J. Cercariae shed by Bulinus tropicus (Krauss, 1848) in the Free State, South Africa. Afr. Zool. 2001, 36, 95–105. [Google Scholar] [CrossRef]
- Van Eeden, J.; Combrinck, C. Distributional trends of four species of freshwater snails in South Africa with special reference to the intermediate hosts of bilharzia. Afr. Zool. 1966, 2, 95–109. [Google Scholar] [CrossRef] [Green Version]
- De Kock, K.; Wolmarans, C. Distribution, habitats and role as intermediate host of the freshwater snail, Bulinus forskalii, in South Africa. Onderstepoort J. Vet. Res. 2005, 72, 165–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appleton, C. Alien and invasive fresh water Gastropoda in South Africa. Afr. J. Aquat. Sci. 2003, 28, 69–81. [Google Scholar] [CrossRef]
- Ndlovu, M.; Clulow, A.D.; Savage, M.J.; Nhamo, L.; Magidi, J.; Mabhaudhi, T. An Assessment of the Impacts of Climate Variability and Change in KwaZulu-Natal Province, South Africa. Atmosphere 2021, 12, 427. [Google Scholar] [CrossRef]
- Ndlovu, M.S.; Demlie, M. Assessment of meteorological drought and wet conditions using two drought indices across KwaZulu-Natal Province, South Africa. Atmosphere 2020, 11, 623. [Google Scholar] [CrossRef]
- Allan, F.; Ame, S.M.; Tian-Bi, Y.-N.T.; Hofkin, B.V.; Webster, B.L.; Diakité, N.R.; N’Goran, E.K.; Kabole, F.; Khamis, I.S.; Gouvras, A.N. Snail-related contributions from the Schistosomiasis Consortium for Operational Research and Evaluation program including xenomonitoring, focal mollusciciding, biological control, and modeling. Am. J. Trop. Med. Hyg. 2020, 103, 66–79. [Google Scholar] [CrossRef]
- Brown, D.; Kristensen, T. A field guide to African freshwater snails, southern African species. Dan. Bilharz. Lab. Publ. Number 1989, 383. [Google Scholar]
- Mereta, S.T.; Bedewi, J.; Yewhalaw, D.; Mandefro, B.; Abdie, Y.; Tegegne, D.; Birke, W.; Mulat, W.L.; Kloos, H. Environmental determinants of distribution of freshwater snails and trematode infection in the Omo Gibe River Basin, southwest Ethiopia. Infect. Dis. Poverty 2019, 8, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, U.B.; Midzi, N.; Mduluza, T.; Soko, W.; Stensgaard, A.-S.; Vennervald, B.J.; Mukaratirwa, S.; Kristensen, T.K. Modelling spatial distribution of snails transmitting parasitic worms with importance to human and animal health and analysis of distributional changes in relation to climate. Geospat. Health 2014, 8, 335–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.R.; Gorley, R.; Somerfield, P.J.; Warwick, R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; PRIMER-E Ltd.: Auckland, New Zealand, 1994; p. 176. [Google Scholar]
- Kiernan, D. Natural Resources Biometrics; Open SUNY Textbooks, Milne Library; State University of New York at Geneseo: New York, NY, USA, 2014. [Google Scholar]
- Leinster, T.; Cobbold, C.A. Measuring diversity: The importance of species similarity. Ecology 2012, 93, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Chimbari, M.J.; Kalinda, C.; Siziba, N. Changing patterns of Schistosoma host snail population densities in Maun, Botswana. Afr. J. Aquat. Sci. 2020, 45, 493–499. [Google Scholar] [CrossRef]
- Usman, A.I.; Adamu, T.; Abdulhamid, A. Studies on distribution and abundance of freshwater snail intermediate hosts of schistosomiasis along Kwanar Areh Dam in Rimi LGA of Katsina State. J. Parasitol. Vector Biol. 2019, 11, 26–35. [Google Scholar]
- Sharma, K.; Bangotra, K.; Saini, M. Diversity and distribution of mollusca in relation to the physico-chemical profile of Gho-Manhasan stream, Jammu (JK). Int. J. Biodivers. Conserv. 2013, 5, 240–249. [Google Scholar]
- Oladejo, M.K.; Oloyede, O.O.; Adesakin, T.A.; Morenikeji, O.A. The abundance, distribution and diversity of invasive and indigenous freshwater snails in a section of the Ogunpa River, southwest Nigeria. Molluscan Res. 2021, 41, 222–234. [Google Scholar] [CrossRef]
- Miranda, N.; Perissinotto, R. Effects of an alien invasive gastropod on native benthic assemblages in coastal lakes of the iSimangaliso Wetland Park, South Africa: Ecology. Afr. Invertebr. 2014, 55, 209–228. [Google Scholar]
- Miranda, N.A.; Perissinotto, R. Benthic assemblages of wetlands invaded by Tarebia granifera (Lamarck, 1822) (Caenogastropoda: Thiaridae) in the iSimangaliso Wetland Park, South Africa. Molluscan Res. 2014, 34, 40–48. [Google Scholar] [CrossRef]
- Miranda, N.A.; Perissinotto, R.; Appleton, C.C. Salinity and temperature tolerance of the invasive freshwater gastropod Tarebia granifera. S. Afr. J. Sci. 2010, 106, 1–7. [Google Scholar] [CrossRef]
- Brown, D.S. Freshwater Snails of Africa and Their Medical Importance; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Odongo-Aginya, E.; Kironde, F.; Kabatereine, N.; Kategere, P.; Kazibwe, F. Effect Of Seasonal Rainfall And Other Environmental Changes, On Snail Density And Infection Rates With Schistosoma mansoni Fifteen Years After The Last Snails Study In Kigungu, Entebbe, Uganda. East Afr. Med. J. 2008, 85, 556–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndione, R.A.; Bakhoum, S.; Haggerty, C.; Jouanard, N.; Senghor, S.; Ndao, P.D.; Riveau, G.; Ba, C.T. Intermediate Host Snails of Human Schistosomes in the Senegal River Delta: Spatial Distribution According to Physicochemical Parameters. Invertebr. -Ecophysiol. Manag. 2019, 71, 1–10. [Google Scholar]
- Ofoezie, I.E. Distribution of freshwater snails in the man-made Oyan Reservoir, Ogun State, Nigeria. Hydrobiologia 1999, 416, 181–191. [Google Scholar] [CrossRef]
- Cañete, R.; Yong, M.; Sánchez, J.; Wong, L.; Gutiérrez, A. Population dynamics of intermediate snail hosts of Fasciola hepatica and some environmental factors in San Juan y Martinez municipality, Cuba. Memórias Do Inst. Oswaldo Cruz 2004, 99, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Brown, D. A review of the freshwater Mollusca of Natal and their distribution. Ann. Natal Mus. 1967, 18, 477–494. [Google Scholar]
- Kalinda, C.; Chimbari, M.J.; Mukaratirwa, S. Effect of temperature on the Bulinus globosus—Schistosoma haematobium system. Infect. Dis. Poverty 2017, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Woolhouse, M.; Chandiwana, S. Population dynamics model for Bulinus globosus, intermediate host for Schistosoma haematobium, in river habitats. Acta Trop. 1990, 47, 151–160. [Google Scholar] [CrossRef]
- Salawu, O.; Odaibo, A. The bionomics and diversity of freshwater snails species in Yewa North, Ogun State, Southwestern Nigeria. Helminthologia 2014, 51, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Oso, O.G.; Odaibo, A.B. Land use/land cover change, physico-chemical parameters and freshwater snails in Yewa North, Southwestern Nigeria. PLoS ONE 2021, 16, e0246566. [Google Scholar] [CrossRef]
- Boelee, E.; Laamrani, H. Environmental control of schistosomiasis through community participation in a Moroccan oasis. Trop. Med. Int. Health 2004, 9, 997–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pointier, J.-P. Invading freshwater snails and biological control in Martinique Island, French West Indies. Memórias Do Inst. Oswaldo Cruz 2001, 96, 67–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez Perez, J.; Vargas, M.; Malek, E.A. Displacement of Biomfhalaria glabrata by thiara granifera under natural conditions in the Dominican Republic. Memórias Do Inst. Oswaldo Cruz 1991, 86, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Pointier, J.; Incani, R.; Balzan, C.; Chrosciechowski, P.; Prypchan, S. Invasion of the rivers of the littoral central region of Venezuela by Thiara granifera and Melanoides tuberculata (Mollusca: Prosobranchia: Thiaridae) and the absence of Biomphalaria glabrata, snail host of Schistosoma mansoni. Nautilus 1994, 107, 124–128. [Google Scholar]
- Brackenbury, T.D.; Appleton, C. Recolonization of the Umsindusi river, Natal, South Africa, by the invasive gastropod, Physa acuta (Basommatophora, Physidae). J. Med. Appl. Malacol. 1993, 5, 39–44. [Google Scholar]
- Zukowski, S.; Walker, K.F. Freshwater snails in competition: Alien Physa acuta (Physidae) and native Glyptophysa gibbosa (Planorbidae) in the River Murray, South Australia. Mar. Freshw. Res. 2009, 60, 999–1005. [Google Scholar] [CrossRef]
Figure 1.
Map of KZN province showing locations where snails were found according to species.


{kind=link}
Table 1.
Summary of the distribution and abundance of freshwater snails collected from 127 sites in 11 districts in KZN between December 2020 to February 2021.
Table 1.
Summary of the distribution and abundance of freshwater snails collected from 127 sites in 11 districts in KZN between December 2020 to February 2021.
| Districts | Number of Sites | Snail Species | Total Snail Abundance | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| B. pfeifferi | B. globosus | B. forskalii | B. tropicus | L. natalensis | T. granifera | P. acuta | Bivalves | |||
| Amajuba | 9 | 0 | 5 | 122 | 83 | 2 | 0 | 0 | 0 | 212 |
| eThekwini | 20 | 1 | 26 | 0 | 123 | 12 | 215 | 0 | 50 | 427 |
| iLembe | 9 | 0 | 6 | 0 | 0 | 50 | 302 | 0 | 0 | 358 |
| King Cetshwayo | 11 | 1 | 117 | 10 | 5 | 0 | 135 | 0 | 0 | 268 |
| Sisonke | 10 | 0 | 0 | 7 | 153 | 100 | 0 | 10 | 45 | 315 |
| Ugu | 10 | 145 | 20 | 0 | 410 | 50 | 320 | 0 | 0 | 945 |
| uMgungundlovu | 10 | 0 | 0 | 0 | 97 | 82 | 0 | 140 | 0 | 319 |
| uMkhanyakude | 13 | 12 | 31 | 0 | 0 | 0 | 847 | 0 | 0 | 890 |
| uMzinyathi | 7 | 0 | 33 | 0 | 20 | 9 | 0 | 0 | 0 | 62 |
| uThukela | 12 | 87 | 136 | 0 | 49 | 12 | 122 | 0 | 0 | 406 |
| Zululand | 16 | 0 | 114 | 0 | 0 | 0 | 260 | 0 | 0 | 374 |
| Total snail abundance | 127 | 246 | 488 | 139 | 940 | 317 | 2201 | 150 | 95 | 4576 |
Table 2.
Biodiversity indices of snail species in 11 districts in KZN between December 2020 and February 2021.
Table 2.
Biodiversity indices of snail species in 11 districts in KZN between December 2020 and February 2021.
| Number of Species | Shannon Index (H′) | Simpson’s Index (D) | Pielou’s Index (J′) | Margalef’s Index (d) | ENS | |
|---|---|---|---|---|---|---|
| Ugu | 8 | 1.63 | 0.74 | 0.78 | 0.84 | 5.10 |
| uMkhanyakude | 3 | 0.22 | 0.09 | 0.20 | 0.29 | 1.25 |
| Zululand | 3 | 0.73 | 0.45 | 0.66 | 0.34 | 2.08 |
| uThukela | 5 | 1.42 | 0.74 | 0.88 | 0.67 | 4.14 |
| iLembe | 2 | 0.10 | 0.04 | 0.14 | 0.17 | 1.11 |
| eThekwini | 6 | 1.24 | 0.65 | 0.69 | 0.83 | 3.46 |
| Sisonke | 5 | 1.19 | 0.64 | 0.74 | 0.70 | 3.29 |
| uMgungundlovu | 4 | 1.09 | 0.65 | 0.79 | 0.52 | 2.97 |
| uMzinyathi | 4 | 0.82 | 0.43 | 0.59 | 0.54 | 2.27 |
| Amajuba | 4 | 0.82 | 0.52 | 0.59 | 0.56 | 2.27 |
| King Cetshwayo | 5 | 0.93 | 0.56 | 0.57 | 0.72 | 2.53 |
Table 3.
Correlation coefficients show relationships between snail species.
| B. globosus | B. pfeifferi | B. tropicus | B. forskalii | L. natalensis | T. granifera | Bivalves | P. acuta | |
|---|---|---|---|---|---|---|---|---|
| B. globosus | 1.00 | |||||||
| B. pfeifferi | 0.713 * | 1.00 | ||||||
| B. tropicus | −0.138 | 0.196 | 1.00 | |||||
| B. forskalii | 0.063 | 0.115 | 0.457 | 1.00 | ||||
| L. natalensis | −0.009 | 0.153 | 0.840 * | 0.128 | 1.00 | |||
| T. granifera | 0.527 | 0.599 | −0.272 | −0.092 | −0.160 | 1.00 | ||
| Bivalves | 0.025 | 0.103 | 0.725 * | 0.092 | 0.680 * | −0.079 | 1.00 | |
| P. acuta | −0.145 | 0.100 | 0.718 * | 0.321 | 0.784 * | −0.030 | 0.593 * | 1.00 |
* Significant correlation at p < 0.05.
Table 4.
Physicochemical parameter values (mean ± SD) of water measured in 11 districts in KZN province between December 2020 and February 2021.
Table 4.
Physicochemical parameter values (mean ± SD) of water measured in 11 districts in KZN province between December 2020 and February 2021.
| Districts | pH | Electrical Conductivity | Salinity (psu) | Pressure (psi) | Dissolved Oxygen (ppm) |
|---|---|---|---|---|---|
| 1. Zululand | 7.98 ± 0.55 | 232.49 ± 163.65 | 0.11 ± 0.08 | 13.34 ± 0.53 | 4.33 ± 3.64 |
| 2. uThukela | 7.80 ± 0.40 | 142.58 ± 120.34 | 0.07 ± 0.06 | 12.94 ± 0.13 | 5.73 ± 1.64 |
| 3. iLembe | 7.74 ± 0.22 | 176.39 ± 100.30 | 0.08 ± 0.05 | 14.10 ± 0.32 | 8.19 ± 1.49 |
| 4. eThekwini | 7.54 ± 0.35 | 695.56 ± 239.27 | 0.32 ± 0.17 | 14.51 ± 0.24 | 6.34 ± 1.87 |
| 5. Ugu | 7.19 ± 0.34 | 358.92 ± 216.67 | 13.12 ± 38.82 | 12.81 ± 4.28 | 9.40 ± 10.27 |
| 6. Sisonke | 7.30 ± 0.27 | 125.09 ± 65.04 | 0.06 ± 0.03 | 13.01 ± 0.66 | 7.71 ± 1.25 |
| 7. uMgungundlovu | 7.24 ± 0.26 | 211.63 ± 313.07 | 0.10 ± 0.15 | 13.14 ± 0.69 | 6.76 ± 0.97 |
| 8. uMzinyathi | 7.42 ± 0.28 | 199.04 ± 79.59 | 0.09 ± 0.04 | 13.02 ± 0.20 | 5.50 ± 1.29 |
| 9. Amajuba | 6.42 ± 1.04 | 1005.30 ± 1108.77 | 0.51 ± 0.59 | 44.13 ± 89.12 | 6.80 ± 1.20 |
| 10. King Cetshwayo | 7.35 ± 0.32 | 695.33 ± 547.55 | 0.34 ± 0.28 | 14.51 ± 0.43 | 5.56 ± 1.70 |
Table 5.
Associations between snail abundance, physicochemical parameters, and climatic factors.
| B. globosus | B. pfeifferi | B. tropicus | B. forskalii | L. natalensis | T. granifera | Bivalves | P. acuta | |
|---|---|---|---|---|---|---|---|---|
| pH | 0.3040 * | 0.0069 | −0.6383 * | −0.7647 * | −0.2683 * | 0.3564 * | −0.3556 * | −0.5891 * |
| EC | 0.1398 | 0.1988 | 0.0851 | 0.4506 * | −0.2378 | 0.1688 | −0.0065 | −0.1715 |
| Salinity | 0.3465 * | 0.3564 * | 0.1824 | 0.5872 * | −0.0671 | 0.3377 * | 0.1358 | 0.0969 |
| DO | −0.2492 * | 0.1234 | 0.5532 * | 0.4370 * | 0.3415 * | 0.2126 | 0.2521 * | 0.5891 * |
| Rainfall | 0.1048 | −0.0348 | −0.7615 * | 0.0316 | −0.8047 * | 0.0419 | −0.8476 * | −0.6706 * |
| NDVI | 0.1595 | 0.2038 | −0.0550 * | 0.2948 * | −0.2621 * | 0.3955 * | 0.2181 | 0.2370 * |
| EVI | −0.1230 | 0.0795 | −0.0459 | −0.0211 | −0.0966 | 0.3443 * | 0.1834 | 0.2948 * |
| NDWI | −0.3872 * | −0.0994 | 0.0000 | 0.0211 | −0.1058 | 0.2466 * | 0.2379 * | 0.3064 * |
| MinTemp | 0.4328 * | 0.4970 * | −0.4404 * | −0.2632 * | −0.4736 * | 0.7583 * | −0.1090 | −0.3237 * |
| MaxTemp | 0.5148 * | 0.2286 * | −0.7615 * | −0.2527 * | −0.7679 * | 0.4141 * | −0.3618 * | −0.6764 * |
* Significant correlation at p < 0.05.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nwoko, O.E.; Kalinda, C.; Manyangadze, T.; Chimbari, M.J. Species Diversity, Distribution, and Abundance of Freshwater Snails in KwaZulu-Natal, South Africa. Water 2022, 14, 2267. https://doi.org/10.3390/w14142267
AMA Style
Nwoko OE, Kalinda C, Manyangadze T, Chimbari MJ. Species Diversity, Distribution, and Abundance of Freshwater Snails in KwaZulu-Natal, South Africa. Water. 2022; 14(14):2267. https://doi.org/10.3390/w14142267
Chicago/Turabian StyleNwoko, Onyekachi Esther, Chester Kalinda, Tawanda Manyangadze, and Moses John Chimbari. 2022. "Species Diversity, Distribution, and Abundance of Freshwater Snails in KwaZulu-Natal, South Africa" Water 14, no. 14: 2267. https://doi.org/10.3390/w14142267
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.