
Reductive Cr(VI) Removal under Different Reducing and Electron Donor Conditions—A Soil Microcosm Study
by
 ,
,
Andriani Galani
, 
Constantinos Noutsopoulos
,
Petra Anastopoulou
,
Alexia Varouxaki
and
Daniel Mamais
* Sanitary Engineering Laboratory, Department of Water Resources and Environmental Engineering, School of Civil Engineering, National Technical University of Athens, Iroon Polytechniou 9, Zografou, 157 80 Athens, Greece
*
Author to whom correspondence should be addressed.
Water 2022, 14(14), 2179; https://doi.org/10.3390/w14142179
Submission received: 20 June 2022
/
Revised: 3 July 2022
/
Accepted: 8 July 2022
/
Published: 10 July 2022
(This article belongs to the Section Water Quality and Contamination)
Abstract
:Increased groundwater and soil contamination by hexavalent chromium have led to the employment of a variety of detoxification methods. Biological remediation of Cr(VI) polluted aquifers is an eco-friendly method that can be performed in situ by stimulating the indigenous microbial population with organic and inorganic electron donors. In order to study the effect of different redox conditions on microbial remediated Cr(VI) reduction to Cr(III), microcosm experiments were conducted under anaerobic, anoxic, and sulfate-reducing conditions and at hexavalent chromium groundwater concentrations in the 0–3000 μg/L range, with groundwater and soil collected from an industrial area (Inofyta region). As electron donors, molasses, emulsified vegetable oil (EVO), and FeSO4 were employed. To quantitatively describe the degradation kinetics of Cr(VI), pseudo-first-order kinetics were adopted. The results indicate that an anaerobic system dosed with simple or complex external organic carbon sources can lead to practically complete Cr(VI) reduction to Cr(III), while the addition of Fe2+ can further increase Cr(VI) removal rate significantly. Furthermore, Cr(VI) microbial reduction is possible in the presence of NO3− at rates comparable to anaerobic Cr(VI) microbial reduction, while high sulfate concentrations have a negative effect on Cr(VI) bioreduction rates in comparison to lower sulfate concentrations.
1. Introduction
High hexavalent chromium [Cr(VI)] concentrations found in the environment during the last decades have raised public concern, owing to its bio-accumulative, carcinogenic, mutagenic, and toxic nature [1,2]. Multiple health and environmental protection agencies consider Cr(VI) a priority pollutant [3] and have set strict limits for its concentration in the aquatic environment [4]. Throughout the years, various pump and treat physicochemical technologies have been employed to remediate Cr(VI) polluted soil and groundwater (ion exchange, chemical precipitation, adsorption, membrane filtration, etc.) [5,6,7,8]. However, these technologies present certain disadvantages, such as secondary pollution, high operational and maintenance cost, and low effectiveness [9,10]. In situ biological remediation is, in contrast, an environmentally friendly and cost-effective method that takes advantage of microbial metabolic processes to reduce Cr(VI) to trivalent chromium [Cr(III)], a stable form of Cr that has been found to be less soluble in water, exists mainly as precipitate and is less toxic for organisms [11,12].
Microbial remediated Cr(VI) reduction can occur under both aerobic and anaerobic conditions and is mediated by a variety of enzymes, being cited extracellularly, intracellularly, or in the cytoplasmic membrane [13,14]. A reduction can also be induced by metabolic byproducts, such as bio-generated Fe(II), hydrogen, and/or sulfides [15,16,17]. One significant advantage of bioremediation is that it can be performed in situ without causing the destruction of the site and that it allows for a continuous Cr(VI) reduction process through the addition of electron donors [18]. However, the effectiveness of Cr(VI) microbial reduction can be affected by the specific physicochemical characteristics of the contaminated area, including pH, temperature, other toxic substances, or different electron acceptors [19]. In many cases, Cr(VI) coexists with other harmful pollutants for public health and the environment. Nitrate (NO3−) is one of the most abundant pollutants in groundwater originating mainly from agricultural activities, i.e., excessive use of nitrogenous fertilizers, as well as discharge of not properly treated urban and industrial wastewater [20]. High concentrations of NO3− can lead to eutrophication of water bodies [21], while its reduction intermediate, nitrite (NO2−), can cause methemoglobinemia, cancer, and thyroid dysfunction in humans [22,23]. Therefore, effective remediation of nitrate-contaminated groundwater is of the utmost importance.
In recent years, simultaneous Cr(VI) and NO3− bioreduction have been the focus of various studies since the interaction mechanisms between those two chemical species are quite complex. To begin with, both chromium and nitrates compete for the supply of electron donors and may be affected by the presence of other competing electron acceptors [24]. Under aerobic conditions, researchers have found that both nitrate and Cr(VI) reduction can occur at the same time, with Cr(VI) removal rates being unaffected by the presence of nitrate, but Cr(VI) concentration affecting and, in certain cases, significantly inhibiting denitrification [25,26], although the reported Cr(VI) threshold values were rather contradictory (ranging from 7.5 mg/L to 30 mg/L). However, in other studies, high concentrations of chromate did not adversely affect heterotrophic denitrification [27,28,29] with specific bacterial strains or mixed cultures. On the other hand, it has been reported that under high nitrate concentration, Cr(VI) reduction is irreversibly inhibited [30]. Nevertheless, the majority of these studies have focused on ex situ applications and, in some cases, were conducted with very specific microbial strains.
Sulfate (SO42−) is another ubiquitous pollutant in groundwater, originating both from natural and anthropogenic sources, such as weathering of sulfide-containing minerals, volcanic activities, rainfall, seawater intrusion, fertilizers, urban and industrial wastes, and wastewater [31,32,33,34]. Under the absence of oxygen, sulfate can be reduced to sulfide by sulfate-reducing bacteria (SRB), anaerobes that use sulfate as a terminal electron acceptor. Produced sulfide has been reported to facilitate the reduction of Cr(VI) to Cr(III) [35], while some strains of SRB have the ability to enzymatically reduce Cr(VI) [36,37]. However, the presence of SO42− could possibly have an inhibitory effect on Cr(VI) reduction due to competition as electron acceptors [38].
In view of the above, the goal of this experimental work was to investigate Cr(VI) reduction rates in groundwater through biostimulation, in completely anaerobic, nitrate-rich, and sulfate-reducing environments and under limiting electron donor conditions. To this end, soil microcosm experiments were carried out under four (4) different reducing conditions—anaerobic, anoxic, low, and high sulfate concentration—with soil and groundwater collected from a deep Cr(VI) contaminated aquifer in the Asopos River basin. In order to obtain results that are representative of the real field conditions, parent cultures were developed under each reducing environment in the presence of low Cr(VI) and organic donor concentrations in order to provide biomass for the batch experiments. Furthermore, various organic and inorganic electron donors were evaluated for their ability to stimulate microbial communities. Emphasis was placed on: (i) the investigation of the effect of electron acceptor on Cr(VI)-reducing microbial communities under anoxic, anaerobic, and different sulfate-reducing conditions and (ii) the evaluation of the effect of a combination of electron donors, including simple and complex organic carbon and iron, on anaerobic or anoxic Cr(VI) reduction and denitrification and on Cr(VI) reduction in the presence of sulfates.
To the best of our knowledge, there are very few studies that have attempted to provide a comprehensive examination of the in situ biological treatment of groundwater polluted with Cr(VI) under different reductive environments. The findings of this study could provide an important basis for in situ anaerobic bioremediation through biostimulation, as well as access to the longevity of the process under limiting electron donor conditions.
2. Materials and Methods
2.1. Soil and Groundwater Source and Characteristics
Soil and groundwater samples used for the microcosm experiments were collected from the deeper aquifer (28–28.5 m) in Inofyta, a heavily industrialized area located in the Asopos River basin in central Greece, where high hexavalent chromium concentrations (exceeding 10,000 μg/L) have been reported [39,40]. The collected soil and groundwater were transported to the laboratory and stored at 4 °C. Groundwater characteristics are presented in Table 1. Groundwater pH was 7.98 and had high NO3-N and SO42− concentrations (11.20 mg/L and 51.00 mg/L respectively), while Cr(VI) was not detected.
Before the commencement of the experiments, soil was air dried and sieved through a 2 mm sieve. The same soil has been used in a previous study and has been described there in detail [41]. It consisted of 58.8% sand, 25.7% clay, and 15.5% silt and had an average organic content matter of 0.16%. The soil pH was 8.31, and its main components were CaO (30.03%), SiO2 (19.04%), MgO (9.50%), and Fe2O3 (5.08%). Cr and Ni concentrations were 2195 mg/kg and 1082 mg/kg, respectively, as the soil originated from an area contaminated by industrial wastewater.
2.2. Parent Culture Development
Biomass, soil, and liquid amendments for the batch experiments were provided by four (4) parent cultures—completely anaerobic (ANAER), anoxic (ANOX), with low sulfate concentration (S100) and high sulfate concentration (S400)—that were developed and maintained in the laboratory. All were exposed to low hexavalent chromium levels (0.5 mg/L) and to different electron acceptors (10 mg/L NO3-N, 100 mg/L SO42−, 400 mg/L SO42− or none). The cultures were developed in two-liter bottles that were sealed with gray butyl rubber stopper and aluminum crimp. They contained a slurry of subsurface material and groundwater (1:4 dry soil to water volume ratio) and 30 mg TSS/L (TSS: total suspended solids) of activated sludge from an anaerobic reactor as inoculum. The parent cultures were acclimatized for a period of at least 135 days prior to providing biomass for the batch experiments. Before the commencement of the acclimatization period, all cultures were equilibrated for two weeks to deplete electron acceptors in groundwater (i.e., NO3-N and SO42−). They were fed weekly with a reduced anaerobic mineral medium (RAMM) [42], selenium-tungsten solution [43], yeast extract, vitamin solution [44], and 30 mg/L butyrate, which served as the organic electron donor. In the anoxic and the two sulfate reducing parent cultures developed, 10 mg/L of NO3-N, 100 mg/L SO42− and 400 mg/L SO42− were added, respectively, as electron acceptors. All solutions were prepared with sterilized water. At the end of each feeding cycle, part of the cultures was replaced by fresh medium to achieve a solid retention time of 45 days. At the end of each weekly cycle, Cr(VI), NO3-N, and SO42− were measured, and the amount to be added was estimated to reach the default concentration for the beginning of the next cycle.
2.3. Microcosms Design
The experiments were conducted according to a modification of the USEPA guidelines concerning anaerobic biotransformation of pollutants in the subsurface [45]. Individual microcosms were constructed in 160 mL serum bottles with the equivalent of 20 g (dry weight) soil to which 80 mL of liquid amendments were added. The soil and the liquid amendments originated from the parent culture. The bottles were flushed with a mixture of N2/CO2 (70%/30%) to remove trapped oxygen from the liquid and solid phases and sealed with gray butyl rubber stoppers and aluminum crimps. Microcosms were incubated in the dark at 22 °C ± 2 without shaking.
Under anaerobic conditions, several types of organic substrates have been reported to be able to reduce hexavalent chromium successfully, such as glucose, lactate, sucrose, milk whey, etc. [13]. Within the context of this study, molasses emulsified vegetable oil (EVO) and ferrous sulfate heptahydrate (FeSO4 ·7H2O) were used as electron donors, independently and as combinations. The organic electron donors were selected for their different properties. Molasses is a byproduct of sugar refining that can be used as a readily available energy source by microbes [46,47,48], while EVOs are slowly soluble substrates that ferment to acetate and hydrogen [49,50,51]. Five biological microcosm treatments were set up for each oxidative condition (anoxic, anaerobic, and sulfate-reducing), and duplicate bottles were prepared for each treatment. After preparation, each bottle was supplied once at the beginning of the experiment, with the selected electron donor (i.e., molasses, emulsified vegetable oil (EVO), ferrous sulfate heptahydrate, or a combination of carbon and the iron source, or none). All the field conditions simulated through the microcosm experiments are shown in Table 2 and Table 3.
Non-biological processes (e.g., sorption) that participate in the removal of Cr(VI) were also accessed by conducting experiments under sterile conditions and without the addition of any e− donor, as soil microcosms are complex systems [52]. Soil and water from the parent cultures were autoclaved twice, 24 h apart, at 121 °C for 20 min to completely inhibit all biological activity. These experiments were performed in duplicate for all conditions studied (anoxic, sulfate-reducing, and pure anaerobic), and the treatments were spiked with Cr(VI), NO3-N (only in the ANOX experiment), and SO42− (only in the S100 and S400 experiments) according to the following experimental program.
The experiment then proceeded by spiking the bottles with Cr(VI) every 2–3 days without the addition of any reducing agent. Initial Cr(VI) concentrations were chosen between 2000–3000 μg/L, well within the reported range of contamination at the Inofyta region [53], and in order to facilitate a reliable investigation of the reductive capabilities of each microbial community. In the anaerobic batch tests, the initial Cr(VI) concentration was chosen at 2000 μg/L, as previous studies have shown that in complete anaerobic conditions, hexavalent chromium is toxic for the microbes in concentrations higher than 2500 μg/L [54]. For the anoxic experiment, initial Cr(VI) and NO3-N concentrations were 2000 μg/L and 10,000 μg/L, respectively, for the first 10 days of the experiment. For the remainder (days 10–60), these concentrations were increased to 3000 μg/L and 20,000 μg/L in order to evaluate the efficacy of the reducing microbial community at higher pollutant concentrations. In the S100 and S400 experiments, initial Cr(VI) concentration was 2000 μg/L for the first 10 days of the experiment and 3000 μg/L for the remainder, while sulfate concentrations were maintained at 100 mg/L and 400 mg/L, respectively, throughout the experiment. On the day of the spike, liquid samples were withdrawn at predetermined time intervals, filtered through 0.45 μm syringe filter and analyzed for Cr(VI), NO3-N and SO42−. Cr(VI), nitrate, and sulfate were added accordingly to reach the default concentrations.
2.4. Chemicals and Reagents
Molasses was provided by Thermo Fischer Scientific (Waltham, MA, USA), while emulsified vegetable oil (EVO) (60% lactate/40% water) by JRW Bioremediation, L.L.C. (Lenexa, KS, USA). The rest of the chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA) and were of high purity. Butyric acid (≥99%), Acetic acid (≥99.8%), Propionate (≥99.8%), isoButyrate (≥99.5%), and Acetone (≥99.5%) were supplied in liquid form and used as received. Within the context of these experiments, 1,5-Diphenylcarbazide (≥98%), NaH2PO4 (≥99.5%), KNO3 (≥99%), and Na2SO4 (≥99%) were also used.
The following chemicals were used for the development of the parent cultures and were purchased in powder form. MgCl2·6H2O (≥99%), CaCl2·6H2O (≥98%), K2HPO4 (≥98%), NH4Cl (≥99.5%), NaHCO3 (≥99.5%), Na2S·9H2O (≥98%), nitrilotriacetic acid (≥98%), FeSO4·7H2O (≥99%), MnSO4·H2O (≥99%), CoCl2·6H2O (≥98%), ZnSO4·7H2O (≥99%), H3BO3 (≥99%), NiCl2 (≥99%), NaMoO4 (≥98%) were used for the preparation of RAMM. The vitamin solution consisted of biotin, DL-pantothenic calcium, folic acid, nicotinic acid, riboflavin, pyridoxine hydrochloride, thiamine hydrochloride, vitamin B12, paminobenzoic acid, while yeast extract solution (EZMixTM) was prepared separately. The selenium-tungsten solution contained Na2SeO3·5H2O (≥99%) and Na2WO4·2H2O (≥99%).
2.5. Analytical Procedures
During the development of the parent cultures and throughout the batch experiments, the following parameters were monitored: pH, dissolved oxygen (DO), oxidation-reduction potential (ORP), hexavalent Cr, chemical oxygen demand (COD), volatile fatty acids (VFAs), nitrate nitrogen (NO3-N), nitrite nitrogen (NO2-N), sulfate (SO42−), sulfide (S2−) and ferrous iron [Fe(II)]. DO and ORP were monitored to ensure that reductive conditions were established. Chromate in solution was determined colorimetrically using the USEPA 7196a method (1,5-diphenylcarbazide method) [55], while COD was measured by USEPA-approved standard method 5220 D [56] and ferrous iron Fe(II) by the 1,10-phenanthroline method [57]. Nitrate nitrogen, nitrite nitrogen, sulfate, and sulfide were measured using Hach methods LCK339, LCK8507, 8051, and 8131, respectively, on a Hach Lange DR2800 spectrophotometer.
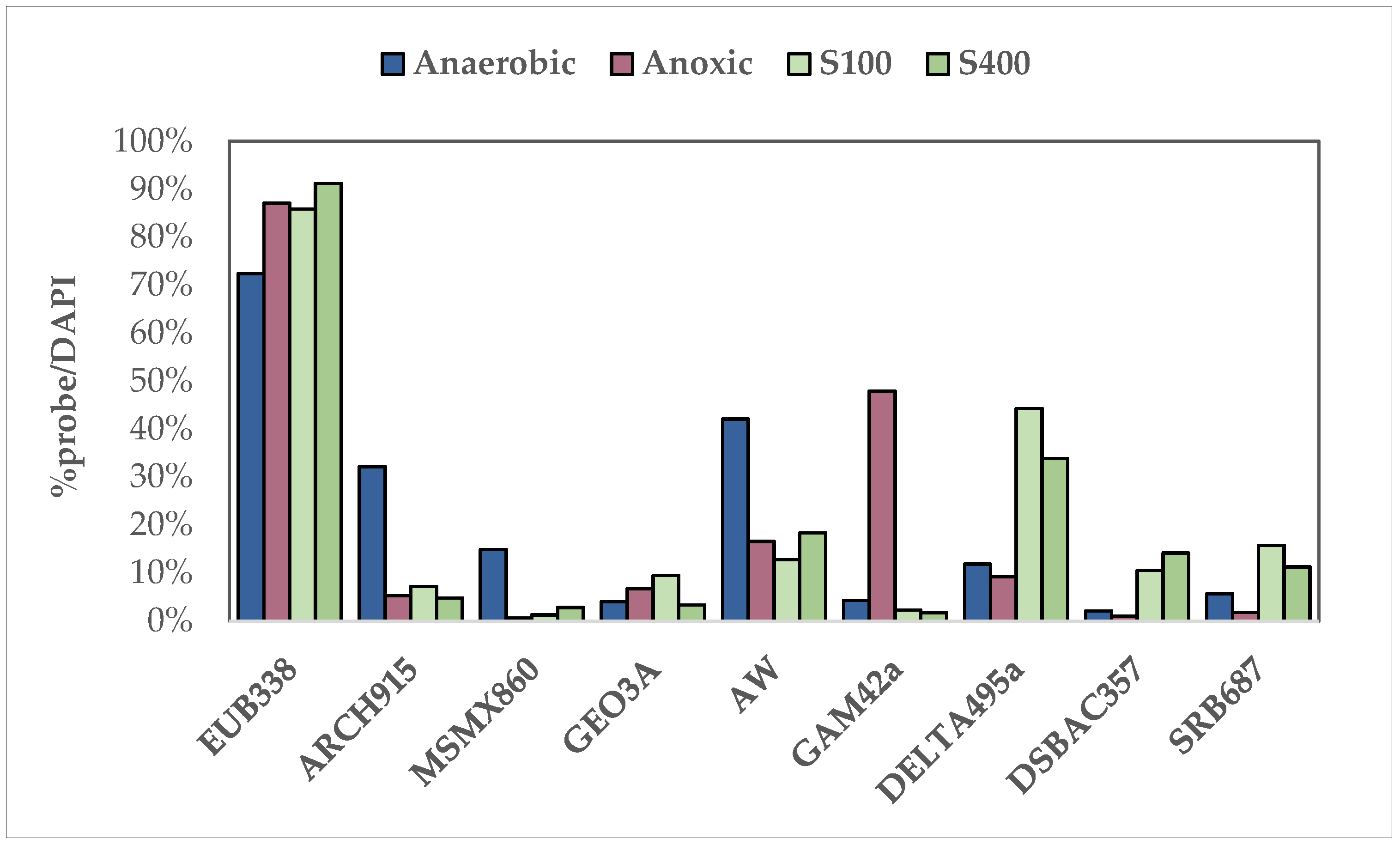
FISH analysis was carried out in liquid samples of the parent cultures after the ending of the acclimatization period following the protocol described by Panousi et al. [54] to estimate the presence of the following groups of microbes (with the probe used in parenthesis): Eubacteria (EUB338) [58], Archaea (ARCH915) [59], Methanosarcina and Methanosaeta spp. (MSMX860) [60], Acetobacterium spp. (AW) [61], Geobacter spp. (GEO3A) [62], Gammaproteobacteria (GAM42a) [63], Deltaproteobacteria (DELTA495a) [64], Desulfobacteraceae and Syntrophobacteraceae families (DSBAC357) [65] and iron-reducing bacteria (SRB687) [66]. Results were expressed as a fraction of the specific binding of hybridized cells out of the number of the total cells (%probe/DAPI).
The presence of four (4) volatile fatty acids was examined: acetate, butyrate, Propionate, and isoButyrate. VFAs analysis was performed on a Gas Chromatograph equipped with a flame ionization detector, a fused-silica capillary Nukol column (15 m, 0.53 mm, Supelco, Inc., Bellefonte, PA, USA), and an autosampler. A total of 0.5 μL of liquid samples were injected in the Chromatograph, while an isothermal method at 90 °C was utilized, with both injector and detector being held at 220 °C. Before injection, samples were filtered through 0.45 μm filters and acidified with 2% v/v HCl (2M).
2.6. Data Calculation
In order to describe the reduction kinetics of hexavalent chromium and nitrate quantitatively, pseudo-first-order kinetics were used. First-order rate constants (k) were estimated by directly fitting the following equation (Equation (1)) until 90% substance removal was achieved:
where Co and C equal the initial substance concentration and substance concentration at time t, respectively. Time profiles of every duplicate of the batch experiments were produced on linear plots of ln(C/Co) versus time, with the slope of the linear trendline being the (k) constant.
All rates reported are mean values of the duplicates of all the batch tests performed at the selected time intervals (i.e., 0–10 days, 10–20 days, 20–30 days, 30–40 days, and 40–60 days), accompanied by standard deviation.
3. Results and Discussion
3.1. Evaluation of Parent Cultures Performance
During the acclimatization period of the parent cultures, Cr(VI), CODs, pH, NO3-N, NO2-N, sulfate, and sulfide were monitored weekly, while VFAs analysis was carried out periodically. As shown in Table 4, the obtained results indicated that the developed microbial communities were able to completely reduce Cr(VI), NO3-N, and NO2-N in the time span of a week. The S400 microbial community was only able to reduce 30% of the initial sulfate concentration, while the S100 community was able to reduce approximately 42% of initial sulfate, although the reduction ability fluctuated throughout the experiment. Measured sulfide concentrations in liquid samples indicated that sulfide production was higher in the S400 culture (13.2 ± 7.1 mg/L) in comparison to the S100 culture (3.1 ± 2.1 mg/L) since, according to published data, higher sulfate concentrations lead to a higher production of sulfide [67]. It should be noted that measured sulfide that corresponded to soluble sulfide was less than the theoretically produced amount from sulfate reduction, probably due to sulfide precipitation with metals. Optical observation in both sulfate-reducing cultures indicated the blackening of the culture due to the formation of dark-hued solids, the result of the precipitation of metal ions present in the soil of the microcosm experiments with the produced sulfide [31]. pH ranged from 8.15–8.30 in all cultures throughout the acclimatization period without fluctuations, owing to the soil’s calcareous composition. GC analysis showed that the VFAs concentrations in the anoxic and sulfate-reducing microbial cultures were more limited in comparison to the anaerobic one. Furthermore, only acetate and butyrate were present in the samples, with the concentration of the latter being less than 15% of total VFAs, which indicated that most of the butyrate added was used as an energy donor by the microbes and did not accumulate.
At the end of the acclimatization period, FISH analysis was carried out on liquid samples from each parent culture (Figure 1). The molecular analysis results demonstrated the differences between each microbial community. In all cultures, bacteria (EUB338) were the most prominent microorganism. In the complete anaerobic culture, Archaea (ARCH915) population reached a percentage of 32.2%, while Methanosarcina and Methanosaeta spp. (MSMX860) were approximately 14.9%. Furthermore, Acetobacterium spp. (AW) was the dominant microbial group in this culture (42.1%). In contrast, in the anoxic parent culture, the presence of Archaea (ARCH915) and methanogens (MSMX860) was negligible as expected, while Gammaproteobacteria (GAM42a), which include some denitrifying bacteria, reached up to 47.9% and played an important role in the reduction of nitrate [68]. The rest of the examined probes (GEO3A, DELTA495a, DSBAC357, SRB687) exhibited low signals (below 10%), which is indicative of the minimal presence of Geobacter spp., Desulfobacteraceae and Syntrophobacteraceae families and iron-reducing bacteria, in both communities. In the sulfate-reducing microbial colonies, the dominant bacterial group was Deltaproteobacteria (DELTA 495a)—44.3% for the S100 and 33.9% for the S400 parent cultures—that contain sulfate-reducing bacteria, anaerobes that use sulfate as a terminal electron acceptor. Archaea (ARCH915) and methanogens (MSMX860) were minimal since sulfide inhibits their growth and sulfate-reducing bacteria outcompete methanogens [69,70,71]. No significant differences between the sulfate-reducing colonies were reported for the rest of the studied microorganisms.
3.2. Evaluation of Cr(VI) Reduction under Complete Anaerobic Conditions
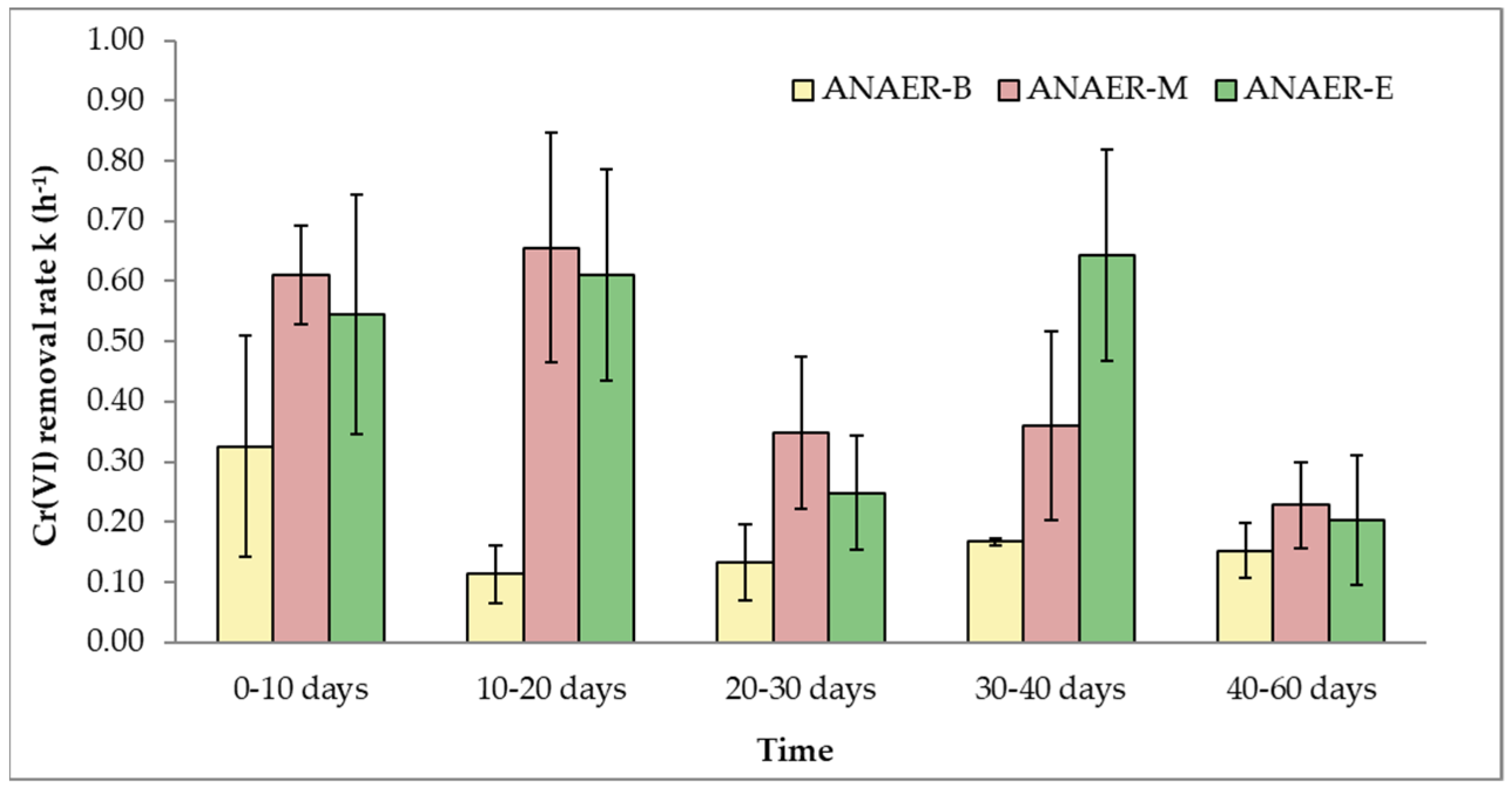
Hexavalent Cr reduction rates were evaluated under anaerobic conditions for a period of 60 days. Batch tests were carried out every 2–3 days, and the initial Cr(VI) concentration was 2000 μg/L. Figure 2 presents the pseudo-first-order Cr(VI) removal rates (k) versus time (days) for two of the conditions studied: (a) the natural soil attenuation (ANAER-B) and (b) the biological enhancement with the addition of a carbon source (ANAER-M and ANAER-E). It should also be underlined that the sterile amendments showed that Cr(VI) removal rates were kSTERILE,ANAER = 0.051 h−1 ± 0.013, which indicates that non-biological processes contributed very little to overall Cr(VI) removal in the amendments where electron donors were added.
As can be observed from the biotic control experiment, hexavalent chromium reduction is possible even without the addition of a reducing agent through endogenous decay. However, compared to batch tests conducted with external organic electron donor addition, the removal rates are 2–6 times slower, which indicates that microbial growth biostimulation was achieved.
In the molasses experiment, the reduction rates show a faster-decreasing pattern, as was expected, since molasses is a readily available organic electron donor that is depleted quickly [72,73]. As a result, molasses is depleted within 20 days, and thereafter Cr(VI) reduction proceeds due to endogenous decay at a lower removal rate. In contrast, in the amendments where EVO was added, the results indicate that although, during the first 30 days of the experiment, the differences between the kANAER-M and kANAER-E are negligible, during the next time period (30–40 days), the kANAER-E removal rate increases by 78% and then decreases rapidly. This finding is in agreement with the expected effect of EVO on the microbes as a slowly depleted electron donor [74]. It is worth noting that all amendments were able to reduce Cr(VI) to levels below 50 μg/L. It should be underlined that according to available studies on Cr speciation in groundwater, Cr(VI) is the major part of chromium in groundwater and represents approximately 70%–99% of total chromium [75,76,77].
Figure 3a,b illustrate the results of the combined biological and chemical Cr(VI) removal. As shown, the addition of Fe2+ and a carbon source can double the removal rates during the first 10 days of the experiment. The rates obtained with Fe2+ addition were 1.25 h−1 ± 0.70 compared to 0.61 h−1 ± 0.08 with molasses as the carbon source and 0.92 h−1 ± 0.38 to 0.54 h−1 ± 0.20 with EVO. This is in accordance with published data, which indicate that chemical reduction of Cr(VI) with ferrous iron is supplementary to microbial reduction [78,79].
Hexavalent chromium and ferrous iron react at a stoichiometric molar ratio of 1/3, producing a nontoxic solid product with the following general molecular formula: Fe(1−×)Crx(OH)3 [80]. Taking into consideration the theoretical ratio, the majority of the Fe(II) was available for Cr(VI) reduction, although it is possible that a small amount could have been passivated due to the soil and groundwater alkaline pH. Furthermore, Fe(II) analysis on liquid samples between the 1st and 10th day of the experiment indicated that soluble ferrous iron was below the detection limit. However, in anaerobic soils and under the presence of microbes and organic e− donors, Fe(III) is reduced to Fe(II), which leads to the formation of highly reactive Fe(II)- bearing minerals, such as magnetite, siderite, and green rusts, that can act as reductants for the abiotic removal of Cr(VI) [81,82,83,84,85]. Between the 10th and 60th day of the experiment, the kinetic rates of the bottles amended with carbon source and Fe(II) were similar to those containing only the corresponding carbon source, so it can be concluded that this mechanism had no significant effect on Cr(VI) removal.
3.3. Evaluation of Cr(VI) Reduction under Anoxic Conditions
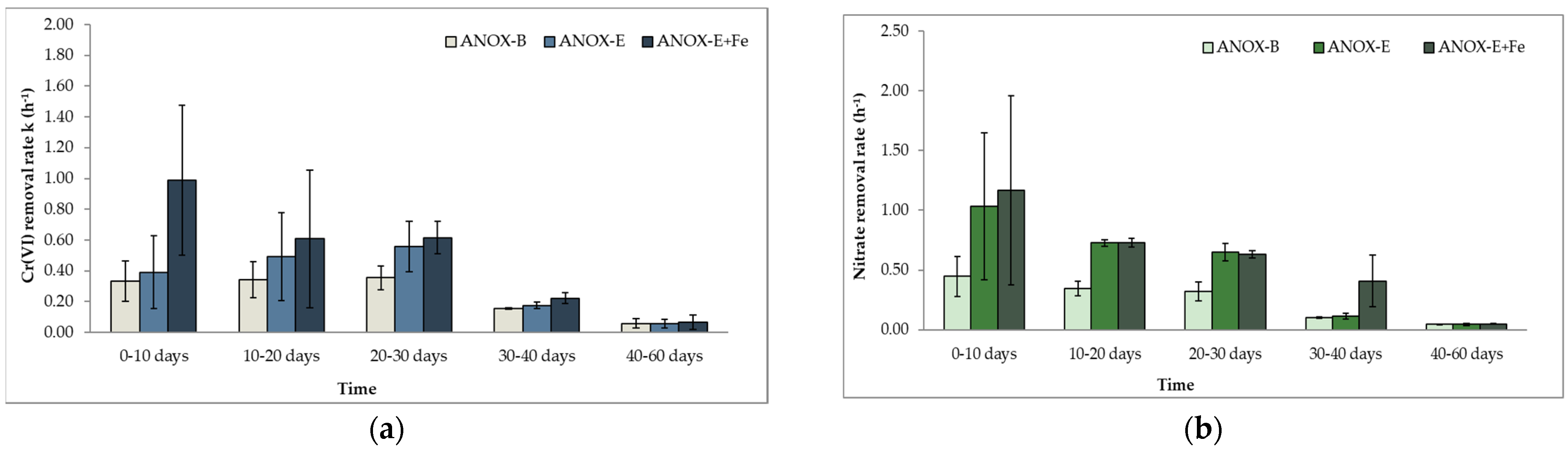
In the anoxic experiment, hexavalent chromium and nitrate removal were evaluated for a period of 60 days. Initial Cr(VI) and NO3-N concentrations were 2000 μg/L and 10,000 μg/L, respectively, for the first 10 days of the experiment, and for the remainder, these concentrations were increased to 3000 μg/L and 20,000 μg/L. The bottles were spiked with Cr(VI) and nitrate every 2–3 days. In Figure 4a,b, the effect of the addition of different organic electron donors on Cr(VI) and NO3-N removal rates is illustrated. Sterile amendments showed that Cr(VI) removal rates were kSTERILE,ANOX = 0.024 h−1± 0.01, which indicates that Cr(VI) removal from non-biological processes was less than 16% in all amendments during the 40 first days of the experiments. According to the obtained results, biostimulation can enhance both mechanisms of biological Cr(VI) reduction and denitrification under anoxic conditions.
In Figure 4a,b, the results from the biotic control experiment (ANOX-B) are presented. For the first 30 days of the experiment, Cr(VI) reduction rates were on average 0.34 h−1 ± 0.01, while nitrate removal rates were 0.37 h−1 ± 0.07. The results indicate that both processes (Cr(VI reduction and denitrification) were possible without the addition of an external reductive agent and were carried out almost simultaneously.
By comparing the kCr(VI) rates in the ANOX-B experiment and the amendments where organic electron donors (molasses and EVO) were added (Figure 4a), it was observed that an increase in kCr(VI) rates in the molasses and EVO batch tests was achieved between the 10th and 30th day of the experiment. During the first 10 days, Cr(VI) reduction rates were similar in all three conditions studied (approximately 0.35 h−1). In contrast, denitrification during the first time interval (0–10 days) seemed to be 2.6–3.0 times faster than Cr(VI) reduction. Nitrate reduction rates obtained in the ANOX-M experiment were 1.07 h−1 ± 0.63 and in the ANOX-E 1.03 h−1 ± 0.62 (Figure 4b). This finding that denitrification rates are higher than Cr(VI) reduction rates is in agreement with published data [86]. However, for the remainder of the experiment, this difference in reduction rates was diminished, with both redox reactions occurring almost simultaneously, although nitrate removal was in the majority of the cases still slightly faster. Similar results, i.e., that Cr(VI) and nitrate microbially remediated reduction can occur concomitantly in the presence of the same electron donor, have also been obtained by Hu et al. [87]. It should also be mentioned that after the 10th day, NO3-N concentration was increased, and it has been reported that in certain cases and within limits, nitrate addition could even be beneficial for Cr(VI) reduction [50,52,88].
Figure 5a,b and Figure 6a,b illustrate the results of the combined biological and chemical Cr(VI) and nitrate removal. In the case of Cr reduction, the simultaneous addition of Fe2+ and a carbon source increases the removal rates by 2.5–3.0 times during the first 10 days of the experiment. The rates obtained with Fe2+ addition were 1.03 h−1 ± 0.59 compared to 0.34 h−1 ± 0.17 with molasses as the carbon source (Figure 5a) and 0.99 h−1 ± 0.48 to 0.39 h−1 ± 0.24 with EVO (Figure 6a). Between the 10th and 60th day of the experiments, the kinetic rates of the bottles amended with carbon source and Fe2+ were similar to those containing only the corresponding organic energy source, as was also the case in the anaerobic experiment.
As mentioned in Section 3.2, the redox conditions favor the formation of reactive Fe(II)—containing minerals. Several studies have suggested that Fe(II) in groundwater (dissolved or in iron-containing minerals) could promote the reduction of NO3-N and NO2-N in contaminated aquifers, with the latter two acting as potential electron acceptors for Fe(II) oxidation [89,90,91]. Denitrification rates with the addition of an iron source (Figure 5b and Figure 6b) were slightly higher than the rates in the amendments where only an organic e− donor was added, although the difference was insignificant, with the exception of rates obtained between the 30th and 40th day. It can be concluded that during the first 10 days of the experiment, the majority of Fe(II) was used for Cr(VI) reduction, while for the remainder, Fe(II)-containing minerals promoted nitrate reduction.
Nitrite measurements showed that during the first 30 days of the experiment, NO2-N was below the detection limit, indicating that nitrite was just an intermediate product and complete denitrification was taking place. However, after the 40th day of the experiment and until termination, NO2-N concentration in all biotic amendments was measured between 1.0–6.0 mg/L. The presence of nitrite at such high concentrations indicated that NO2-N reduction could not be completed, possibly due to an electron donor shortage [92]. After the 30th day, both processes were significantly slower and were carried out through the endogenous decay path, as indicated by Figure 4, Figure 5 and Figure 6.
3.4. Evaluation of Cr(VI) Reduction under Different Sulfate Reducing Conditions
Hexavalent Cr reduction rates were evaluated under low and high sulfate concentrations for a period of 60 days. Initial Cr(VI) concentration was 2000 μg/L for the first 10 days of the experiment and 3000 μg/L for the remainder. The bottles were spiked with Cr(VI) every 2–3 days, while sulfate concentration was measured and kept at 100 mg/L (S100) and 400 mg/L (S400) throughout each experiment. Sterile amendments showed that Cr(VI) removal rates were kSTERILE,S100 = 0.071 h−1 ± 0.03 and kSTERILE,S400 = 0.076 h−1 ± 0.02. These rates indicate that Cr(VI) removal from non-biological processes was less than 11% throughout the 60-day observation period in the amendments where electron donors were added in the S100 experiment, while in the S400 experiment, their contribution was less than 16% during the first 30 experimental days.
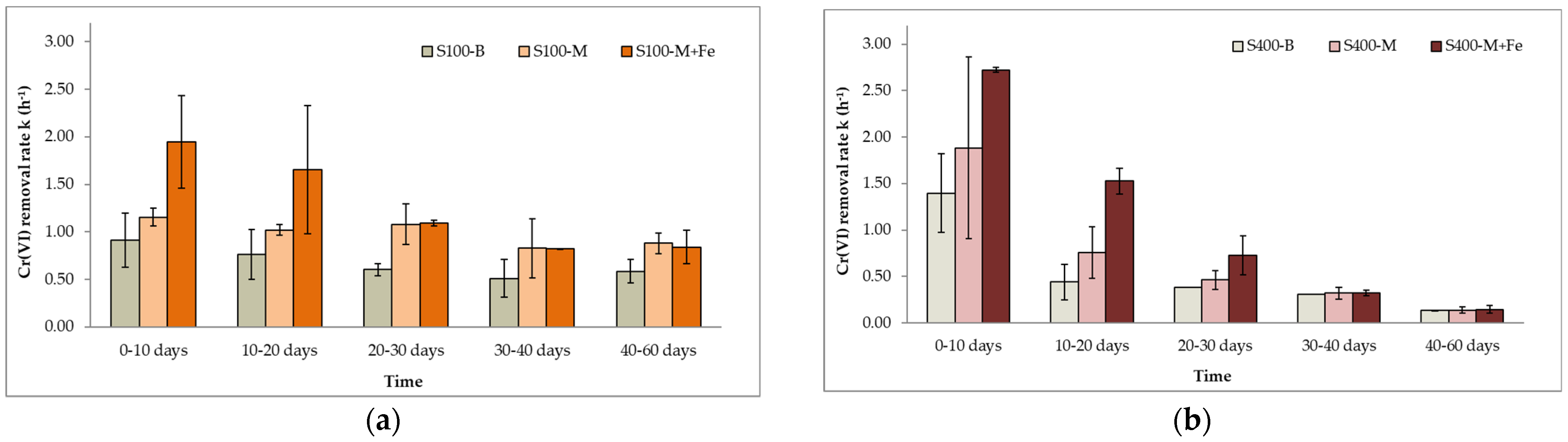
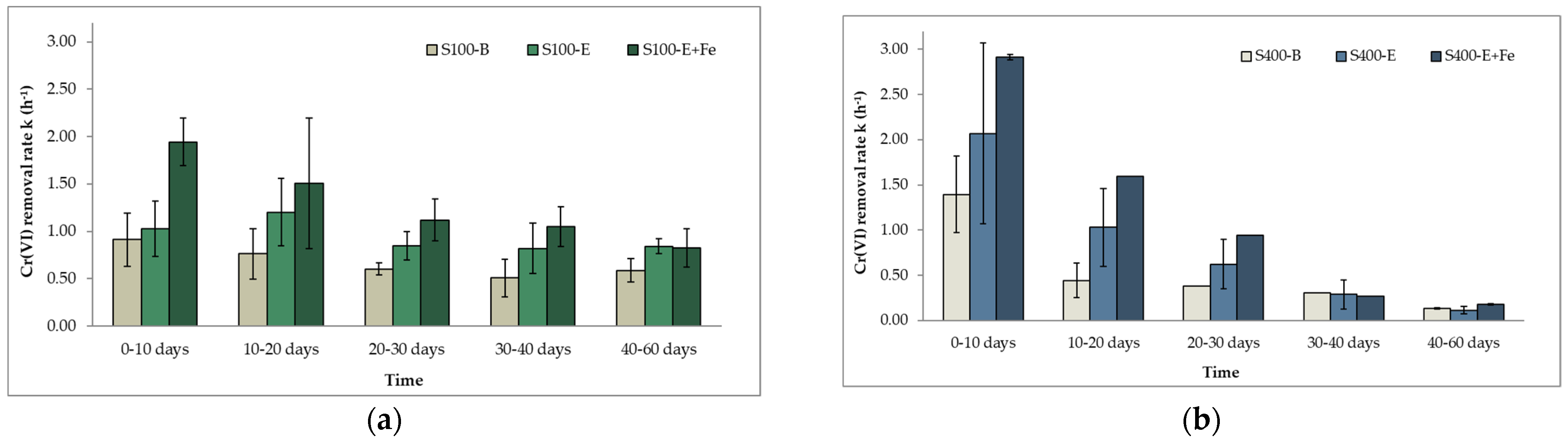
In Figure 7a,b Cr(VI) removal rates from the biotic control experiment and the amendments containing molasses and EVO for both sulfate concentrations are presented. Cr(VI) removal rates without the addition of an organic e− donor were kS100-B = 0.91 h−1 ± 0.03 and kS400-B = 1.39 h−1 ± 0.03 during the first 10 experimental days. During the next period (days 10–60), both reduction rates were slower. In the S100 experiment, kS100-B decreased by 48% and was on average 0.61 h−1 ± 0.11 throughout the experiment without significant fluctuations. In contrast, in the S400 experiment, kS400-B was 0.31 h−1 ± 0.13, indicating a decline of 3.43 times. Nonetheless, Cr(VI) reduction was still possible in both cases through (a) endogenous decay and (b) direct reduction by the produced sulfide from sulfate reduction. The addition of an organic electron donor (molasses or EVO) increased Cr(VI) reduction rates 1.2–2.3 times under both sulfate-reducing conditions. No significant differences between the reduction rates of batch tests amended with molasses and EVO were reported.
Comparing the Cr(VI) removal rates obtained from the S100 and S400 experiments, there is a noticeable difference in their progression. In the S100 experiment (Figure 7a), the rates are almost stable for the duration of the experiment. However, in the S400 experiment (Figure 7b), there is a steep decline between the first- and second-time intervals (0–10 days and 10–20 days, respectively), followed by a slighter decrease. Furthermore, kS400 reduction rates on days 0–10 were approximately 1.5 higher than their counterparts in the S100 experiment, while for the next 30 days (days 10–40), kS400 rates were 10–70% lower than kS100. The addition of an electron donor in the S400 experiment promoted the completion of microbial metabolic processes at a higher rate, including sulfate and Cr(VI) reduction, which led, however, to the consumption of the electron donors at a faster pace. Observation of the kS100 removal rates beyond the 60-day period (results not shown) indicates that reduction of Cr(VI) through the endogenous decay path occurred 80 days after the addition of the electron donor. In contrast, in the S400 experiment, endogenous decay was reached faster in approximately only 40 days. It can be concluded that the overall Cr(VI) reduction proceeded at a slightly higher rate in the S100 experiment.
During the first 10 days of the experiments, measured sulfide concentrations (Table 5) in the S400 experiment ranged from 12.2 mg/L to 19.0 mg/L in amendments where an organic electron donor was added. Thereafter sulfide concentration decreased following the pattern described above. In the S100 biotic amendments, sulfide concentrations were on average 7.7 mg/L in the first time interval but did not significantly decline during the rest of the experiment. Comparing the sulfide concentrations in amendments where electron donors were added during days 0–10, sulfide concentrations in the S400 experiment were, on average, two times higher than the ones in the S100. As already mentioned, sulfide is a reductant of Cr(VI) to Cr(III) at the pH range of the microcosms experiments. As reported by Kim et al. [93], this chemical reaction can take place at a pH higher than 8, while Cr(VI) reduction rate is positively correlated to initial sulfide concentration. Therefore the possibility of abiotic Cr(VI) reduction in the presence of sulfate-reducing conditions where sulfides act as electron acceptors should not be excluded.
Cr(VI) reduction rates with the addition of ferrous sulfate are presented in Figure 8 and Figure 9. The addition of a ferrous source can increase Cr(VI) reduction rates by 41%–99% during the first 20–30 days of both experiments since redox conditions favor the creation of reactive minerals, as described in Section 3.1 and Section 3.2. For the rest of the experiment, reduction rates were similar to the ones containing only the corresponding organic electron donor.
3.5. Comparison of Cr(VI) Reduction Rates under Different Reducing Conditions
An important aim of this experimental work was the evaluation of Cr(VI) microbial reduction under different reducing and limiting electron donor conditions and their effect on in situ bioremediation applications. Cr(VI) reduction rates obtained under anaerobic and anoxic conditions were comparable in all amendments, even with the increase in initial Cr(VI) concentration at 3 mg/L in the anoxic experiment. Furthermore, biostimulation appears to favor Cr(VI) reduction under sulfate reduction conditions. In the S100 experiment, Cr(VI) reduction was in the majority of the amendments at least two times faster than their counterpart’s in the anaerobic experiment throughout the 60-day observation period and in the biotic control batch test four to seven times faster. Similar results were also obtained in the S400 experiment during the 30 first experimental days, although for the remainder, Cr(VI) removal rates did not exhibit significant differences compared to their anaerobic counterparts. It is not clear whether microorganisms directly reduce Cr(VI) via an enzymatic mechanism or whether a significant amount of Cr(VI) reduction proceeds indirectly by microorganisms reducing sulfates which then abiotically reduce Cr(VI). In either event, it is clear that in the sulfate range studied, the addition of an organic electron donor is an effective mechanism to stimulate Cr(VI) reduction in the subsurface under sulfate-reducing conditions. Overall, the most favorable conditions for microbial remediated Cr(VI) reduction in terms of reduction rates and longevity of the process are low sulfate-reducing conditions (100 mg/L).
The obtained results have significant importance for in situ Cr(VI) bioremediation applications. Cr(VI) bioreduction under sulfate-reducing conditions is the most favorable environment since, even without the addition of a carbon donor, Cr(VI) removal can be carried out through direct chemical reduction by bio-produced sulfide. However, special emphasis should be placed on the creation of a balanced ratio between provided COD and sulfate concentration to ensure the longevity of the process. Anoxic environments would require more frequent injections of organic electron donors since the reduction of nitrates is a high-energy demanding process.
4. Conclusions
The aim of this study was to examine the effect of two different organic carbon sources and an iron source on biological hexavalent chromium reduction under nitrate-rich, complete anaerobic, and sulfate-reducing conditions. The results of the experiments can be summarized in the following conclusions.
Biostimulation by the addition of an organic electron donor can increase the rate of Cr(VI) reduction up to six times in comparison to the natural soil capacity rates under anaerobic conditions. Comparing the two Cr(VI) reduction rates obtained by the two different carbon sources, they did not exhibit any significant differences. Furthermore, a combination of organic carbon and iron electron donors can further increase the removal rate of hexavalent chromium through biotic and abiotic processes. However, this biostimulatιon did not have a long-lasting effect on Cr(VI) reduction and gradually diminished following the depletion of the electron donors.
Cr(VI) reduction is also feasible in the presence of nitrate ions and, according to the microcosm results, is carried out almost simultaneously with denitrification when electron donors are available. Microbes that have been acclimatized under anoxic conditions can completely mitigate water containing Cr(VI) and NO3-N concentrations, equal to 3 mg/L and 20 mg/L, respectively, through biostimulation, employing either an organic or a combination of organic and inorganic electron donors. However, following electron donor depletion, both Cr(VI) reduction and denitrification rates are significantly reduced.
Sulfate presence in a Cr(VI)-contaminated aquifer significantly improves microbially induced Cr(VI) reduction, even without the addition of an electron donor. However, at high sulfate concentrations (400 mg/L), Cr(VI) bioreduction is slightly impaired compared to lower sulfate concentrations (100 mg/L) due to the faster consumption of the organic electron donor.
Since anaerobic, anoxic, and sulfate-reducing conditions are quite common in groundwater aquifers, the findings of this study provide significant information on Cr(VI) bioreduction under these redox conditions through biostimulation and could be used as a basis for a possible larger-scale application for bioremediation purposes.
Author Contributions
A.G.: conceptualization, methodology, experimental investigation, experimental work and data analysis, writing. C.N.: conceptualization, supervision, writing review and editing. P.A. and A.V.: experimental work and data analyses. D.M.: conceptualization, supervision, data analyses, writing review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available within the article. For any additional data the corresponding authors may be contacted.
Acknowledgments
This research is co-financed by Greece and the European Union (European Social Fund-ESF) through the Operational Programme «Human Resources Development, Education and Lifelong Learning» in the context of the project Strengthening Human Resources Research Potential via Doctorate Research (MIS-5000432), implemented by the State Scholarships Foundation (ΙΚΥ).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Laxmi, V.; Kaushik, G. Toxicity of Hexavalent Chromium in Environment, Health Threats, and Its Bioremediation and Detoxification from Tannery Wastewater for Environmental Safety. In Bioremediation of Industrial Waste for Environmental Safety; Saxena, G., Bharagava, R.N., Eds.; Springer: Singapore, 2020; pp. 223–243. [Google Scholar] [CrossRef]
- Kapoor, R.T.; Bani Mfarrej, M.F.; Alam, P.; Rinklebe, J.; Ahmad, P. Accumulation of chromium in plants and its repercussion in animals and humans. Environ. Pollut. 2022, 301, 119044. [Google Scholar] [CrossRef] [PubMed]
- EPA. National Primary Drinking Water Regulations; EPA 816-F-09-004; EPA: Washington, DC, USA, 2009.
- Vaiopoulou, E.; Gikas, P. Regulations for chromium emissions to the aquatic environment in Europe and elsewhere. Chemosphere 2020, 254, 126876. [Google Scholar] [CrossRef]
- Chen, Y.G.; Sofińska-Chmiel, W.; Lv, G.Y.; Kołodyńska, D.; Chen, S.H. Application of modern research methods for the physicochemical characterization of ion exchangers. Materials 2021, 14, 7067. [Google Scholar] [CrossRef]
- Djedidi, Z.; Bouda, M.; Souissi, M.A.; Cheikh, R.B.; Mercier, G.; Tyagi, R.D.; Blais, J.F. Metals removal from soil, fly ash and sewage sludge leachates by precipitation and dewatering properties of the generated sludge. J. Hazard. Mater. 2009, 172, 1372–1382. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.; Alharbi, N.S.; Ren, X.; Chen, C. A comprehensive review on emerging natural and tailored materials for chromium-contaminated water treatment and environmental remediation. J. Environ. Chem. Eng. 2022, 10, 107325. [Google Scholar] [CrossRef]
- Ren, X.; Zhao, C.; Du, S.; Wang, T.; Luan, Z.; Wang, J.; Hou, D. Fabrication of asymmetric poly (m-phenylene isophthalamide) nanofiltration membrane for chromium(VI) removal. J. Environ. Sci. 2010, 22, 1335–1341. [Google Scholar] [CrossRef]
- Owlad, M.; Aroua, M.K.; Daud, W.A.W.; Baroutian, S. Removal of hexavalent chromium-contaminated water and wastewater: A review. Water Air Soil Pollut. 2009, 200, 59–77. [Google Scholar] [CrossRef]
- Nur-E.-Alam, M.; Mia, M.A.S.; Ahmad, F.; Rahman, M.M. An overview of chromium removal techniques from tannery effluent. Appl. Water Sci. 2020, 10, 205. [Google Scholar] [CrossRef]
- Hininger, I.; Benaraba, R.; Osman, M.; Faure, H.; Marie Roussel, A.; Anderson, R.A. Safety of trivalent chromium complexes: No evidence for DNA damage in human HaCaT keratinocytes. Free Radic. Biol. Med. 2007, 42, 1759–1765. [Google Scholar] [CrossRef]
- Sharma, A.; Kapoor, D.; Wang, J.; Shahzad, B.; Kumar, V.; Bali, A.S.; Jasrotia, S.; Zheng, B.; Yuan, H.; Yan, D. Chromium bioaccumulation and its impacts on plants: An overview. Plants 2020, 9, 100. [Google Scholar] [CrossRef] [Green Version]
- Rahman, Z.; Thomas, L. Chemical-Assisted Microbially Mediated Chromium (Cr) (VI) Reduction Under the Influence of Various Electron Donors, Redox Mediators, and Other Additives: An Outlook on Enhanced Cr(VI) Removal. Front. Microbiol. 2021, 11, 619766. [Google Scholar] [CrossRef]
- Huang, Y.; Zeng, Q.; Hu, L.; Zhong, H.; He, Z. Bioreduction performances and mechanisms of Cr(VI) by Sporosarcina saromensis W5, a novel Cr(VI)-reducing facultative anaerobic bacteria. J. Hazard. Mater. 2021, 413, 125411. [Google Scholar] [CrossRef] [PubMed]
- Brookshaw, D.R.; Coker, V.S.; Lloyd, J.R.; Vaughan, D.J.; Pattrick, R.A.D. Redox interactions between Cr(VI) and Fe(II) in bioreduced biotite and chlorite. Environ. Sci. Technol. 2014, 48, 11337–11342. [Google Scholar] [CrossRef] [PubMed]
- Faybishenko, B.; Hazen, T.C.; Long, P.E.; Brodie, E.L.; Conrad, M.E.; Hubbard, S.S.; Christensen, J.N.; Joyner, D.; Borglin, S.E.; Chakraborty, R.; et al. In situ long-term reductive bioimmobilization of Cr(VI) in groundwater using hydrogen release compound. Environ. Sci. Technol. 2008, 42, 8478–8485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Fan, M.; Liu, L.; Chang, J.; Zhang, J. Treatment of high-concentration chromium-containing wastewater by sulfate-reducing bacteria acclimated with ethanol. Water Sci. Technol. 2019, 80, 2362–2372. [Google Scholar] [CrossRef] [PubMed]
- Fernández, P.M.; Viñarta, S.C.; Bernal, A.R.; Cruz, E.L.; Figueroa, L.I.C. Bioremediation strategies for chromium removal: Current research, scale-up approach and future perspectives. Chemosphere 2018, 208, 139–148. [Google Scholar] [CrossRef]
- Song, X.; Wang, Q.; Jin, P.; Chen, X.; Tang, S.; Wei, C.; Li, K.; Ding, X.; Tang, Z.; Fu, H. Enhanced biostimulation coupled with a dynamic groundwater recirculation system for Cr(VI) removal from groundwater: A field-scale study. Sci. Total Environ. 2021, 772, 145495. [Google Scholar] [CrossRef]
- Abascal, E.; Gómez-Coma, L.; Ortiz, I.; Ortiz, A. Global diagnosis of nitrate pollution in groundwater and review of removal technologies. Sci. Total Environ. 2022, 810, 152233. [Google Scholar] [CrossRef]
- Romanelli, A.; Soto, D.X.; Matiatos, I.; Martínez, D.E.; Esquius, S. A biological and nitrate isotopic assessment framework to understand eutrophication in aquatic ecosystems. Sci. Total Environ. 2020, 715, 136909. [Google Scholar] [CrossRef]
- Fewtrell, L. Drinking-water nitrate, methemoglobinemia, and global burden of disease: A discussion. Environ. Health Perspect. 2004, 112, 1371–1374. [Google Scholar] [CrossRef] [Green Version]
- Bahadoran, Z.; Mirmiran, P.; Ghasemi, A.; Kabir, A.; Azizi, F.; Hadaegh, F. Is dietary nitrate/nitrite exposure a risk factor for development of thyroid abnormality? A systematic review and meta-analysis. Nitric Oxide-Biol. Chem. 2015, 47, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Middleton, S.S.; Latmani, R.B.; Mackey, M.R.; Ellisman, M.H.; Tebo, B.M.; Criddle, C.S. Cometabolism of Cr(VI) by Shewanella oneidensis MR-1 produces cell-associated reduced chromium and inhibits growth. Biotechnol. Bioeng. 2003, 83, 627–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, R.; Wang, B.; Liu, G.; Wang, Y.; Zhou, J.; Wang, J. Bioreduction of Cr(VI) by acinetobacter sp. WB-1 during simultaneous nitrification/denitrification process. J. Chem. Technol. Biotechnol. 2017, 92, 649–656. [Google Scholar] [CrossRef]
- He, D.; Zheng, M.; Ma, T.; Li, C.; Ni, J. Interaction of Cr(VI) reduction and denitrification by strain Pseudomonas aeruginosa PCN-2 under aerobic conditions. Bioresour. Technol. 2015, 185, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Sahinkaya, E.; Kilic, A. Heterotrophic and elemental-sulfur-based autotrophic denitrification processes for simultaneous nitrate and Cr(VI) reduction. Water Res. 2014, 50, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Liao, R.; Zhang, X.X.; Wang, Y.; Wang, Z.; Shi, P.; Liu, B.; Li, A. Metagenomic insights into Cr(VI) effect on microbial communities and functional genes of an expanded granular sludge bed reactor treating high-nitrate wastewater. Water Res. 2015, 76, 43–52. [Google Scholar] [CrossRef]
- Yu, X.; Jiang, Y.; Huang, H.; Shi, J.; Wu, K.; Zhang, P.; Lv, J.; Li, H.; He, H.; Liu, P.; et al. Simultaneous aerobic denitrification and Cr(VI) reduction by Pseudomonas brassicacearum LZ-4 in wastewater. Bioresour. Technol. 2016, 221, 121–129. [Google Scholar] [CrossRef]
- Zhong, L.; Lai, C.Y.; Shi, L.D.; Wang, K.D.; Dai, Y.J.; Liu, Y.W.; Ma, F.; Rittmann, B.E.; Zheng, P.; Zhao, H.P. Nitrate effects on chromate reduction in a methane-based biofilm. Water Res. 2017, 115, 130–137. [Google Scholar] [CrossRef]
- Chang, I.S.; Kim, B.H. Effect of sulfate reduction activity on biological treatment of hexavalent chromium [Cr(VI)] contaminated electroplating wastewater under sulfate-rich condition. Chemosphere 2007, 68, 218–226. [Google Scholar] [CrossRef]
- Samborska, K.; Halas, S.; Bottrell, S.H. Sources and impact of sulphate on groundwaters of Triassic carbonate aquifers, Upper Silesia, Poland. J. Hydrol. 2013, 486, 136–150. [Google Scholar] [CrossRef]
- Torres-Martínez, J.A.; Mora, A.; Knappett, P.S.K.; Ornelas-Soto, N.; Mahlknecht, J. Tracking nitrate and sulfate sources in groundwater of an urbanized valley using a multi-tracer approach combined with a Bayesian isotope mixing model. Water Res. 2020, 182, 115962. [Google Scholar] [CrossRef] [PubMed]
- Sadeghalvad, B.; Khorshidi, N.; Azadmehr, A.; Sillanpää, M. Sorption, mechanism, and behavior of sulfate on various adsorbents: A critical review. Chemosphere 2021, 263, 128064. [Google Scholar] [CrossRef] [PubMed]
- Márquez-Reyes, J.M.; López-Chuken, U.J.; Valdez-González, A.; Luna-Olvera, H.A. Removal of chromium and lead by a sulfate-reducing consortium using peat moss as carbon source. Bioresour. Technol. 2013, 144, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Wei, L.; Liu, R.; Jiang, F.; Hao, X.; Chen, G.H. An exploratory study on the pathways of Cr (VI) reduction in sulfate-reducing Up-flow Anaerobic Sludge Bed (UASB) reactor. Sci. Rep. 2016, 6, 23694. [Google Scholar] [CrossRef] [Green Version]
- Goulhen, F.; Gloter, A.; Guyot, F.; Bruschi, M. Cr(VI) detoxification by Desulfovibrio vulgaris strain Hildenborough: Microbe-metal interactions studies. Appl. Microbiol. Biotechnol. 2006, 71, 892–897. [Google Scholar] [CrossRef]
- Lv, P.L.; Zhong, L.; Dong, Q.Y.; Yang, S.L.; Shen, W.W.; Zhu, Q.S.; Lai, C.Y.; Luo, A.C.; Tang, Y.; Zhao, H.P. The effect of electron competition on chromate reduction using methane as electron donor. Environ. Sci. Pollut. Res. 2018, 25, 6609–6618. [Google Scholar] [CrossRef]
- Kazakis, N.; Kantiranis, N.; Kalaitzidou, K.; Kaprara, E.; Mitrakas, M.; Frei, R.; Vargemezis, G.; Vogiatzis, D.; Zouboulis, A.; Filippidis, A. Environmentally available hexavalent chromium in soils and sediments impacted by dispersed fly ash in Sarigkiol basin (Northern Greece). Environ. Pollut. 2018, 235, 632–641. [Google Scholar] [CrossRef]
- Dermatas, D.; Panagiotakis, I.; Mpouras, T.; Tettas, K. The Origin of Hexavalent Chromium as a Critical Parameter for Remediation of Contaminated Aquifers. Bull. Environ. Contam. Toxicol. 2017, 98, 331–337. [Google Scholar] [CrossRef]
- Galani, A.; Mamais, D.; Noutsopoulos, C.; Anastopoulou, P.; Varouxaki, A. Biotic and Abiotic Biostimulation for the Reduction of Hexavalent Chromium in Contaminated Aquifers. Water 2022, 14, 89. [Google Scholar] [CrossRef]
- Tandoi, V.; DiStefano, T.D.; Bowser, P.A.; Gossett, J.M.; Zinder, S.H. Reductive Dehalogenation of Chlorinated Ethenes and Halogenated Ethanes by a High-Rate Anaerobic Enrichment Culture. Environ. Sci. Technol. 1994, 28, 973–979. [Google Scholar] [CrossRef]
- Widdel, F. Microbiology and Ecology of Sulfate-and Sulfur-Reducing Bacteria. In Biology of Anaerobic Microorganisms; Zehnder, A.J.B., Ed.; John Wiley & Sons: New York, USA, 1988; pp. 469–585. [Google Scholar]
- Balch, W.E.; Fox, G.E.; Magrum, L.J.; Woese, C.R.; Wolfe, R.S. Methanogens: Reevaluation of a unique biological group. Microbiol. Rev. 1979, 43, 260–296. [Google Scholar] [CrossRef] [PubMed]
- USEPA Fate. Transport and Transformation Test Guidelines OPPTS 835.3300 Soil Biodegradation; EPA: Washington, DC, USA, 1998.
- Yang, X.; Liu, P.; Yao, M.; Sun, H.; Liu, R.; Xie, J.; Zhao, Y. Mechanism and enhancement of Cr(VI) contaminated groundwater remediation by molasses. Sci. Total Environ. 2021, 780, 146580. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Lan, J.; Du, Y.; Guo, L.; Du, D.; Chen, S.; Ye, H.; Zhang, T.C. Chromium(VI) bioreduction and removal by Enterobacter sp. SL grown with waste molasses as carbon source: Impact of operational conditions. Bioresour. Technol. 2020, 302, 121974. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Lan, J.; Du, Y.; Li, Z.; Liao, X.; Du, D.; Ye, H.; Zhang, T.C.; Chen, S. Efficient removal of heavy metals by synergistic actions of microorganisms and waste molasses. Bioresour. Technol. 2020, 302, 122797. [Google Scholar] [CrossRef]
- Harkness, M.; Fisher, A. Use of emulsified vegetable oil to support bioremediation of TCE DNAPL in soil columns. J. Contam. Hydrol. 2013, 151, 16–33. [Google Scholar] [CrossRef]
- Ding, L.; Song, J.; Huang, D.; Lei, J.; Li, X.; Sun, J. Simultaneous removal of nitrate and hexavalent chromium in groundwater using indigenous microorganisms enhanced by emulsified vegetable oil: Interactions and remediation threshold values. J. Hazard. Mater. 2021, 406, 124708. [Google Scholar] [CrossRef]
- Wen, C.; Sheng, H.; Ren, L.; Dong, Y.; Dong, J. Study on the removal of hexavalent chromium from contaminated groundwater using emulsified vegetable oil. Process Saf. Environ. Prot. 2017, 109, 599–608. [Google Scholar] [CrossRef]
- Kourtev, P.S.; Nakatsu, C.H.; Konopka, A. Inhibition of nitrate reduction by chromium(VI) in anaerobic soil microcosms. Appl. Environ. Microbiol. 2009, 75, 6249–6257. [Google Scholar] [CrossRef] [Green Version]
- Perraki, M.; Vasileiou, E.; Bartzas, G. Tracing the origin of chromium in groundwater: Current and new perspectives. Curr. Opin. Environ. Sci. Health 2021, 22, 100267. [Google Scholar] [CrossRef]
- Panousi, E.; Mamais, D.; Noutsopoulos, C.; Antoniou, K.; Koutoula, K.; Mastrantoni, S.; Koutsogiannis, C.; Gkioni, A. Biological treatment of groundwater with a high hexavalent chromium content under anaerobic and anoxic conditions. J. Chem. Technol. Biotechnol. 2016, 91, 1681–1687. [Google Scholar] [CrossRef]
- USEPA Method 7196A for Chromium Hexavalent (colorimetric). USEPA Method 7196A; 1992; pp. 1–6. Available online: https://www.epa.gov/sites/default/files/2015-12/documents/7196a.pdf (accessed on 20 June 2022).
- Baird, R.; Bridgewater, L. 5220 CHEMICAL OXYGEN DEMAND (COD). In Standard Methods for the Examination of Water and Wastewater, 23rd ed.; American Public Health Association: Washington, DC, USA, 2017. [Google Scholar]
- Baird, R.; Bridgewater, L. 3500-Fe IRON. In Standard Methods for the Examination of Water and Wastewater, 23rd ed.; American Public Health Association: Washington, DC, USA, 2017. [Google Scholar]
- Amann, R.I.; Krumholz, L.; Stahl, D.A. Fluorescent-oligonucleotide probing of whole cells for determinative, phylogenetic, and environmental studies in microbiology. J. Bacteriol. 1990, 172, 762–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, D.A.; Amann, R. Development and application of nucleic acid probes in bacterial systematics. In Nucleic Acid Techniques in Bacterial Systematics, 1st ed.; Stackebrandt, E., Goodfellow, M., Eds.; Wiley & Sons Ltd.: Chichester, UK, 1991; pp. 205–248. [Google Scholar]
- Raskin, L.; Stromley, J.M.; Rittmann, B.E.; Stahl, D.A. Group-specific 16S rRNA hybridization probes to describe natural communities of methanogens. Appl. Environ. Microbiol. 1994, 60, 1232–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Küsel, K.; Pinkart, H.C.; Drake, H.L.; Devereux, R. Acetogenic and sulfate-reducing bacteria inhabiting the rhizoplane and deep cortex cells of the sea grass Halodule wrightii. Appl. Environ. Microbiol. 1999, 65, 5117–5123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, D.E.; Giloteaux, L.; Barlett, M.; Chavan, M.A.; Smith, J.A.; Williams, K.H.; Wilkins, M.; Long, P.; Lovley, D.R. Molecular analysis of the In situ growth rates of subsurface geobacter species. Appl. Environ. Microbiol. 2013, 79, 1646–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manz, W.; Amann, R.; Ludwig, W.; Wagner, M.; Schleifer, K. Phylogenetic oligodeoxynucleotide probes for the major subclasses of proteobacteria: Problems and solutions. Syst. Appl. Microbiol. 1992, 15, 593–600. [Google Scholar] [CrossRef]
- Loy, A.; Lehner, A.; Lee, N.; Adamczyk, J.; Meier, H.; Ernst, J.; Schleifer, K.H.; Wagner, M. Oligonucleotide microarray for 16S rRNA gene-based detection of all recognized lineages of sulfate-reducing prokaryotes in the environment. Appl. Environ. Microbiol. 2002, 68, 5064–5081. [Google Scholar] [CrossRef] [Green Version]
- Lücker, S.; Steger, D.; Kjeldsen, K.U.; MacGregor, B.J.; Wagner, M.; Loy, A. Improved 16S rRNA-targeted probe set for analysis of sulfate-reducing bacteria by fluorescence in situ hybridization. J. Microbiol. Methods 2007, 69, 523–528. [Google Scholar] [CrossRef]
- Nielsen, J.L.; Juretschko, S.; Wagner, M.; Nielsen, P.H. Abundance and phylogenetic affiliation of iron reducers in activated sludge as assessed by fluorescence in situ hybridization and microautoradiography. Appl. Environ. Microbiol. 2002, 68, 4629–4636. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.H.; Chen, C.C.; Ou, J.H.; Sheu, Y.T.; Hou, D.; Kao, C.M. Bioremediation of hexavalent-chromium contaminated groundwater: Microcosm, column, and microbial diversity studies. Chemosphere 2022, 295, 133877. [Google Scholar] [CrossRef]
- Qu, W.; Suo, L.; Liu, R.; Liu, M.; Zhao, Y.; Xia, L.; Fan, Y.; Zhang, Q.; Gao, Z. Influence of Temperature on Denitrification and Microbial Community Structure and Diversity: A Laboratory Study on Nitrate Removal from Groundwater. Water 2022, 14, 436. [Google Scholar] [CrossRef]
- Vavilin, V.A.; Vasiliev, V.B.; Rytov, S.V.; Ponomarev, A.V. Self-oscillating coexistence of methanogens and sulfate-reducers under hydrogen sulfide inhibition and the pH-regulating effect. Bioresour. Technol. 1994, 49, 105–119. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of anaerobic digestion process: A review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef] [PubMed]
- Dar, S.A.; Kleerebezem, R.; Stams, A.J.M.; Kuenen, J.G.; Muyzer, G. Competition and coexistence of sulfate-reducing bacteria, acetogens and methanogens in a lab-scale anaerobic bioreactor as affected by changing substrate to sulfate ratio. Appl. Microbiol. Biotechnol. 2008, 78, 1045–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeyasingh, J.; Philip, L. Bioremediation of chromium contaminated soil: Optimization of operating parameters under laboratory conditions. J. Hazard. Mater. 2005, 118, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Michailides, M.K.; Tekerlekopoulou, A.G.; Akratos, C.S.; Coles, S.; Pavlou, S.; Vayenas, D. V Molasses as an efficient low-cost carbon source for biological Cr(VI) removal. J. Hazard. Mater. 2015, 281, 95–105. [Google Scholar] [CrossRef]
- Dong, J.; Yu, J.; Bao, Q. Simulated reactive zone with emulsified vegetable oil for the long-term remediation of Cr(VI)-contaminated aquifer: Dynamic evolution of geological parameters and groundwater microbial community. Environ. Sci. Pollut. Res. 2018, 25, 34392–34402. [Google Scholar] [CrossRef]
- Ball, J.W.; Izbicki, J.A. Occurrence of hexavalent chromium in ground water in the western Mojave Desert, California. Appl. Geochem. 2004, 19, 1123–1135. [Google Scholar] [CrossRef]
- Kumar, A.R.; Riyazuddin, P. Chromium speciation in a contaminated groundwater: Redox processes and temporal variability. Environ. Monit. Assess. 2011, 176, 647–662. [Google Scholar] [CrossRef]
- Izbicki, J.A.; Wright, M.T.; Seymour, W.A.; McCleskey, R.B.; Fram, M.S.; Belitz, K.; Esser, B.K. Cr(VI) occurrence and geochemistry in water from public-supply wells in California. Appl. Geochem. 2015, 63, 203–217. [Google Scholar] [CrossRef] [Green Version]
- Wielinga, B.; Mizuba, M.M.; Hansel, C.M.; Fendorf, S. Iron promoted reduction of chromate by dissimilatory iron-reducing bacteria. Environ. Sci. Technol. 2001, 35, 522–527. [Google Scholar] [CrossRef]
- Fendorf, S.; Wielinga, B.W.; Hansel, C.M. Chromium Transformations in Natural Environments: The Role of Biological and Abiological Processes in Chromium(VI) Reduction. Int. Geol. Rev. 2000, 42, 691–701. [Google Scholar] [CrossRef]
- Papassiopi, N.; Vaxevanidou, K.; Christou, C.; Karagianni, E.; Antipas, G.S.E. Synthesis, characterization and stability of Cr(III) and Fe(III) hydroxides. J. Hazard. Mater. 2014, 264, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Génin, J.M.R.; Refait, P.H.; Abdelmoula, M. Green rusts and their relationship to iron corrosion; a key role in microbially influenced corrosion. Hyperfine Interact. 2002, 119–131. [Google Scholar] [CrossRef]
- Cundy, A.B.; Hopkinson, L.; Whitby, R.L.D. Use of iron-based technologies in contaminated land and groundwater remediation: A review. Sci. Total Environ. 2008, 400, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Gheju, M.; Balcu, I. Removal of chromium from Cr(VI) polluted wastewaters by reduction with scrap iron and subsequent precipitation of resulted cations. J. Hazard. Mater. 2011, 196, 131–138. [Google Scholar] [CrossRef]
- Borch, T.; Kretzschmar, R.; Skappler, A.; Van Cappellen, P.; Ginder-Vogel, M.; Voegelin, A.; Campbell, K. Biogeochemical redox processes and their impact on contaminant dynamics. Environ. Sci. Technol. 2010, 44, 15–23. [Google Scholar] [CrossRef]
- Bryce, C.; Blackwell, N.; Schmidt, C.; Otte, J.; Huang, Y.M.; Kleindienst, S.; Tomaszewski, E.; Schad, M.; Warter, V.; Peng, C.; et al. Microbial anaerobic Fe(II) oxidation—Ecology, mechanisms and environmental implications. Environ. Microbiol. 2018, 20, 3462–3483. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Liu, T.; Chen, N.; Feng, C.; Lu, W.; Guo, H. Simultaneous bio-reduction of nitrate and Cr(VI) by mechanical milling activated corn straw. J. Hazard. Mater. 2022, 429, 128258. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, T.; Chen, N.; Feng, C. Changes in microbial community diversity, composition, and functions upon nitrate and Cr(VI) contaminated groundwater. Chemosphere 2022, 288, 132476. [Google Scholar] [CrossRef]
- He, Y.; Lin, H.; Luo, M.; Liu, J.; Dong, Y.; Li, B. Highly efficient remediation of groundwater co-contaminated with Cr(VI) and nitrate by using nano-Fe/Pd bimetal-loaded zeolite: Process product and interaction mechanism. Environ. Pollut. 2020, 263, 114479. [Google Scholar] [CrossRef]
- Margalef-Marti, R.; Carrey, R.; Benito, J.A.; Marti, V.; Soler, A.; Otero, N. Nitrate and nitrite reduction by ferrous iron minerals in polluted groundwater: Isotopic characterization of batch experiments. Chem. Geol. 2020, 548, 119691. [Google Scholar] [CrossRef]
- Smith, R.L.; Kent, D.B.; Repert, D.A.; Böhlke, J.K. Anoxic nitrate reduction coupled with iron oxidation and attenuation of dissolved arsenic and phosphate in a sand and gravel aquifer. Geochim. Cosmochim. Acta 2017, 196, 102–120. [Google Scholar] [CrossRef]
- Zhu, I.; Getting, T. A review of nitrate reduction using inorganic materials. Environ. Technol. Rev. 2012, 1, 46–58. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Hu, Y.; Chen, N.; He, Q.; Feng, C. High redox potential promotes oxidation of pyrite under neutral conditions: Implications for optimizing pyrite autotrophic denitrification. J. Hazard. Mater. 2021, 416, 125844. [Google Scholar] [CrossRef]
- Kim, C.; Zhou, Q.; Deng, B.; Thornton, E.C.; Xu, H. Chromium(VI) reduction by hydrogen sulfide in aqueous media: Stoichiometry and kinetics. Environ. Sci. Technol. 2001, 35, 2219–2225. [Google Scholar] [CrossRef]
Figure 1.
FISH analysis results describing the composition of the microbial communities participating in Cr(VI) reduction under anaerobic, anoxic and sulfate-reducing conditions.
Figure 1.
FISH analysis results describing the composition of the microbial communities participating in Cr(VI) reduction under anaerobic, anoxic and sulfate-reducing conditions.

Figure 2.
Cr(VI) kinetic rate constants (k) of microcosms during the anaerobic experiment (error bars are equal to standard deviation).
Figure 2.
Cr(VI) kinetic rate constants (k) of microcosms during the anaerobic experiment (error bars are equal to standard deviation).

Figure 3.
Cr(VI) kinetic rate constants (k) of microcosms with the addition of (a) molasses and a combination of molasses and Fe and (b) EVO and a combination of EVO and Fe, during the anaerobic experiment (error bars are equal to standard deviation).
Figure 3.
Cr(VI) kinetic rate constants (k) of microcosms with the addition of (a) molasses and a combination of molasses and Fe and (b) EVO and a combination of EVO and Fe, during the anaerobic experiment (error bars are equal to standard deviation).

Figure 4.
Cr(VI) (a) and nitrate kinetic rate constants (b) of microcosms during the anoxic experiment (error bars are equal to standard deviation).
Figure 4.
Cr(VI) (a) and nitrate kinetic rate constants (b) of microcosms during the anoxic experiment (error bars are equal to standard deviation).

Figure 5.
Comparison (a) of Cr(VI) kinetic rate constants and (b) of nitrate kinetic rate constants between molasses and a combination of molasses and Fe (error bars are equal to standard deviation).
Figure 5.
Comparison (a) of Cr(VI) kinetic rate constants and (b) of nitrate kinetic rate constants between molasses and a combination of molasses and Fe (error bars are equal to standard deviation).

Figure 6.
Comparison (a) of Cr(VI) kinetic rate constants and (b) of nitrate kinetic rate constants between EVO and a combination of EVO and Fe (error bars are equal to standard deviation).
Figure 6.
Comparison (a) of Cr(VI) kinetic rate constants and (b) of nitrate kinetic rate constants between EVO and a combination of EVO and Fe (error bars are equal to standard deviation).

Figure 7.
Cr(VI) kinetic rate constants of microcosms during the (a) S100 and (b) S400 experiment (error bars are equal to standard deviation).
Figure 7.
Cr(VI) kinetic rate constants of microcosms during the (a) S100 and (b) S400 experiment (error bars are equal to standard deviation).

Figure 8.
Cr(VI) kinetic rate constants (k) of microcosms with the addition of molasses and a combination of molasses and Fe during the (a) S100 and (b) S400 experiment (error bars are equal to standard deviation).
Figure 8.
Cr(VI) kinetic rate constants (k) of microcosms with the addition of molasses and a combination of molasses and Fe during the (a) S100 and (b) S400 experiment (error bars are equal to standard deviation).

Figure 9.
Cr(VI) kinetic rate constants (k) of microcosms with the addition of EVO and a combination of EVO and Fe during the (a) S100 and (b) S400 experiment (error bars are equal to standard deviation).
Figure 9.
Cr(VI) kinetic rate constants (k) of microcosms with the addition of EVO and a combination of EVO and Fe during the (a) S100 and (b) S400 experiment (error bars are equal to standard deviation).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Groundwater characteristics used for the development of the parent cultures.
| Parameter | Value |
|---|---|
| pH | 7.98 |
| ORP, mV | 76.50 |
| EC, μS/cm | 1865.00 |
| NO3-N, mg/L | 11.20 |
| NO2-N, mg/L | 0.14 |
| SO42−, mg/L | 51.00 |
| Cl−, mg/L | 342.00 |
| Cr6+, mg/L | <0.03 |
| CODs *, mg/L | <15.00 |
| Ca, mg/L | 68.00 |
| Mg, mg/L | 155.00 |
| K, mg/L | 1.40 |
| Hardness, mg CaCO3/L | 805.50 |
| Alkalinity, mg CaCO3/L | 382.00 |
* CODs: soluble chemical oxygen demand.
Table 2.
Experimental conditions studied in the anaerobic and anoxic microcosms tests.
| Experimental Set | Batch Test | Cr(VI) | NO3-N | Molasses | EVO | Fe2+ |
|---|---|---|---|---|---|---|
| mg/L | mg/L | mg/L CODt * | mg/L CODt * | mg/L | ||
| Anaerobic | ANAER-S (Sterile) | 2 | - | - | - | - |
| ANAER-B (Biotic control) | - | - | - | - | ||
| ANAER-M | - | 100 | - | - | ||
| ANAER-M+Fe | - | 100 | - | 8 | ||
| ANAER-E | - | - | 100 | - | ||
| ANAER-E+Fe | - | - | 100 | 8 | ||
| Anoxic | ANOX-S (Sterile) | 2 (days 0–10) 3 (days 10–60) | 10 (days 0–10) 20 (days 10–60) | - | - | - |
| ANOX-B (Biotic control) | - | - | - | |||
| ANOX-M | 100 | - | - | |||
| ANOX-M+Fe | 100 | - | 8 | |||
| ANOX-E | - | 100 | - | |||
| ANOX-E+Fe | - | 100 | 8 |
* CODt: total COD.
Table 3.
Experimental conditions studied in the sulfate reducing microcosms tests.
| Experimental Set | Batch Test | Cr(VI) | SO42− | Molasses | EVO | Fe2+ |
|---|---|---|---|---|---|---|
| mg/L | mg/L | mg/L CODt * | mg/L CODt * | mg/L | ||
| Low sulfate concentration | S100-S (Sterile) | 2 (days 0–10) 3 (days 10–60) | 100 | - | - | - |
| S100-B (Biotic control) | - | - | - | |||
| S100-M | 100 | - | - | |||
| S100-M+Fe | 100 | - | 8 | |||
| S100-E | - | 100 | - | |||
| S100-E+Fe | - | 100 | 8 | |||
| High sulfate concentration | S400-S (Sterile) | 2 (days 0–10) 3 (days 10–60) | 400 | - | - | - |
| S400-B (Biotic control) | - | - | - | |||
| S400-M | 100 | - | - | |||
| S400-M+Fe | 100 | - | 8 | |||
| S400-E | - | 100 | - | |||
| S400-E+Fe | - | 100 | 8 |
* CODt: total COD.
Table 4.
Results of the parent cultures’ performance during the acclimatization period (Weekly average results).
Table 4.
Results of the parent cultures’ performance during the acclimatization period (Weekly average results).
| Parameter | Parent Culture | |||
|---|---|---|---|---|
| Anaerobic | Anoxic | Sulfate 100 mg/L | Sulfate 400 mg/L | |
| CODs * (mg/L) | 86.8 ± 20.1 | 48.0 ± 8.3 | 57.3 ± 18.1 | 49.3 ± 16.5 |
| VFAs (mg CODs/L) | 27.5 ± 8.3 | 13.1 ± 5.0 | 11.2 ± 2.0 | 9.9 ± 0.9 |
| pH | 8.28 ± 0.16 | 8.30 ± 0.13 | 8.23 ± 0.12 | 8.15 ± 0.22 |
| Cr(VI) (μg/L) | ND | ND | ND | ND |
| NO3-N (mg/L) | - | <0.23 | - | - |
| NO2-N (mg/L) | - | <0.01 | - | - |
| remaining SO42− (mg/L) | - | - | 58.6 ± 45.3 | 280.0 ± 58.0 |
| S2− (mg/L) | - | - | 3.1 ± 2.1 | 13.2 ± 7.1 |
* CODs: soluble COD.
Table 5.
Average sulfide concentrations under sulfate reducing conditions (S100 and S400 experiments).
Table 5.
Average sulfide concentrations under sulfate reducing conditions (S100 and S400 experiments).
| Experimental Set | Sulfide Concentration (mg/L) | ||
|---|---|---|---|
| 0–10 Days | 10–20 Days | 20–60 Days | |
| S100-B | 4.6 ± 0.0 | 3.1 ± 0.1 | 1.6 ± 1.1 |
| S100-M | 7.1 ± 0.2 | 4.3 ± 0.8 | 3.7 ± 0.8 |
| S100-E | 7.5 ± 0.5 | 3.1 ± 1.5 | 3.0 ± 1.1 |
| S100-M+Fe | 7.1 ± 0.6 | 5.4 ± 1.4 | 3.2 ± 1.6 |
| S100-E+Fe | 9.1 ± 1.2 | 5.8 ± 0.8 | 3.3 ± 2.2 |
| S400-B | 2.1 ± 0.1 | 1.7 ± 0.6 | 1.0 ± 0.5 |
| S400-M | 19.0 ± 1.8 | 6.9 ± 3.9 | 2.3 ± 1.0 |
| S400-E | 12.2 ± 2.8 | 7.0 ± 5.1 | 1.9 ± 1.0 |
| S400-M+Fe | 16.2 ± 5.3 | 3.6 ± 2.8 | 2.0 ± 0.7 |
| S400-E+Fe | 13.3 ± 2.6 | 5.3 ± 0.5 | 3.9 ± 2.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Galani, A.; Noutsopoulos, C.; Anastopoulou, P.; Varouxaki, A.; Mamais, D. Reductive Cr(VI) Removal under Different Reducing and Electron Donor Conditions—A Soil Microcosm Study. Water 2022, 14, 2179. https://doi.org/10.3390/w14142179
AMA Style
Galani A, Noutsopoulos C, Anastopoulou P, Varouxaki A, Mamais D. Reductive Cr(VI) Removal under Different Reducing and Electron Donor Conditions—A Soil Microcosm Study. Water. 2022; 14(14):2179. https://doi.org/10.3390/w14142179
Chicago/Turabian StyleGalani, Andriani, Constantinos Noutsopoulos, Petra Anastopoulou, Alexia Varouxaki, and Daniel Mamais. 2022. "Reductive Cr(VI) Removal under Different Reducing and Electron Donor Conditions—A Soil Microcosm Study" Water 14, no. 14: 2179. https://doi.org/10.3390/w14142179
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.