
The Inland Water Copepod Fauna of a Traditional Rural Landscape in a Mediterranean Island (Crustacea, Copepoda)


Department of Biological, Chemical and Pharmaceutical Sciences and Technologies (STEBICEF), University of Palermo, Via Archirafi 18, 90123 Palermo, Italy
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Water 2022, 14(14), 2168; https://doi.org/10.3390/w14142168
Submission received: 16 June 2022
/
Revised: 5 July 2022
/
Accepted: 6 July 2022
/
Published: 8 July 2022
(This article belongs to the Special Issue Zooplankton Diversity in Water Bodies)
Abstract
:Although the Mediterranean area is a well-known hotspot of biological diversity, the crustacean assemblages inhabiting inland waters of the Mediterranean islands are to date unevenly known, and detailed information is missing for most taxa and areas. In the frame of this paper, we provide a checklist and a characterization of the copepod fauna of the lentic water bodies occurring in a traditional rural landscape of Sicily, where the co-existence of agriculture, woodlands, and pastoral activities lead to the presence of a wide range of different aquatic habitats. Overall, 22 copepod species belonging to the orders Calanoida, Cyclopoida, and Harpacticoida have been found in the 92 surveyed sites, stressing the conservation value of the area. In the study area, species widespread in the west Palaearctic region co-exist with strictly Mediterranean elements and a small but biogeographically significant group of species with northern or Balkan affinities, which support the role of the investigated area as a refugium for species that colonised Sicily during Pleistocene climate fluctuations and are now restricted to the more wet parts of the island. A single non-native species has been found, and its distribution is currently limited to permanent, man-made reservoirs.
Keywords:
Calanoida; Cyclopoida; Harpacticoida; inland waters; Sicily; biological invasions; temporary ponds1. Introduction
Thanks to a complex combination of climatic, geological, and paleogeographic factors, the Mediterranean Basin, characterised by a pronounced seasonality and a mosaic of ecosystems and communities, is a well-known hotspot of biological diversity [1]. However, the biota inhabiting the inland waters of the Mediterranean area is unevenly known, with several taxa and areas poorly known or data-deficient. This is particularly unfortunate since inland water bodies host a high biological diversity due to the intrinsically fragmented nature of these habitats, which promotes endemism and differentiation phenomena [2]. The inland waterbodies of the Mediterranean area can thus be correctly considered “hotspots within hotspots”. Unfortunately, Mediterranean inland water biota is nowadays highly threatened by a combination of climatic change, biological invasions, and anthropic pressure, and several taxa are exposed to local or global extinction even before their presence is surveyed or they are formally described [3]. A sound assessment of the biodiversity of Mediterranean inland waters is thus urgent to describe and characterise this unique biota before it vanishes and to create a sound reference point which allows to explore the long-term biological changes that might interest the area.
Copepod crustaceans are amongst the most relevant organisms in inland waters in terms of diversity, numbers, and biomass and constitutes one of the major groups among inland water microcrustaceans, where they are represented by three orders of mostly free-living species: Calanoida, Cyclopoida, and Harpacticoida [4]. In Sicilian inland waters, calanoids have been sufficiently investigated, with special reference to the families Diaptomidae (e.g., [5,6,7,8,9,10]) and Pseudodiaptomidae [11]. Conversely, the cyclopoid and harpacticoid copepods inhabiting the surface inland waters of Sicily have been to date the object of few dedicated studies (e.g., [12,13,14,15]), and most of the available records are occasionally included in papers dealing with other topics (e.g., [5,16,17,18]).
In the frame of this work, we provide a checklist and distribution of the copepods occurring in an area characterised by a mixed agricultural, rangeland, and wood landscape, which can be considered representative of the land use, climate, and physiography of the hilly and mountainous inner part of Sicily, the largest island of the Mediterranean region and a renowned “crossroad” for circum-Mediterranean freshwater microcrustaceans ([19,20] and references therein). The crustacean fauna inhabiting the surface lentic inland water bodies of Sicily is in fact a prime example of an understudied but greatly interesting group, and no recent synoptical data are available for Sicilian cyclopoid and harpacticoid copepods. Moreover, the few available lists should be critically revised, and the realisation of sound checklists supported by accurate, possibly geo-referenced, distributional data is highly desirable.
2. Materials and Methods
2.1. The Study Area
The study area occurs within the province of Palermo, and it is included approximately between 37.82° and 37.92° latitude N and 13.30° and 13.45° longitude E, within an altitudinal range comprised between 350 and 1613 m a.s.l. Most of the investigated water bodies are located within the Nature Reserve “Bosco della Ficuzza, Rocca Busambra, Bosco del Cappelliere e Gorgo del Drago”, one of the largest protected areas of Sicily (Figure 1).
The landscape of the area is dominated by broadleaf woods, rangelands, agricultural fields, and rocky outcrops occurring on calcareous, siliceous, and clayey substrates. The average annual rainfall in the study area ranges between 700 and 1000 mm y−1 and the average annual temperature from 10 to 16 °C; the bioclimate ranges from the thermo-Mediterranean lower subhumid to the supra-Mediterranean humid belts [21,22]. A detailed characterization of the climate, flora, vegetation, and geology of the area is provided by Gianguzzi and La Mantia [22], and further information can be retrieved in Gianguzzi [23], whereas an extensive census of both lentic and lotic water bodies of the study area and their botanical characterization is provided by Caldarella [24]. Despite its long-term anthropic exploitation, the area still maintains a high level of plant diversity and a well-preserved rural landscape [25,26], hosting a peculiar and rare flora [27].
To date, some information is available for the ostracod [28,29], branchiopod [30], and malacostracan ([31,32] and references therein) faunas of the study area. Conversely, apart from the occasional records of two copepod species by Calvo et al. [15] and Marrone et al. [33], no information is currently available for Copepoda.
Sparse information is also available for freshwater molluscs (e.g., [34]), water mites [35], leeches [36], and insects (e.g., [37,38]). Among the aquatic vertebrates, some information is available for amphibians (See references in Lo Valvo et al. [39]), the Sicilian pond turtle Emys trinacris [40,41], and the freshwater blenny Salariopsis fluviatilis (Asso, 1801) (sub Salaria fluviatilis in [42,43]; see Vecchioni et al. [44]).
2.2. Sampling Methods
In the frame of this survey, only surface lentic water bodies ranging from a large dam reservoir to small pools on dirt roads were investigated (Figure 2). Both natural (pools and ponds) and man-made (reservoirs and concrete channel) water bodies with different hydroperiods were included in the sampling campaigns. Distribution maps were realised using the QGIS freeware software v. 3.18 (QGIS Development Team, 2022 [45]).
The majority of sites were sampled only once, whereas about 20% of the sampled sites were visited several times in different months and years in order to account for the seasonal successions and the possible inter-annual differences of the copepod assemblages (e.g., [46]). The coordinates of the sampled water bodies were registered with GPS along with information about their hydroperiods and the possible occurrence of vertebrate predators (i.e., fish or Xenopus laevis tadpoles). In each site and on each sampling date, electric conductivity and water temperature were recorded with a Hanna Instrument HI9835 multiprobe; moreover, an estimate of the abundance of macrophytes and of water turbidity was assessed by using three arbitrary qualitative classes (see Marrone et al. [47]).
Studied crustacean samples originate from two different sampling campaigns: from 2004 to 2019, samples were collected with a 200 μm hand net in littoral areas and with a 100 μm towing net in the open waters of larger water bodies. In the frame of a second sampling campaign (2020–2022), samples were collected with 250 μm hand and towing nets. In both cases, attention was paid to collect samples in all the microhabitats possibly present in each site. All used nets were manufactured and provided by NHBS Ltd. (Totnes, UK—https://www.nhbs.com/). Samples collected prior to 2010 were preserved in 4% buffered formalin, while those collected from 2010 to 2022 were preserved in 90% ethanol. All samples are now stored in Federico Marrone crustacean collection at the Department “STEBICEF” (Scienze e Tecnologie Biologiche Chimiche e Farmaceutiche—University of Palermo, Palermo, Italy) and are available for loan on request.
Identification of collected copepods was performed based on Dussart [48,49], Kiefer [50], Einsle [51,52], Karaytug [53], Hołyńska et al. [54], Hołyńska [55], Alekseev and Defaye [56], and Miracle et al. [57]. Copepod nomenclature here follows Ruffo and Stoch [58], with the exceptions of the genera Arctodiaptomus Kiefer, 1932, and Acanthocyclops Kiefer, 1927, for which we follow Alfonso et al. [59] and Miracle et al. [57], respectively. Notwithstanding the doubts raised by some authors (e.g., [60,61]), we here consider D. bicuspidatus (Claus, 1857) and D. lubbocki (Brady, 1869) as different taxa of species level.
Sample-based rarefaction curves [62] and the rarefaction curve of the mean values of “uniques” (i.e., species present only in a single sample) were computed to evaluate if sampling effort was exhaustive enough to be representative of the total copepod diversity of the study area. The non-parametric species richness estimators ICE (Incidence Coverage-based Estimator), Jack1 (first order Jackknife-based estimator), and the bias-corrected formula of Chao2 were calculated. Analyses were done using the EstimateS software version 9.1.0 (Boulder, USA - http://viceroy.eeb.uconn.edu/EstimateS - accessed on 1 May 2022), based on the analytical formulas of Colwell et al. [63].
3. Results
Overall, 122 samples were collected in 92 lentic water bodies characterised by different origin (68 natural and 24 man-made sites) and hydroperiod (66 temporary, 6 astatic, and 20 permanent sites), located from 470 to 1165 m a.s.l. (Table 1). A synopsis of the environmental data and collected copepods for each site and sampling date is reported in Appendix A, Table A1.
Copepods were observed in 88 out of the 92 sampled sites. All the copepod taxa could be identified to species level with the exceptions of the diaptomid Arctodiaptomus sp., a species new to science whose formal description is still pending (see Alfonso et al. [59]), and a cyclopoid species belonging to the genus Megacyclops Kiefer, 1927. The Megacyclops populations studied in the frame of this survey showed a morphology close but not identical to M. viridis (Jurine, 1820). Pending a revision of the genus Megacyclops, we have thus conservatively opted for identifying the Megacyclops populations of the study area as Megacyclops viridis s.l.
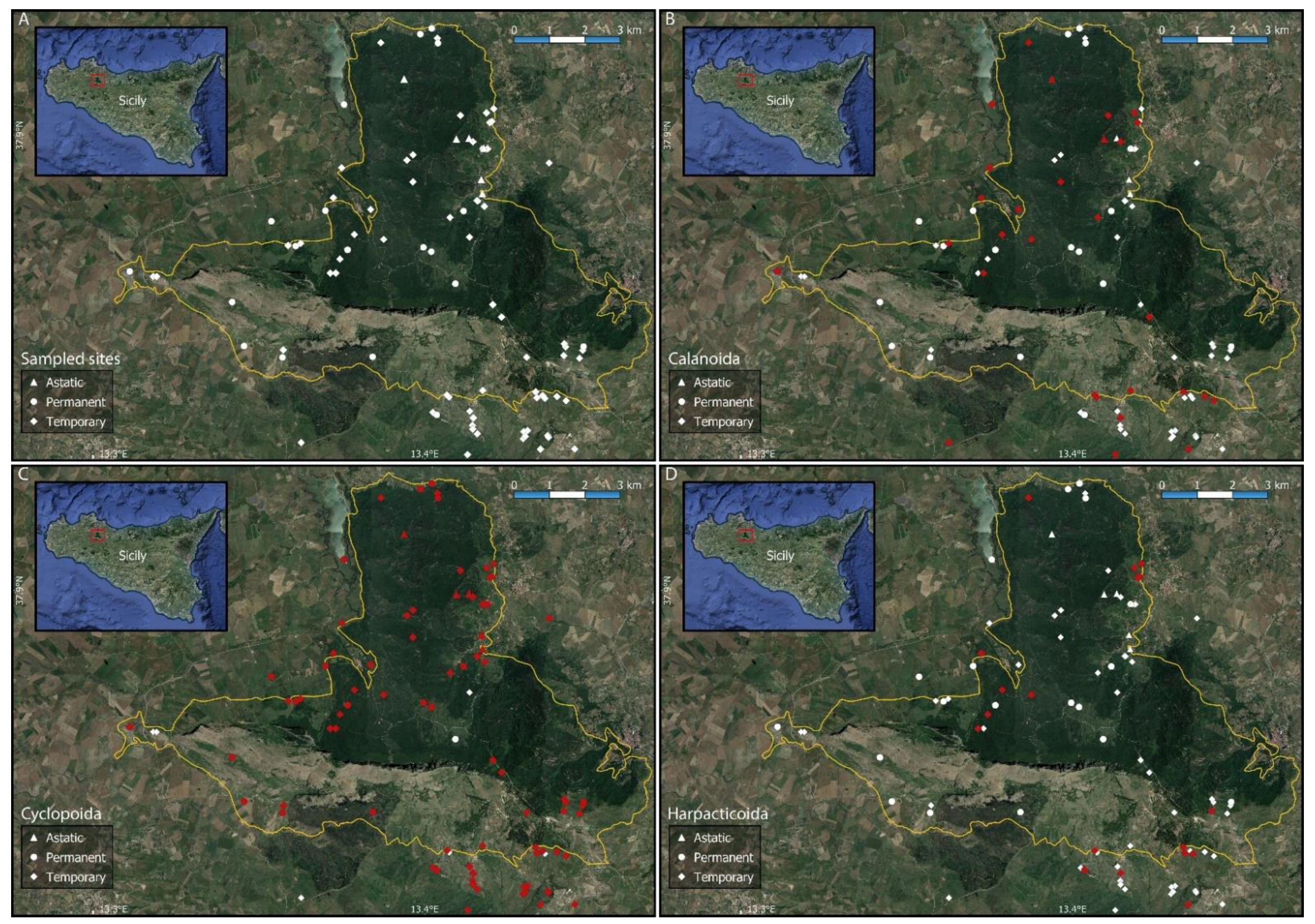
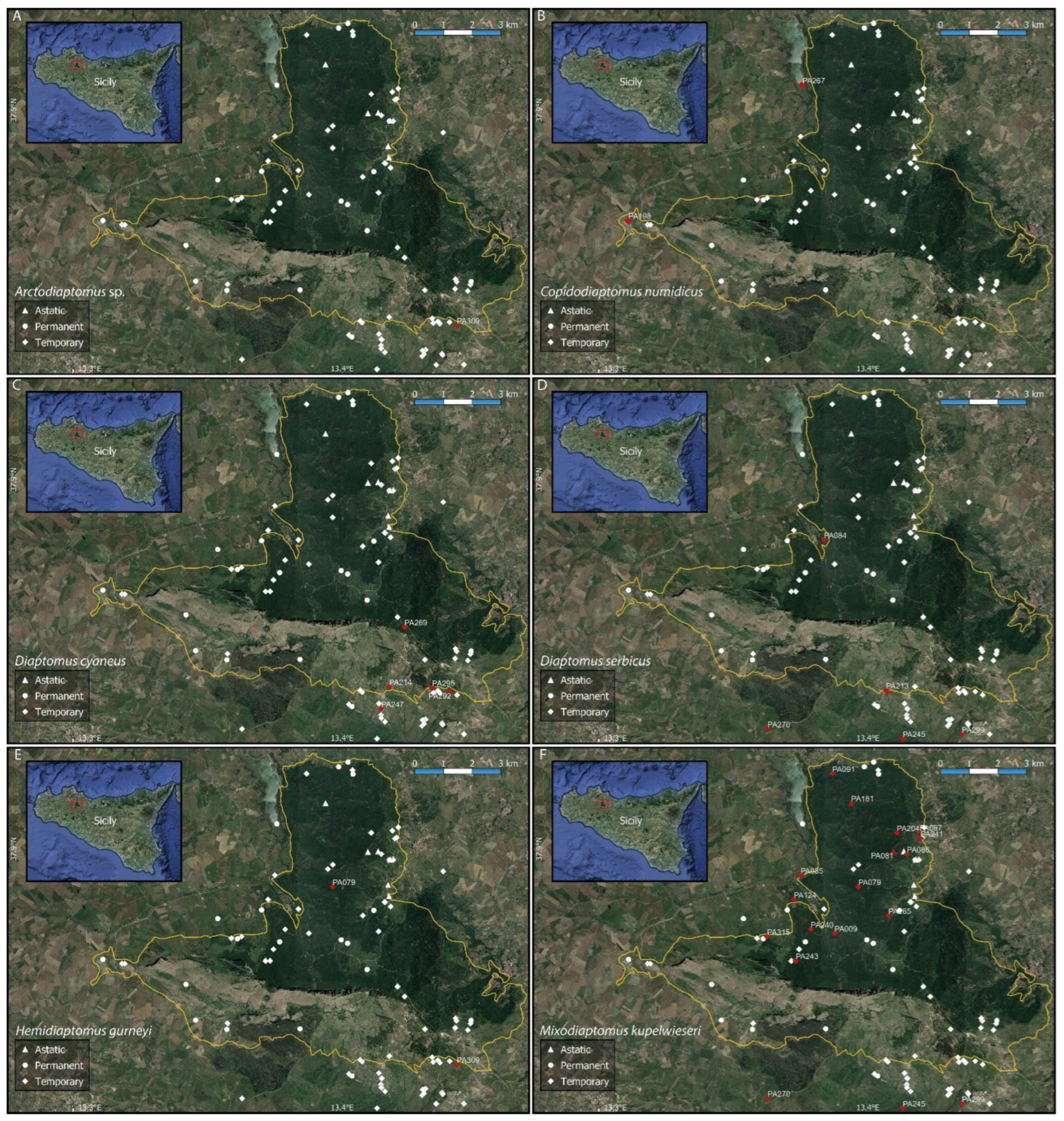
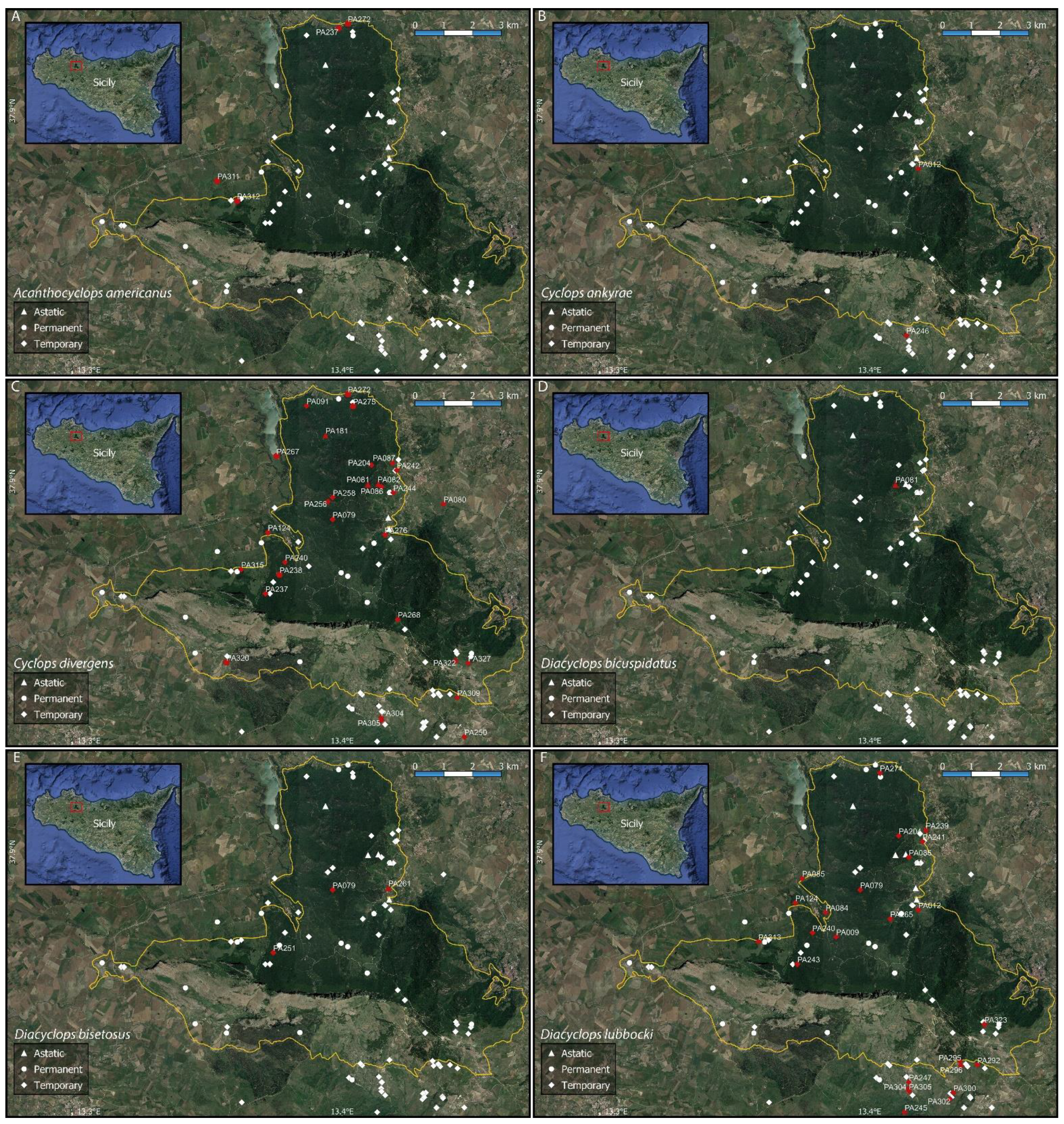
The obtained checklist includes 22 species belonging to 18 genera. Calanoid, cyclopoid, and harpacticoid copepods were observed in 29, 85, and 18 of the sampled water bodies, respectively (Figure 1). The occurrence sites for each of the recorded species are listed in Table 2 and shown in Appendix B, Figure A1, Figure A2, Figure A3 and Figure A4; synoptical data on the number of occurrence localities (N) and the observed altitudinal, electric conductivity (EC), and hydroperiod ranges of each species are reported in Table 3.
The most frequently occurring copepod species within the study area (i.e., those occurring in at least 20% of the sampled sites) were the cyclopoids Cyclops divergens Lindberg, 1936; Megacyclops viridis s.l.; Diacyclops lubbocki (Brady, 1869); Eucyclops serrulatus (Fischer, 1851); and the calanoid Mixodiaptomus kupelwieseri (Brehm, 1907) (Table 2).
The occurrence in the study area of both the copepod species reported in the literature, i.e., the diaptomid Hemidiaptomus gurneyi (Roy, 1927) and the cyclopid Diacyclops bicuspidatus (Claus, 1857), was confirmed during the present survey.
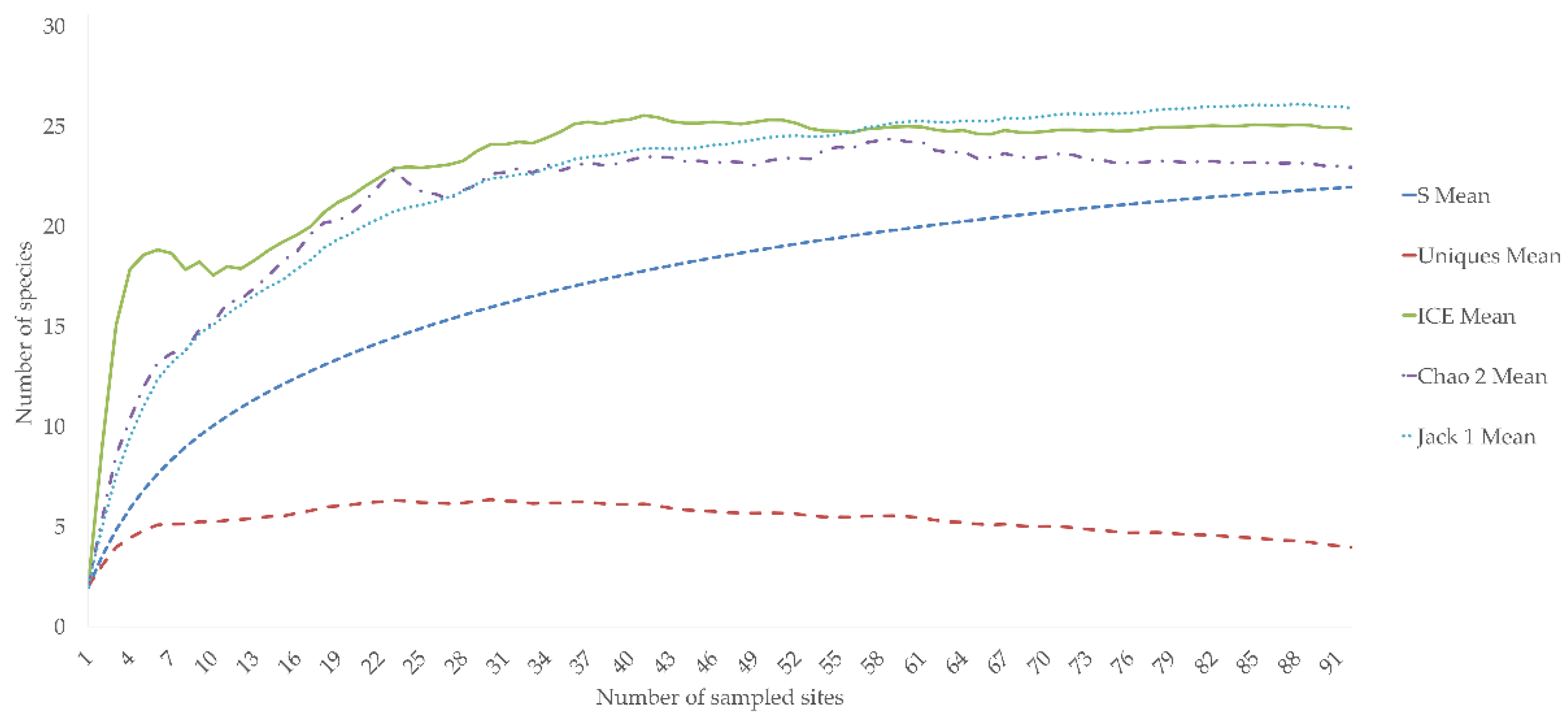
Rarefaction curves and species richness estimators are reported in Figure 3. The sample-based rarefaction curve of mean species richness based on the collected data approaches the asymptote without reaching it, thus suggesting that the recorded species richness is a bit lower than the real copepod diversity occurring in the study area. The trend of uniqueness is stable at four species. The implemented non-parametric species richness estimators show an expected overall species richness between 23 (bias-corrected formula of Chao2 equation—Chao2) and 26 (first-order Jackknife richness estimator—Jack1) in the study area. These values are slightly higher than the recorded copepod species richness value of 22.
4. Discussion
The current crustacean fauna of Sicily is characterised by the sympatric (but mostly allotopic) coexistence of mesophilous taxa of northern and north-eastern origin, which likely colonised the island during Pleistocene climatic fluctuations and are now confined to higher altitudes and microthermal refuges, and termophilous taxa of southern origin, which colonised Sicily during the Holocene and whose expansion in the island is still underway [20]. Due to its physiography, geology, and climate, the study area is expected to be characterised by the occurrence of taxa with a northern or north-eastern affinity, as also suggested by its flora and vegetation [24].
Most of the collected harpacticoid and cyclopoid copepods are here ascribed to euryecious species, which are rather widespread in the west Palaearctic region and thus biogeographically poorly informative. However, the unresolved taxonomy of most copepod taxa belonging to these orders leads to overestimate their distribution ranges and ecological niches, thus biasing any ecological and biogeographical analysis [64], and it is likely that under several binomia are actually lumped complexes of species characterised by narrower distribution areas and ecology (e.g., [65,66]). The only cyclopoid species with a restricted distribution range collected in the frame of the present survey is Cyclops ankyrae Mann, 1940, belonging to a Turanic-Mediterranean chorotype sensu Vigna Taglianti et al. [67]. Conversely, the relatively stable taxonomy of the Calanoida and the fairly accurate degree of knowledge about their large-scale distribution allow to draft some biogeographical inferences. Within this group, the occurrence of species with European (Mixodiaptomus kupelwieseri) or Balkan (Hemidiaptomus gurneyi, Diaptomus serbicus Gjorgjewič, 1907, and Arctodiaptomus sp.) distribution barycentres confirms the role of the area as a refugium where relic copepod taxa that colonised the island during the Pleistocene glaciations might have survived during the current post-glacial phase, as also observed for other aquatic taxa in the same area (e.g., [35,68]). This group of mesophilous species occurs in association with some primarily west Mediterranean species linked with Mediterranean “rainy areas” (sensu Gauthier [69], see also Marrone et al. [70]), here represented by the diaptomids Copidodiaptomus numidicus (Gurney, 1909) and Diaptomus cyaneus Gurney, 1909.
In accordance with the expectations, no strictly Maghrebian elements are present in the study area since these are confined to the arid and semi-arid parts of the island [5,6].
Based on the currently available data, the copepod fauna of the study area is thus composed by a numerically small but biogeographically significant group of taxa with a Balkan affinity, a few Mediterranean taxa, and a larger group of species with wider distribution areas.
The absence of consolidated checklists for Sicilian cyclopoids and harpacticoids prevents from a sound comparison between the regional fauna and that of the study area. However, most of the species reported for Sicilian surface waters by Ruffo and Stoch [58] were collected also in the frame of this study, whereas the identity of some taxa listed by Ruffo and Stoch [58] but not found in the study area needs to be thoroughly verified; e.g., the populations of Cyclops abyssorum G.O. Sars, 1863, reported by Calvo et al. [15] are possibly to be ascribed to C. divergens, a species occurring in the study area.
With reference to the order Calanoida, for which more representative data are available (cf. Alfonso et al. [59] for the Diaptomidae and Vecchioni et al. [11] for the Pseudodiaptomidae), only 6 of the 11 species reported for Sicily were found in the frame of this study. Only those species restricted to highly mineralised water bodies, such as Arctodiaptomus salinus (Daday, 1885) and Calanipeda aquaedulcis Kritschagin, 1873, and those linked with arid and semi-arid areas, such as Hemidiaptomus ingens (Gurney, 1909) and Metadiaptomus chevreuxi (Guerne and Richard, 1894), along with and Arctodiaptomus kerkyrensis (Pesta, 1935), a rare species currently known for a single site in eastern Sicily, proved to be absent.
Based on the available data, the study area is thus characterised by a noteworthy copepod diversity and high conservation value. Moreover, as suggested by the sample-based rarefaction curves and the non-parametric species richness estimators, further sampling surveys might bring to the finding of further copepod species, with an estimated occurrence of one to four species more than those currently known.
All the collected copepod species are already known to occur in Sicily (see references above), but most of them are regionally rare. In fact, except for E. serrulatus, T. prasinus (Fischer, 1866), M. viridis, C. divergens, and D. lubbocki, all the other observed copepod species were to date known in Sicily from few sites each, and their status and distribution on the island are currently unknown. Among the Cyclopoida, the finding of Cyclops ankyrae, a rare cyclopid species currently known to occur in temporary water bodies in Sicily [17], peninsular Italy [58,71,72], the southern Balkan peninsula [47], Turkey [55], and Iran [52] is particularly noteworthy; however, it is probable that the populations of C. furcifer Claus, 1857, a mostly North European species reported for Sicilian temporary ponds by Margaritora et al. [12], are in fact to be ascribed to C. ankyrae so that the regional distribution of the species could be underestimated. As already stressed for Cyclops abyssorum, the copepod populations belonging to the genus Cyclops in Sicily need a careful revision to ascertain their actual identity.
All the calanoid copepods observed in the study area were collected in astatic or temporary water bodies, with the only exception of the diaptomid Copidodiaptomus numidicus, which was collected only in permanent reservoirs. Among the Cyclopoida, six species were only observed in temporary or astatic ponds, three were only observed in permanent ones, and five species were collected both in permanent and temporary water bodies (Table 3). Cyclops ankyrae and Metacyclops minutus were usually observed in small, temporary water bodies. Conversely, Tropocyclops prasinus, Acanthocyclops americanus (Marsh, 1893), and Thermocyclops dybowskii (Landé, 1890) were only observed in permanent water bodies. However, the limited number of occurrence localities available for several cyclopoid species prevents from any sound ecological inference for the Sicilian populations of these taxa, and it is desirable that further distribution and ecological data are collected. Acanthocyclops americanus is the only non-native copepod species found in the study area. It is a widespread invader of Nearctic origin, which colonised most of the Palaearctic region from Madeira to Central Siberia, where it became one of the dominant zooplankters in eutrophic reservoirs [73,74]. In the study area, A. americanus was found in four permanent reservoirs, where it was often the only occurring copepod. In Sicily, the species was already reported for a concrete reservoir within the city of Palermo (Schifani et al. [18], sub A. trajani Mirabdullayev and Defaye, 2004), thus confirming the preference of this non-native species for permanent, man-made water bodies; in fact, artificial reservoirs are known to act as invasion hubs favouring invasive species because of their recent origin and the lack of structured biological communities [75,76]. A. americanus is currently limited to the western and northern fringes of the study area, and its possible colonisation of the natural water bodies of the area should be monitored. However, it should also be stressed that, although the synonymy of A. trajani and A. americanus and their non-native status in the Palaearctic region are accepted by most authors (e.g., [57,73,74]), these issues have been discussed and criticized by Anufriieva et al. [77], and further research should desirably be performed to test these hypotheses.
Only two harpacticoids have been found in the frame of this survey, i.e., Attheyella trispinosa (Brady, 1880) and Canthocamptus staphylinus (Jurine, 1820). Both species were collected mostly in temporary water bodies although A. trispinosa, was also found in a single permanent site out of the nine occurrence localities of the species. Prior to this survey, both species had been only occasionally recorded in Sicily [17,78]; however, they proved to be rather common in the study area, thus suggesting that their alleged rarity on the island might be ascribed to the lack or paucity of samplings aimed at collecting harpacticoids in Sicily rather than to their actual rarity.
5. Conclusions
The results of present work stress the noteworthy diversity of the copepod fauna of the investigated traditional rural landscape and the magnitude of the so-called Wallacean and Hutchinsonian shortfalls (see Hortal et al. [79]), which are hindering current knowledge and comprehension of the inland water biota of the Mediterranean islands. Apart for a few studies [19,29,30,80,81,82,83,84,85], the non-malacostracan inland water crustacean fauna of Mediterranean islands is to date scarcely known, and synoptical data are missing. We hope that this paper might stimulate further, detailed faunistic studies, which are the necessary bases for any further ecological, taxonomical, and biogeographical inference, and help to better understand and manage the unique inland water biological diversity of the Mediterranean area.
Author Contributions
Conceptualization, F.M. and M.A.; methodology, M.M., L.V., G.B. and F.M.; writing—original draft preparation, M.M., L.V., G.B. and F.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partly supported by the Research Project MEMOLA—Mediterranean Mountainous Landscapes: a historical approach to cultural heritage based on traditional agrosystems, funding from the European Union’s Seventh Framework Programme for Research, Technological Development, and Demonstration under grant agreement no. 613265.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
NHBS Ltd. (www.nhbs.com) is kindly acknowledged for having provided the nets used in the frame of this survey. The authors wish to thank Fabio Stoch (Université libre de Bruxelles, Belgium) for the stimulating discussions about inland water copepods and Federica Vicari, Luca Montevago, and Francesco Barna for the support they provided in the frame of the fieldwork. Orazio Caldarella is warmly acknowledged for the information he generously shared about the location and characteristics of the water bodies occurring in the study area. The comments of three anonymous reviewers allowed to improve the first draft of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Collected copepods and ecological characteristics of the sampled sites at different sampling dates. EC, electrical conductivity (µS/cm); TEMP, water temperature (°C); n.a., not available. FM, Federico Marrone; FV, Federica Vicari; LM, Luca Montevago; LV, Luca Vecchioni; MM, Marta Marchese.
Table A1.
Collected copepods and ecological characteristics of the sampled sites at different sampling dates. EC, electrical conductivity (µS/cm); TEMP, water temperature (°C); n.a., not available. FM, Federico Marrone; FV, Federica Vicari; LM, Luca Montevago; LV, Luca Vecchioni; MM, Marta Marchese.
| Code | Sampling Date | Calanoida | Cyclopoida | Harpacticoida | EC | TEMP | Collector(s) |
|---|---|---|---|---|---|---|---|
| PA009 | 09/03/2004 | M. kupelwieseri | M. viridis s.l., D. lubbocki | A. trispinosa | 112 | 10 | FM |
| PA009 | 08/05/2015 | M. kupelwieseri | M. viridis s.l. | A. trispinosa | 140 | 17 | FM |
| PA009 | 30/04/2019 | M. kupelwieseri | M. viridis s.l. | A. trispinosa | n.a. | n.a. | FV and LM |
| PA009 | 24/04/2021 | M. kupelwieseri | M. viridis s.l. | A. trispinosa | 180 | 16 | MM |
| PA012 | 20/12/2003 | C. ankyrae, D. lubbocki | 250 | 10 | FM | ||
| PA079 | 10/03/2004 | M. kupelwieseri | D. lubbocki | 53 | 6 | FM | |
| PA079 | 02/05/2004 | M. kupelwieseri | D. lubbocki | 300 | 18 | FM | |
| PA079 | 28/01/2012 | juveniles | D. lubbocki | 309 | 4.8 | FM | |
| PA079 | 06/03/2012 | H. gurneyi, M. kupelwieseri | D. lubbocki | 208 | 13.4 | FM | |
| PA079 | 03/01/2021 | C. divergens, D. bisetosus | 71 | 8.0 | MM | ||
| PA080 | 20/12/2003 | M. viridis s.l., C. divergens, M. minutus | 340 | 9 | FM | ||
| PA081 | 02/05/2004 | M. kupelwieseri | C. divergens, D. bicuspidatus | 109 | 18.8 | FM | |
| PA081 | 27/05/2010 | M. kupelwieseri | M. viridis s.l., E. serrulatus | 145 | 16 | FM | |
| PA081 | 26/04/2019 | M. kupelwieseri | M. viridis s.l. | n.a. | n.a. | FV and LM | |
| PA081 | 03/01/2021 | M. kupelwieseri | M. viridis s.l., E. serrulatus | 94 | 6.0 | MM | |
| PA082 | 06/05/2018 | M. viridis s.l., C. divergens, E. serrulatus | n.a. | n.a. | FM | ||
| PA082 | 26/04/2019 | C. divergens, E. serrulatus | n.a. | n.a. | FV and LM | ||
| PA082 | 03/01/2021 | 1372 | 7.5 | MM | |||
| PA082 | 06/05/2010 | 678 | 18 | FM | |||
| PA082 | 08/05/2015 | C. divergens | 340 | 21 | FM | ||
| PA084 | 19/04/2021 | D. serbicus | Diacyclops lubbocki | 467 | 13 | MM, FM, LV | |
| PA085 | 22/02/2005 | M. kupelwieseri | D. lubbocki | 565 | 9.2 | FM | |
| PA085 | 13/02/2021 | M. kupelwieseri | E. serrulatus | 543 | 12 | MM | |
| PA086 | 26/04/2019 | M. kupelwieseri | D. lubbocki, C. divergens | n.a. | n.a. | FV and LM | |
| PA086 | 03/01/2021 | 160 | 8 | MM | |||
| PA087 | 20/12/2003 | M. kupelwieseri | C. divergens, M. minutus | C. staphylinus | 170 | 9 | FM |
| PA091 | 01/11/2010 | M. kupelwieseri | M. viridis s.l., C. divergens | A. trispinosa | n.a. | n.a. | FM |
| PA120 | 10/10/2020 | E. serrulatus | n.a. | n.a. | FM | ||
| PA121 | 21/08/2008 | E. serrulatus | 273 | 23.5 | FM | ||
| PA124 | 17/03/2022 | M. kupelwieseri | C. divergens, D. lubbocki | A. trispinosa | 158 | 11.3 | FM and LV |
| PA133 | 29/11/2020 | 230 | 13.0 | MM | |||
| PA181 | 01/11/2010 | M. kupelwieseri | C. divergens | 1124 | n.a. | FM | |
| PA198 | 09/01/2013 | E. serrulatus | 714 | 10 | FM | ||
| PA198 | 19/04/2021 | C. numidicus | T. dybowskii | 482 | 15 | MM, FM, LV | |
| PA204 | 17/03/2022 | M. kupelwieseri | M. viridis s.l., C. divergens, D. lubbocki | 222 | 6.5 | FM and LV | |
| PA212 | 29/01/2021 | D. serbicus | 1305 | 11.2 | MM | ||
| PA213 | 29/01/2021 | D. serbicus | M. minutus | A. trispinosa | 1028 | 12.5 | MM |
| PA214 | 07/03/2021 | D. cyaneus | M. viridis s.l. | 314 | 15.4 | MM | |
| PA235 | 26/04/2019 | M. viridis s.l. | n.a. | n.a. | FV and LM | ||
| PA236 | 26/04/2019 | M. albidus, E. serrulatus | n.a. | n.a. | FV and LM | ||
| PA237 | 30/04/2018 | C. divergens | A. trispinosa | n.a. | n.a. | FV and LM | |
| PA238 | 30/04/2019 | C. divergens | n.a. | n.a. | FV and LM | ||
| PA238 | 24/04/2021 | C. divergens | 159 | 18.5 | MM | ||
| PA239 | n.a. | D. lubbocki | A. trispinosa | n.a. | n.a. | FV and LM | |
| PA240 | 30/04/2019 | M. kupelwieseri | M. viridis s.l., D. lubbocki | C. staphylinus | n.a. | n.a. | FV and LM |
| PA240 | 03/01/2021 | M. kupelwieseri | C. divergens, D. lubbocki | 121 | 9.2 | MM | |
| PA241 | 26/04/2019 | M. kupelwieseri | M. viridis s.l. | C. staphylinus | n.a. | n.a. | FV and LM |
| PA241 | 13/02/2021 | M. kupelwieseri | D. lubbocki | C. staphylinus | 142 | 8.7 | MM |
| PA242 | 13/02/2021 | C. divergens, M. viridis s.l. | 290 | 9.5 | MM | ||
| PA242 | n.a. | E. serrulatus, M. viridis s.l. | C. staphylinus | n.a. | n.a. | FV and LM | |
| PA243 | 30/04/2019 | M. kupelwieseri | D. lubbocki | n.a. | n.a. | FV and LM | |
| PA244 | 26/04/2019 | C. divergens | n.a. | n.a. | FV and LM | ||
| PA245 | 29/03/2020 | D. serbicus, M. kupelwieseri | D. lubbocki | 470 | 12 | MM | |
| PA246 | 07/03/2021 | Cyclops ankyrae | 1335 | 14.5 | MM | ||
| PA247 | 29/03/2020 | D. cyaneus | 1369 | 14 | MM | ||
| PA247 | 29/01/2021 | D. cyaneus | M. viridis s.l., D. lubbocki | C. staphylinus | 992 | 10.8 | MM |
| PA247 | 12/02/2021 | D. cyaneus | D. lubbocki | 960 | 9.9 | MM | |
| PA247 | 07/03/2021 | D. cyaneus | M. viridis s.l. | 1287 | 15.7 | MM | |
| PA247 | 05/04/2021 | D. cyaneus | A. trispinosa | 1405 | 14.5 | MM | |
| PA248 | 12/02/2021 | 734 | 9.8 | MM | |||
| PA248 | 05/04/2021 | M. viridis s.l., M. minutus | 935 | 14.5 | MM | ||
| PA249 | 22/11/2020 | M. viridis s.l., E. serrulatus | 360 | 13 | MM | ||
| PA250 | 12/02/2021 | C. divergens | 541 | 15 | MM | ||
| PA251 | 24/04/2021 | D. bisetosus | C. staphylinus | 160 | 13 | MM | |
| PA252 | 13/02/2021 | M. minutus | 560 | 5.7 | MM | ||
| PA253 | 13/02/2021 | 64 | 6.2 | MM | |||
| PA253 | 19/04/2021 | 246 | 16.2 | MM, FM, LV | |||
| PA256 | 19/04/2021 | C. divergens | 4130 | 10.2 | MM, FM, LV | ||
| PA258 | 03/01/2021 | C. divergens | 112 | 8.0 | MM | ||
| PA259 | 29/01/2021 | M. viridis s.l., T. prasinus | A. trispinosa | 491 | 11.2 | MM | |
| PA261 | 13/02/2021 | M. viridis s.l., D. bisetosus | 585 | 13.6 | MM | ||
| PA264 | 19/04/2021 | M. albidus, M. leuckarti | 317 | 12.6 | MM, FM, LV | ||
| PA265 | 19/04/2021 | M. kupelwieseri | D. lubbocki | 220 | 15 | MM, FM, LV | |
| PA266 | 19/04/2021 | 413 | 12.1 | MM, FM, LV | |||
| PA267 | 19/04/2021 | C. numidicus | C. divergens | 419 | 12.1 | MM, FM, LV | |
| PA268 | 19/04/2021 | C. divergens | 123 | 13 | MM, FM, LV | ||
| PA269 | 19/04/2021 | D. cyaneus | M. viridis s.l. | 219 | 13.2 | MM, FM, LV | |
| PA270 | 07/03/2020 | D. serbicus, M. kupelwieseri | 360 | 10 | MM | ||
| PA271 | 13/02/2021 | E. serrulatus | 535 | 13 | MM | ||
| PA272 | 24/04/2021 | C. divergens, A. americanus | 626 | 18.3 | MM | ||
| PA273 | 24/04/2021 | A. americanus | 486 | 17.5 | MM | ||
| PA274 | 24/04/2021 | D. lubbocki | 89 | 16 | MM | ||
| PA275 | 24/04/2021 | C. divergens | 301 | 16.2 | MM | ||
| PA276 | 13/02/2021 | C. divergens | 83 | 10.5 | MM | ||
| PA277 | 13/02/2021 | C. divergens | 64 | 10 | MM | ||
| PA278 | 29/11/2020 | 190 | 13.0 | MM | |||
| PA292 | 05/12/2021 | juveniles | - | 43 | 11.4 | MM | |
| PA292 | 05/01/2022 | D. cyaneus | D. lubbocki | 41 | 14 | MM | |
| PA293 | 05/12/2021 | C. staphylinus | 41 | 12 | MM | ||
| PA294 | 05/12/2021 | E. serrulatus | n.a | n.a. | MM | ||
| PA295 | 05/12/2021 | juveniles | M. viridis s.l., D. lubbocki | A. trispinosa | 51 | 9.4 | MM |
| PA295 | 05/01/2022 | D. cyaneus | D. lubbocki | A. trispinosa | 40 | 12.5 | MM |
| PA296 | 05/12/2021 | M. viridis s.l., D. lubbocki | C. staphylinus | 57 | 9 | MM | |
| PA297 | 19/12/2021 | M. albidus, E. serrulatus | 27 | 8 | MM | ||
| PA298 | 19/12/2021 | E. serrulatus | 21 | 8.7 | MM | ||
| PA299 | 19/12/2021 | juveniles | M. varicans | A. trispinosa | 90 | 9 | MM |
| PA299 | 05/01/2022 | D. serbicus, M. kupelwieseri | 50 | 12.3 | MM | ||
| PA300 | 19/12/2021 | D. lubbocki | 40 | 9.2 | MM | ||
| PA301 | 19/12/2021 | M. viridis s.l., E. serrulatus | 94 | 9.3 | MM | ||
| PA302 | 19/12/2021 | D. lubbocki | 98 | 9.6 | MM | ||
| PA303 | 19/12/2021 | M. viridis s.l., M. leuckarti | 83 | 9.6 | MM | ||
| PA304 | 19/12/2021 | C. divergens, D. lubbocki | 97 | 9.5 | MM | ||
| PA305 | 19/12/2021 | C. divergens, D. lubbocki | 123 | 8.8 | MM | ||
| PA306 | 19/12/2021 | M. viridis s.l. | 120 | 8.8 | MM | ||
| PA308 | 19/12/2021 | M. viridis s.l. | 98 | 8.2 | MM | ||
| PA309 | 05/01/2022 | H. gurneyi, D. cyaneus, Arctodiaptomus sp. | M. viridis s.l., C. divergens | 39 | 13 | MM | |
| PA311 | 17/03/2022 | A. americanus | 505 | 13.5 | FM and LV | ||
| PA312 | 17/03/2022 | A. americanus | 478 | 10 | FM and LV | ||
| PA313 | 17/03/2022 | D. lubbocki | 152 | 12.2 | FM and LV | ||
| PA314 | 17/03/2022 | T. prasinus | 296 | 11.2 | FM and LV | ||
| PA315 | 17/03/2022 | M. kupelwieseri | M. viridis s.l., C. divergens | 1100 | 10.2 | FM and LV | |
| PA317 | 02/05/2021 | E. serrulatus | 438 | 20 | MM | ||
| PA318 | 19/03/2022 | E. serrulatus | 56 | 5 | MM | ||
| PA319 | 19/03/2022 | M. viridis s.l., E. serrulatus | 64 | 6 | MM | ||
| PA320 | 19/03/2022 | C. divergens | 54 | 6 | MM | ||
| PA321 | 19/03/2022 | M. albidus | 66 | 5.7 | MM | ||
| PA322 | 20/03/2022 | C. divergens | C. staphylinus | 20 | 7 | MM | |
| PA323 | 20/03/2022 | D. lubbocki | 66 | 6.8 | MM | ||
| PA324 | 20/03/2022 | E. serrulatus | 45 | 7.2 | MM | ||
| PA325 | 20/03/2022 | E. serrulatus | 73 | 7.5 | MM | ||
| PA326 | 20/03/2022 | M. viridis s.l. | 45 | 7.3 | MM | ||
| PA327 | 20/03/2022 | M. viridis s.l., C. divergens | 60 | 7.2 | MM |
Appendix B
Figure A1.
Distribution maps of the six collected calanoid copepod species ((A) Arctodiaptomus sp.; (B) Copidodiaptomus numidicus; (C) Diaptomus cyaneus; (D) Diaptomus serbicus; (E) Hemidiaptomus gurneyi; (F) Mixodiaptomus kupelwieseri). The solid yellow line shows the boundaries of the Nature Reserve “Bosco della Ficuzza, Rocca Busambra, Bosco del Cappelliere e Gorgo del Drago”.
Figure A1.
Distribution maps of the six collected calanoid copepod species ((A) Arctodiaptomus sp.; (B) Copidodiaptomus numidicus; (C) Diaptomus cyaneus; (D) Diaptomus serbicus; (E) Hemidiaptomus gurneyi; (F) Mixodiaptomus kupelwieseri). The solid yellow line shows the boundaries of the Nature Reserve “Bosco della Ficuzza, Rocca Busambra, Bosco del Cappelliere e Gorgo del Drago”.

Figure A2.
Distribution maps of some of the collected cyclopoid copepod species ((A) Acanthocyclops americanus; (B) Cyclops ankyrae; (C) Cyclops divergens; (D) Diacyclops bicuspidatus; (E) Diacyclops bisetosus; (F) Diacyclops lubbocki). The solid yellow line shows the boundaries of the Nature Reserve “Bosco della Ficuzza, Rocca Busambra, Bosco del Cappelliere e Gorgo del Drago”.
Figure A2.
Distribution maps of some of the collected cyclopoid copepod species ((A) Acanthocyclops americanus; (B) Cyclops ankyrae; (C) Cyclops divergens; (D) Diacyclops bicuspidatus; (E) Diacyclops bisetosus; (F) Diacyclops lubbocki). The solid yellow line shows the boundaries of the Nature Reserve “Bosco della Ficuzza, Rocca Busambra, Bosco del Cappelliere e Gorgo del Drago”.

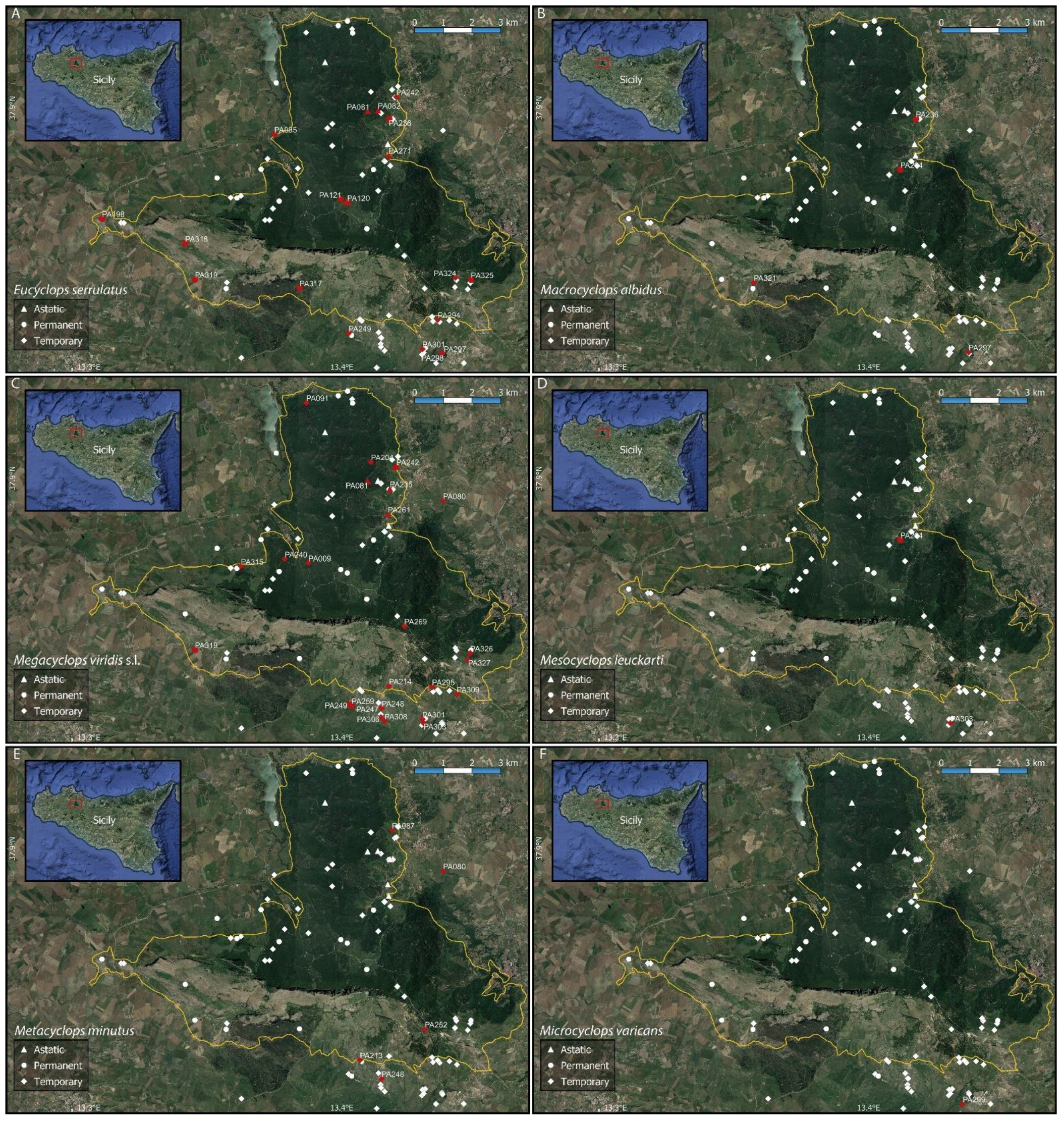
Figure A3.
Distribution maps of some of the collected cyclopoid copepod species ((A) Eucyclops serrulatus; (B) Macrocyclops albidus; (C) Megacyclops viridis s.l.; (D) Mesocyclops leuckarti; (E) Metacyclops minutus; (F) Microcyclops varicans). The solid yellow line shows the boundaries of the Nature Reserve “Bosco della Ficuzza, Rocca Busambra, Bosco del Cappelliere e Gorgo del Drago”.
Figure A3.
Distribution maps of some of the collected cyclopoid copepod species ((A) Eucyclops serrulatus; (B) Macrocyclops albidus; (C) Megacyclops viridis s.l.; (D) Mesocyclops leuckarti; (E) Metacyclops minutus; (F) Microcyclops varicans). The solid yellow line shows the boundaries of the Nature Reserve “Bosco della Ficuzza, Rocca Busambra, Bosco del Cappelliere e Gorgo del Drago”.

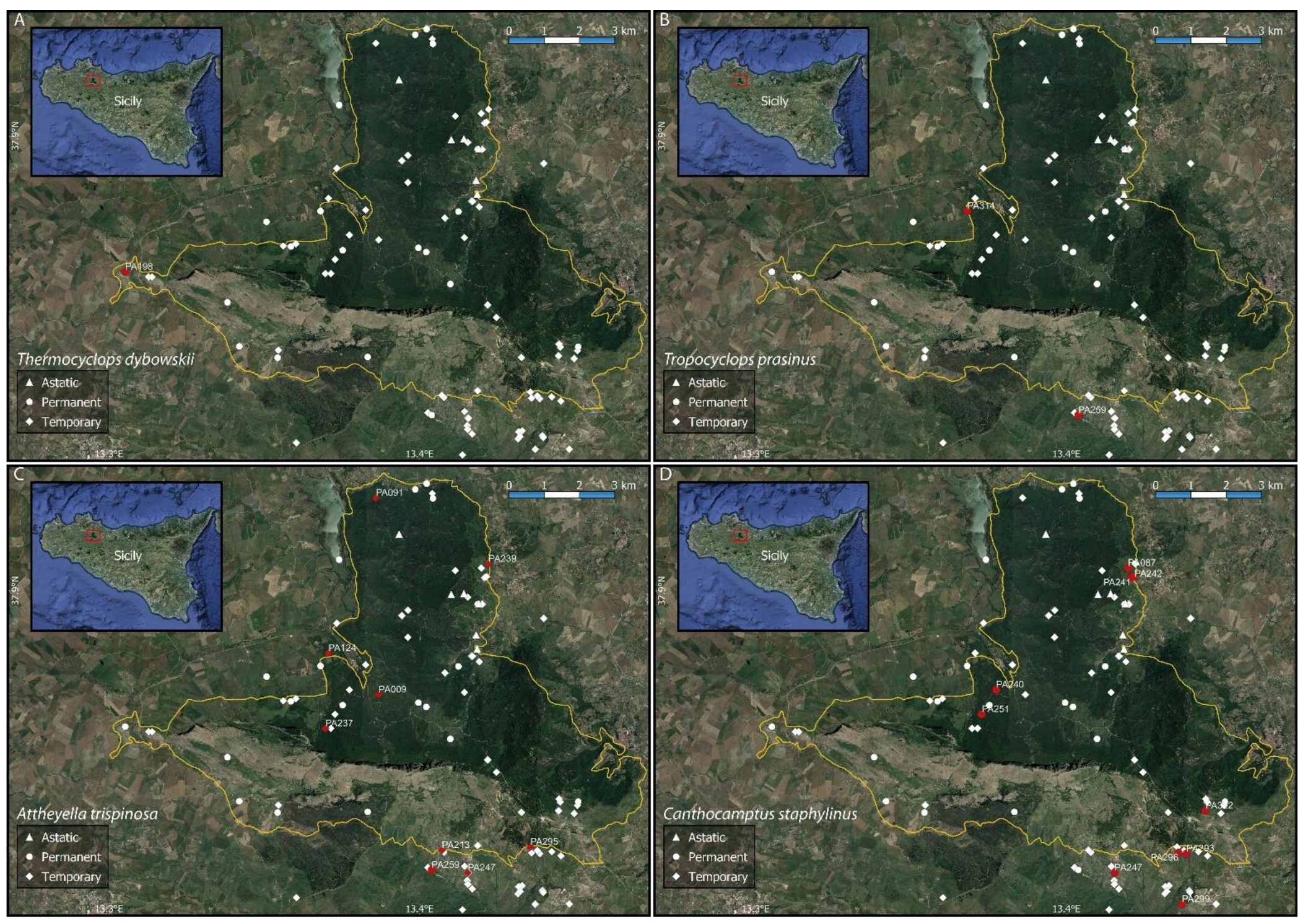
Figure A4.
Distribution maps of Thermocyclops dybowskii, (A), and Tropocyclops prasinus, (B), (Cyclopoida) and Attheyella trispinosa, (C), and Canthocamptus staphylinus, (D), (Harpacticoida). The solid yellow line shows the boundaries of the Nature Reserve “Bosco della Ficuzza, Rocca Busambra, Bosco del Cappelliere e Gorgo del Drago”.
Figure A4.
Distribution maps of Thermocyclops dybowskii, (A), and Tropocyclops prasinus, (B), (Cyclopoida) and Attheyella trispinosa, (C), and Canthocamptus staphylinus, (D), (Harpacticoida). The solid yellow line shows the boundaries of the Nature Reserve “Bosco della Ficuzza, Rocca Busambra, Bosco del Cappelliere e Gorgo del Drago”.

References
- Blondel, J.; Aronson, J.; Bodiou, J.Y.; Boeuf, G. The Mediterraneaen Region—Biological Diversity in Space and Time; Oxford University Press: Oxford, UK, 2010; ISBN 0-19-955798-5. [Google Scholar]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. Camb. Philos. Soc. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Cantonati, M.; Poikane, S.; Pringle, C.M.; Stevens, L.E.; Turak, E.; Heino, J.; Richardson, J.S.; Bolpagni, R.; Borrini, A.; Cid, N.; et al. Characteristics, main impacts, and stewardship of natural and artificial freshwater environments: Consequences for biodiversity conservation. Water 2020, 12, 260. [Google Scholar] [CrossRef] [Green Version]
- Dussart, B.H.; Defaye, D. Guides to the Identification of the Microinvertebrates of the Continental Waters of the World, 16. In Introduction to the Copepoda, 2nd ed.; SPB Academic Publishing: Leiden, The Netherlands, 2001. [Google Scholar]
- Marrone, F.; Naselli-Flores, L. First record and morphological features of Hemidiaptomus (Occidodiaptomus) ingens (Gurney, 1909) (Copepoda Calanoida) in Italy. J. Limnol. 2004, 63, 250–255. [Google Scholar] [CrossRef] [Green Version]
- Marrone, F.; Naselli-Flores, L. First record of a representative of the subfamily Paradiaptominae (Copepoda Calanoida Diaptomidae) in Italy: Metadiaptomus chevreuxi (Guerne and Richard, 1894). J. Limnol. 2005, 64, 89–92. [Google Scholar] [CrossRef]
- Marrone, F.; Castelli, G.; Barone, R.; Naselli-Flores, L. Ecology and distribution of calanoid copepods in Sicilian inland waters (Italy). SIL Proc. 2006, 29, 2150–2156. [Google Scholar] [CrossRef] [Green Version]
- Marrone, F.; Ortega, F.; Mesquita-Joanes, F.; Guerrero, F. On the occurrence of Metadiaptomus chevreuxi (Calanoida, Diaptomidae, Paradiaptominae) in the Iberian Peninsula, with notes on the ecology and distribution of its European populations. Water 2020, 12, 1989. [Google Scholar] [CrossRef]
- Marrone, F.; Lo Brutto, S.; Arculeo, M. Molecular evidence for the presence of cryptic evolutionary lineages in the freshwater copepod genus Hemidiaptomus G.O. Sars, 1903 (Calanoida, Diaptomidae). Hydrobiologia 2010, 644, 115–125. [Google Scholar] [CrossRef]
- Marrone, F.; Lo Brutto, S.; Hundsdoerfer, A.K.; Arculeo, M. Overlooked cryptic endemism in copepods: Systematics and natural history of the calanoid subgenus Occidodiaptomus Borutzky 1991 (Copepoda, Calanoida, Diaptomidae). Mol. Phylogenet. Evol. 2013, 66, 190–202. [Google Scholar] [CrossRef] [Green Version]
- Vecchioni, L.; Marrone, F.; Naselli-Flores, L. On the occurrence and distribution of Calanipeda aquaedulcis Kritschagin, 1873 (Copepoda, Calanoida, Pseudodiaptomidae) in Sicily, Italy, with some notes on coexistence and species replacement in calanoid copepods. Adv. Oceanogr. Limnol. 2019, 10, 19–23. [Google Scholar] [CrossRef]
- Margaritora, F.G.; Mastrantuono, L.; Crosetti, D.; Lombardi, F. Contributo allo studio della fauna ad entomostraci delle acque interne della Sicilia. Animalia 1982, 9, 87–102. [Google Scholar]
- Vecchioni, L.; Marrone, F.; Deidun, A.; Adepo-Gourene, B.; Froglia, C.; Sciberras, A.; Bariche, M.; Burak, A.Ç.; Foka-Corsini, M.; Arculeo, M. DNA Taxonomy Confirms the Identity of the Widely-Disjunct Mediterranean and Atlantic Populations of the Tufted Ghost Crab Ocypode cursor (Crustacea: Decapoda: Ocypodidae). Zool. Sci. 2019, 36, 322. [Google Scholar] [CrossRef] [PubMed]
- Vecchioni, L.; Arculeo, M.; Cottarelli, V.; Marrone, F. Range-wide phylogeography and taxonomy of the marine rock pools dweller Tigriopus fulvus (Fischer, 1860) (Copepoda, Harpacticoida). J. Zool. Syst. Evol. Res. 2021, 59, 839–857. [Google Scholar] [CrossRef]
- Calvo, S.; Barone, R.; Naselli-Flores, L.; Fradà Orestano, C.; Dongarrà, G.; Lugaro, A.; Genchi, G. Limnological studies on lakes and reservoirs of Sicily. Nat. Sicil. 1993, XVII, 288. [Google Scholar]
- Naselli-Flores, L.; Barone, R. Importance of water-level fluctuation on population dynamics of cladocerans in a hypertrophic reservoir (Lake Arancio, south-west Sicily, Italy). Hydrobiologia 1997, 360, 223–232. [Google Scholar] [CrossRef]
- Troia, A.; Adragna, F.; Campisi, P.; Campo, G.; Dia, M.G.; Ilardi, V.; La Mantia, T.; La Rosa, A.; Lo Valvo, M.; Muscarella, C.; et al. I pantani di Anguillara (Calatafimi Segesta, Trapani): Dati preliminari sulla biodiversità a supporto della tutela del biotopo. Nat. Sicil. 2016, XL, 171–200. [Google Scholar]
- Schifani, E.; Viviano, A.; Viviano, R.; Naselli-Flores, L.; Marrone, F. Different lineages of freshwater jellyfishes (Cnidaria, Olindiidae, Craspedacusta) invading Europe: Another piece of the puzzle from Sicily, Italy. Limnology 2019, 20, 143–151. [Google Scholar] [CrossRef]
- Marrone, F. The microcrustacean fauna of Sicily and the Central Mediterranean Sea area—Current knowledge and gaps to be filled. Polish J. Ecol. 2006, 54, 681–685. [Google Scholar]
- Marrone, F.; Castelli, G.; Naselli-Flores, L. Sicilian temporary ponds: An overview of the composition and affinities of their Crustacean biota. Int. Conf. Mediterr. Tempor. Ponds 2009, 189–202. [Google Scholar] [CrossRef]
- Drago, A.; Cartabellotta, D.; Lo Bianco, B.; Lombardo, M. Atlante Climatologico della Sicilia e Carta Climatica Della Sicilia (Scala 1:250000); Regione Siciliana, Assessorato Agricoltura e Foreste, Servizi allo Sviluppo, Unità Operativa di Agrometeorologia: Palermo, Italy, 2000. [Google Scholar]
- Gianguzzi, L.; La Mantia, T. Le serie di vegetazione della Riserva Naturale Orientata “Bosco Ficuzza, Rocca Busambra, Bosco del Cappelliere e Gorgo del Drago” con allegata carta della vegetazione (scala 1:20000). Nat. Sicil. 2004, XXVIII, 205–242. [Google Scholar]
- Gianguzzi, L. Il Paesaggio Vegetale Della Riserva Naturale Orientata “Bosco della Ficuzza, Rocca Busambra, Bosco del Cappelliere, Gorgo del Drago”. La Flora, le Fitocenosi, le Serie di Vegetazione; Officine Grafiche Riunite: Palermo, Italy, 2004. [Google Scholar]
- Caldarella, O. Censimento degli ambienti lentici e note distributive sulla flora idroigrofitica nell’area di Bosco Ficuzza (Sicilia Occidentale). Nat. Sicil. 2014, XXXVIII, 193–244. [Google Scholar]
- Bazan, G.; Castrorao Barba, A.; Rotolo, A.; Marino, P. Vegetation series as a marker of interactions between rural settlements and landscape: New insights from the archaeological record in Western Sicily. Landsc. Res. 2020, 45, 484–502. [Google Scholar] [CrossRef]
- Bazan, G.; Speciale, C.; Barba, A.C.; Cambria, S.; Miccichć, R.; Marino, P. Historical suitability and sustainability of Sicani mountains landscape (western Sicily): An integrated approach of phytosociology and archaeobotany. Sustain 2020, 12, 3201. [Google Scholar] [CrossRef] [Green Version]
- Troia, A.; Bazan, G.; Schicchi, R. Micromorphological approach to the systematics of Mediterranean Isoëtes species (Isoëtaceae, Lycopodiophyta): Analysis of the megaspore surface. Grana 2012, 51, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Pieri, V.; Martens, K.; Naselli-Flores, L.; Marrone, F.; Rossetti, G. Distribution of recent ostracods in inland waters of Sicily (Southern Italy). J. Limnol. 2006, 65, 1–8. [Google Scholar] [CrossRef]
- Pieri, V.; Marrone, F.; Martens, K.; Rossetti, G. An updated checklist of recent ostracods (Crustacea: Ostracoda) from inland waters of Sicily and adjacent small islands with notes on their distribution and ecology. Eur. Zool. J. 2020, 87, 714–740. [Google Scholar] [CrossRef]
- Marrone, F.; Vecchioni, L. The genus Daphnia in Sicily and Malta (Crustacea, Branchiopoda, Anomopoda). In Life on Islands. 1. Biodiversity in Sicily and Surrounding Islands. Studies Dedicated to Bruno Massa; La Mantia, T., Badalamenti, E., Carapezza, A., Lo Cascio, P., Troia, A., Eds.; Edizioni Danaus: Palermo, Italy, 2020; pp. 105–123. ISBN 978-88-97603-26-9. [Google Scholar]
- Vecchioni, L.; Deidun, A.; Sciberras, J.; Sciberras, A.; Marrone, F.; Arculeo, M. The late Pleistocene origin of the Italian and Maltese populations of Potamon fluviatile (Malacostraca: Decapoda): Insights from an expanded sampling of molecular data. Eur. Zool. J. 2017, 84, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Vecchioni, L.; Faraone, F.P.; Stoch, F.; Arculeo, M.; Marrone, F. Diversity and Distribution of the Inland Water Decapods of Sicily (Crustacea, Malacostraca). Diversity 2022, 14, 246. [Google Scholar] [CrossRef]
- Marrone, F.; Havenstein, K.; Tiedemann, R.; Ketmaier, V. Identification and characterization of five polymorphic microsatellite loci in the freshwater copepod Hemidiaptomus gurneyi (Copepoda: Calanoida: Diaptomidae). Ital. J. Zool. 2016, 83, 146–150. [Google Scholar] [CrossRef] [Green Version]
- Sparacio, I. Il Planorbis moquini (Requien, 1848) in Sicilia (Gastropoda, Pulmonata). Nat. Sicil. 1992, XVI, 129–133. [Google Scholar]
- Gerecke, R.; Marrone, F.; Sorgi, G.; Dossena, M.; Stoch, F. The water mites (Acari: Hydrachnidia) of the standing waters of Corsica, Sardinia and Sicily: Review and new data. Ital. J. Zool. 2014, 81, 389–408. [Google Scholar] [CrossRef] [Green Version]
- Marrone, F.; Sacco, F.; Kehlmaier, C.; Arizza, V.; Arculeo, M. Some like it cold: The glossiphoniid parasites of the Sicilian endemic pond turtle Emys trinacris (Testudines, Emydidae), an example of “parasite inertia”? J. Zool. Syst. Evol. Res. 2016, 54, 60–66. [Google Scholar] [CrossRef]
- Cianficconi, F.; de Pietro, R.; Gerecke, R.; Moretti, G. Catalogo dei Tricotteri della Sicilia. Mem. Della Soc. Entomol. Ital. 1999, 77, 259–309. [Google Scholar]
- Muscarella, C. Analisi Corologica, Biogeografica e del Popolamento. Master’s Thesis, University of Palermo, Palermo, Italy, 2014. [Google Scholar]
- Lo Valvo, M.; Faraone, F.P.; Giacalone, G.; Lillo, F. Fauna di Sicilia. Anfibi. Monografie Naturalistiche, 5; Danaus, Edizioni: Palermo, Italy, 2017. [Google Scholar]
- Vamberger, M.; Stuckas, H.; Sacco, F.; D’Angelo, S.; Arculeo, M.; Cheylan, M.; Corti, C.; Lo Valvo, M.; Marrone, F.; Wink, M.; et al. Differences in gene flow in a twofold secondary contact zone of pond turtles in southern Italy (Testudines: Emydidae: Emys orbicularis galloitalica, E. o. hellenica, E. trinacris). Zool. Scr. 2015, 44, 233–249. [Google Scholar] [CrossRef]
- Vecchioni, L.; Marrone, F.; Arculeo, M.; Fritz, U.; Vamberger, M. Stand out from the crowd: Small-scale genetic structuring in the endemic Sicilian pond turtle. Diversity 2020, 12, 343. [Google Scholar] [CrossRef]
- Belaiba, E.; Marrone, F.; Vecchioni, L.; Bahri-Sfar, L.; Arculeo, M. An exhaustive phylogeny of the combtooth blenny genus Salaria (Pisces, Blenniidae) shows introgressive hybridization and lack of reciprocal mtDNA monophyly between the marine species Salaria basilisca and Salaria pavo. Mol. Phylogenet. Evol. 2019, 135, 210–221. [Google Scholar] [CrossRef]
- Wagner, M.; Zogaris, S.; Berrebi, P.; Freyhof, J.; Koblmüller, S.; Magnan, P.; Laporte, M. Diversity and biogeography of Mediterranean freshwater blennies (Blenniidae, Salaria). Divers. Distrib. 2021, 27, 1832–1847. [Google Scholar] [CrossRef]
- Vecchioni, L.; Arculeo, M.; Hundt, P.J.; Marrone, F. The valid genus name of the European freshwater blennies, Ichthyocoris or Salariopsis (Teleostei: Blenniidae)? Zootaxa 2022, 5162, 99–100. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System; Open-Source Geospatial Foundation Project, 2022. [Google Scholar]
- Florencio, M.; Fernández-Zamudio, R.; Lozano, M.; Díaz-Paniagua, C. Interannual variation in filling season affects zooplankton diversity in Mediterranean temporary ponds. Hydrobiologia 2020, 847, 1195–1205. [Google Scholar] [CrossRef]
- Marrone, F.; Arculeo, M.; Georgiadis, C.; Stoch, F. On the non-malacostracan crustaceans (Crustacea: Branchiopoda, Copepoda, Ostracoda) from the inland waters of Fthiotida (Greece). Biogeographia 2019, 34, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Dussart, B.H. Les copépodes des Eaux Continentales d’Europe Occidentale, Tome I: Calanoïdes et Harpacticoïdes; Boubée et Cie: Paris, France, 1967. [Google Scholar]
- Dussart, B.H. Les Copépodes des Eaux Continentales d’Europe Occidentale, Tome 2: Cyclopöides et Biologie; Boubée et Cie: Paris, France, 1969. [Google Scholar]
- Kiefer, F. Das Zooplankton der Binnengewässer. Freilebende Copepoda. Die Binnengewässer, 26/2; Schweizerbart’sche Verlagbuchhandlung: Stuttgart, Germany, 1978. [Google Scholar]
- Einsle, U. Crustacea Copepoda. Calanoida und Cyclopoida. Süsswasserfauna Mitteleuropas, 8/4-1; Verlag, Gustav Fisher: Stuttgart, Germany, 1993. [Google Scholar]
- Einsle, U. Copepoda: Cyclopoida. Genera Cyclops, Megacyclops, Acanthocyclops. 10. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Dumont, H.J.F., Ed.; Backhuys Publishers: Leiden, The Netherlands, 1996. [Google Scholar]
- Karaytug, S. Copepoda: Cyclopoida. Genera Paracyclops, Ochridacyclops and key to the Eucyclopinae. 14. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Dumont, H.J.F., Ed.; Backhuys Publishers: Leiden, The Netherlands, 1999. [Google Scholar]
- Hołyńska, M.; Mirabdullayev, I.M.; Reid, J.W.; Ueda, H. Copepoda: Cyclopoida. Genera Mesocyclops and Thermocyclops. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Dumont, H.J.F., Ed.; Backhuys Publishers: Leiden, The Netherlands, 2003. [Google Scholar]
- Hołyńska, M. On the morphology and geographical distribution of some problematic South Palearctic Cyclops (Copepoda: Cyclopidae). J. Nat. Hist. 2008, 42, 2011–2039. [Google Scholar] [CrossRef]
- Alekseev, V.R.; Defaye, D. Taxonomic differentiation and world geographical distribution Of the Eucyclops serrulatus Group (Copepoda, Cyclopidae, Eucyclopinae). Stud. Freshw. Copepoda Vol. Honour Bernard Dussart 2011, 16, 41–72. [Google Scholar]
- Miracle, M.R.; Alekseev, V.; Monchenko, V.; Sentandreu, V.; Vicente, E. Molecular-genetic-based contribution to the taxonomy of the Acanthocyclops robustus group. J. Nat. Hist. 2013, 47, 863–888. [Google Scholar] [CrossRef]
- Ruffo, S.; Stoch, F. Checklist and Distribution of the Italian Fauna: 10,000 Terrestrial and Inland Water Species. Memorie del Museo Civico di Storia Naturale di Verona, 2.Serie, Sezione Scienze della Vita 17, with CD-ROM; Ruffo, S., Stoch, F., Eds.; Comune di Verona: Verona, Italy, 2006; Volume 91, ISBN 8889230096. [Google Scholar]
- Alfonso, G.; Stoch, F.; Marrone, F. An annotated checklist and bibliography of the Diaptomidae (Copepoda, Calanoida) of Italy, Corsica, and the Maltese islands. J. Limnol. 2021, 80, 257–272. [Google Scholar] [CrossRef]
- Gurney, R. British Fresh-Water Copepoda—Volume III; Ray Society: London, UK, 1933. [Google Scholar]
- Monchenko, V.I. [Cyclopidae Fauna of the Ukraine] Fauna Ukrainii, 27; Naukova Dumka: Kyiv, Ukraine, 1974. [Google Scholar]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Colwell, R.K.; Mao, C.X.; Chang, J. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 2004, 85, 2717–2727. [Google Scholar] [CrossRef] [Green Version]
- Marrone, F.; Fontaneto, D.; Naselli-Flores, L. Cryptic diversity, niche displacement and our poor understanding of taxonomy and ecology of aquatic microorganisms. Hydrobiologia 2022. [Google Scholar] [CrossRef]
- Alekseev, V.R.; Dumont, H.J.; Pensaert, J.; Baribwegure, D.; Vanfleteren, J.R. A redescription of Eucyclops serrulatus (Fischer, 1851) (Crustacea: Copepoda: Cyclopoida) and some related taxa, with a phylogeny of the E. serrulatus-group. Zool. Scr. 2006, 35, 123–147. [Google Scholar] [CrossRef]
- Sukhikh, N.; Alekseev, V. Genetic and morphological heterogeneity within Eucyclops serrulatus (Fischer, 1851) (Crustacea: Copepoda: Cyclopidae). J. Nat. Hist. 2015, 49, 2929–2953. [Google Scholar] [CrossRef]
- Vigna Taglianti, A.; Audisio, P.A.; Belfiore, C.; Biondi, M.; Bologna, M.A.; Carpaneto, G.M.; De Biase, A.; De Felici, S.; Piattella, E.; Racheli, T.; et al. Riflessioni di gruppo sui corotipi fondamentali della fauna W-paleartica ed in particolare italiana. Biogeogr. J. Integr. Biogeogr. 1992, 16, 159–179. [Google Scholar] [CrossRef] [Green Version]
- Vecchioni, L.; Arculeo, M.; Marrone, F. Molecular data attest to the occurrence of autochthonous Daphnia pulex (Crustacea, Branchiopoda) populations in Sicily, Italy. Adv. Oceanogr. Limnol. 2021, 12, 29–35. [Google Scholar] [CrossRef]
- Gauthier, H. Recherches sur la Faune des eaux Continentales de l’Algérie et de la Tunisie; Imprimerie Minerva: Alger, Algeria, 1928. [Google Scholar]
- Marrone, F.; Korn, M.; Stoch, F.; Naselli-Flores, L.; Turki, S. Updated checklist and distribution of large Branchiopods (Branchiopoda: Anostraca, Notostraca, Spinicaudata) in Tunisia. Biogeographia 2016, 31, 27–53. [Google Scholar] [CrossRef] [Green Version]
- Alfonso, G.; Beccarisi, L.; Pieri, V.; Frassanito, A.; Belmonte, G. Using crustaceans to identify different pond types. A case study from the Alta Murgia National Park, Apulia (South-eastern Italy). Hydrobiologia 2016, 782, 53–69. [Google Scholar] [CrossRef]
- Seminara, M.; Vagaggini, D.; Stoch, F. A comparison of Cladocera and Copepoda as indicators of hydroperiod length in Mediterranean ponds. Hydrobiologia 2016, 782, 71–80. [Google Scholar] [CrossRef]
- Vecchioni, L.; Sala, J.; Arculeo, M.; Marrone, F. On the occurrence of Eudiaptomus gracilis (G.O. Sars, 1863) (Copepoda: Diaptomidae) in Madeira (Portugal). Limnetica 2020, 39, 571–578. [Google Scholar] [CrossRef]
- Alekseev, V.R. Confusing invader: Acanthocyclops americanus (Copepoda: Cyclopoida) and its biological, anthropogenic and climate-dependent mechanisms of rapid distribution in Eurasia. Water 2021, 13, 1423. [Google Scholar] [CrossRef]
- Naselli-Flores, L.; Marrone, F. Different invasibility of permanent and temporary waterbodies in a semiarid Mediterranean Island. Inland Waters 2019, 9, 411–421. [Google Scholar] [CrossRef]
- Marrone, F.; Naselli-Flores, L. A review on the animal xenodiversity in sicilian inland waters (Italy). Adv. Oceanogr. Limnol. 2015, 6, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Anufriieva, E.; Hołyńska, M.; Shadrin, N. Current invasions of Asian Cyclopid species (Copepoda: Cyclopidae) in Crimea, with taxonomical and zoogeographical remarks on the hypersaline and freshwater fauna. Ann. Zool. 2014, 64, 109–130. [Google Scholar] [CrossRef]
- Pesce, G.L.; Galassi, D.P. Copepodi di acque sotterranee della Sicilia. Animalia 1987, 14, 193–235. [Google Scholar]
- Hortal, J.; De Bello, F.; Diniz-Filho, J.A.F.; Lewinsohn, T.M.; Lobo, J.M.; Ladle, R.J. Seven shortfalls that beset large-scale knowledge of biodiversity. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 523–549. [Google Scholar] [CrossRef] [Green Version]
- Marrone, F.; Alfonso, G.; Stoch, F.; Pieri, V.; Alonso, M.; Dretakis, M.; Naselli-Flores, L. An account on the non-malacostracan crustacean fauna from the inland waters of Crete, Greece, with the synonymization of Arctodiaptomus piliger Brehm, 1955 with Arctodiaptomus alpinus (Imhof, 1885) (Copepoda: Calanoida). Limnetica 2019, 38, 167–187. [Google Scholar] [CrossRef]
- Stella, E.; Margaritora, F.G.; Cottarelli, V. La fauna ad entomostraci di acque astatiche della Sardegna nord orientale. Ric. Biol. Ecol. Rend. Accad. Naz. XL 1972, 4, 3–50. [Google Scholar]
- Stella, E.; Margaritora, F.G. Contributo alla conoscenza della fauna ad entomostraci di acque astatiche della Sardegna (zone Nord occidentale e Centrale). Considerazioni ecologiche e biogeografiche. Rend. Accad. Naz. dei XL 1975, 5, 1–11. [Google Scholar]
- Pretus, J.L. A commented check-list of the Balearic Branchiopoda (Crustacea). Limnetica 1990, 6, 157–164. [Google Scholar] [CrossRef]
- Stephanides, T. A survey of the freshwater biology of Corfu and of certain other regions of Greece. Prakt. Hell. Hydrobiol. Inst. 1948, 2, 1–263. [Google Scholar]
- Jaume, D. Calanoides (Crustacea: Copepoda) de las aigues continentals baleariques. Bolletí Soc. d’Història Nat. Balear. 1989, 33, 207–219. [Google Scholar]
Figure 1.
Distribution of the sampling sites in the study area (A) and local distribution of Calanoida (B), Cyclopoida (C), and Harpacticoida (D). The solid yellow line shows the boundaries of the Nature Reserve “Bosco della Ficuzza, Rocca Busambra, Bosco del Cappelliere e Gorgo del Drago”.
Figure 1.
Distribution of the sampling sites in the study area (A) and local distribution of Calanoida (B), Cyclopoida (C), and Harpacticoida (D). The solid yellow line shows the boundaries of the Nature Reserve “Bosco della Ficuzza, Rocca Busambra, Bosco del Cappelliere e Gorgo del Drago”.

Figure 2.
An example of the different habitat type sampled (see Table 1 and Appendix A for further details). (A) PA124 on 17 March 2022 (inhabited by Mixodiaptomus kupelwieseri, Cyclops divergens, Diacyclops lubbocki, Attheyella trispinosa); (B) PA214 on 7 March 2021 (Diaptomus cyaneus, Megacyclops viridis s.l.); (C) PA238 on 24 April 2021 (C. divergens); (D) PA246 on 7 March 2021 (Cyclops ankyrae); (E) PA253 on 13 February 2021 (no copepods); (F) PA312 on 17 March 2021 (Acanthocyclops americanus).
Figure 2.
An example of the different habitat type sampled (see Table 1 and Appendix A for further details). (A) PA124 on 17 March 2022 (inhabited by Mixodiaptomus kupelwieseri, Cyclops divergens, Diacyclops lubbocki, Attheyella trispinosa); (B) PA214 on 7 March 2021 (Diaptomus cyaneus, Megacyclops viridis s.l.); (C) PA238 on 24 April 2021 (C. divergens); (D) PA246 on 7 March 2021 (Cyclops ankyrae); (E) PA253 on 13 February 2021 (no copepods); (F) PA312 on 17 March 2021 (Acanthocyclops americanus).

Figure 3.
Species accumulation curves (S Mean) and performance of estimators of copepod species richness (ICE Mean, Jack 1 Mean, Chao 2 Mean) and of uniqueness (Uniques Mean). Number of sampled sites is on the x-axis; cumulative species richness is on the y-axis.
Figure 3.
Species accumulation curves (S Mean) and performance of estimators of copepod species richness (ICE Mean, Jack 1 Mean, Chao 2 Mean) and of uniqueness (Uniques Mean). Number of sampled sites is on the x-axis; cumulative species richness is on the y-axis.

Table 1.
List of the sampled sites. When available, the codes used by Caldarella [24] are included between brackets. Geographical coordinates are expressed as decimal degrees (Map Datum: WGS84). Hydroperiod = A, astatic; P, permanent; T, temporary.
Table 1.
List of the sampled sites. When available, the codes used by Caldarella [24] are included between brackets. Geographical coordinates are expressed as decimal degrees (Map Datum: WGS84). Hydroperiod = A, astatic; P, permanent; T, temporary.
| Code | Site Name | Latitude N | Longitude E | Elevation (m a.s.l.) | Habitat Type | Hydroperiod |
|---|---|---|---|---|---|---|
| PA009 | Gorgo dei palermitani (SSP1) | 37.875203 | 13.385595 | 770 | Pond | T |
| PA012 | Valle Maria 1 | 37.884058 | 13.417820 | 614 | Pool | T |
| PA079 | Gorgo Cerro (SSP2) | 37.889991 | 13.394688 | 750 | Pond | T |
| PA080 | Gorgo di Glaviano (SSP6) | 37.895409 | 13.438412 | 470 | Pond | T |
| PA081 | Gorgo Lungo (SP1) | 37.901131 | 13.408438 | 904 | Pond | A |
| PA082 | Gorgo Tondo (Gorgo del Drago, SP2) | 37.901389 | 13.412501 | 854 | Pond | A |
| PA084 | Pozza degli incidenti | 37.882777 | 13.381256 | 680 | Pool | T |
| PA085 | Pozza di C.da Castellaccio | 37.893280 | 13.371701 | 593 | Pool | T |
| PA086 | C.da Cannitello (PT18) | 37.900496 | 13.413772 | 868 | Pool | T |
| PA087 | Pozza dello Zù Santino (PT20) | 37.907907 | 13.417980 | 845 | Pool | T |
| PA091 | Gorgo Margiazzu (PT10) | 37.925365 | 13.383588 | 526 | Pond | T |
| PA120 | Laghetto Coda di Riccio 1 (AI16) | 37.872286 | 13.400995 | 876 | Reservoir | P |
| PA121 | Laghetto Coda di Riccio 2 (AI15) | 37.873333 | 13.398449 | 870 | Reservoir | P |
| PA124 | Gorgo di Gaetanello (PT7) | 37.885585 | 13.369187 | 671 | Pond | T |
| PA133 | Pozza di Rocca Argenteria (FIA1) | 37.864722 | 13.311705 | 553 | Pool | T |
| PA181 | Invaso dell’Arcera (SCA3) | 37.916233 | 13.391296 | 596 | Reservoir | A |
| PA198 | Stagno agricolo del Frattina (AI1) | 37.865893 | 13.304113 | 482 | Reservoir | P |
| PA204 | Stagno sotto cozzo Bileo (AZ9) | 37.907135 | 13.409628 | 943 | Pond | T |
| PA212 | Pozza 1 della Busambra | 37.835266 | 13.407517 | 831 | Pool | T |
| PA213 | Pozza 2 della Busambra | 37.835778 | 13.406812 | 837 | Pool | T |
| PA214 | Pozza 3 della Busambra (SPP4) | 37.837101 | 13.418269 | 859 | Pond | T |
| PA235 | Casa Franco (SSP5) | 37.898886 | 13.417320 | 782 | Reservoir | A |
| PA236 | Casa Franco (AI23) | 37.898747 | 13.417050 | 749 | Reservoir | P |
| PA237 | C.da Balata (PT6) | 37.866354 | 13.368611 | 918 | Pool | T |
| PA238 | Laghetto forestale di C.da Ramusa (AI12) | 37.872371 | 13.374066 | 807 | Reservoir | P |
| PA239 | C.da Sovarita (PT23) | 37.909001 | 13.420300 | 832 | Pool | T |
| PA240 | Gorgo della Porcaria (PT9) | 37.876317 | 13.376156 | 752 | Pool | T |
| PA241 | C.da Cannitello (PT21) | 37.905479 | 13.419079 | 817 | Pool | T |
| PA242 | C.da Cannitello 2 (PT22) | 37.905731 | 13.419733 | 812 | Pool | T |
| PA243 | C.da Balata (AZ5) | 37.866396 | 13.370395 | 940 | Pond | T |
| PA244 | C.da Cannitello (AZ17) | 37.898713 | 13.418518 | 780 | Pond | T |
| PA245 | Marosella-Ramusa | 37.820679 | 13.413760 | 713 | Pond | T |
| PA246 | Pozza 3 del recinto | 37.831756 | 13.414239 | 795 | Pool | T |
| PA247 | Pozza Pippinello bassa | 37.830114 | 13.415278 | 750 | Pool | T |
| PA248 | Pozza Pippinello alta | 37.830064 | 13.415484 | 752 | Pool | T |
| PA249 | Pozza Junchi | 37.831304 | 13.402387 | 747 | Pool | T |
| PA250 | Pozza ai cipressi | 37.822586 | 13.448174 | 718 | Pond | T |
| PA251 | C.da Ramusa (PT8) | 37.870010 | 13.371656 | 832 | Pool | T |
| PA252 | Strada P.no Tramontana (DCT1) | 37.845778 | 13.432219 | 1165 | Pool | T |
| PA253 | Valle Maria (ANT1) | 37.876156 | 13.413202 | 802 | Pool | T |
| PA256 | Piano Carduna (PT12) | 37.895505 | 13.392657 | 706 | Pool | T |
| PA258 | Zotta Frascino (PT14) | 37.896893 | 13.394595 | 773 | Pool | T |
| PA259 | Pozza profonda | 37.830691 | 13.403638 | 742 | Pool | P |
| PA261 | C.da Bivieri (SSP3) | 37.890910 | 13.416672 | 653 | Pond | A |
| PA264 | Stagno artificiale del vivaio forestale (AI19) | 37.882770 | 13.411161 | 651 | Reservoir | P |
| PA265 | Stagno presso vivaio forestale (AZ8) | 37.881071 | 13.406820 | 706 | Pond | T |
| PA266 | Stagno artificiale sopra Alpe Cucco (AI18) | 37.864211 | 13.408923 | 1002 | Reservoir | P |
| PA267 | Lago Scanzano | 37.909439 | 13.372096 | 518 | Reservoir | P |
| PA268 | Piano Prani 1 (AZ21) | 37.859039 | 13.421083 | 1068 | Pool | T |
| PA269 | Piano Prani 2 (AZ24) | 37.855905 | 13.423935 | 1071 | Pond | T |
| PA270 | Pozza “strada Corleone” | 37.823028 | 13.360222 | 838 | Pool | T |
| PA271 | C.da Bivieri (AZ15) | 37.887333 | 13.417115 | 613 | Pond | A |
| PA272 | Laghetto agricolo Massariota | 37.929219 | 13.399892 | 615 | Reservoir | P |
| PA273 | C.da Massariota (AI14) | 37.927702 | 13.396248 | 598 | Pond | P |
| PA274 | Pozza di C.da Pertuso (PT16) | 37.926722 | 13.401774 | 669 | Pool | T |
| PA275 | Laghetto di Piano Cancemi (AI17) | 37.925492 | 13.401974 | 677 | Reservoir | P |
| PA276 | Strada Valle Agnese 1 | 37.885479 | 13.415342 | 610 | Pool | T |
| PA277 | Strada Valle Agnese 2 | 37.885430 | 13.415636 | 610 | Pool | T |
| PA278 | Rocca Argenteria (FIA2) | 37.864707 | 13.312803 | 568 | Pool | T |
| PA292 | Gorgo 4 Tummini (SSP7) | 37.835880 | 13.442202 | 925 | Pond | T |
| PA293 | Pozza Tonda (PPS2) | 37.835343 | 13.438344 | 891 | Pool | T |
| PA294 | Pozza Grande | 37.836062 | 13.437582 | 890 | Pool | T |
| PA295 | Pozza Bosco Acero | 37.836986 | 13.435305 | 913 | Pool | T |
| PA296 | Pozza Rovo | 37.835482 | 13.435739 | 887 | Pool | T |
| PA297 | Pozza Carrareccia Giardinello | 37.825890 | 13.439420 | 742 | Pool | T |
| PA298 | Canaletta della gebbia | 37.825462 | 13.439689 | 738 | Concrete channel | T |
| PA299 | Pozza 4 Giardinello | 37.822380 | 13.437167 | 662 | Pool | T |
| PA300 | Pozza 5 Giardinello | 37.827061 | 13.432886 | 715 | Pool | T |
| PA301 | Pozza 6 Giardinello | 37.826803 | 13.431717 | 703 | Pool | T |
| PA302 | Pozza 7 Giardinello | 37.825008 | 13.431589 | 665 | Pool | T |
| PA303 | Pozza 8 Giardinello | 37.825619 | 13.432246 | 675 | Pool | T |
| PA304 | Letto fiume alto 1 | 37.828162 | 13.415172 | 765 | Pool | T |
| PA305 | Letto fiume alto 2 | 37.827262 | 13.415334 | 702 | Pool | T |
| PA306 | Letto fiume alto 3 | 37.827196 | 13.415326 | 702 | Pool | T |
| PA308 | Letto fiume alto 5 | 37.826046 | 13.416791 | 677 | Pool | T |
| PA309 | Stagno di Piano Guddemi (AI26) | 37.834858 | 13.445156 | 951 | Reservoir | T |
| PA311 | Stagno artificiale | 37.879307 | 13.349360 | 651 | Reservoir | P |
| PA312 | Contrada Bifarera (AI11) | 37.873129 | 13.357364 | 790 | Reservoir | P |
| PA313 | Contrada Bifarera (PT3) | 37.873220 | 13.354886 | 798 | Pool | T |
| PA314 | Stagno artificiale | 37.882280 | 13.366696 | 672 | Reservoir | P |
| PA315 | Contrada Bifarera (PPS1) | 37.873802 | 13.358911 | 757 | Pond | T |
| PA317 | C.da Pirrello (AI13) | 37.845225 | 13.382714 | 891 | Reservoir | P |
| PA318 | C.da Casale (AI4) | 37.858561 | 13.337228 | 784 | Reservoir | P |
| PA319 | C.da Casale 1 (AI5) | 37.847380 | 13.341298 | 547 | Reservoir | P |
| PA320 | C.da Casale 2 (AI10) | 37.844661 | 13.353724 | 710 | Reservoir | P |
| PA321 | C.da Casale 3 | 37.846572 | 13.353964 | 715 | Pool | T |
| PA322 | Pozza | 37.846346 | 13.444175 | 1097 | Pool | T |
| PA323 | Pozza Roccia | 37.848604 | 13.444704 | 1069 | Rock pool | T |
| PA324 | Pizzo Angelo (ANT3) | 37.849362 | 13.444248 | 1030 | Reservoir | T |
| PA325 | C.da Acqua Jenco (AZ26) | 37.848741 | 13.450455 | 928 | Reservoir | P |
| PA326 | C.da Acqua Jenco 1 (PT27) | 37.847783 | 13.450237 | 959 | Pool | T |
| PA327 | C.da Acqua Jenco 2 (PT26) | 37.845733 | 13.449234 | 1023 | Pool | T |
Table 2.
Checklist and distribution of the copepods collected in the study area.
| Calanoida | Occurrence Sites |
|---|---|
| Arctodiaptomus sp. | PA309 |
| Copidodiaptomus numidicus (Gurney, 1909) | PA198; PA267 |
| Diaptomus cyaneus Gurney, 1909 | PA214; PA247; PA269; PA292; PA295; PA309 |
| Diaptomus serbicus Gjorgjevič, 1907 | PA084; PA212; PA213; PA245; PA299; PA270 |
| Hemidiaptomus gurneyi (Roy, 1927) | PA079; PA309 |
| Mixodiaptomus kupelwieseri (Brehm, 1907) | PA009; PA079; PA081; PA085; PA086; PA087; PA091; PA124; PA181; PA204; PA240; PA241; PA243; PA245; PA265; PA270; PA299; PA315 |
| Cyclopoida | |
| Acanthocyclops americanus (Marsh, 1893) | PA272; PA273; PA311; PA312 |
| Cyclops ankyrae Mann, 1940 | PA012; PA246 |
| Cyclops divergens Lindberg, 1936 | PA079; PA080; PA081; PA082; PA082; PA086; PA087; PA091; PA124; PA181; PA204; PA237; PA238; PA240; PA242; PA244; PA250; PA256; PA258; PA267; PA268; PA272; PA275; PA276; PA277; PA304; PA305; PA309; PA315; PA320; PA322; PA327 |
| Diacyclops bicuspidatus (Claus, 1857) | PA081 |
| Diacyclops bisetosus (Rehberg, 1880) | PA079; PA251; PA261 |
| Diacyclops lubbocki (Brady, 1869) | PA009; PA012; PA079; PA084; PA085; PA086; PA124; PA204; PA239; PA240; PA241; PA243; PA245; PA247; PA265; PA274; PA292; PA295; PA296; PA300; PA302; PA304; PA305; PA313; PA323 |
| Eucyclops serrulatus (Fischer, 1851) | PA081; PA082; PA085; PA120; PA121; PA198; PA236; PA242; PA249; PA271; PA294; PA297; PA298; PA301; PA317; PA318; PA319; PA324; PA325 |
| Macrocyclops albidus (Jurine, 1820) | PA236; PA264; PA297; PA321 |
| Megacyclops viridis s.l. (Jurine, 1820) | PA009; PA080; PA081; PA082; PA091; PA204; PA214; PA235; PA240; PA241; PA242; PA247; PA248; PA249; PA259; PA261; PA269; PA295; PA296; PA301; PA303; PA306; PA308; PA309; PA315; PA319; PA326; PA327 |
| Mesocyclops leuckarti (Claus, 1857) | PA264; PA303 |
| Metacyclops minutus (Claus, 1863) | PA080; PA087; PA213; PA248; PA252 |
| Microcyclops varicans (Sars, 1863) | PA299 |
| Thermocyclops dybowskii (Landé, 1890) | PA198 |
| Tropocyclops prasinus (Fischer, 1866) | PA314 |
| Harpacticoida | |
| Attheyella trispinosa (Brady, 1880) | PA009; PA091; PA124; PA213; PA237; PA239; PA247; PA259; PA295; PA299 |
| Canthocamptus staphylinus (Jurine, 1820) | PA087; PA240; PA241; PA242; PA247; PA251; PA293; PA296; PA322 |
Table 3.
Ranges of environmental conditions under which the collected copepod species were found in the study area. N, number of occurrence localities; EC, electrical conductivity (µS/cm). Hydroperiod = A, astatic; P, permanent; T, temporary.
Table 3.
Ranges of environmental conditions under which the collected copepod species were found in the study area. N, number of occurrence localities; EC, electrical conductivity (µS/cm). Hydroperiod = A, astatic; P, permanent; T, temporary.
| Taxon | N | Elevation (m a.s.l.) | EC (µS/cm) | Water Temperature (°C) | Hydroperiod | |||
|---|---|---|---|---|---|---|---|---|
| Range | Mean ± SD | Range | Mean ± SD | Range | Mean ± SD | |||
| Calanoida Sars, 1903 | ||||||||
| Arctodiaptomus sp. | 1 | 951 | 39 | 13 | T | |||
| Copidodiaptomus numidicus | 2 | 482–518 | 500 ± 25.5 | 419–482 | 450.5 ± 44.5 | 12.1–15 | 13.5 ± 2 | P |
| Diaptomus cyaneus | 6 | 750–1071 | 847 ± 114.8 | 39–1405 | 666.6 ± 588.6 | 9.9–15.7 | 13.3 ± 1.9 | T |
| Diaptomus serbicus | 6 | 662–838 | 760 ± 83.9 | 50–1305 | 613.3 ± 463.4 | 10–13 | 11.8 ± 1.1 | T |
| Hemidiaptomus gurneyi | 2 | 750–951 | 850 ± 142.1 | 93–208 | 123.5 ± 119.5 | 13–13.4 | 13.2 ± 0.3 | T |
| Mixodiaptomus kupelwieseri | 18 | 526–943 | 769 ± 109.1 | 50–1124 | 299.4 ± 300.6 | 6–18.8 | 11.7 ± 3.9 | A-T |
| Cyclopoida Burmeister, 1834 | ||||||||
| Acanthocyclops americanus | 4 | 598–790 | 663 ± 87.2 | 478–626 | 523.7 ± 69.1 | 10–18.3 | 14.8 ± 3.8 | P |
| Cyclops ankyrae | 2 | 614–795 | 704 ± 128 | 250–1335 | 792.5 ± 767.2 | 10–14.5 | 12.2 ± 3.2 | T |
| Cyclops divergens | 31 | 470–1097 | 774 ± 151.7 | 20–4130 | 407.3 ± 797.8 | 6–21 | 10.2 ± 4.2 | A-P-T |
| Diacyclops bicuspidatus | 1 | 904 | 109 | 18.8 | A | |||
| Diacyclops bisetosus | 3 | 653–832 | 741 ± 89.5 | 71–585 | 272 ± 274.7 | 8–13.6 | 11.5 ± 3.1 | A-T |
| Diacyclops lubbocki | 25 | 593–1069 | 780 ± 108.5 | 40–992 | 237.1 ± 254.9 | 4.8–18 | 10.5 ± 3.1 | T |
| Eucyclops serrulatus | 19 | 482–1030 | 786 ± 139.1 | 21–714 | 232.1 ± 228.8 | 5–23.5 | 11 ± 5.4 | A-P-T |
| Macrocylops albidus | 4 | 651–749 | 714 ± 44.6 | 27–317 | 136.7 ± 157.4 | 5.7–12.6 | 8.8 ± 3.5 | P-T |
| Megacyclops viridis s.l. | 27 | 470–1071 | 790 ± 130.9 | 39–1287 | 315.4 ± 356.1 | 6–17 | 10.9 ± 3.3 | A-P-T |
| Mesocyclops leuckarti | 2 | 651–675 | 663 ± 17 | 83–317 | 200 ± 165.5 | 9.6–12.6 | 11.1 ± 2.1 | P-T |
| Metacyclops minutus | 5 | 470–1165 | 814 ± 248.4 | 170–1028 | 606.6 ± 370.6 | 5.7–14.5 | 10.1 ± 3.4 | T |
| Microcyclops varicans | 1 | 662 | 90 | 9 | T | |||
| Thermocyclops dybowskii | 1 | 482 | 182 | 15 | P | |||
| Tropocyclops prasinus | 2 | 672 | 296 | 11.2 | P | |||
| Harpacticoida Sars, 1903 | ||||||||
| Attheyella trispinosa | 9 | 526–918 | 775 ± 108.5 | 40–1405 | 369.5 ± 490.1 | 9–17 | 12.3 ± 2.7 | P-T |
| Canthocamptus staphylinus | 10 | 750–1097 | 850 ± 98.7 | 20–992 | 226 ± 343.1 | 7–13 | 9.9 ± 2.1 | T |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marchese, M.; Vecchioni, L.; Bazan, G.; Arculeo, M.; Marrone, F. The Inland Water Copepod Fauna of a Traditional Rural Landscape in a Mediterranean Island (Crustacea, Copepoda). Water 2022, 14, 2168. https://doi.org/10.3390/w14142168
AMA Style
Marchese M, Vecchioni L, Bazan G, Arculeo M, Marrone F. The Inland Water Copepod Fauna of a Traditional Rural Landscape in a Mediterranean Island (Crustacea, Copepoda). Water. 2022; 14(14):2168. https://doi.org/10.3390/w14142168
Chicago/Turabian StyleMarchese, Marta, Luca Vecchioni, Giuseppe Bazan, Marco Arculeo, and Federico Marrone. 2022. "The Inland Water Copepod Fauna of a Traditional Rural Landscape in a Mediterranean Island (Crustacea, Copepoda)" Water 14, no. 14: 2168. https://doi.org/10.3390/w14142168
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.