
Effects of Ethyl Lauroyl Arginate (LAE) on Biofilm Detachment: Shear Rate, Concentration, and Dosing Time
by
Peng-Fei Sun
1,2,†, 
Taek-Seung Kim
1,3,†,
So-Young Ham
1,
Yong-Sun Jang
1 and
Hee-Deung Park
1,4,* 1
School of Civil, Environmental and Architectural Engineering, Korea University, Seoul 02841, Korea
2
Department of Civil Engineering, The University of Hong Kong, Hong Kong 999077, China
3
Research Solution Center, Institute for Basic Science, Daejeon 34126, Korea
4
KU-KIST Graduate School of Converging Science and Technology, Korea University, Seoul 02841, Korea
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Water 2022, 14(14), 2158; https://doi.org/10.3390/w14142158
Submission received: 7 June 2022
/
Revised: 5 July 2022
/
Accepted: 6 July 2022
/
Published: 7 July 2022
(This article belongs to the Special Issue Membrane Technology for Water Treatment and Desalination)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Biofilm formation is one of the main obstacles in membrane treatment. The non-oxidizing biocide ethyl lauroyl arginate (LAE) is promising for mitigating biofilm development on membrane surfaces. However, the operating conditions of LAE and their impact on biofilm detachment are not comprehensively understood. In this study, a real-time in vitro flow cell system was utilized to observe biofilm dispersal caused by the shear rate, concentration, and treatment time of LAE. This confirmed that the biofilm was significantly reduced to 68.2% at a shear rate of 3.42 s−1 due to the increased physical lifting force. LAE exhibited two different mechanisms for bacterial inactivation and biofilm dispersal. Biofilms treated with LAE at sub-growth inhibitory concentrations for a longer time could effectively detach the biofilm formed on the surface of the glass slides, which can be attributed to the increased motility of microorganisms. However, a high concentration (i.e., bactericidal concentration) of LAE should be seriously considered because of the inactivated sessile bacteria and their residual debris remaining on the surface. This study sheds light on the effect of LAE on biofilm detachment and provides insights into biofouling mitigation during the membrane process.
1. Introduction
Membrane-based technologies are widely applied in seawater desalination, wastewater treatment, water reclamation, and resource and energy recovery [1,2,3]. However, the performance of membrane processes is largely limited by membrane fouling, which can increase the feed pressure drop, deteriorate the permeate quality, and reduce the membrane lifespan [4,5]. Compared to other fouling types (organic and inorganic) and scaling, biofouling caused by microorganism deposition and biofilm formation on membrane surfaces has attracted special attention [6,7,8,9]. Because the feed solution inevitably contains various nutrients and microorganisms even after rigorous pretreatment, bacteria can establish an adhesive biofilm by secreting extracellular polymeric substances (EPS) to protect them from the surrounding environment, which is highly intractable for sustainable membrane operation [5,10,11,12,13]. Previous studies revealed that biofilm-enhanced concentration polarization (CP) can greatly impair the overall performance of membranes in both pressure-driven and osmotically driven membrane processes [6,14]. Hence, it is necessary to develop effective membrane biofouling control strategies and novel characterization approaches.
In addition to the pretreatment of the feed solution to prolong the time at which biofilm forms on the membrane and spacer surfaces, substantial efforts were made to mitigate biofouling, such as physical flushing, osmotic backwashing, chemical cleaning, and membrane and spacer modification [7,8,15,16,17]. The prevention of initial biofilm deposition on the membrane surface is important for maintaining sustainable water production, which is why many studies focused on membrane surface modification to mitigate bacterial adhesion [18,19,20]. Nevertheless, there is an urgent need to develop effective approaches (e.g., treatment and characteristic methods) to control the biofilm formed inside the membrane systems, such as triggering bacterial dispersion and biofilm detachment [21,22]. For instance, biocides can effectively minimize biofouling in seawater reverse osmosis (SWRO) plants because they can inactivate microorganisms and induce biofilm matrix dispersion from the membrane surface [23]. Biocides can be classified into oxidizing biocides (e.g., chlorine, bromine, and chlorine dioxide) and non-oxidizing biocides (e.g., quaternary ammonium compounds and glutaraldehyde). It is well known that most oxidizing biocides have compatibility issues with polyamide membranes because free chlorine can attack the amine functional group. Chlorination and de-chlorination occasionally enhance biofouling rather than control it [24,25,26].
In contrast, non-oxidizing biocides with broad-spectrum antimicrobial effects and good compatibility with polyamide membranes, such as 2,2-dibromo-3-nitrilopropionamide (DBNPA) [27], are successfully applied in some reverse osmosis (RO) plants [28]. Ethyl lauroyl arginate (LAE), a preservative and cationic biocide, is widely used in food and medical fields, and was approved by the Food and Drug Administration (FDA) in 2005 [29]. Previous studies [30,31] revealed that the cationic liposomes (e.g., stearylamine) or biocides (e.g., quaternary ammonium) can specifically electrostatically bond with bacterial cell walls and biofilms. Indeed, the LAE is an amphiphilic molecule consisting of a positively charged head (i.e., L-arginine) and an aliphatic tail (i.e., lauric acid). For this reason, the positively charged LAE can interact with the negatively charged surface of a bacteria cell and cause disturbances in the cell membrane potential and structural changes, thus inducing the loss of cell viability [32,33]. This action mode of LAE rationally explains why the LAE has high antibacterial activity against a wide range of microorganisms (e.g., bacteria, fungi, yeast, and algae). LAE is also proven to be safe and non-toxic in mammals and the human body [34,35]. In our previous studies, LAE was proposed as an effective non-oxidizing biocide for direct dosing in the RO process and showed high inhibitory effects on biofilm formation as well as good compatibility with the membrane [36,37].
Generally, the biofilm development consists of four stages, including (1) the initial adhesion of microbials to a surface, (2) formation of microcolonies, (3) maturation of the biofilm, and (4) the dispersal or decomposition of the biofilm, which accompanies the bacteria itself to switch from a solitary planktonic lifestyle to the communal biofilm lifestyle [38]. It was recognized that the switch from motile to sessile cells is critically important for its ability to colonize on the surface. For the planktonic cells in liquid, the movement behavior of cells is controlled by the swimming motility, which is powered by rotating flagella. In contrast, the swarming and twitching play dominant roles in the cell’s movement under a solid surface or biofilm lifestyle, which are mediated by the helical flagella and type IV pili, respectively [39]. It was evidenced that the migration of bacteria can affect the phenotype and structure of biofilm [38]. Previous study also reported that the cationic peptide can effectively inhibit the biofilm by altering or simulating the swarming and twitching motilities [40].
However, previous studies lack a sufficient understanding of the mechanism of LAE in the control of attached microorganisms in biofilms [36]. Additionally, operational guidelines for LAE treatment, such as applicable concentrations, dosing times, and dosing methods, should be systematically investigated. Generally, the lab-scale RO unit has limitations and difficulties in the real-time observation of bacterial transfer properties near the membrane surface. Recently, transmission flow cells with a viewing window combined with a microscope were successfully utilized in biological and membrane fields, which can be used in in vitro assays of initial bacterial adhesion [41] and deposition [42,43] on various material surfaces. We speculate that a continuous flow cell system, featuring real-time biofilm monitoring, is critically important and useful for exploring the operational conditions of LAE treatment.
In this study, the operational guidelines for LAE (e.g., dosing methods, times, and concentrations) and the shear rate effects on biofilm detachment were systematically determined. The mechanisms underlying the ability of LAE to mitigate biofilm formation were also critically discussed. This study not only sheds light on the role of LAE concentration on biofilm detachment, but also provides an operation guide of non-oxidizing biocide for biofouling mitigation. Moreover, this study developed a novel in situ and real-time biofilm monitoring method, which can be used as a research paradigm for biofilm inhibition and detachment in the field of membrane-based water treatment and industrial cooling water systems.
2. Materials and Methods
2.1. Bacteria, Biofilm Formation, and Preparation of LAE Solution
Pseudomonas aeruginosa strain PA14 (PA14) was used as a model gram-negative bacterium for the flow cell test in this study. PA14 was cultured overnight in a shaking incubator at 250 rpm and 37 °C in AB medium (300 mM NaCl, 50 mM MgSO4, 0.2% vitamin-free casamino acids, 10 mM potassium phosphate, 1 mM L-arginine, and 1% glucose, pH 7.5). LAE powder (95%, CDI, Hwaseong, South Korea) was dissolved in deionized (DI) water. LAE solutions were filtered using a 0.22 µm syringe filter and adjusted to pH 7 using 1 M NaOH and 1 M HNO3 solutions. The LAE solutions were prepared at concentrations of 0.42, 4.2, and 42 mg/L.
2.2. Setup of Flow Cell System and Calculation of Shear Rate
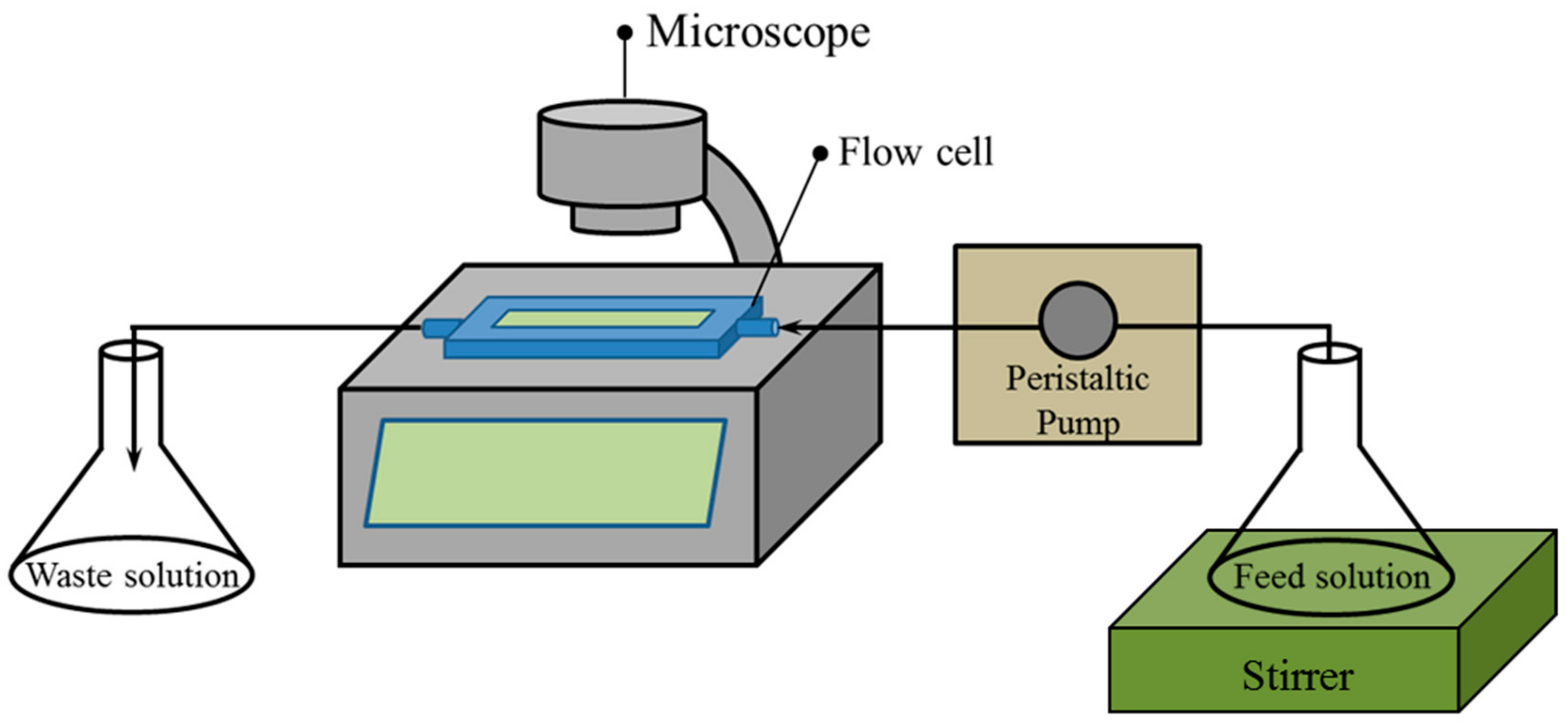
Real-time bacterial detachment was observed using a flow cell system. Briefly, the glass slides were first immersed in a Petri dish containing 20 mL of AB medium with 1 mL of PA14 suspension (optical density (OD) at 595 nm = ~1.0) and then incubated for 24 h at 37 °C to form biofilm on the glass slide surface. A flow cell reactor (FC 81, BioSurface Technologies Corporation, Bozeman, MT, USA) was adopted as the flow channel for dosing the LAE solution and phosphate-buffered saline (PBS) to observe cell detachment. The dimensions of the flow channel were 13 mm × 3 mm × 50 mm (W × H × L). The flow cell system (Figure 1) consisted of a flow cell reactor, live cell imaging microscope (JuLITM, NanoEnTek, Inc., Newton, MA, USA), peristaltic pump (Masterflex L/S, Cole-Parmer, Vernon Hills, IL, USA), magnetic stirrer, feed tank, and waste tank. Considering that the thickness of the formed biofilm in this study was much thinner than that in dynamic conditions [44,45], only three relatively low shear rates were adopted in this study to better investigate the effects of LAE concentrations on biofilm detachment. The flow rates were set at 1, 2, and 4 mL/min. The shear rate of the flow cell was calculated from the volumetric flow rate and the size of the rectangular flow channel using the following equation [46,47]:
where
is the wall shear rate,
. is the volumetric flow rate, and
and
are the height and width of the flow channel, respectively.
2.3. Image Capture and Data Processing
The impact of LAE treatment on biofilm detachment from glass slides was continuously observed using time-lapse microscopy. The JuLITM microscope was focused centrally on the surface of the flow cell channel for all time-lapse tests. The interval of image capture was set to 2 min, and the total duration for observing biofilm detachment was 2 h. Time-lapse images and movies were captured using JuLITM image view software (version 1.01). The captured images and movies were stored on a JuLITM SD card and transferred to a personal computer for data processing. The normalized biofilm area (A/A0; A, biofilm area according to time-lapse; A0, initial biofilm area) on the surface was calculated based on the pixels of the threshold images using the ImageJ program (National Institutes of Health, Bethesda, MD, USA). The mean and standard deviation of the biofilm areas were calculated using at least three independent tests.
2.4. Motility Test
The PA14 solution after 12 h of cultivation was diluted with fresh AB medium (OD at 595 nm = 0.1). The diluted bacterial suspension (5 μL) was then dropped onto the surface of a BM2 swarm agar plate (62 mM potassium, phosphate buffer [pH 7], 2 mM MgSO4, 10 μM FeSO4, 0.4% [wt/vol] glucose, 0.5% [wt/vol] casamino acids, and 0.5% [wt/vol] Bacto agar) [48], which included LAE at different concentrations (0, 0.42, 4.2, and 42 mg/L). After incubation for 32 h at room temperature, PA14 motility on the agar plate surface was determined by measuring the colony radius of the swarming zones. The swarming motility experiments were repeated three times for each sample.
2.5. Confocal Laser Scanning Microscopy (CLSM)
The cultivated PA14 solution (OD at 595 nm = 0.1) was diluted with fresh AB medium (1:20). Then, the glass slides were immersed in the AB medium and incubated for 24 h at 37 °C without agitation in order to form biofilms on the surface. The formed biofilms were treated with LAE solutions (0, 0.42, 4.2, and 42 mg/L) for 60 min. The remaining biofilms were stained using a LIVE/DEAD BacLight bacterial viability kit (Invitrogen, Carlsbad, CA, USA). After washing the unbound dye, live (green) and dead (red) cells were observed using confocal laser scanning microscopy (CLSM) (LSM700, Carl Zeiss, Jena, Germany). CLSM images were obtained with green and red fluorescent lights using a 20× objective lens (Plan-Apochromat 20×/0.8) and Z-stack mode in the Zen 2011 software (Carl Zeiss).
3. Results and Discussion
3.1. Effects of Shear Rate on Biofilm Detachment
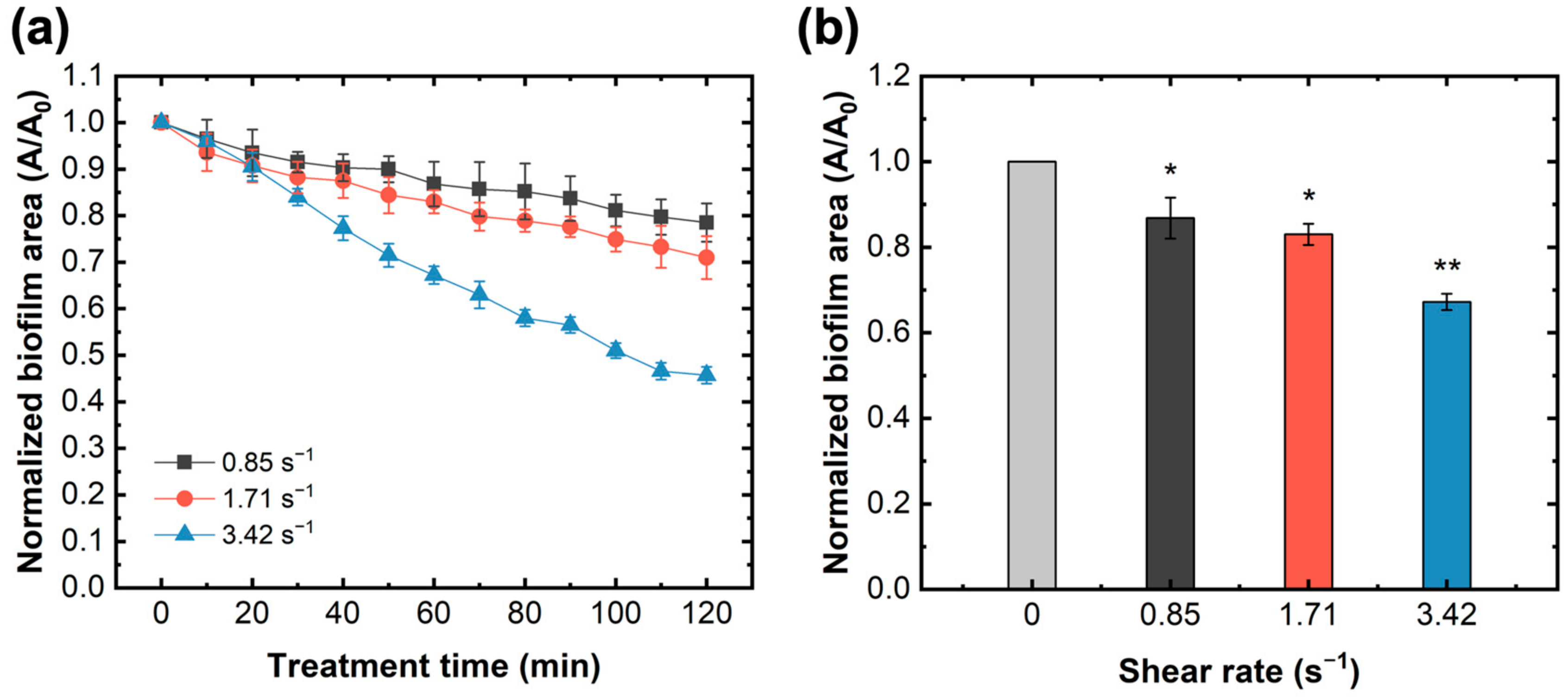
Before investigating the impacts of the LAE treatment, the shear rate inside the flow cell channel was varied by altering the flow velocity, and the resulting effects on biofilm detachment were explored. The detachment efficiency can be determined by the ratio of the biofilm area before and after LAE or PBS treatment, that is, the normalized biofilm area. Figure 2 shows the reduction trends of the biofilm area formed on the glass slide surface under different shear rates without the LAE treatment. Generally, it is shown that the normalized biofilm area rapidly decreased with the increase in shear rate (Figure 2a). When PBS flowed for over 60 min, the normalized biofilm area decreased from approximately 13.2% to 32.8% at shear rates ranging from 0.85 s−1 to 3.42 s−1 (Figure 2b). Although the formed biofilm was not directly exposed to the biocide of LAE, the physical shear stress caused by the flow velocity could still trigger biofilm dispersion to some degree. A biofilm on the membrane surface is a dynamic balance between the adhesive rate of planktonic cells, the growth rate of attached microorganisms, and the detachment rate of the biofilm [49]. Previous studies proposed that shear rate plays multiple roles in biofilm formation [50] and its structure, as well as affects membrane performance [51,52]. For instance, a high shear rate can facilitate the transfer of mass (e.g., nutrients) to the biofilm, resulting in faster biofilm development. The biofilm formed under high shear rate conditions generally possessed a flatter and more compact structure, whereas a looser and fluffy biofilm formed at a low shear rate [49,51].
In contrast, a high shear rate can easily reduce the cell attachment rate. In this study, biofilms and their structures were identically formed on the glass slide surface and then subjected to varied shear stress. Therefore, shear rate was confirmed as the only influencing factor causing sessile bacterial reduction in biofilms. A higher shear rate (3.42 s−1) could induce a greater lifting force and weaken the binding force between sessile bacteria and the surface, resulting in a high biofilm detachment rate during identical treatment times. Based on these results, the shear rate in the following tests was set to 3.42 s−1 to reduce the dose of biocide.
3.2. Effects of LAE Concentration and Dosing Time on Biofilm Detachment
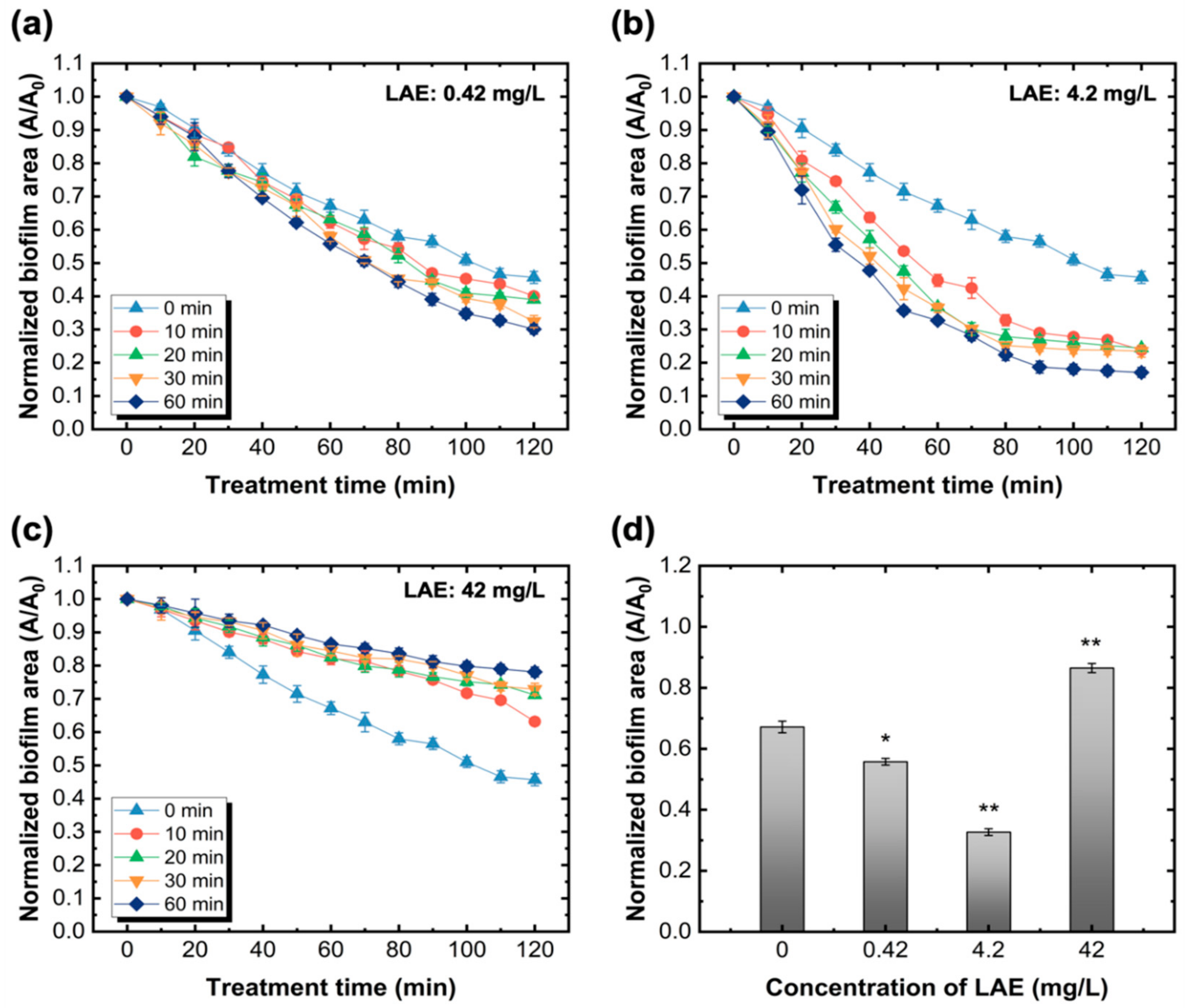
A previous study [36] revealed that the minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of LAE were both approximately less than 32 mg/L. To explore the influence of LAE concentrations on biofilm detachment, concentrations of LAE of three orders of magnitude (0.42, 4.2, and 42 mg/L) were selected and then continuously injected into the flow cell. The trends of biofilm area change were interval determined, and the results are shown in Figure 3. Generally, the biofilm treated with 0.42 mg/L and 4.2 mg/L of LAE possessed similar decreasing trends with the increase in LAE dosing time ranging from 10 to 60 min. For example, for the same treatment time of LAE (e.g., 60 min), the normalized biofilm areas were eventually reduced to 0.30 and 0.17 after dosing with 0.42 mg/L and 4.2 mg/L of LAE, respectively. However, compared with the biofilm that was only exposed to the PBS buffer, it was noted that the biofilm area increased as the LAE treatment time was prolonged when its concentration equaled 42 mg/L. Figure 3d also shows the normalized biofilm area declined with respect to the LAE concentration. The reduction of biofilms treated with 4.2 mg/L of LAE for 60 min was statistically significant (p < 0.005) compared with the control group. Additionally, Figure 3d also reveals that there is no direct correlation between the decline in biofilm and the increase in LAE concentration.
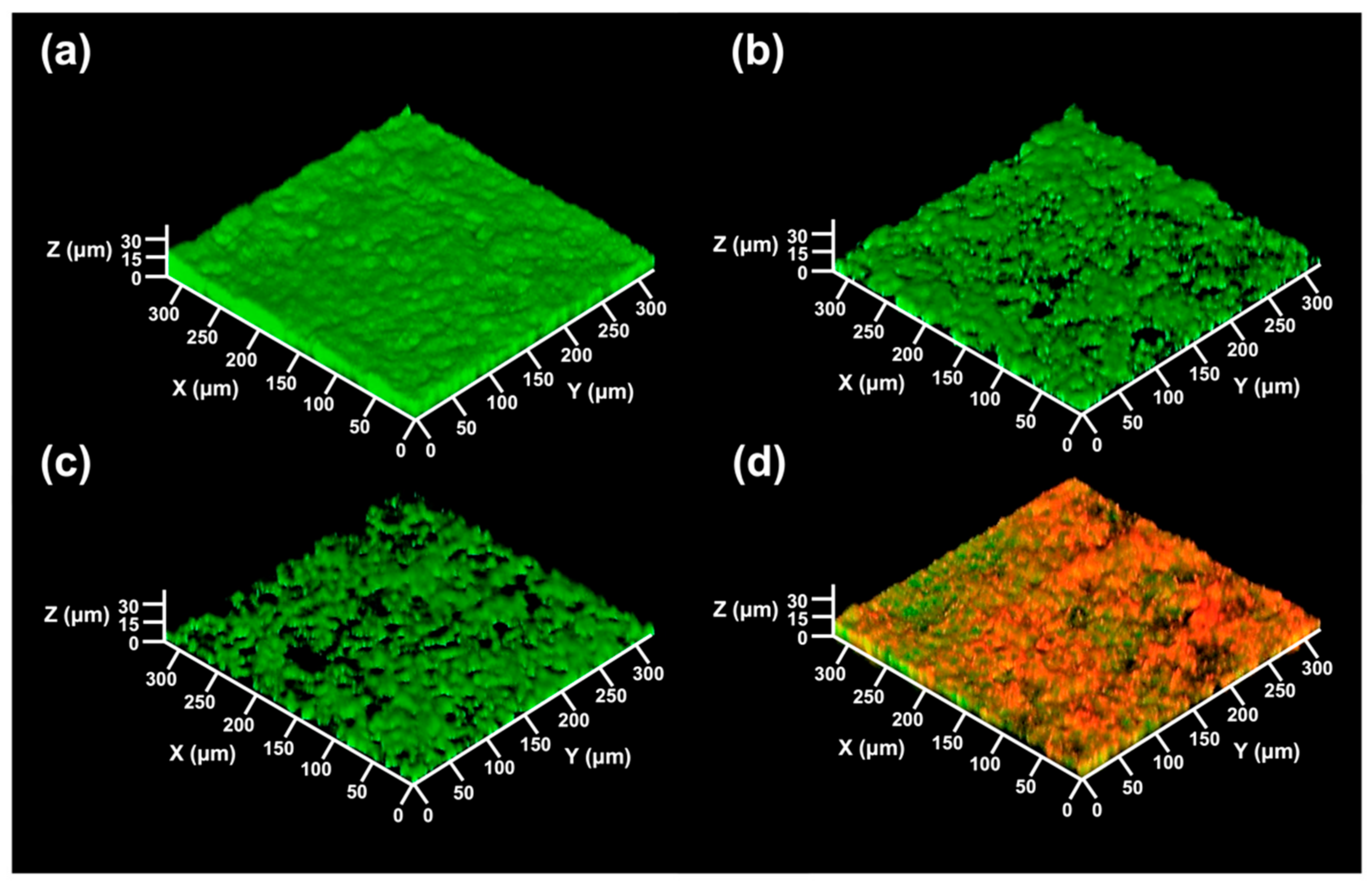
It seems that the biocidal and sub-growth inhibitory concentrations against PA14 have different effects and mechanisms of action on biofilm detachment. In these results, a low LAE concentration was able to induce detachment of sessile bacteria in the biofilm, which was more effective than treatment with high LAE concentrations greater than the MBC value (32 mg/L). The detachment of sessile bacteria by LAE treatment was associated with bacterial movement, such as twitching motility, owing to the chemical properties of LAE. A previous study [53] demonstrated that LAE against PA14 increased the twitching motility of bacteria on the surface by iron chelating activity. The rapid decrease of formed biofilms during the 60 min treatment of 0.42 mg/L and 4.2 mg/L of LAE suggests that the sub-growth concentration of LAE may enhance the cell mobility (Figure 4). In contrast, Figure 4 shows that biofilms treated with 42 mg/L of LAE had no obvious changes on the glass slide surface for 120 min. We rationally speculated that LAE treatment over biocidal concentrations (MBC > 32 mg/L of LAE with PA14) would induce the direct death of sessile bacteria in biofilms and loss of mobility, thus leaving its debris on the surface. The CLSM images directly support the above explanation, wherein the biofilm consisted almost entirely of dead cells (red color in Figure 5d) when the biofilm was treated with 42 mg/L of LAE. Additionally, the formed biofilm without treatment of LAE exhibited a dense, compact, and flat morphology. In contrast, the biofilms were fragmentary and sporadic once treated with LAE, regardless of concentrations. The thickness of the biofilm also decreased from 16.0 ± 1.3 μm of control to 11.9 ± 1.1, 8.1 ± 1.2, 12.8 ± 0.4 μm as it was treated with 0.42, 4.2 and 42 mg/L of LAE, respectively.
3.3. Effects of LAE on the Motility of Pseudomonas aeruginosa
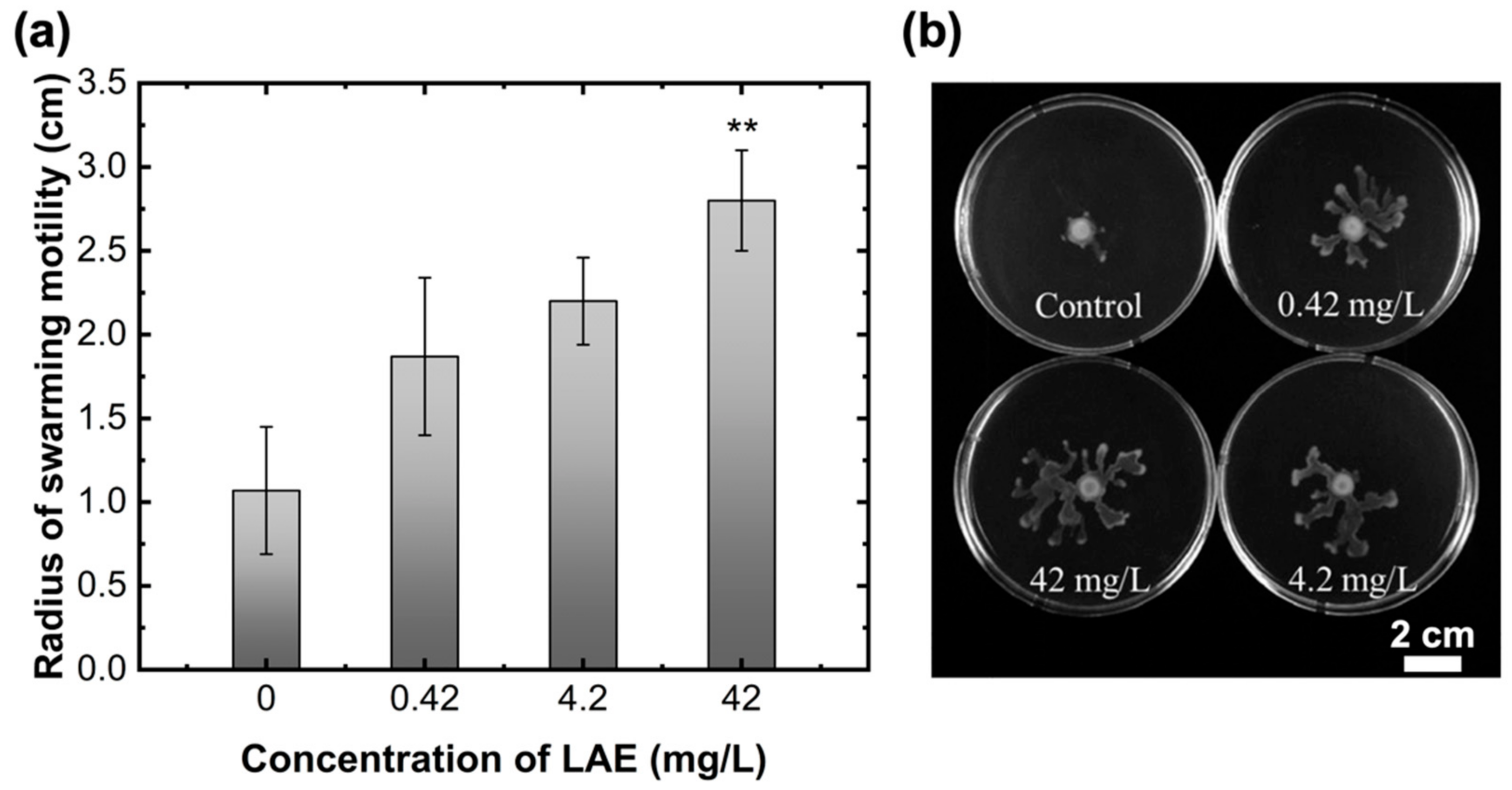
To further investigate the impact of LAE concentration on the motility of PA14, LAE-containing plate agar was used to determine the radius of swarming motility, as shown in Figure 6. Swarming motility is well known as the rapid multicellular movement of bacteria on the surface and is generated by rotating helical flagella [39]. Swarming motility was visibly correlated with LAE treatment concentration. As shown in Figure 6a, the radii of swarming motility were 1.07 ± 0.38, 1.87 ± 0.47, 2.20 ± 0.26, and 2.80 ± 0.30 cm, as the BM2 swarm agar contained 0, 0.42, 4.2, and 42 mg/L of LAE concentrations, respectively. Therefore, LAE, including BM2 swarm agar, induced an increase in swarming motility with increasing concentration. An increase in swarming motility has an antagonistic effect on biofilm formation on the surface [39,54]. Additionally, our previous study [53] demonstrated that the LAE at sub-growth inhibitory concentration can chelate irons because the LAE molecules contain two types of donor groups (i.e., carbonyl group and guanidine group) that can potentially form ferric and ferrous iron chelae complexes. Another study [55] also revealed that iron-limiting conditions can increase the twitching motility of P. aeruginosa in the presence of the iron-chelating molecules of deferoxamine. LAE treatment caused biofilm detachment on the surface due to the generation of continuous bacterial movement. LAE was assumed to act as a surfactant secretion (e.g., rhamnolipid) to reduce the surface tension, and thus enhancing the movement of multicellular bacteria and detachment of biofilms [36,56].
In fact, 42 mg/L of LAE against PA14 was biocidal in a nutrient broth medium, such as tryptic soy broth. Nevertheless, 42 mg/L of LAE in the solid phase, such as BM2 swarm agar, increased the swarming motility of PA14. This result may be attributed to the increased resistance levels against the LAE of swarming cells on the agar surface under more aerobic conditions compared with planktonic cells. Moreover, the diffusion resistance of biocides in the solid agar medium are higher than that in the broth medium [57,58]. Hence, 42 mg/L LAE in swarm agar may not influence the growth of PA14. Lai et al. [59] observed that swarm cells of five different species (P. aeruginosa, E. coli, S. marcescens, B. subtilis, and B. thailandensis) can grow at concentrations higher than their planktonic counterparts. For example, the resistance of B. subtilis swarm cells to tobramycin was 85 times higher than that of planktonic cells. Nevertheless, although a larger radius of swarming motility under a higher concentration of LAE would be formed, the bacterial cells and their formed biofilm can be directly exposed to the LAE solution, and thus are more susceptible to damage at the same concentration.
3.4. Implications and Prospects
Biofilm detachment from the membrane surface is critical for mitigating and realizing sustainable operation. Real-time and in vitro observations of biofilms are urgently needed to explore the operational conditions for biofilm detachment or removal. Although some devices (e.g., optical coherence tomography [60]) can continuously observe and analyze the dynamic variation of biofouling, their cost is relatively high. Compact and ingenious transmission flow cells combined with microscopy are easier to access and control. A modified flow cell can also mimic the operating conditions (e.g., chemical concentration, treatment time, and hydrodynamic parameters) applied in real processes [43,61].
Although the biofilm can be partially removed by physically elevating the crossflow velocity, the bacterial cells protected by extracellular polymeric substances (EPS) are difficult to address [10,62]. Generally, the biofilm formation on the substrate includes two stages: bacterial adhesion and bacterial colonization on the substrate [63,64]. Bacterial adhesion on the substrate can be mediated by the short-range interactions, such as hydrophobic interactions, hydrogen bonding, electrostatic interactions, as well as van der Waals interactions. At the same time, the extracellular polymeric substances (EPS) are secreted by bacterial cells themselves, which act as a “bio-glue” that can encase the cells from external distractions and enhance the bonding force to the substrate, since it has many functional groups [63,65].
In this study, we speculated that the LAE act as a cationic surfactant, and biocide may play two important roles in the biofilm dispersal, including altering the binding forces between the biofilm and substrate, as well as the bacterial swimming mobility. An appropriate dose of chemicals can enhance the treatment effectiveness because the microorganism can respond differently to the biocide. The bactericidal concentration of LAE can directly damage bacteria, which overshadows its role in enhancing cell movement. Instead, the sub-growth inhibitory concentrations of LAE can effectively increase bacterial motility and the biofilm detachment rate. Based on this consideration, the dose of biocides should be carefully optimized during different stages of real membrane operations. For instance, a high concentration of biocide is preferred to kill microorganisms in the feed solution at the initial stage. However, once the biofilm develops after long-term operation, it should be treated with an appropriate concentration of biocide to increase cell motility and trigger biofilm dispersion rather than kill the microorganisms. Additionally, the biocide adsorption on the biofilm surface, diffusion-reaction inhibition, and concentration polarization phenomena can affect the temporal and spatial distribution of biocides in the biofilm matrix, thus influencing the treatment efficacy. Therefore, future studies should consider and perform a more comprehensive evaluation based on the realistic properties of the feed solution (e.g., nutrient concentration and type), membrane surface (e.g., hydrophobicity and surface charge), and hydrodynamic conditions (e.g., shear rates) for a wide range of real applications.
4. Conclusions
In this study, LAE was selected as a model non-oxidizing biocide with high antibacterial effectiveness to explore the effects of shear rate, chemical concentration, and treatment time on biofilm detachment. The biofilm detachment was enhanced with the increase in shear rate owing to the higher shear force, which easily lifted the biofilm from the surface. LAE concentration played two opposite roles in biofilm detachment. Under a high concentration of LAE (i.e., higher than the MIC or MBC), the sessile bacteria in the biofilm were susceptible to damage, and their debris remained on the surface; thus, only a part of the biofilm could eventually be detached. In contrast, LAE treatment at sub-growth inhibitory concentrations increased bacterial movement due to the increase in swarming motility on the surface, resulting in enhanced biofilm disintegration. We also propose that future studies should pay more attention to real conditions in order to achieve a more comprehensive evaluation.
Author Contributions
Conceptualization, T.-S.K. and P.-F.S.; methodology, T.-S.K. and S.-Y.H.; validation, T.-S.K. and P.-F.S.; investigation, T.-S.K.; data curation, P.-F.S. and Y.-S.J.; writing—original draft preparation, P.-F.S. and T.-S.K.; writing—review and editing, P.-F.S.; visualization, P.-F.S. and T.-S.K.; supervision, H.-D.P.; funding acquisition, H.-D.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2020R1A6A1A03045059).
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Logan, B.E.; Elimelech, M. Membrane-Based Processes for Sustainable Power Generation Using Water. Nature 2012, 488, 313–319. [Google Scholar] [CrossRef]
- Elimelech, M.; Phillip, W.A. The Future of Seawater Desalination: Energy, Technology, and the Environment. Science 2011, 333, 712–717. [Google Scholar] [CrossRef]
- Tang, C.Y.; Yang, Z.; Guo, H.; Wen, J.J.; Nghiem, L.D.; Cornelissen, E. Potable Water Reuse through Advanced Membrane Technology. Environ. Sci. Technol. 2018, 52, 10215–10223. [Google Scholar] [CrossRef] [Green Version]
- Tijing, L.D.; Woo, Y.C.; Choi, J.-S.; Lee, S.; Kim, S.-H.; Shon, H.K. Fouling and Its Control in Membrane Distillation—A Review. J. Membr. Sci. 2015, 475, 215–244. [Google Scholar] [CrossRef]
- Meng, S.; Meng, X.; Fan, W.; Liang, D.; Wang, L.; Zhang, W.; Liu, Y. The Role of Transparent Exopolymer Particles (TEP) in Membrane Fouling: A Critical Review. Water Res. 2020, 181, 115930. [Google Scholar] [CrossRef]
- Bogler, A.; Lin, S.; Bar-Zeev, E. Biofouling of Membrane Distillation, Forward Osmosis and Pressure Retarded Osmosis: Principles, Impacts and Future Directions. J. Membr. Sci. 2017, 542, 378–398. [Google Scholar] [CrossRef]
- Kucera, J. Biofouling of Polyamide Membranes: Fouling Mechanisms, Current Mitigation and Cleaning Strategies, and Future Prospects. Membranes 2019, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Firouzjaei, M.D.; Seyedpour, S.F.; Aktij, S.A.; Giagnorio, M.; Bazrafshan, N.; Mollahosseini, A.; Samadi, F.; Ahmadalipour, S.; Firouzjaei, F.D.; Esfahani, M.R.; et al. Recent Advances in Functionalized Polymer Membranes for Biofouling Control and Mitigation in Forward Osmosis. J. Membr. Sci. 2020, 596, 117604. [Google Scholar] [CrossRef]
- Sun, P.-F.; Jang, Y.; Ham, S.-Y.; Ryoo, H.; Park, H.-D. Effects of Reverse Solute Diffusion on Membrane Biofouling in Pressure-Retarded Osmosis Processes. Desalination 2021, 512, 115145. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The Biofilm Matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Lin, H.; Zhang, M.; Wang, F.; Meng, F.; Liao, B.Q.; Hong, H.; Chen, J.; Gao, W. A Critical Review of Extracellular Polymeric Substances (EPSs) in Membrane Bioreactors: Characteristics, Roles in Membrane Fouling and Control Strategies. J. Membr. Sci. 2014, 460, 110–125. [Google Scholar] [CrossRef]
- Sun, P.-F.; Kim, T.-S.; Kim, H.-S.; Ham, S.-Y.; Jang, Y.; Park, Y.-G.; Tang, C.Y.; Park, H.-D. Improved Anti-biofouling Performance of Pressure Retarded Osmosis (PRO) by Dosing with Chlorhexidine Gluconate. Desalination 2020, 481, 114376. [Google Scholar] [CrossRef]
- Herzberg, M.; Kang, S.; Elimelech, M. Role of Extracellular Polymeric Substances (EPS) in Biofouling of Reverse Osmosis Membranes. Environ. Sci. Technol. 2009, 43, 4393–4398. [Google Scholar] [CrossRef]
- Herzberg, M.; Elimelech, M. Biofouling of Reverse Osmosis Membranes: Role of Biofilm-Enhanced Osmotic Pressure. J. Membr. Sci. 2007, 295, 11–20. [Google Scholar] [CrossRef]
- Gao, Y.; Zhao, S.; Qiao, Z.; Zhou, Y.; Song, B.; Wang, Z.; Wang, J. Reverse Osmosis Membranes with Guanidine and Amine Enriched Surface for Biofouling and Organic Fouling Control. Desalination 2018, 430, 74–85. [Google Scholar] [CrossRef]
- Bucs, S.S.; Farhat, N.; Kruithof, J.C.; Picioreanu, C.; van Loosdrecht, M.C.M.; Vrouwenvelder, J.S. Review on Strategies for Biofouling Mitigation in Spiral Wound Membrane Systems. Desalination 2018, 434, 189–197. [Google Scholar] [CrossRef]
- Park, K.-H.; Sun, P.-F.; Kang, E.H.; Han, G.D.; Kim, B.J.; Jang, Y.; Lee, S.-H.; Shim, J.H.; Park, H.-D. Photocatalytic Anti-biofouling Performance of Nanoporous Ceramic Membranes Treated by Atomic Layer Deposited ZnO. Sep. Purif. Technol. 2021, 272, 118935. [Google Scholar] [CrossRef]
- Liu, C.; Faria, A.F.; Ma, J.; Elimelech, M. Mitigation of Biofilm Development on Thin-Film Composite Membranes Functionalized with Zwitterionic Polymers and Silver Nanoparticles. Environ. Sci. Technol. 2017, 51, 182–191. [Google Scholar] [CrossRef]
- Khan, R.; Wang, H.; Li, Y.; Yu, S.; Khan, M.K.; Xiao, K.; Huang, X. Surface Grafting of Reverse Osmosis Membrane with Chlorhexidine Using Biopolymer Alginate Dialdehyde as a Facile Green Platform for In Situ Biofouling Control. ACS Appl. Mater. Interfaces 2020, 12, 37515–37526. [Google Scholar] [CrossRef]
- Yang, Z.; Wu, Y.; Wang, J.; Cao, B.; Tang, C.Y. In Situ Reduction of Silver by Polydopamine: A Novel Antimicrobial Modification of a Thin-Film Composite Polyamide Membrane. Environ. Sci. Technol. 2016, 50, 9543–9550. [Google Scholar] [CrossRef]
- Oh, H.-S.; Constancias, F.; Ramasamy, C.; Tang, P.Y.P.; Yee, M.O.; Fane, A.G.; McDougald, D.; Rice, S.A. Biofouling Control in Reverse Osmosis by Nitric Oxide Treatment and Its Impact on the Bacterial Community. J. Membr. Sci. 2018, 550, 313–321. [Google Scholar] [CrossRef]
- Bar-Zeev, E.; Elimelech, M. Reverse Osmosis Biofilm Dispersal by Osmotic Back-Flushing: Cleaning via Substratum Perforation. Environ. Sci. Technol. Lett. 2014, 1, 162–166. [Google Scholar] [CrossRef]
- Kim, D.; Jung, S.; Sohn, J.; Kim, H.; Lee, S. Biocide Application for Controlling Biofouling of SWRO Membranes—An Overview. Desalination 2009, 238, 43–52. [Google Scholar] [CrossRef]
- Gohil, J.M.; Suresh, A.K. Chlorine Attack on Reverse Osmosis Membranes: Mechanisms and Mitigation Strategies. J. Membr. Sci. 2017, 541, 108–126. [Google Scholar] [CrossRef]
- Al-Abri, M.; Al-Ghafri, B.; Bora, T.; Dobretsov, S.; Dutta, J.; Castelletto, S.; Rosa, L.; Boretti, A. Chlorination Disadvantages and Alternative Routes for Biofouling Control in Reverse Osmosis Desalination. NPJ Clean Water 2019, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.T.; Hong, P.Y.; Nada, N.; Croue, J.P. Does Chlorination of Seawater Reverse Osmosis Membranes Control Biofouling? Water Res. 2015, 78, 84–97. [Google Scholar] [CrossRef] [Green Version]
- Bertheas, U.; Majamaa, K.; Arzu, A.; Pahnke, R. Use of DBNPA to Control Biofouling in RO Systems. Desalin. Water Treat. 2009, 3, 175–178. [Google Scholar] [CrossRef] [Green Version]
- Da-Silva-Correa, L.H.; Smith, H.; Thibodeau, M.C.; Welsh, B.; Buckley, H.L. The Application of Non-oxidizing Biocides to Prevent Biofouling in Reverse Osmosis Polyamide Membrane Systems: A Review. J. Water Supply Res. Technol. Aqua 2022, 71, 261–292. [Google Scholar] [CrossRef]
- Fernández, C.E.; Aspiras, M.; Dodds, M.W.; González-Cabezas, C.; Rickard, A.H. Combinatorial Effect of Magnolia Bark Extract and Ethyl Lauroyl Arginate against Multi-species Oral Biofilms: Food Additives with the Potential to Prevent Biofilm-Related Oral Diseases. J. Funct. Foods. 2018, 47, 48–55. [Google Scholar] [CrossRef]
- Scriboni, A.B.; Couto, V.M.; Ribeiro, L.N.d.M.; Freires, I.A.; Groppo, F.C.; de Paula, E.; Franz-Montan, M.; Cogo-Müller, K. Fusogenic Liposomes Increase the Antimicrobial Activity of Vancomycin Against Staphylococcus aureus Biofilm. Front Pharmacol. 2019, 10, 1401. [Google Scholar] [CrossRef] [PubMed]
- Simões, M.; Pereira, M.O.; Vieira, M.J. Action of a cationic surfactant on the activity and removal of bacterial biofilms formed under different flow regimes. Water Res. 2005, 39, 478–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, E.; Seguer, J.; Rocabayera, X.; Manresa, A. Cellular effects of monohydrochloride of l-arginine, Nα-lauroyl ethylester (LAE) on exposure to Salmonella typhimurium and Staphylococcus aureus. J. Appl. Microbiol. 2004, 96, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Manso, S.; Wrona, M.; Salafranca, J.; Nerín, C.; Alfonso, M.J.; Caballero, M.Á. Evaluation of New Antimicrobial Materials Incorporating Ethyl Lauroyl Arginate or Silver into Different Matrices, and Their Safety in Use as Potential Packaging. Polymers 2021, 13, 355. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, D.R.; Rocabayera, X.; Ruckman, S.; Segret, R.; Shaw, D. Metabolism and Pharmacokinetics of Ethyl N(Alpha)-Lauroyl-L-Arginate Hydrochloride in Human Volunteers. Food Chem. Toxicol. 2009, 47, 2711–2715. [Google Scholar] [CrossRef] [PubMed]
- Ruckman, S.A.; Rocabayera, X.; Borzelleca, J.F.; Sandusky, C.B. Toxicological and Metabolic Investigations of the Safety of N-α-Lauroyl-l-arginine Ethyl Ester Monohydrochloride (LAE). Food Chem. Toxicol. 2004, 42, 245–259. [Google Scholar] [CrossRef]
- Kim, T.-S.; Park, H.-D. Lauroyl Arginate Ethyl: An Effective Antibiofouling Agent Applicable for Reverse Osmosis Processes Producing Potable Water. J. Membr. Sci. 2016, 507, 24–33. [Google Scholar] [CrossRef]
- Kim, T.-S.; Antoinette, M.; Park, H.-D. Combination of Lauroyl Arginate Ethyl and Nisin for Biofouling Control in Reverse Osmosis Processes. Desalination 2018, 428, 12–20. [Google Scholar] [CrossRef]
- Tolker-Nielsen, T.; Ghannoum, M.; Parsek, M.; Whiteley, M.; Mukherjee, P. Biofilm Development. Microbiol Spectr. 2015, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Kearns, D.B. A Field Guide to Bacterial Swarming Motility. Nat. Rev. Microbiol. 2010, 8, 634–644. [Google Scholar] [CrossRef] [Green Version]
- Fuente-Núñez, C.d.l.; Korolik, V.; Bains, M.; Nguyen, U.; Breidenstein, E.B.M.; Horsman, S.; Lewenza, S.; Burrows, L.; Hancock, R.E.W. Inhibition of Bacterial Biofilm Formation and Swarming Motility by a Small Synthetic Cationic Peptide. Antimicrob. Agents Chemother. 2012, 56, 2696–2704. [Google Scholar] [CrossRef] [Green Version]
- Semião, A.J.C.; Habimana, O.; Cao, H.; Heffernan, R.; Safari, A.; Casey, E. The Importance of Laboratory Water Quality for Studying Initial Bacterial Adhesion during NF Filtration Processes. Water Res. 2013, 47, 2909–2920. [Google Scholar] [CrossRef] [Green Version]
- Sweity, A.; Oren, Y.; Ronen, Z.; Herzberg, M. The Influence of Antiscalants on Biofouling of RO Membranes in Seawater Desalination. Water Res. 2013, 47, 3389–3398. [Google Scholar] [CrossRef] [PubMed]
- Ziemba, C.; Khavkin, M.; Priftis, D.; Acar, H.; Mao, J.; Benami, M.; Gottlieb, M.; Tirrell, M.; Kaufman, Y.; Herzberg, M. Antifouling Properties of a Self-Assembling Glutamic Acid-Lysine Zwitterionic Polymer Surface Coating. Langmuir 2019, 35, 1699–1713. [Google Scholar] [CrossRef] [PubMed]
- Kwan, S.E.; Bar-Zeev, E.; Elimelech, M. Biofouling in Forward Osmosis and Reverse Osmosis: Measurements and Mechanisms. J. Membr. Sci. 2015, 493, 703–708. [Google Scholar] [CrossRef]
- Farhat, N.M.; Vrouwenvelder, J.S.; Van Loosdrecht, M.C.M.; Bucs, S.S.; Staal, M. Effect of Water Temperature on Biofouling Development in Reverse Osmosis Membrane Systems. Water Res. 2016, 103, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Hoek, E.M.V.; Elimelech, M. Cake-Enhanced Concentration Polarization: A New Fouling Mechanism for Salt-Rejecting Membranes. Environ. Sci. Technol. 2003, 37, 5581–5588. [Google Scholar] [CrossRef]
- Nejadnik, M.R.; van der Mei, H.C.; Busscher, H.J.; Norde, W. Determination of the Shear Force at the Balance between Bacterial Attachment and Detachment in Weak-Adherence Systems, Using a Flow Displacement Chamber. Appl. Environ. Microbiol. 2008, 74, 916–919. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.-C. Reverse Osmosis Membrane Biofouling. Exp. Therm. Fluid Sci. 1997, 14, 382–391. [Google Scholar] [CrossRef]
- Ying, W.; Gitis, V.; Lee, J.; Herzberg, M. Effects of Shear Rate on Biofouling of Reverse Osmosis Membrane during Tertiary Wastewater Desalination. J. Membr. Sci. 2013, 427, 390–398. [Google Scholar] [CrossRef]
- Paramonova, E.; Kalmykowa, O.J.; van der Mei, H.C.; Busscher, H.J.; Sharma, P.K. Impact of Hydrodynamics on Oral Biofilm Strength. J. Dent. Res. 2009, 88, 922–926. [Google Scholar] [CrossRef]
- Radu, A.I.; Vrouwenvelder, J.S.; van Loosdrecht, M.C.M.; Picioreanu, C. Effect of Flow Velocity, Substrate Concentration and Hydraulic Cleaning on Biofouling of Reverse Osmosis Feed Channels. Chem. Eng. J. 2012, 188, 30–39. [Google Scholar] [CrossRef]
- Al Ashhab, A.; Gillor, O.; Herzberg, M. Biofouling of Reverse-Osmosis Membranes under Different Shear Rates during Tertiary Wastewater Desalination: Microbial Community Composition. Water Res. 2014, 67, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.S.; Ham, S.Y.; Park, B.B.; Byun, Y.; Park, H.D. Lauroyl Arginate Ethyl Blocks the Iron Signals Necessary for Pseudomonas aeruginosa Biofilm Development. Front. Microbiol. 2017, 8, 970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topa, S.H.; Subramoni, S.; Palombo, E.A.; Kingshott, P.; Rice, S.A.; Blackall, L.L. Cinnamaldehyde Disrupts Biofilm Formation and Swarming Motility of Pseudomonas aeruginosa. Microbiology 2018, 164, 1087–1097. [Google Scholar] [CrossRef]
- Singh, P.K.; Parsek, M.R.; Greenberg, E.P.; Welsh, M.J. A Component of Innate Immunity Prevents Bacterial Biofilm Development. Nature 2002, 417, 552–555. [Google Scholar] [CrossRef]
- Boles, B.R.; Thoendel, M.; Singh, P.K. Rhamnolipids Mediate Detachment of Pseudomonas aeruginosa from Biofilms. Mol. Microbiol. 2005, 57, 1210–1223. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, F.; Han, F.; Prinyawiwatkul, W.; No, H.K.; Ge, B. Evaluation of Diffusion and Dilution Methods to Determine the Antimicrobial Activity of Water-Soluble Chitosan Derivatives. J. Appl. Microbiol. 2013, 114, 956–963. [Google Scholar] [CrossRef] [Green Version]
- Klančnik, A.; Piskernik, S.; Jeršek, B.; Možina, S.S. Evaluation of Diffusion and Dilution Methods to Determine the Antibacterial Activity of Plant Extracts. J. Microbiol. Methods 2010, 81, 121–126. [Google Scholar] [CrossRef]
- Lai, S.; Tremblay, J.; Déziel, E. Swarming Motility: A Multicellular Behaviour Conferring Antimicrobial Resistance. Environ. Microbiol. 2009, 11, 126–136. [Google Scholar] [CrossRef]
- Wagner, M.; Horn, H. Optical Coherence Tomography in Biofilm Research: A Comprehensive Review. Biotechnol. Bioeng. 2017, 114, 1386–1402. [Google Scholar] [CrossRef]
- Teodósio, J.S.; Simões, M.; Melo, L.F.; Mergulhão, F.J. Flow Cell Hydrodynamics and Their Effects on E. coli Biofilm Formation under Different Nutrient Conditions and Turbulent Flow. Biofouling 2011, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, D.; Ziemba, C.; Herzberg, M. Revisiting Interrelated Effects of Extracellular Polysaccharides during Biofouling of Reverse Osmosis Membranes: Viscoelastic Properties and Biofilm Enhanced Osmotic Pressure. J. Membr. Sci. 2017, 523, 394–401. [Google Scholar] [CrossRef]
- Flemming, H.C.; Schaule, G.; Griebe, T.; Schmitt, J.; Tamachkiarowa, A. Biofouling—The Achilles Heel of Membrane Processes. Desalination 1997, 113, 215–225. [Google Scholar] [CrossRef]
- Pang, C.M.; Hong, P.; Guo, H.; Liu, W.-T. Biofilm Formation Characteristics of Bacterial Isolates Retrieved from a Reverse Osmosis Membrane. Environ. Sci. Technol. 2005, 39, 7541–7550. [Google Scholar] [CrossRef]
- Habimana, O.; Semião, A.J.C.; Casey, E. The Role of Cell-Surface Interactions in Bacterial Initial Adhesion and Consequent Biofilm Formation on Nanofiltration/Reverse Osmosis Membranes. J. Membr. Sci. 2014, 454, 82–96. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic diagram of the flow cell system for in vitro and real-time observation of biofilm detachment.
Figure 1.
Schematic diagram of the flow cell system for in vitro and real-time observation of biofilm detachment.

Figure 2.
Normalized biofilm area formed on glass slide surface under different shear rates. (a) Variations of normalized biofilm area depending on different shear rates without LAE treatment and with PBS buffer for 120 min. (b) Reduction of biofilm under varied shear rates after a 60 min flow of PBS buffer. Error bars indicate the standard deviations of three independent measurements. * p < 0.05 versus control, ** p < 0.005 versus control.
Figure 2.
Normalized biofilm area formed on glass slide surface under different shear rates. (a) Variations of normalized biofilm area depending on different shear rates without LAE treatment and with PBS buffer for 120 min. (b) Reduction of biofilm under varied shear rates after a 60 min flow of PBS buffer. Error bars indicate the standard deviations of three independent measurements. * p < 0.05 versus control, ** p < 0.005 versus control.

Figure 3.
Detachment of biofilm formed on the glass slide surface under different LAE concentration and dosing time. (a–c) are the effects of LAE dosing time (0, 10, 20, 30, and 60 min) on the biofilm area reduction under different LAE concentrations of 0.42 mg/L, 4.2 mg/L, and 42 mg/L, respectively. (d) Changes of normalized biofilm area treated with different LAE concentrations for 60 min. Other experimental conditions: the biofilm was continuously flowed with PBS buffer after treating with LAE until the total treatment time reached 120 min. All shear rates were set to 3.42 s−1. Error bars indicate the standard deviations of three independent measurements. * p < 0.05 versus control, ** p < 0.005 versus control.
Figure 3.
Detachment of biofilm formed on the glass slide surface under different LAE concentration and dosing time. (a–c) are the effects of LAE dosing time (0, 10, 20, 30, and 60 min) on the biofilm area reduction under different LAE concentrations of 0.42 mg/L, 4.2 mg/L, and 42 mg/L, respectively. (d) Changes of normalized biofilm area treated with different LAE concentrations for 60 min. Other experimental conditions: the biofilm was continuously flowed with PBS buffer after treating with LAE until the total treatment time reached 120 min. All shear rates were set to 3.42 s−1. Error bars indicate the standard deviations of three independent measurements. * p < 0.05 versus control, ** p < 0.005 versus control.

Figure 4.
Microscopy images of biofilm formed on the glass slide surface treated with different LAE concentrations and time. Other experimental conditions: all the LAE treatment times were identical (60 min) and followed by PBS dosing (60 min). The shear rates in all tests were equal to 3.42 s−1.
Figure 4.
Microscopy images of biofilm formed on the glass slide surface treated with different LAE concentrations and time. Other experimental conditions: all the LAE treatment times were identical (60 min) and followed by PBS dosing (60 min). The shear rates in all tests were equal to 3.42 s−1.

Figure 5.
Presented are 3D CLSM images of the biofilm treated with varied concentrations of LAE. (a) The biofilm formed on the glass slide without LAE treatment. (b–d) The formed biofilms were treated with 0.42, 4.2, and 42 mg/L of LAE for 60 min, respectively. The green and red colors refer to the live and dead PA14 cells, respectively.
Figure 5.
Presented are 3D CLSM images of the biofilm treated with varied concentrations of LAE. (a) The biofilm formed on the glass slide without LAE treatment. (b–d) The formed biofilms were treated with 0.42, 4.2, and 42 mg/L of LAE for 60 min, respectively. The green and red colors refer to the live and dead PA14 cells, respectively.

Figure 6.
The radius of the swarming motility of P. aeruginosa with LAE treatment. (a) The radius of swarming motility as a function of different concentrations of LAE included in the BM2 swarm agar. (b) Colony patterns formed by swarming motility with the treatment of different concentrations of LAE. Error bars indicate standard deviations of three independent measurements. ** p < 0.005 versus control.
Figure 6.
The radius of the swarming motility of P. aeruginosa with LAE treatment. (a) The radius of swarming motility as a function of different concentrations of LAE included in the BM2 swarm agar. (b) Colony patterns formed by swarming motility with the treatment of different concentrations of LAE. Error bars indicate standard deviations of three independent measurements. ** p < 0.005 versus control.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sun, P.-F.; Kim, T.-S.; Ham, S.-Y.; Jang, Y.-S.; Park, H.-D. Effects of Ethyl Lauroyl Arginate (LAE) on Biofilm Detachment: Shear Rate, Concentration, and Dosing Time. Water 2022, 14, 2158. https://doi.org/10.3390/w14142158
AMA Style
Sun P-F, Kim T-S, Ham S-Y, Jang Y-S, Park H-D. Effects of Ethyl Lauroyl Arginate (LAE) on Biofilm Detachment: Shear Rate, Concentration, and Dosing Time. Water. 2022; 14(14):2158. https://doi.org/10.3390/w14142158
Chicago/Turabian StyleSun, Peng-Fei, Taek-Seung Kim, So-Young Ham, Yong-Sun Jang, and Hee-Deung Park. 2022. "Effects of Ethyl Lauroyl Arginate (LAE) on Biofilm Detachment: Shear Rate, Concentration, and Dosing Time" Water 14, no. 14: 2158. https://doi.org/10.3390/w14142158
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.