
Structural Characteristics of Periphytic Algal Community and Its Relationship with Environmental Factors in the Taiyuan Region of the Fenhe River


Abstract
:1. Introduction
2. Materials and Methods
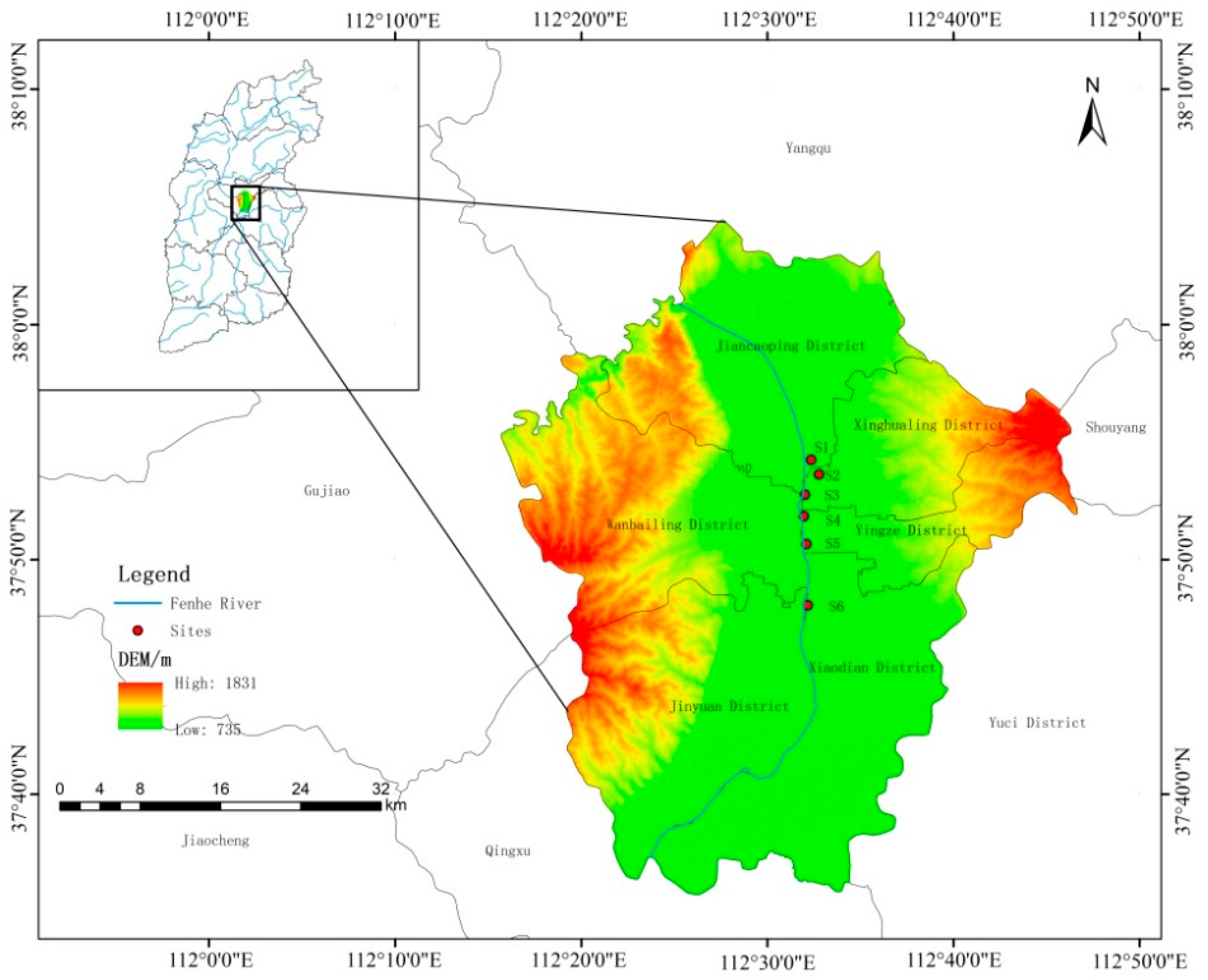
2.1. Study Area
2.2. Sample Collection
2.3. Determination of Environmental Parameters
2.4. Data Analysis
- Shannon–Wiener diversity index (H’):
- 2.
- Pielou evenness index (J):
- 3.
- Margalef richness index (d):
- 4.
- Dominance index formula (Y):
3. Results
3.1. Physical and Chemical Conditions
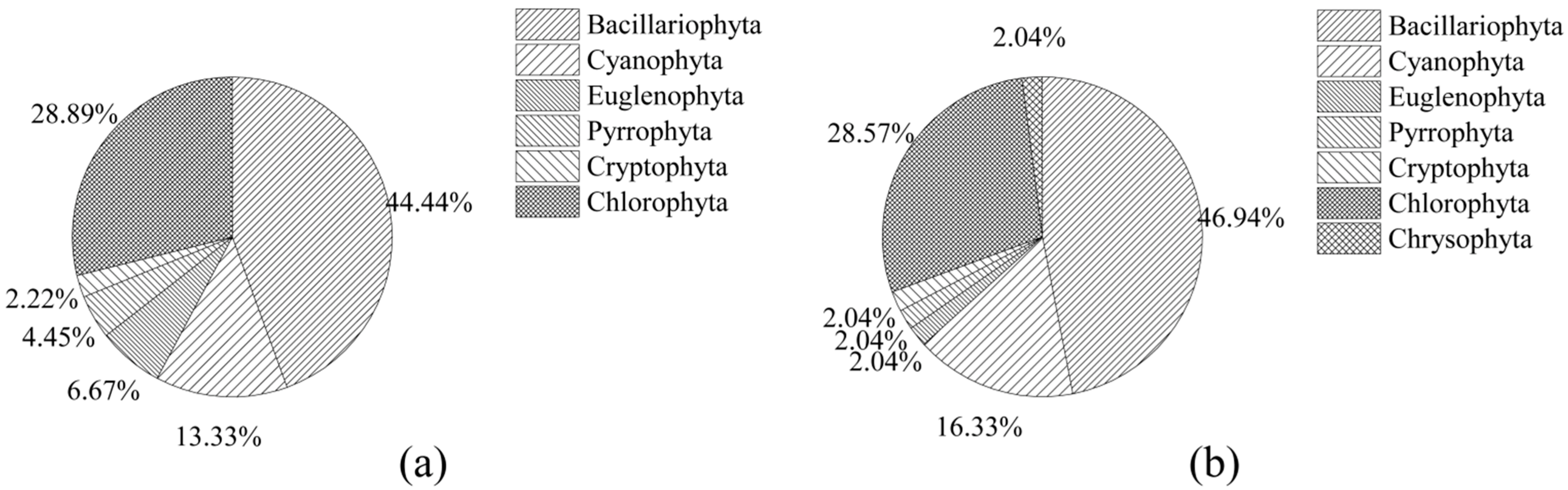
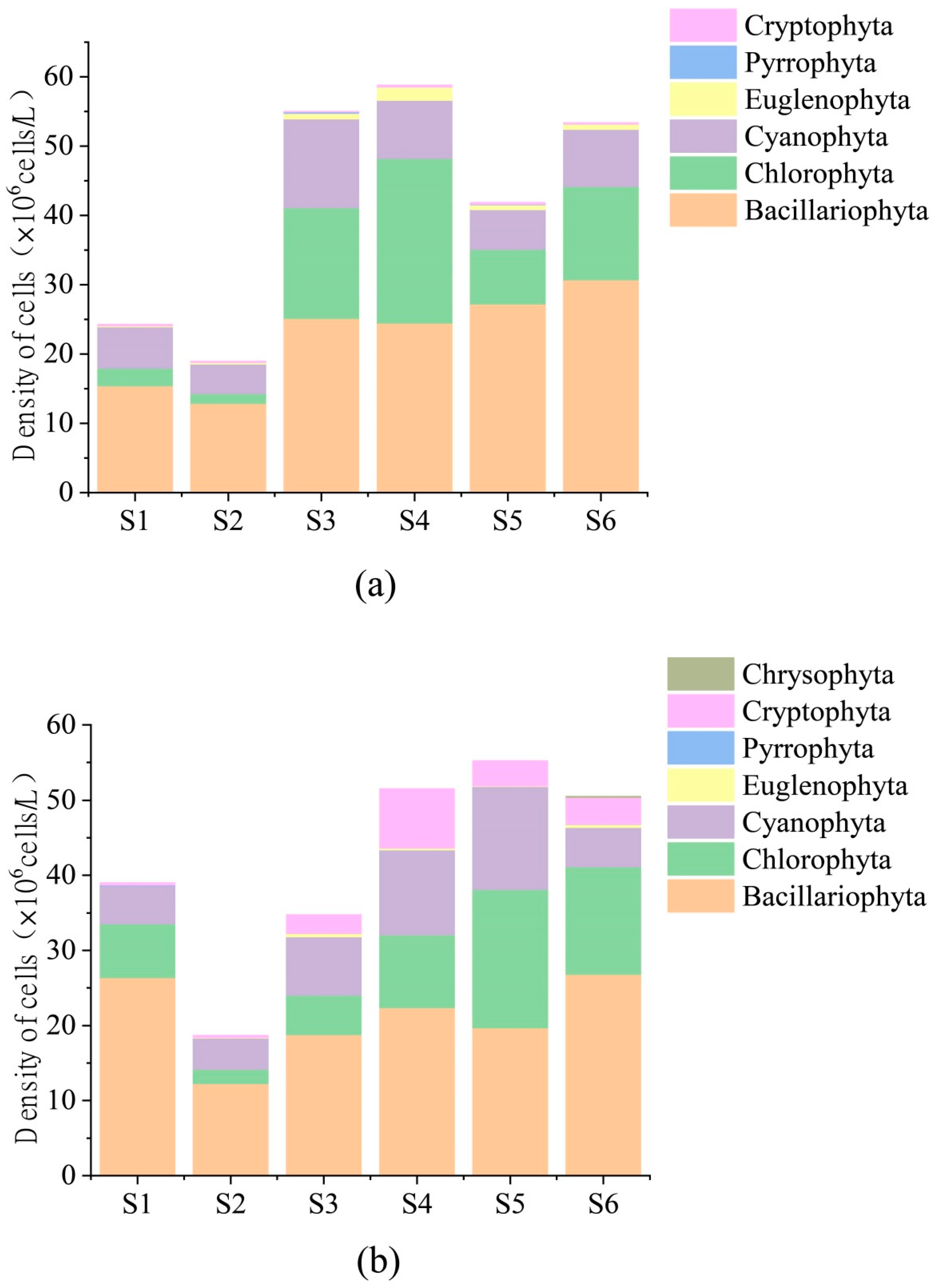
3.2. Characteristics of Periphytic Algal Community Structure
3.3. Diversity of Periphytic Algal Community and Water Quality Assessment
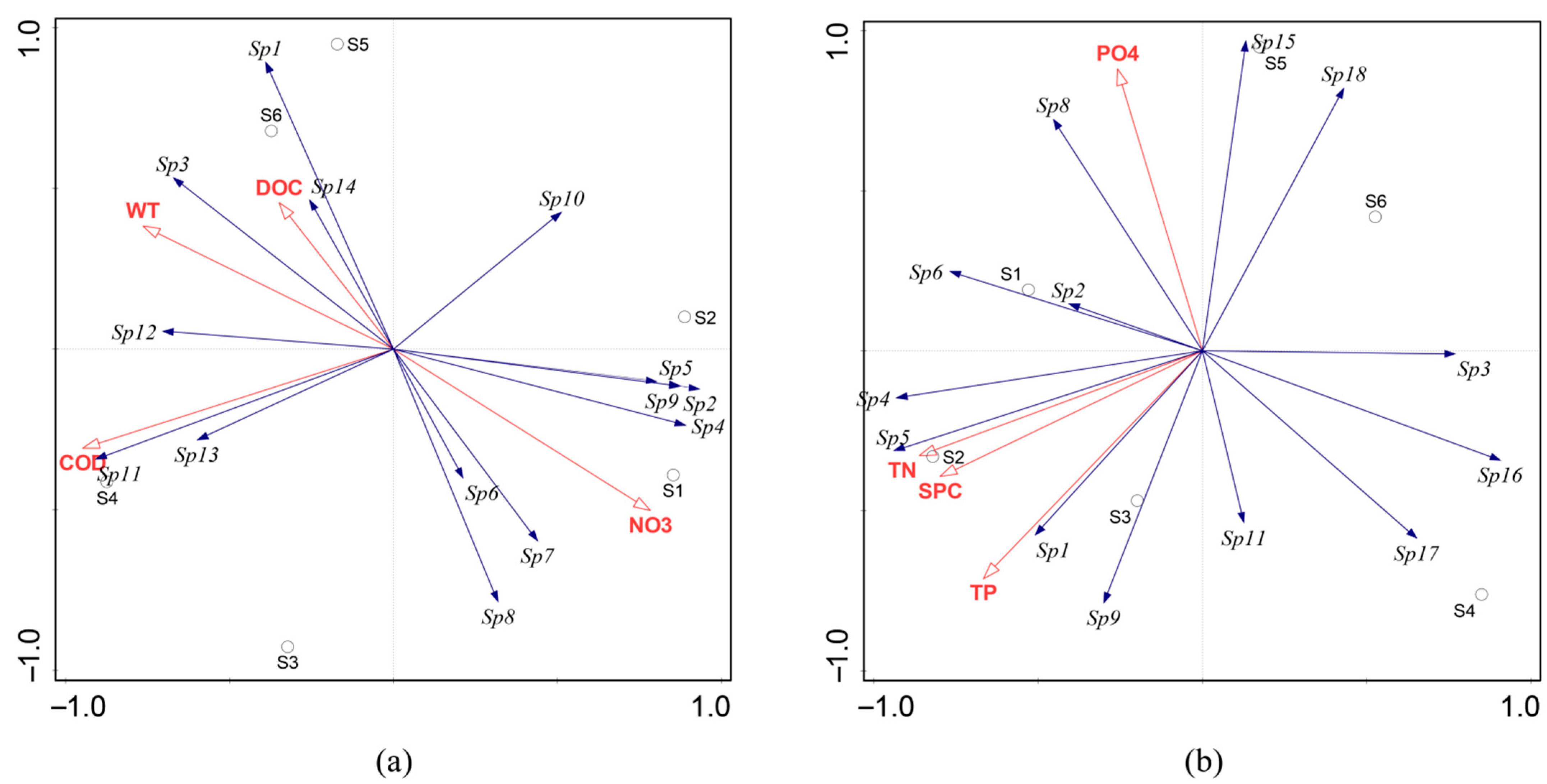
3.4. Correlation between Periphytic Algal Community Structure and Environmental Factors
3.5. Redundancy Analysis of Periphytic Algal Community Structure and Environmental Factors
4. Discussion
4.1. Structural Characteristics of Periphytic Algae Community
4.2. Biodiversity Index of Periphytic Algae and Water Quality Evaluation
4.3. The Relationship between Periphytic Algae and Environmental Factors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nalley, J.O.; O’Donnell, D.R.; Litchman, E. Temperature effects on growth rates and fatty acid content in freshwater algae and cyanobacteria. Algal Res. 2018, 35, 500–507. [Google Scholar] [CrossRef]
- Song, Y.Z.; Zhang, Y.D.; Deng, J.W. Periphytic algae ecology in freshwater lake: A review. Chin. J. Ecol. 2016, 35, 534–541. [Google Scholar]
- DeNicola, D.M.; Kelly, M. Role of periphyton in ecological assessment of lakes. Freshw. Sci. 2014, 33, 619–638. [Google Scholar] [CrossRef]
- Yang, S.; Bertuzzo, E.; Büttner, O.; Borchardt, D.; Rao, P.S.C. Emergent spatial patterns of competing benthic and pelagic algae in a river network: A parsimonious basin-scale modeling analysis. Water Res. 2021, 193, 116887. [Google Scholar] [CrossRef] [PubMed]
- Jäger, C.G.; Borchardt, D. Longitudinal patterns and response lengths of algae in riverine ecosystems: A model analysis emphasising benthic-pelagic interactions. J. Theor. Biol. 2018, 442, 66–78. [Google Scholar] [CrossRef]
- Dunck, B.; Felisberto, S.A.; Nogueira, I.S. Effects of freshwater eutrophication on species and functional beta diversity of periphytic algae. Hydrobiologia 2019, 837, 195–204. [Google Scholar] [CrossRef]
- Casartelli, M.R.; Ferragut, C. The efects of habitat complexity on periphyton biomass accumulation and taxonomic structure during colonization. Hydrobiologia 2018, 807, 233–246. [Google Scholar] [CrossRef]
- Liu, Y.; Fu, J.; Cheng, D.; Lin, Q.; Su, P.; Wang, X.; Sun, H. The spatial pattern of periphytic algae communities and its corresponding mechanism to environmental variables in the Weihe River Basin, China. Hydrol. Res. 2020, 51, 1036–1047. [Google Scholar] [CrossRef]
- Wang, J.; Shi, Y.; Liu, Q.; Li, Z.; Zhang, M.; Xie, S.L. Taxonomic and molecular phylogenetics of bloom-forming algae from the Taiyuan section of Fenhe River, China. J. Lake Sci. 2018, 30, 1332–1342. [Google Scholar]
- Wang, J.; Feng, J.; Xie, S.L.; Zhang, J.M.; Cheng, G.; Lian, Y.J. Phytoplankton diversity and off-flavor-producing Microcystis in the Taiyuan region of the Fenhe River. Acta Ecol. Sin. 2015, 35, 3357–3363. [Google Scholar]
- Liu, Y.Y.; Ayi, Q.L.; Zhang, S.R.; Wu, X.R.; Wan, B.N.; Zhang, X.P.; Zeng, B. Comparative study on the suitability of periphytic algae and phytoplankton in river health assessment. Acta Ecol. Sin. 2020, 40, 3833–3843. [Google Scholar]
- Gao, J.H.; Liu, Y.; Li, K.N.; Yuan, Y.; Li, Y.J.; Zhang, L. Dynamic changes in rotifer diversity and community structure in the Taiyuan section of Fenhe River. J. Hydroecol. 2021, 42, 77–84. [Google Scholar]
- Vlaičević, B.; Matoničkin Kepčija, R.; Gulin, V.; Turković Čakalić, I.; Kepec, M.; Čerba, D. Key drivers influencing the colonization of periphytic ciliates and their functional role in hydrologically dynamic floodplain lake ecosystem. Knowl. Manag. Aquat. Ecosyst. 2021, 422, 33. [Google Scholar] [CrossRef]
- Yang, J.; Wang, F.; Lv, J.; Liu, Q.; Nan, F.; Liu, X.; Xu, L.; Xie, S.; Feng, J. The spatiotemporal contribution of the phytoplankton community and environmental variables to the carbon sequestration potential in an urban river. Environ. Sci. Pollut. Res. Int. 2020, 27, 4814–4829. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.J.; Wang, Y.; Yang, L.L.; Yuan, L.W.; Peng, D.C. Relationship between phytoplankton and environmental factors in landscape water supplemented with reclaimed water. Ecol. Indic. 2015, 58, 113–121. [Google Scholar] [CrossRef]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China: Systematics, Taxonomy and Ecology; Beijing Science Press: Beijing, China, 2006. [Google Scholar]
- Wang, X.; Zhang, F.; Kung, H.T.; Ghulam, A.; Trumbo, A.L.; Yang, J.; Ren, Y.; Jing, Y. Evaluation and estimation of surface water quality in an arid region based on EEM-PARAFAC and 3D fluorescence spectral index: A case study of the Ebinur Lake Watershed, China. Catena 2017, 155, 62–74. [Google Scholar] [CrossRef]
- Das, M.; Semy, K.; Kuotsu, K. Seasonal monitoring of algal diversity and spatiotemporal variation in water properties of Simsang river at South Garo Hills, Meghalaya, India. Sustain. Water Resour. Manag. 2022, 8, 16. [Google Scholar] [CrossRef]
- Zhang, Z.S.; Huang, X.F. Research Methods of Freshwater Plankton; Beijing Press: Beijing, China, 1991. [Google Scholar]
- Song, J.; Yang, X.; Zhang, J.; Long, Y.; Zhang, Y.; Zhang, T. Assessing the variability of heavy metal concentrations in liquid-solid two-phase and related environmental risks in the Weihe River of Shaanxi province, China. Int. J. Environ. Res. Public Health 2015, 12, 8243–8262. [Google Scholar] [CrossRef]
- Mirzahasanlou, J.P.; Ramezanpour, Z.; Nejadsattari, T.; Namin, J.I.; Asri, Y. Temporal and spatial distribution of diatom assemblages and their relationship with environmental factors in Balikhli River (NW Iran). Ecohydrol. Hydrobiol. 2020, 20, 102–111. [Google Scholar] [CrossRef]
- Stevenson, J.; Graham, L. Ecological assessments with algae: A review and synthesis. J. Phycol. 2014, 50, 437–461. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Osorio, N.C.; Cunha, E.R.; Tramonte, R.P.; Mormul, R.P.; Rodrigues, L. Habitat complexity drives the turnover and nestedness patterns in a periphytic algae community. Limnology 2019, 20, 297–307. [Google Scholar] [CrossRef]
- Blanco, S.; Olenici, A.; Ortega, F.; Jiménez-Gómez, F.; Guerrero, F. Identifying environmental drivers of benthic diatom diversity: The case of Mediterranean mountain ponds. PeerJ 2020, 8, e8825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunde-Cimerman, N.; Plemenitaš, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. FEMS Microbiol. Rev. 2018, 42, 353–375. [Google Scholar] [CrossRef]
- Jiao, J.D.; Deng, J.F.; Li, F.K.; Sun, L.P. Analysis for growth characteristics and phosphorus nutrient dynamics of alage in mesohalinous water. Environ. Pollut. Control. 2017, 39, 726–729. [Google Scholar]
- Korneva, L.G.; Solovyeva, V.V. Golden algae (Chrysophyta) in plankton of the Volga River reservoirs: Taxonomic structure, dynamics of diversity, and abundance. Inland Water Biol. 2017, 10, 168–175. [Google Scholar] [CrossRef]
- Seeta, Y.; Reddy, M. Algal diversity and water quality in the rivers of Telangana. Int. J. Recent Sci. Res. 2020, 11, 37702–37704. [Google Scholar]
- Filstrup, C.T.; Hillebrand, H.; Heathcote, A.J.; Harpole, W.S.; Downing, J.A. Cyanobacteria dominance influences resource use efficiency and community turnover in phytoplankton and zooplankton communities. Ecol. Lett. 2014, 17, 464–474. [Google Scholar] [CrossRef]
- Ozer, T.; Erkaya, I.A.; Solak, C.N.; Udoh, A.U. Diversity and ecology of algae from Melen River (western black sea river catchment) in Turkey. Turk. J. Fish. Aquat. Sci. 2018, 18, 1187–1194. [Google Scholar] [CrossRef]
- Jun, S.; Wang, D.B.; Zhou, J.H.; Bai, X.; Bai, K. Community structures of phytoplankton and its relationship with environmental factors in the Lhasa River. Acta Ecol. Sin. 2019, 39, 787–798. [Google Scholar]
- Zhao, G.N.; Pan, B.Z.; Li, Y.; He, H.R.; Zhang, L. Research progress on phytoplankton and periphyton ecology of the waterbodies in the Loess Plateau. Sichuan Environ. 2021, 40, 235–240. [Google Scholar]
- Hao, B.; Wu, H.; Li, W.; Xing, W. Periphytic algae mediate interactions between neighbor and target submerged macrophytes along a nutrient gradient. Ecol. Indic. 2020, 110, 1–8. [Google Scholar] [CrossRef]
- Prieto-Barajas, C.M.; Valencia-Cantero, E.; Santoyo, G. Microbial mat ecosystems: Structure types, functional diversity, and biotechnological application. Electron. J. Biotechnol. 2018, 31, 48–56. [Google Scholar] [CrossRef]
- Medeiros, G.; Padial, A.A.; Amaral, M.W.W.; Ludwig, T.A.V.; Bueno, N.C. Environmental variables likely influence the periphytic diatom community in a subtropical lotic environment. Limnologica 2020, 80, 125718. [Google Scholar] [CrossRef]
- Shevchenko, T.F.; Klochenko, P.D.; Timchenko, V.M.; Dubnyak, S.S. Epiphyton of a cascade plain reservoir under different hydrodynamic conditions. Ecohydrol. Hydrobiol. 2019, 19, 407–416. [Google Scholar] [CrossRef]
- Wang, Q.; Li, X.; Yan, T.; Song, J.; Yu, R.; Zhou, M. Laboratory simulation of dissolved oxygen reduction and ammonia nitrogen generation in the decay stage of harmful algae bloom. J. Oceanol. Limnol. 2021, 39, 500–507. [Google Scholar] [CrossRef]
- Baragi, L.V.; Khandeparker, L.; Anil, A.C. Influence of elevated temperature and pCO2 on the marine periphytic diatom Navicula distans and its associated organisms in culture. Hydrobiologia 2015, 762, 127–142. [Google Scholar] [CrossRef]
- Rhomad, H.; Khalil, K.; Neves, R.; Bougadir, B.; Elkalay, K. Modeling investigation of the nutrients and phytoplankton dynamics in the Moroccan Atlantic coast: A case study of Agadir coast. Ecol. Model. 2021, 447, 109510. [Google Scholar] [CrossRef]
- Chou, Q.; Nielsen, A.; Andersen, T.K.; Hu, F.; Chen, W.; Zhang, X.; Cao, T.; Ni, L.; Jeppesen, E.; Trolle, D. Assessing impacts of changes in external nutrient loadings on a temperate Chinese drinking water reservoir. Front. Environ. Sci. 2021, 2, 9–23. [Google Scholar] [CrossRef]
- Du, Y.N. The Effect of Dissolved Organic Matter and Aquatic Plant Humidification on Chemical Oxygen Demand in Yilong Lake. Master’s Thesis, Kunming University of Scince and Technology, Kunming, China, 2019. [Google Scholar]
- Shevchenko, T.F.; Klochenko, P.D.; Bilous, O.P. Response of epiphytic algae to heavy pollution of water bodies. Water Environ. Res. 2018, 90, 706–718. [Google Scholar] [CrossRef]
- Oliveira, L.F.V.; Margis, R. The source of the river as anursery for microbial diversity. PLoS ONE 2015, 10, e0120608. [Google Scholar]
- Wang, L.; Zhang, J.; Li, H.; Yang, H.; Peng, C.; Peng, Z.; Lu, L. Shift in the microbial community composition of surface water and sediment along an urban river. Sci. Total Environ. 2018, 627, 600–612. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.L.; Chen, W.; Zhao, R.; Li, Z.Y.; Dong, Y.L.; Zhang, R.; Wang, Y.Y.; Jin, X.W. Periphytic algae community structure and its relation to environment factors in the main stream of the Songhua River from 2014 to 2019. Huan Jing Ke Xue 2021, 42, 819–830. [Google Scholar] [PubMed]
- Yang, L.; Li, J.X.; Qian, T.Y.; Yin, X.W.; Ruan, Y.J. Quantitative analysis of nitrogen and phosphorus nutrients for periphyton population in Taizi River. Asian J. Ecotoxicol. 2019, 14, 104–117. [Google Scholar]
- Wang, Z.; Wang, W.K.; Zhang, Z.Y.; Hou, X.; Ma, Z.; Chen, B. River-groundwater interaction affected species composition and diversity perpendicular to a regulated river in an arid riparian zone. Glob. Ecol. Conserv. 2021, 27, e01595. [Google Scholar] [CrossRef]
- Cui, L.; Lu, X.; Dong, Y.; Cen, J.; Cao, R.; Pan, L.; Lu, S.; Ou, L. Relationship between phytoplankton community succession and environmental parameters in Qinhuangdao coastal areas, China: A region with recurrent brown tide outbreaks. Ecotoxicol. Environ. Saf. 2018, 159, 85–93. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | S1 | S2 | S3 | S4 | S5 | S6 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Summer | Winter | Summer | Winter | Summer | Winter | Summer | Winter | Summer | Winter | Summer | Winter | |
| WT (°C) | 25.05 ± 0.55 | 4.50 ± 1.80 | 27.10 ± 1.00 | 4.05 ± 1.25 | 27.15 ± 0.85 | 5.05 ± 1.85 | 28.15 ± 0.15 | 4.15 ± 1.15 | 27.75 ± 0.15 | 5.45 ± 2.45 | 28.05 ± 0.05 | 6.10 ± 2.30 |
| pH | 8.61 ± 0.32 | 8.31 ± 0.69 | 8.69 ± 0.34 | 8.02 ± 0.38 | 9.11 ± 0.05 | 7.91 ± 0.42 | 8.81 ± 0.36 | 7.87 ± 0.30 | 9.12 ± 0.06 | 7.87 ± 0.42 | 8.76 ± 0.30 | 7.27 ± 0.04 |
| SPC (us/cm) | 869.50 ± 17.50 | 1036.05 ± 316.95 | 855.00 ± 19.00 | 990.65 ± 340.35 | 848.00 ± 39.00 | 795.90 ± 227.10 | 809.50 ± 38.50 | 653.90 ± 142.70 | 797.00 ± 11.00 | 538.50 ± 36.10 | 860.00 ± 14.00 | 661.35 ± 1.05 |
| SAL (ppt) | 0.43 ± 0.01 | 0.51 ± 0.160 | 0.42 ± 0.01 | 0.49 ± 0.17 | 0.41 ± 0.02 | 0.39 ± 0.11 | 0.40 ± 0.02 | 0.32 ± 0.07 | 0.38 ± 0.02 | 0.26 ± 0.02 | 0.42 ± 0.01 | 0.32 ± 0.01 |
| DO (mg/L) | 7.75 ± 0.77 | 15.40 ± 0.62 | 8.71 ± 3.14 | 17.70 ± 2.77 | 8.97 ± 3.33 | 18.71 ± 4.94 | 8.84 ± 3.52 | 19.93 ± 2.03 | 9.30 ± 1.47 | 17.54 ± 1.99 | 9.43 ± 3.79 | 13.01 ± 0.20 |
| TDS (mg/L) | 565.55 ± 12.95 | 675.68 ± 208.33 | 605.75 ± 40.25 | 643.83 ± 220.68 | 552.50 ± 26.00 | 516.43 ± 146.58 | 529.75 ± 22.75 | 425.10 ± 92.95 | 520.00 ± 6.50 | 350.35 ± 23.40 | 559.01 ± 6.45 | 429.65 ± 0.65 |
| SD (cm) | 0.60 ± 0.10 | 0.76 ± 0.14 | 0.46 ± 0.08 | 0.68 ± 0.16 | 0.55 ± 0.20 | 0.63 ± 0.03 | 0.48 ± 0.06 | 0.95 ± 0.15 | 0.50 ± 0.02 | 0.94 ± 0.22 | 0.51 ± 0.01 | 0.95 ± 0.15 |
| TN (mg/L) | 0.53 ± 0.06 | 3.67 ± 2.44 | 0.59 ± 0.18 | 3.16 ± 2.14 | 0.53 ± 0.11 | 2.98 ± 2.39 | 0.51 ± 0.05 | 1.94 ± 1.16 | 0.49 ± 0.05 | 1.83 ± 0.94 | 0.78 ± 0.04 | 1.61 ± 0.72 |
| (mg/L) | 0.31 ± 0.05 | 1.14 ± 0.05 | 0.32 ± 0.06 | 0.94 ± 0.24 | 0.30 ± 0.02 | 0.38 ± 0.32 | 0.38 ± 0.07 | 0.30 ± 0.26 | 0.34 ± 0.03 | 0.09 ± 0.01 | 0.50 ± 0.25 | 0.04 ± 0.01 |
| (mg/L) | 0.17 ± 0.01 | 0.16 ± 0.02 | 0.11 ± 0.05 | 0.15 ± 0.03 | 0.09 ± 0.02 | 0.13 ± 0.03 | 0.07 ± 0.01 | 0.12 ± 0.02 | 0.06 ± 0.01 | 0.10 ± 0.01 | 0.06 ± 0.01 | 0.13 ± 0.01 |
| TP (mg/L) | 0.04 ± 0.01 | 0.10 ± 0.03 | 0.07 ± 0.03 | 0.13 ± 0.06 | 0.06 ± 0.03 | 0.12 ± 0.01 | 0.07 ± 0.03 | 0.10 ± 0.01 | 0.09 ± 0.02 | 0.08 ± 0.01 | 0.15 ± 0.09 | 0.08 ± 0.01 |
| (mg/L) | 0.02 ± 0.01 | 0.01 ± 0.01 | 0.05 ± 0.04 | 0.01 ± 0.01 | 0.05 ± 0.04 | 0.01 ± 0.01 | 0.06 ± 0.04 | 0.01 ± 0.00 | 0.05 ± 0.03 | 0.01 ± 0.01 | 0.04 ± 0.02 | 0.01 ± 0.00 |
| COD (mg/L) | 21.67 ± 13.20 | 14.67 ± 5.67 | 21.17 ± 5.74 | 15.67 ± 7.67 | 34.33 ± 12.47 | 22.17 ± 5.50 | 37.00 ± 4.29 | 16.00 ± 7.00 | 26.83 ± 19.28 | 21.67 ± 6.67 | 28.83 ± 20.04 | 17.00 ± 6.33 |
| DOC (mg/L) | 6.08 ± 0.15 | 5.93 ± 0.37 | 5.69 ± 0.18 | 6.28 ± 0.56 | 5.72 ± 0.07 | 5.70 ± 0.02 | 6.08 ± 0.55 | 5.70 ± 0.12 | 6.08 ± 0.23 | 5.70 ± 0.08 | 6.13 ± 0.40 | 5.64 ± 0.15 |
| Number | Dominant Species | Number | Dominant Species |
|---|---|---|---|
| Sp1 | Cyclotella | Sp10 | Fragilaria |
| Sp2 | Gomphonema | Sp11 | Scenedesmus |
| Sp3 | Melosira | Sp12 | Pediastrum |
| Sp4 | Navicula | Sp13 | Tetrastrum |
| Sp5 | Nitzschia | Sp14 | Oocystis |
| Sp6 | Synedra | Sp15 | Ulothrix |
| Sp7 | Pinnularia | Sp16 | Cryptomonas |
| Sp8 | Oscillatoria | Sp17 | Merismopedia |
| Sp9 | Planktothrix | Sp18 | Oedogonium |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, K.; Yang, J.; Lv, J.; Liu, Q.; Liu, X.; Xie, S.; Feng, J. Structural Characteristics of Periphytic Algal Community and Its Relationship with Environmental Factors in the Taiyuan Region of the Fenhe River. Water 2022, 14, 2151. https://doi.org/10.3390/w14142151
Zhao K, Yang J, Lv J, Liu Q, Liu X, Xie S, Feng J. Structural Characteristics of Periphytic Algal Community and Its Relationship with Environmental Factors in the Taiyuan Region of the Fenhe River. Water. 2022; 14(14):2151. https://doi.org/10.3390/w14142151
Chicago/Turabian StyleZhao, Kangxu, Jing Yang, Junping Lv, Qi Liu, Xudong Liu, Shulian Xie, and Jia Feng. 2022. "Structural Characteristics of Periphytic Algal Community and Its Relationship with Environmental Factors in the Taiyuan Region of the Fenhe River" Water 14, no. 14: 2151. https://doi.org/10.3390/w14142151