
The Effect of Algicidal and Denitrifying Bacteria on the Vertical Distribution of Cyanobacteria and Nutrients
by
Qing Zhou
1,2,*, 
Yan Wang
1,
Xuezheng Wen
1,
Haiqin Liu
1,
Yingying Zhang
1,2 and
Zhiyong Zhang
1,2,* 1
Institute of Agricultural Resources and Environment, Liuhe Observation and Experimental Station of National Agricultural Environment, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China
2
School of the Environment and Safety Engineering, Jiangsu University, Zhenjiang 212013, China
*
Authors to whom correspondence should be addressed.
Water 2022, 14(13), 2129; https://doi.org/10.3390/w14132129
Submission received: 20 May 2022
/
Revised: 30 June 2022
/
Accepted: 30 June 2022
/
Published: 4 July 2022
(This article belongs to the Special Issue Wastewater Bio-Ecological Treatment)
Abstract
:Algicidal bacteria combined with the ability of aerobic denitrification is considered to be a promising way to control harmful cyanobacterial bloom and remove nitrogen. However, the effect of these bacteria on the vertical distribution of colonial cyanobacteria and nutrients remained unknown. In this study, two algicidal and denitrifying bacteria were respectively co-cultured with the colonial Microcystis aeruginosa to construct the microcosm systems, and then the cyanobacteria number, the ratio of bacterial to cyanobacterial abundance, the content of dissolved nitrogen, phosphorus and organic carbon in different water layers were investigated. The results showed that the distribution difference of Microcystis among the vertical water layers was further enlarged due to the short-term influence of algicidal bacteria Brevundimonas diminuta and Pseudomonas stutzeri. The number of Microcystis in the lower layer was further reduced by the inhibitory effect of the algicidal bacteria. However, there was a dramatic increase in the number of Microcystis in the upper layer, even when the ratio of algicidal bacteria to cyanobacteria increased significantly. B. diminuta and P. stutzeri both greatly promoted the removal of dissolved total nitrogen in the upper and middle layers of cyanobacteria blooming water, but they also boosted the release of dissolved phosphorus in all layers. These results enable us to better understand the possible limitations of algicidal bacteria in their application to control cyanobacteria blooms.
1. Introduction
Cyanobacterial blooms have been known to be a serious threat to public health and aquatic ecosystem functioning [1]. Toxic Microcystis aeruginosa is the most primary bloom-forming cyanobacteria in eutrophic freshwaters [2]. It can produce hepatotoxic and hepatocarcinogenic microcystins. Some water crises, such as the Wuxi drinking water crisis in China and the recreational water risk in Portugal, further increased public health concerns [3,4]. The control and mitigation measures of Microcystis bloom have been developed and highlighted by researchers. Many algicidal bacteria have been isolated [5,6] and constitute the major group of microorganisms showing inhibitory effects on cyanobacteria [7]. Algicidal bacteria, as a biological control method, are expected to be environmentally friendly and feasible for bloom control [8]. The research about algicidal bacteria mostly focuses on cyanobacteria species specificity [9], their attacking mode [10], the relationship between their concentration threshold and cyanobacteria decay [11], definite cyanobactericidal substances [12] and their cyanobactericidal mechanism [13]. Most Microcystis strains tested in the laboratory are not in a colonial state [6]. During coexistence with algicidal bacteria, vertical stratification of these cyanobacteria will not happen in glass Erlenmeyer flasks. However, Microcystis in the natural water environment tends to migrate vertically through changing cell density and regulating buoyancy. The size of colonies and wind speed have significant effects on the migration, which leads to the stratified distribution of Microcystis [14]. Then, the physical and chemical environments in different water layers, such as dissolved oxygen (DO) and pH, are affected differently [15]. It is not clear whether the vertically stratified distribution of cyanobacteria will interfere with the inhibitory activity of algicidal bacteria. In addition, algicidal bacteria combined with the ability of denitrification are considered to have the dual effects of nitrogen and cyanobacteria bloom control [16]. However, it remains poorly defined how these bacteria further affect the vertical distribution of cyanobacteria and nutrients in the corresponding water layer.
The aims of this study were to explore the response of algicidal and denitrifying bacteria to the vertical distribution of cyanobacteria, and their effects on the distribution of cyanobacteria and nutrients in different water layers. For this, two algicidal bacteria were respectively co-cultured with the colonial Microcystis; then, the cyanobacteria number, the ratio of bacterial to cyanobacterial abundance, the content of dissolved nitrogen, phosphorus and organic carbon in different water layers were examined. The results of this study enable us to better understand the possible limitations of algicidal bacteria in their application to control cyanobacterial blooms as well as help to seek strategies for more effective use of them.
2. Materials and Methods
2.1. Strains and Culture Conditions
M. aeruginosa (FACHB 912) was purchased from Institute of Hydrobiology, Chinese Academy of Sciences (Wuhan, China). This strain was cultivated in 10% modified Hoagland’s medium (pH 7.0) supplemented with Ca2+ and Fe2+ at the final concentration of 36 mg/L and 2.8 mg/L to maintain colony status. The incubator was maintained at 28 °C and illuminated with a light intensity of 2800 lux by cool-white, fluorescent lamps in 12 h diurnal cycles.
Brevundimonas diminuta Z1 (CGMCC 20909) and Pseudomonas stutzeri ZJ4 (CGMCC 20910) were isolated from a water bloom sample in the aquaculture ponds of the Taihu Lake Basin and deposited to the China General Microbiological Culture Collection Center (Beijing, China). They both exhibited algicidal activity (Table 1). These strains were grown aerobically in Luria-Bertani (LB) medium at 28 °C, respectively. All media were sterilized at 121 °C for 20 min before use.
2.2. Co-Existence Experiments
M. aeruginosa was amplified in 10% modified Hoagland’s medium (pH 6.8) supplemented with Ca2+ and Fe2+ in aseptic conical flasks (2 L) for 15 days with a thorough shake once a day. When it grew exponentially and reached the density of 1.07 × 107 cells/mL, it was transferred into cylindrical plastic containers (2.8 L, 50 cm high). These containers were pre-disinfected with 75% alcohol. The upper one-third part of the container was transparent, and the lower two-thirds part was shielded with aluminum foil. B. diminuta Z1 and P. stutzeri ZJ4 were individually collected during the exponential growth period and washed twice with 10% modified Hoagland’s medium supplemented with Ca2+ and Fe2+. Then, B. diminuta Z1 and P. stutzeri ZJ4 were respectively added into M. aeruginosa cultures and mixed well in cylindrical containers at a final density of 2.93 × 108 cells/mL (B treatment) and 2.75 × 108 cells/mL (P treatment) in the microcosm systems, while the M. aeruginosa cultures without B. diminuta Z1 or P. stutzeri ZJ4 were mixed and used as controls. All microcosm experiments were conducted with three independent replicates. All treatments were maintained under the culturing conditions described above. The cool-white, fluorescent lamps were located above the culture containers.
At day 0, initial mixed samples were collected. After 3 days of static cultures, DO and pH were measured in three vertical stratified sampling sites with pre-fixed multi-parameter water quality monitoring sondes (HQ40d, HACH, Loveland, USA). The upper sampling site was located in the near-surface layer of the fluid column (0–2 cm). The middle site was in the middle of the fluid column (23–25 cm). The lower site was at the bottom of the fluid column (46–48 cm). Vertically stratified water samples were conducted by the slow siphon method without breaking the stratification.
2.3. Measures of Cyanobacteria Cell Density
M. aeruginosa cells were fixed by formaldehyde (2% final conc.) and counted in a hemocytometer using a microscope (CX41, Olympus, Tokyo, Japan), after dispersion by ultrasonication (5 s).
2.4. The Ratio of Bacterial to Cyanobacterial Abundance
Water samples were filtered through a 0.22 μm Millipore membrane. The filtered membrane was used for subsequent DNA extraction. Total genomic DNA was extracted by using a PowerWater® Sterivex™ DNA Isolation Kit (MO BIO Laboratories, Carlsbad, CA, USA). The concentration of DNA was determined via spectrophotometer analysis. Amplification and sequencing of the 16S rRNA gene V4 region were performed at Shanghai Personal Biotechnology Co., Ltd. (Shanghai, China). PCR was carried out using Q5™ High-Fidelity DNA Polymerase (NEB, Ipswich, MA, USA) with forward primer 520F (5′-AYTGGGYDTAAAGNG-3′) and reverse primer 802R (5′-TACNVGGGTATCTAATCC-3′). The amplicon libraries were generated using TruSeq Nano DNA LT Library Prep Kit (Illumina, San Diego, CA, USA) and then sequenced on the Illumina MiSeq, using MiSeq Reagent Kit V3 (600 cycles) for a 2 × 250 bp paired-end reads. The sequences with 97% similarity were assigned into the same operational taxonomic unit (OTU) by using the Qiime software platform. The relative abundances were calculated by the abundance of each OTU at specific taxonomic ranks, and then the ratio of bacterial to cyanobacterial abundance was obtained.
2.5. Chemical Analyses
Water samples were filtered through a 0.45 μm Millipore membrane. Nitrate-N (NO3−-N) concentration was analyzed using the UV spectrophotometric screening method. Ammonium-N (NH4+-N) concentration was determined using Nessler’s reagent spectrophotometry method. Nitrite-N (NO2−-N) concentration was determined using N-(1-naphthyl)-1, 2-diaminoethane dihydrochloride method. Dissolved total nitrogen (DTN) was measured using alkaline potassium persulfate digestion-UV spectrophotometric method. Phosphate-phosphorous (PO43−-P) was measured using ammonium molybdate spectrophotometric method. Dissolved total phosphorous (DTP) was measured using alkaline potassium persulfate digestion-ammonium molybdate spectrophotometric method. Dissolved organic carbon (DOC) was determined using a TOC analyzer (Vario TOC, Langenselbold, Germany).
2.6. Statistical Analysis
Data were expressed as mean ± standard deviation (SD). Differences between controls and treatments were determined by one-way ANOVA. Post hoc test was used to perform statistical multiple comparisons. LSD adjustment was adopted when the assumption of equal variances was met, and Games-Howell adjustment was adopted when the assumption of equal variances was not met. Results were expressed as letters. Different letters were marked in histograms to indicate significant differences (p < 0.05). Student’s t-test was used to assess the correlation between groups of parameters and the significance level (two-tailed; p < 0.05 was considered significant). All statistical analyses were performed using IBM SPSS Statistics 20.0 (IBM SPSS Inc., Chicago, IL, USA).
3. Results
3.1. Spatial Distribution of Colonial Microcystis
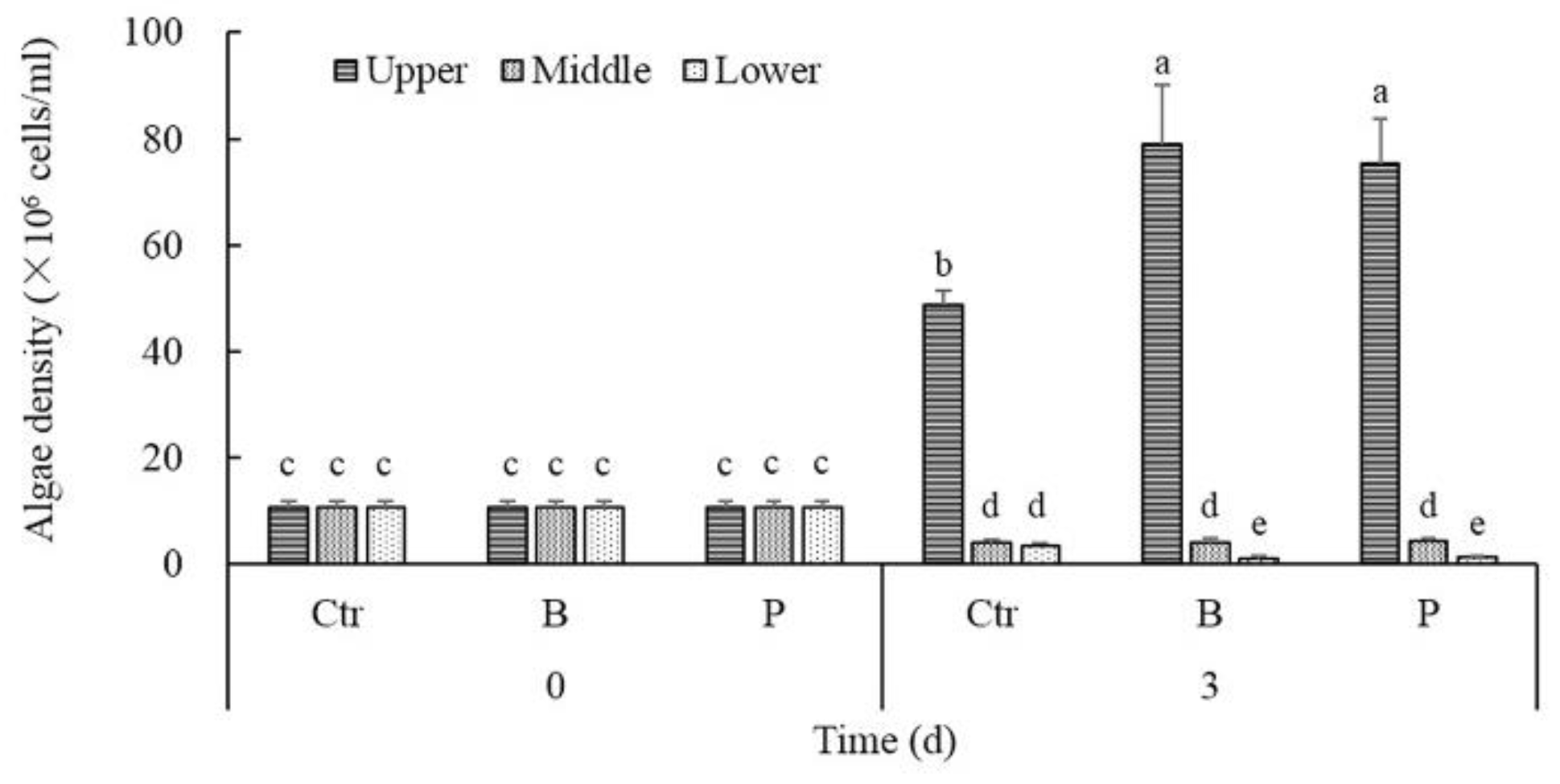
Based on the different cyanobacteria concentrations in the vertical layers of water column under calm conditions for 3 days, colonial M. aeruginosa could migrate to the near-surface region and form surface scum even after thorough mixing. As shown in Figure 1, the density of cyanobacteria cells in the upper layer was up to 12.98 times that of the middle or lower layer in controls after 3 days of static cultures, unlike the initial homogeneous state. Compared to the controls, the density of cyanobacteria cells in the upper layer of the B treatments and P treatments at day 3 increased significantly by an average of 62.33% and 54.79%, respectively. On the contrary, the density of cyanobacteria cells in the lower layer of the B treatments and P treatments were on average 70.09% and 63.55% lower than controls, respectively. There was no significant difference between the cyanobacteria density in the middle layer of treatments and controls.
3.2. Characteristics of Environmental Parameters in Vertical Space
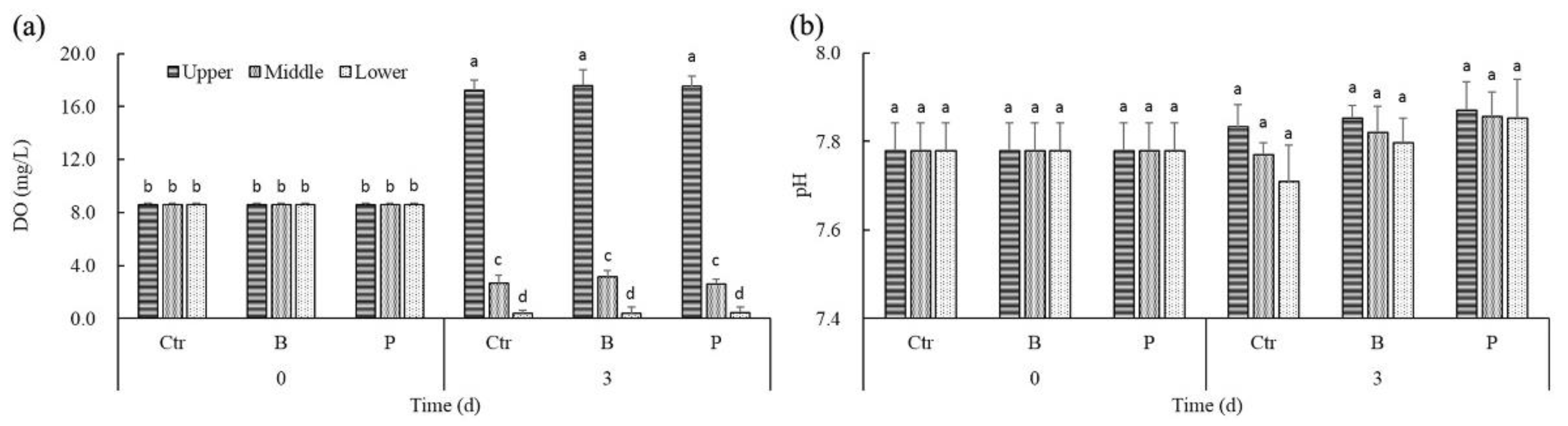
Colonial M. aeruginosa was positively buoyant. After 3 days of static cultures, DO concentrations varied between 0.02 mg/L and 18.43 mg/L, depending on the vertical position and cyanobacteria density in water column (Figure 2a). There was no significant difference among DO concentrations of the same water layers in controls and treatments. Correlation analysis revealed that there existed significantly positive correlation between DO and the density of cyanobacteria cells, no matter in controls or treatments (rctr = 0.989, p < 0.05; rB = 0.977, p < 0.05; rP = 0.984, p < 0.05). However, along with the significant vertical gradients of oxygen at day 3, the difference of pH among water layers was not remarkable (Figure 2b). It might be due to the use of modified Hoagland’s medium.
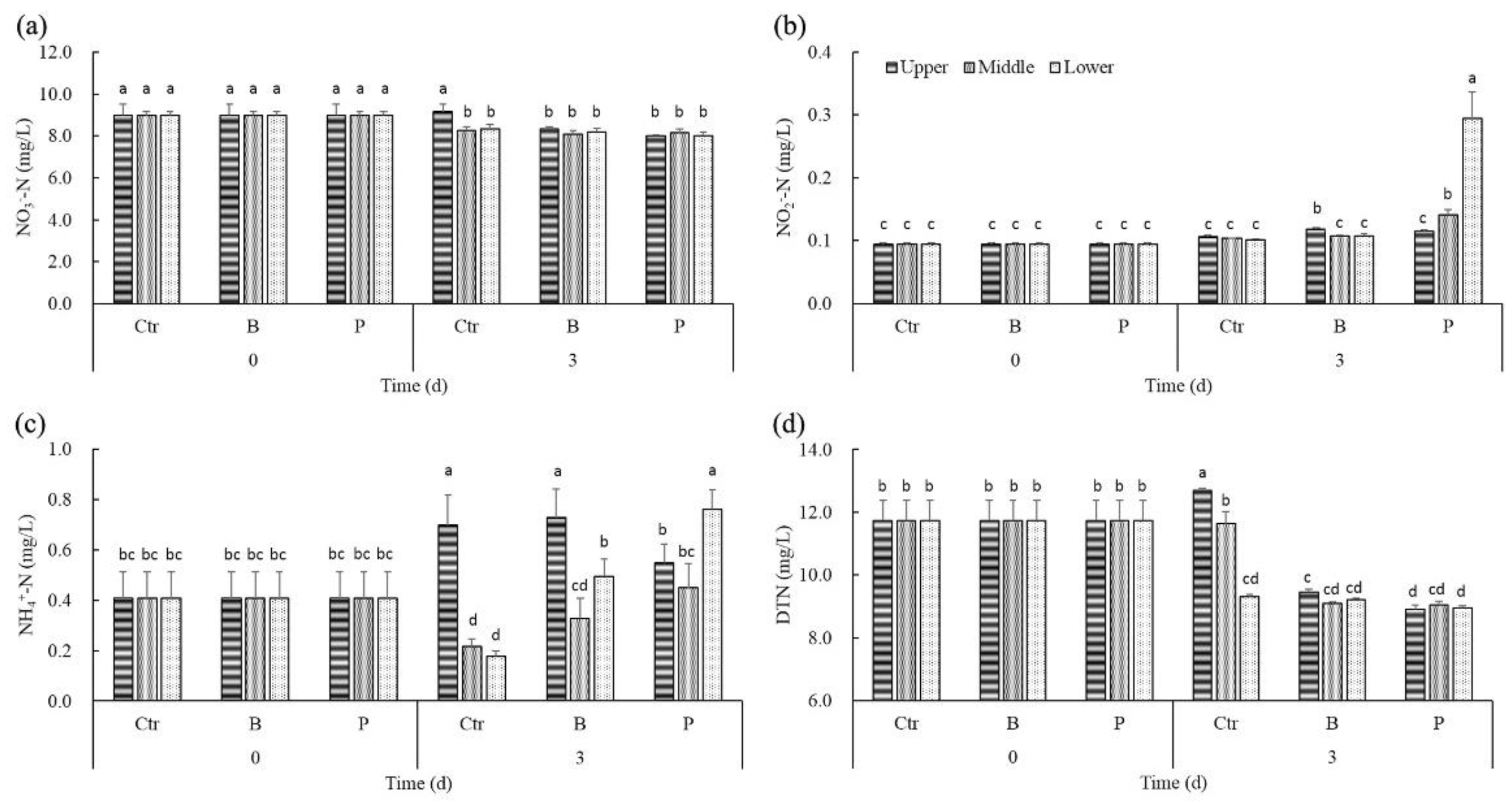
In the medium, the main form of nitrogen was NO3−-N. After 3 days, NO3−-N concentrations of the upper water layer were significantly higher than that of the middle and lower layer in controls (Figure 3a). However, there was no obvious difference among the NO3−-N concentrations in different water layers of B treatments and P treatments. Significant positive correlations were found between NO3−-N concentrations and the density of cyanobacteria cells only in controls (rctr = 0.920, p < 0.05).
NO2−-N distribution in controls after 3 days was the same as the initial homogeneous state (Figure 3b). However, NO2−-N concentrations of the middle and lower water layers were significantly higher than that of the upper layer in P treatments. Significant negative correlations were found between NO2−-N concentrations and DO in P treatments (rP = −0.675, p < 0.05), while significant positive correlations were found between NO2−-N concentrations and DO in B treatments (rB = 0.920, p < 0.05).
NH4+-N concentrations of the upper layer amounted to about three to four times that of the middle or lower layer in controls, unlike the initial homogeneous state. NH4+-N distribution in B treatments was similar to that in controls after 3 days, except that NH4+-N concentrations of the lower layer in B treatments were enhanced as compared to controls (Figure 3c). However, NH4+-N concentrations of the lower layer were significantly higher than that of the upper and middle layers in P treatments, while its NH4+-N concentrations of upper layer were cut down as compared to controls at day 3. Correlation analysis indicated that NH4+-N concentrations had a significantly positive correlation with the density of cyanobacteria cells only in controls (rctr = 0.954, p < 0.05).
The trends of DTN distribution after 3 days in controls and treatments were similar to NO3−-N. Unlike the significant vertical gradients of DTN in controls, there was no marked difference among the DTN concentrations in different water layers of B treatments or P treatments (Figure 3d). DTN concentrations in the upper layer of the B treatments were on average 23.76% lower than controls, while DTN concentrations in the upper layer of the P treatments were significantly lower than B treatments. Significant positive correlations were found between DTN concentrations and the density of cyanobacteria cells only in controls (rctr = 0.738, p < 0.05).
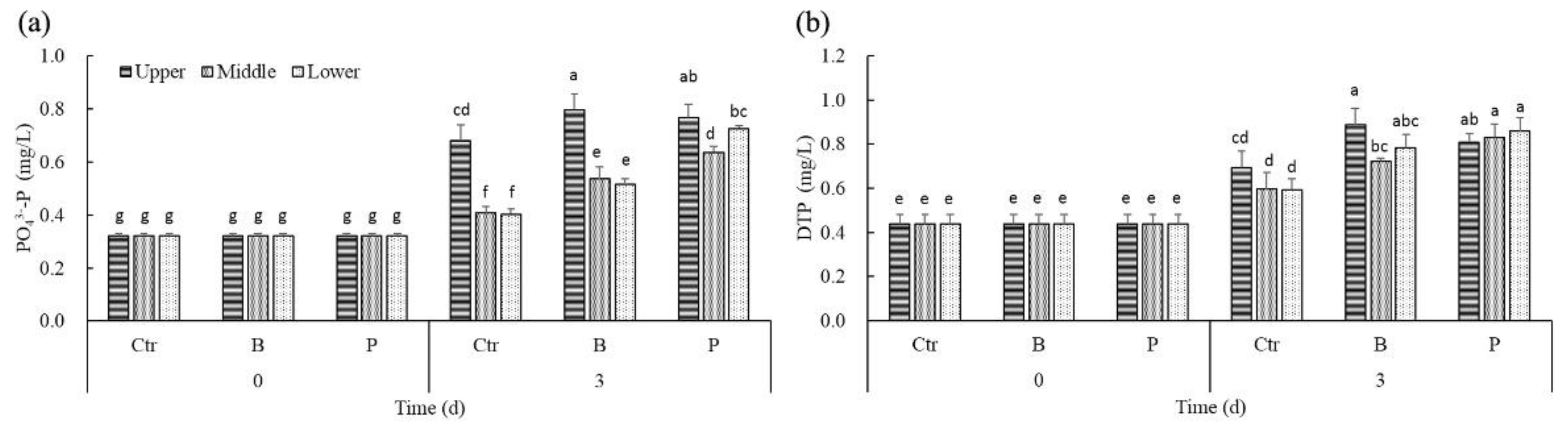
Different from the initial homogeneous state, PO43−-P concentrations in the upper layer of controls and treatments were higher than that of the middle and lower layers after 3 days (Figure 4a). The PO43−-P concentrations of the different water layers both in B treatments and P treatments were significantly higher than the corresponding concentrations in controls. Significant positive correlations were found between PO43−-P concentrations and the density of cyanobacteria cells only in controls (rctr = 0.965, p < 0.05).
Just like PO43−-P, the DTP concentrations of the different water layers both in B treatments and P treatments were also significantly higher than the corresponding concentrations in controls (Figure 4b). Significant positive correlations were found between DTP concentrations and the density of cyanobacteria cells only in controls (rctr = 0.661, p < 0.05).
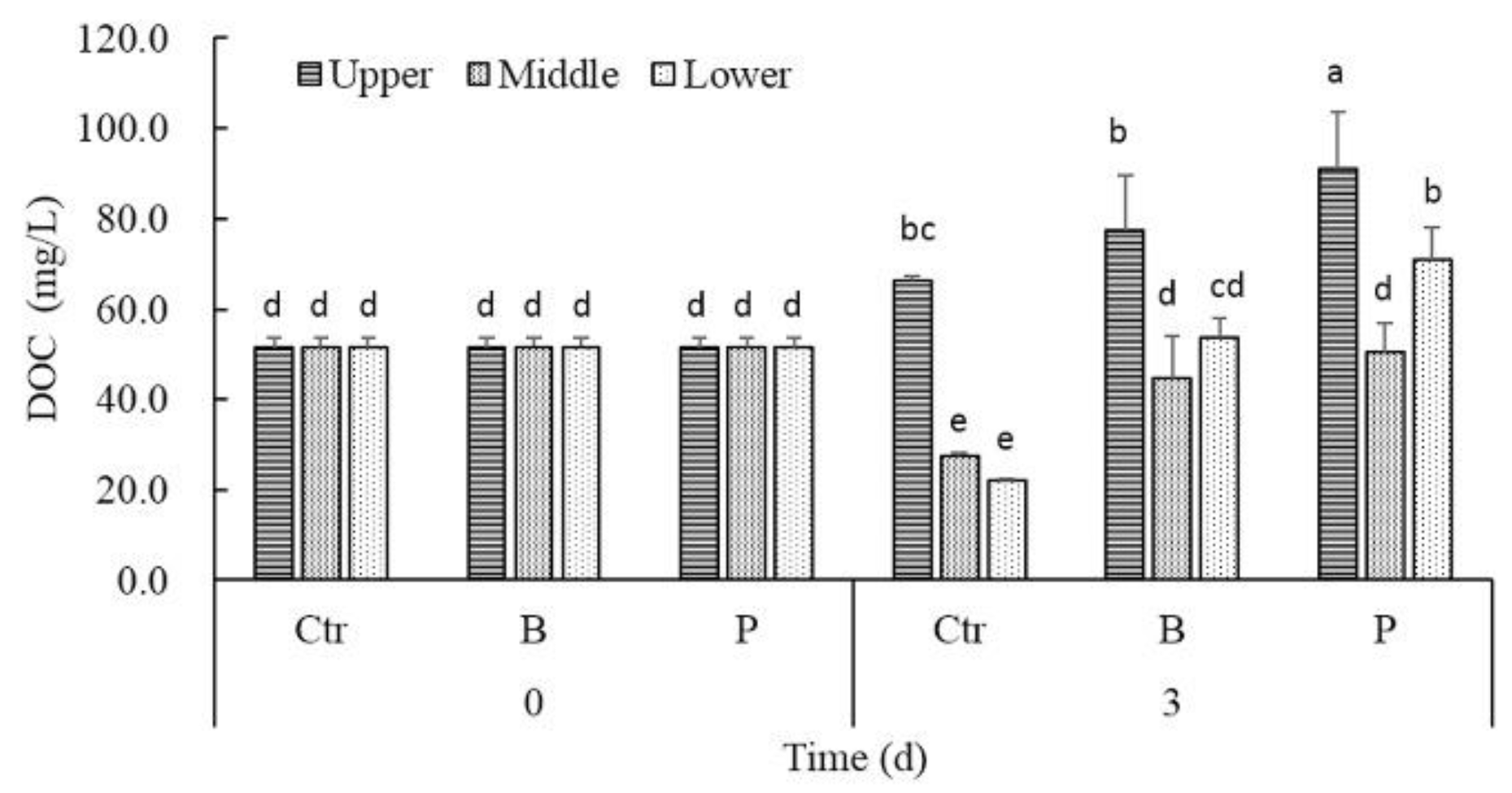
The trends of DOC gradients in controls and treatments after 3 days were similar to PO43−-P. The DOC concentrations in the middle or lower layer of both B treatments and P treatments were significantly higher than that of controls (Figure 5). Correlation analysis revealed that there existed significantly positive correlation between DOC and the density of cyanobacteria cells only in controls (rctr = 0.990, p < 0.05).
3.3. Shifts in Ratios of Bacteria to Microcystis
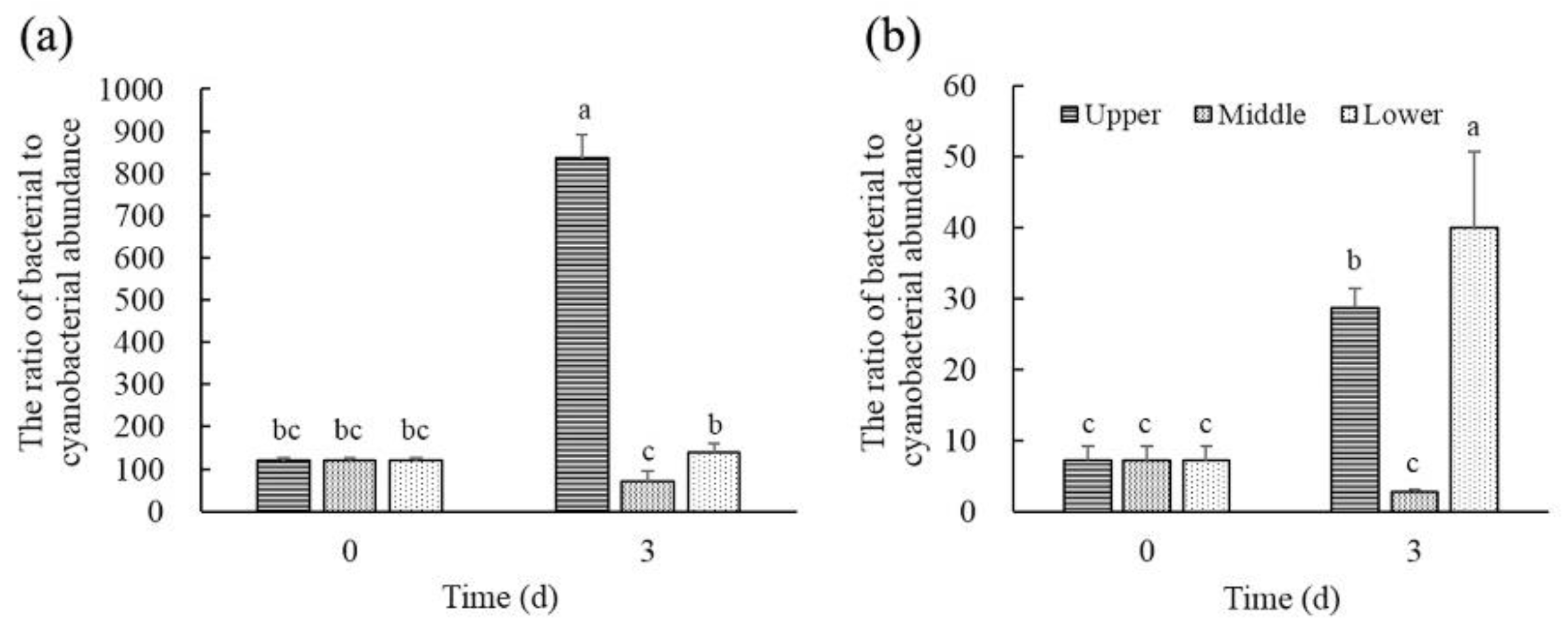
As shown in Figure 6, the average ratio of bacterial to cyanobacterial abundance in the upper layer of B treatments and P treatments at day 3 remarkably increased to 6.95 times and 4.02 times of the initial, respectively. The average ratio of bacterial to cyanobacterial abundance in the middle layer of B treatments and P treatments at day 3 respectively decreased by 42.06% and 61.29% compared with their initial value. Moreover, the ratio of bacterial to cyanobacterial abundance of the middle layer was significantly lower than that of the upper and lower layers both in B treatments and P treatments. It was different from B treatments that the ratio of bacterial to cyanobacterial abundance in the lower layer of P treatments was significantly higher than that in the upper and middle layers. Correlation analysis indicated that there was no significant correlation between the ratio of bacterial to cyanobacterial abundance and the density of cyanobacteria cells in B treatments and P treatments. Moreover, the ratio of bacterial to cyanobacterial abundance showed a significant correlation with NO3−-N concentrations (rB = 0.896, p < 0.05; rP = −0.694, p < 0.05), NH4+-N concentrations (rB = 0.850, p < 0.05; rP = 0.828, p < 0.05) and PO43−-P concentrations (rB = 944, p < 0.05; rP = 0.693, p < 0.05) both in B and P treatments.
4. Discussion
Under the influence of low wind speed and low wave height, the vertical distribution of cyanobacteria was uneven in many inland waters and cyanobacterial surface scum was often detected [17,18]. In this experiment, the vertical gradient distribution of colonial cyanobacteria was successfully simulated by the induction of high calcium ions in the water column. The data showed that the heterogeneous distribution of cyanobacteria also affected the physical and chemical properties of different water layers.
The biological control of cyanobacteria by using algicidal bacteria is a promising approach in an aquatic environment [11,19]. The two bacteria we used have been verified to be able to have algicidal effects on the unicellular Microcystis of the same strain in a short time, which was in accordance with previous reports on other Microcystis strains [20,21,22]. P. stutzeri ZJ4 belongs to one of the top three genera in cyanobactericidal bacteria [6]. Interestingly, however, they were found to have different effects on the populations of colonial Microcystis distributed in the upper, middle and lower layers. The biomass of colonial Microcystis in the upper layer was significantly elevated in B treatments and P treatments, while the number of the corresponding bacteria in the same layer was also increased significantly. The density of colonial Microcystis in the middle layer was not influenced in B treatments and P treatments. The lysis of the colonial Microcystis was only detected in the lower layer of B treatments and P treatments during the short coexisting period. The dissolved oxygen of the lower layer was much lower than that of the middle layer and the upper layer due to the vertical gradient distribution of colonial Microcystis, but Microcystis was proved to be able to tolerate the dark and anoxic environment [23]. Based on the nitrogen and phosphorus concentration in our experiment, nutrients’ limitation for Microcystis were not realized by the algicidal bacteria. Therefore, it was speculated that the inhibition of Microcystis in the lower layer and the release of ammonia nitrogen and phosphorus were mainly due to the algicidal effect of the two bacteria.
Many researches indicated that the ratio of algicidal bacteria to cyanobacteria was set at least 1:1 for the effective suppression or lysis of cyanobacteria [22,24,25]. The cell ratio of each bacteria to cyanobacteria initially added in this experiment has exceeded 25:1. After three days of coexistence experiments, the ratio of bacteria to cyanobacteria in the upper layer was further elevated both in B treatments and P treatments. However, the algicidal effect of the two bacteria did not appear in the upper layer. It was reported that the Microcystis colonies larger than 180 μm made a major contribution to the formation of surface bloom scum, while the colonies smaller than 100 μm made a rather limited contribution to the surface scum [26]. Colony size was proved to be correlated with extracellular polysaccharides (EPS) levels. The presence of higher EPS in the population was also found to protect Microcystis cells against chemical stress [27,28]. Our preliminary experiments showed that the cell-free filtrates from these two bacteria had algicidal activity on M. aeruginosa (Table S1, Supplementary Materials). It indicated that the two bacteria we used could secrete algicidal substances. 1-methyl-β-carboline have been identified as a algicidal compound in Pseudomonas [29]. However, the amount of algicidal substances usually needed to be large enough to meet the effective threshold for bloom control [6]. Therefore, the large-sized Microcystis colonies in the upper layer were likely to be less susceptible to algicidal bacteria. Moreover, some studies suggested that the production of algicidal substances was stimulated under the condition of limited growth, instead of in optimum growth conditions [30,31]. Excessive photosynthetic products could be released into the water by cyanobacteria as an overflow mechanism, which were also important carbon sources and energy sources for heterotrophic bacteria [32,33]. Our data showed that high concentrations of cyanobacteria in the upper layer indeed created a high DOC environment which was conducive to bacterial growth. Furthermore, bacterial quorum-sensing system was found to control algicide production [34]. More interestingly, bacteria could switch from algicidal mode to symbiotic mode based on nutrient acquisition and bacterial density [35]. Just like the bacteria we used, it was reported for its algicidal effect [21,36], but it was also reported that it has the effect to promote nutrient exchange with cyanobacteria and further promote cyanobacteria growth [37]. These opposite phenomena were somewhat similar to the different growth of cyanobacteria in different water layers during the coexistence with algicidal bacteria in our results. However, the key points of the transformation between algicidal mode and symbiotic mode were still unclear. It was likely to increase the uncertainty of the large-scale application of algicidal bacteria.
For a long time, the research on the relationship between algicidal bacteria and cyanobacteria has mainly focused on the one-sided algicidal effect of bacteria on cyanobacteria [11,38]. However, it was also found in some reports that the growth of algicidal bacteria was inhibited by cyanobacteria [39,40]. Further data on the differential gene expression confirmed that algicidal bacteria also suffered from cell damage and oxidative damage caused by cyanobacteria [24,41]. Our results showed that the number of Microcystis in the middle layer was not affected by the two algicidal bacteria, but the ratios of each bacteria to cyanobacteria in the middle layer were significantly lower than that in the upper and lower layers. Moreover, the release of ammonia nitrogen and phosphorus in the middle layer was elevated. It was speculated that the bacteria in the middle layer of the water column might be under certain stress. Whether this stress was caused by carbon limitation or allelopathic stress needs to be further studied.
Aerobic denitrifying bacteria was considered to have application potential for bioremediation of eutrophic water bodies due to their nitrogen removal ability under aerobic conditions [42]. The two bacteria we used have the ability of aerobic denitrification, which was consistent with the previous reports [43,44]. Our results showed that P. stutzeri and B. diminuta both greatly promoted the removal of dissolved nitrogen in the cyanobacteria blooming waters rich in dissolved oxygen. P. stutzeri was good at remove NO3−-N, NH4+-N and DTN. B. diminuta had good removal performance at NO3−-N and DTN. Nitrite reductase was found sensitive to oxygen [45] and high oxygen environment was detected to easily lead to the accumulation of nitrite [46]. Our data showed that B. diminuta exhibited nitrite accumulation under high oxygen conditions, while P. stutzeri exhibited nitrite accumulation under anoxic conditions. Moreover, some potential risks should be paid attention to. Firstly, the number of cyanobacteria in surface water could further be increased under the influence of these bacteria. Once the cyanobacteria population declined, the release of nutrients and microcystins from cyanobacteria would not be ignored. Secondly, according to the previous report [47], the concentrations of TP showed an increase at algae-rich stations. In our experiment, the soluble phosphorus, especially in the upper layer of cyanobacteria blooming water increased under the coexistence with these bacteria, which might be primarily related to the high density of cyanobacteria. Cyanobacteria could release phosphorus in the whole life cycle, especially in the stationary and declining phase [37]. The coexistence of low concentrations of cyanobacteria and bacteria was shown to contribute to the removal of phosphorus from water [48]. Therefore, it was recommended that in order to better use denitrifying bacteria to improve water quality in cyanobacteria blooming waters, the application time of bacteria should be paid attention to, and the combination with mechanized salvage measures of cyanobacteria should also be considered.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w14132129/s1, Table S1: Algicidal effects of bacterial LB cultures and cell-free filtrates against unicellular M. aeruginosa.
Author Contributions
Conceptualization, Q.Z. and Z.Z.; methodology, Q.Z. and Y.Z.; formal analysis, Q.Z. and Y.W.; investigation, Q.Z. and Y.W.; resources, X.W.; data curation, Q.Z. and Y.W.; writing—original draft preparation, Q.Z.; writing—review and editing, Q.Z. and Z.Z.; visualization, Q.Z. and X.W.; supervision, Z.Z.; project administration, H.L.; funding acquisition, Q.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Jiangsu Agriculture Science and Technology Innovation Fund, grant number CX(20)2027 and the National Natural Science Foundation of China, grant number 31800426.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting the findings of this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Amorim, C.A.; Moura, A. Ecological impacts of freshwater algal blooms on water quality, plankton biodiversity, structure, and ecosystem functioning. Sci. Total Environ. 2021, 758, 143605. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Qin, B.; Brookes, J.D.; Shi, K.; Zhu, G.; Zhu, M.; Yan, W.; Zhen, W. The influence of changes in wind patterns on the areal extension of surface cyanobacterial blooms in a large shallow lake in China. Sci. Total Environ. 2015, 518–519, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Qin, B. The changing environment of Lake Taihu and its ecosystem responses. J. Freshw. Ecol. 2015, 30, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Menezes, C.; Churro, C.; Dias, E. Risk levels of toxic cyanobacteria in Portuguese recreational freshwaters. Toxins 2017, 9, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, G.; Zhang, M.; Gao, P.; Wang, J.; Sun, D. Algicidal efficiency and genotoxic effects of Phanerochaete chrysosporium against Microcystis aeruginosa. Int. J. Env. Res. Public Health 2020, 17, 4029. [Google Scholar] [CrossRef]
- Yang, C.; Hou, X.; Wu, D.; Chang, W.; Zhang, X.; Dai, X.; Du, H.; Zhang, X.; Igarashi, Y.; Luo, F. The characteristics and algicidal mechanisms of cyanobactericidal bacteria, a review. World J. Microbiol. Biotechnol. 2020, 36, 188. [Google Scholar] [CrossRef]
- Bi, X.; Dai, W.; Wang, X.; Dong, S.; Zhang, S.; Zhang, D.; Shi, H. Effects of Bacillus subtilis on the growth, colony maintenance, and attached bacterial community composition of colonial cyanobacteria. Environ. Sci. Pollut. Res. 2019, 26, 14977–14987. [Google Scholar] [CrossRef]
- Imai, I.; Inaba, N.; Yamamoto, K. Harmful algal blooms and environmentally friendly control strategies in Japan. Fish. Sci. 2021, 87, 437–464. [Google Scholar] [CrossRef]
- Van Le, V.; Ko, S.R.; Kang, M.; Lee, S.A.; Oh, H.M.; Ahn, C.Y. Algicide capacity of Paucibacter aquatile DH15 on Microcystis aeruginosa by attachment and non-attachment effects. Environ. Pollut. 2022, 302, 119079. [Google Scholar] [CrossRef]
- Kong, Y.; Wang, Y.; Miao, L.; Mo, S.; Li, J.; Zheng, X. Recent advances in the research on the anticyanobacterial effects and biodegradation mechanisms of Microcystis aeruginosa with microorganisms. Microorganisms 2022, 10, 1136. [Google Scholar] [CrossRef]
- Liu, J.; Yang, C.; Chi, Y.; Wu, D.; Dai, X.; Zhang, X.; Igarashi, Y.; Luo, F. Algicidal characterization and mechanism of Bacillus licheniformis Sp34 against Microcystis aeruginosa in Dianchi Lake. J. Basic Microbiol. 2019, 59, 1112–1124. [Google Scholar] [CrossRef]
- Lin, S.; Geng, M.; Liu, X.; Tan, J.; Yang, H. On the control of Microcystis aeruginosa and Synechococccus species using an algicidal bacterium, Stenotrophomonas F6, and its algicidal compounds cyclo-(Gly-Pro) and hydroquinone. J. Appl. Phycol. 2016, 28, 345–355. [Google Scholar] [CrossRef]
- Yu, Y.; Zeng, Y.; Li, J.; Yang, C.; Zhang, X.; Luo, F.; Dai, X. An algicidal Streptomyces amritsarensis strain against Microcystis aeruginosa strongly inhibits microcystin synthesis simultaneously. Sci. Total Environ. 2019, 650, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.C.; Wu, S.C.; Chen, W.C.; Chou, C.C. Model simulation of diurnal vertical migration patterns of different-sized colonies of Microcystis employing a particle trajectory approach. Environ. Eng. Sci. 2013, 30, 179–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziallas, C.; Grossart, H.P. Microbial interactions with the cyanobacterium Microcystis aeruginosa and their dependence on temperature. Mar. Biol. 2012, 159, 2389–2398. [Google Scholar] [CrossRef]
- Su, J.; Shao, S.; Huang, T.; Ma, F.; Lu, J.; Zhang, K. Algicidal effects and denitrification activities of Acinetobacter sp. J25 against Microcystis aeruginosa. J. Environ. Chem. Eng. 2016, 4, 1002–1007. [Google Scholar] [CrossRef]
- Bosse, K.R.; Sayers, M.J.; Shuchman, R.A.; Fahnenstiel, G.L.; Ruberg, S.A.; Fanslow, D.L.; Stuart, D.G.; Johengen, T.H.; Burtner, A.M. Spatial-temporal variability of in situ cyanobacteria vertical structure in Western Lake Erie: Implications for remote sensing observations. J. Great Lakes Res. 2019, 45, 480–489. [Google Scholar] [CrossRef]
- Moore, T.S.; Churnside, J.H.; Sullivan, J.M.; Twardowski, M.S.; Nayak, A.R.; Mcfarland, M.N.; Stockley, N.D.; Gould, R.W.; Johengen, T.H.; Ruberg, S.A. Vertical distributions of blooming cyanobacteria populations in a freshwater lake from LIDAR observations. Remote Sens. Environ. 2019, 225, 347–367. [Google Scholar] [CrossRef]
- Sun, R.; Sun, P.; Zhang, J.; Esquivel-Elizondo, S.; Wu, Y. Microorganisms-based methods for harmful algal blooms control: A review. Bioresour. Technol. 2018, 248, 12–20. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Huang, J.; Fan, Q.; Wei, J.; Wang, F.; Jia, Z.; Xiang, W.; Liang, W. Inhibition of Microcystis aeruginosa using Brevundimonas sp. AA06 immobilized in polyvinyl alcohol-sodium alginate beads. Desalination Water Treat. 2018, 111, 192–200. [Google Scholar] [CrossRef]
- Zhou, S.; Yin, H.; Tang, S.; Peng, H.; Yin, D.; Yang, Y.; Liu, Z.; Dang, Z. Physiological responses of Microcystis aeruginosa against the algicidal bacterium Pseudomonas aeruginosa. Ecotoxicol. Environ. Saf. 2016, 127, 214–221. [Google Scholar] [CrossRef]
- Gumbo, J.R.; Ross, G.; Cloete, T.E. The isolation and identification of predatory bacteria from a Microcystis algal bloom. Afr. J. Biotechnol. 2010, 9, 663–671. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Huang, Y.; Chen, G.; Li, P.; Shen, Y.; Davis, T.W. The secretion of organics by living Microcystis under the dark/anoxic condition and its enhancing effect on nitrate removal. Chemosphere 2018, 196, 280–287. [Google Scholar] [CrossRef]
- Ndlela, L.L.; Oberholster, P.J.; Van Wyk, J.H.; Cheng, P.H. Bacteria as biological control agents of freshwater cyanobacteria: Is it feasible beyond the laboratory? Appl. Microbiol. Biotchnol. 2018, 102, 9911–9923. [Google Scholar] [CrossRef] [PubMed]
- Mayali, X.; Doucette, G.J. Microbial community interactions and population dynamics of an algicidal bacterium active against Karenia brevis (Dinophyceae). Harmful Algae 2002, 1, 277–293. [Google Scholar] [CrossRef]
- Wu, X.; Yang, T.; Feng, S.; Li, L.; Xiao, B.; Song, L.; Sukenik, A.; Ostrovsky, I. Recovery of Microcystis surface scum following a mixing event: Insights from a tank experiment. Sci. Total Environ. 2020, 728, 138727. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Nkrumah, P.N.; Peng, Q. Different tolerances to chemical contaminants between unicellular and colonial morph of Microcystis aeruginosa: Excluding the differences among different strains. J. Hazard. Mater. 2015, 285, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony formation in the cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodani, S.; Imoto, A.; Mitsutani, A.; Murakami, M. Isolation and identification of the antialgal compound, harmane (1-methyl-β-carboline) produced by the antialgal bacterium, Pseudomonas sp. K44–1. J. Appl. Phycol. 2002, 14, 109–114. [Google Scholar] [CrossRef]
- Ray, S.; Bagchi, S.N. Nutrients and pH regulate algicide accumulation in cultures of the cyanobacterium Oscillatoria laetevirens. New Phytol. 2001, 149, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Granéli, E.; Weberg, M.; Salomon, P.S. Harmful algal blooms of allelopathic microalgal species: The role of eutrophication. Harmful Algae 2008, 8, 94–102. [Google Scholar] [CrossRef]
- Wyatt, K.H.; Turetsky, M.R.; Rober, A.R.; Giroldo, D.; Kane, E.S.; Stevenson, R.J. Contributions of algae to GPP and DOC production in an Alaskan fen: Effects of historical water table manipulations on ecosystem responses to a natural flood. Oecologia 2012, 169, 821–832. [Google Scholar] [CrossRef]
- Wyatt, K.H.; Turetsky, M.R. Algae alleviate carbon limitation of heterotrophic bacteria in a boreal peatland. J. Ecol. 2015, 103, 1165–1171. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Guo, X.; Liu, X.; Yang, H. NprR-NprX Quorum-sensing system Regulates the algicidal activity of Bacillus sp. strain S51107 against bloom-forming cyanobacterium Microcystis aeruginosa. Front. Microbiol. 2017, 8, 1968. [Google Scholar] [CrossRef] [PubMed]
- Meyer, N.; Bigalke, A.; Kaulfuß, A.; Pohnert, G. Strategies and ecological roles of algicidal bacteria. FEMS Microbiol. Rev. 2017, 41, 880–899. [Google Scholar] [CrossRef] [Green Version]
- Berthold, D.E.; Shetty, K.G.; Jayachandran, K.; Laughinghouse IV, H.D.; Gantar, M. Enhancing algal biomass and lipid production through bacterial co-culture. Biomass Bioenergy 2019, 122, 280–289. [Google Scholar] [CrossRef]
- Jiang, L.; Yang, L.; Xiao, L.; Shi, X.; Gao, G.; Qin, B. Quantitative studies on phosphorus transference occuring between Microcystis aeruginosa and its attached bacterium (Pseudomonas sp.). Hydrobiologia 2007, 581, 161–165. [Google Scholar] [CrossRef]
- Yang, K.; Chen, Q.; Zhang, D.; Zhang, H.; Lei, X.; Chen, Z.; Li, Y.; Hong, Y.; Ma, X.; Zheng, W.; et al. The algicidal mechanism of prodigiosin from Hahella sp. KA22 against Microcystis aeruginosa. Sci. Rep. 2017, 7, 7750. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Pan, J.; Li, Z.; Liu, X.; Tan, J.; Yang, H. Characterization of an algicidal bacterium Brevundimonas J4 and chemical defense of Synechococcus sp. BN60 against bacterium J4. Harmful Algae 2014, 37, 1–7. [Google Scholar] [CrossRef]
- Shao, J.; Jiang, Y.; Wang, Z.; Peng, L.; Luo, S.; Gu, J.; Li, R. Interactions between algicidal bacteria and the cyanobacterium Microcystis aeruginosa: Lytic characteristics and physiological responses in the cyanobacteria. Int. J. Environ. Sci. Technol. 2014, 11, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Osman, O.A.; Beier, S.; Grabherr, M.; Bertilsson, S. Freshwater cyanobacterial interactions with bacterial antagonists. Appl. Environ. Microbiol. 2017, 83, e02634-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Ren, Y.X.; Zhao, S.Q.; Liang, X.; Wang, J.P. Isolation and characterization of three heterotrophic nitrifying-aerobic denitrifying bacteria from a sequencing batch reactor. Ann. Microbiol. 2016, 66, 737–747. [Google Scholar] [CrossRef]
- Agustiyani, D.; dan Sarjiya Antonius, N.L. Physiological growth, potential activity of N2O production and their functional gene of some isolate of denitrifying bacteria. Ber. Biol. 2018, 17, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Qing, H.; Donde, O.O.; Tian, C.; Wang, C.; Wu, X.; Feng, S.; Liu, Y.; Xiao, B. Novel heterotrophic nitrogen removal and assimilation characteristic of the newly isolated bacterium Pseudomonas stutzeri AD-1. J. Biosci. Bioeng. 2018, 126, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; He, J.; Chen, M.; Lin, L.; Zhang, F. Factors dominating bacterioplankton abundance and production in the Nordic seas and the Chukchi Sea in summer 2012. Acta Oceanol. Sin. 2017, 36, 153–162. [Google Scholar] [CrossRef]
- Sun, Y.; Li, A.; Zhang, X.; Ma, F. Regulation of dissolved oxygen from accumulated nitrite during the heterotrophic nitrification and aerobic denitrification of Pseudomonas stutzeri T13. Appl. Microbiol. Biotechnol. 2015, 99, 3243–3248. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Lu, T.; Song, H.; Lavoie, M.; Xu, J.; Fan, X.; Pan, X. Spatial variability of cyanobacteria and heterotrophic bacteria in Lake Taihu (China). Bull. Environ. Contam. Toxicol. 2017, 99, 380–384. [Google Scholar] [CrossRef]
- Ji, X.; Jiang, M.; Zhang, J.; Jiang, X.; Zheng, Z. The interactions of algae-bacteria symbiotic system and its effects on nutrients removal from synthetic wastewater. Bioresour. Technol. 2018, 247, 44–50. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The effect of algicidal and denitrifying bacteria on the vertical distribution of M. aeruginosa.
Figure 1.
The effect of algicidal and denitrifying bacteria on the vertical distribution of M. aeruginosa.

Figure 2.
The effect of algicidal and denitrifying bacteria on DO (a) and pH (b) in the different layers of water.
Figure 2.
The effect of algicidal and denitrifying bacteria on DO (a) and pH (b) in the different layers of water.

Figure 3.
The effect of algicidal and denitrifying bacteria on NO3−-N (a), NO2−-N (b), NH4+-N (c) and DTN (d) concentrations in the different layers of water.
Figure 3.
The effect of algicidal and denitrifying bacteria on NO3−-N (a), NO2−-N (b), NH4+-N (c) and DTN (d) concentrations in the different layers of water.

Figure 4.
The effect of algicidal and denitrifying bacteria on PO43−-P (a) and DTP (b) concentrations in the different layers of water.
Figure 4.
The effect of algicidal and denitrifying bacteria on PO43−-P (a) and DTP (b) concentrations in the different layers of water.

Figure 5.
The effect of algicidal and denitrifying bacteria on DOC concentrations in the different layers of water.
Figure 5.
The effect of algicidal and denitrifying bacteria on DOC concentrations in the different layers of water.

Figure 6.
The ratio of bacteria to cyanobacteria in the different layers of water. (a) B. diminuta and (b) P. stutzeri.
Figure 6.
The ratio of bacteria to cyanobacteria in the different layers of water. (a) B. diminuta and (b) P. stutzeri.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Algicidal effects of B. diminuta Z1 and P. stutzeri ZJ4 washed cells against unicellular M. aeruginosa.
Table 1.
Algicidal effects of B. diminuta Z1 and P. stutzeri ZJ4 washed cells against unicellular M. aeruginosa.
| Algicidal Bacteria | Concentration (Cells/mL) | Inhibitory Rate after 3 Days (%) |
|---|---|---|
| B. diminuta Z1 | 1.36 × 108 | 24.29 ± 3.29% |
| P. stutzeri ZJ4 | 1.02 × 108 | 10.07 ± 0.94% |
Note: Initial concentration of unicellular M. aeruginosa (FACHB 912) was 7.18 × 106 cells/mL.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhou, Q.; Wang, Y.; Wen, X.; Liu, H.; Zhang, Y.; Zhang, Z. The Effect of Algicidal and Denitrifying Bacteria on the Vertical Distribution of Cyanobacteria and Nutrients. Water 2022, 14, 2129. https://doi.org/10.3390/w14132129
AMA Style
Zhou Q, Wang Y, Wen X, Liu H, Zhang Y, Zhang Z. The Effect of Algicidal and Denitrifying Bacteria on the Vertical Distribution of Cyanobacteria and Nutrients. Water. 2022; 14(13):2129. https://doi.org/10.3390/w14132129
Chicago/Turabian StyleZhou, Qing, Yan Wang, Xuezheng Wen, Haiqin Liu, Yingying Zhang, and Zhiyong Zhang. 2022. "The Effect of Algicidal and Denitrifying Bacteria on the Vertical Distribution of Cyanobacteria and Nutrients" Water 14, no. 13: 2129. https://doi.org/10.3390/w14132129
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.