
Effects of Protein Level on the Production and Growth Performance of Juvenile Chinese Mitten Crab (Eriocheir sinensis) and Environmental Parameters in Paddy Fields

Abstract
:1. Introduction
2. Materials and Methods
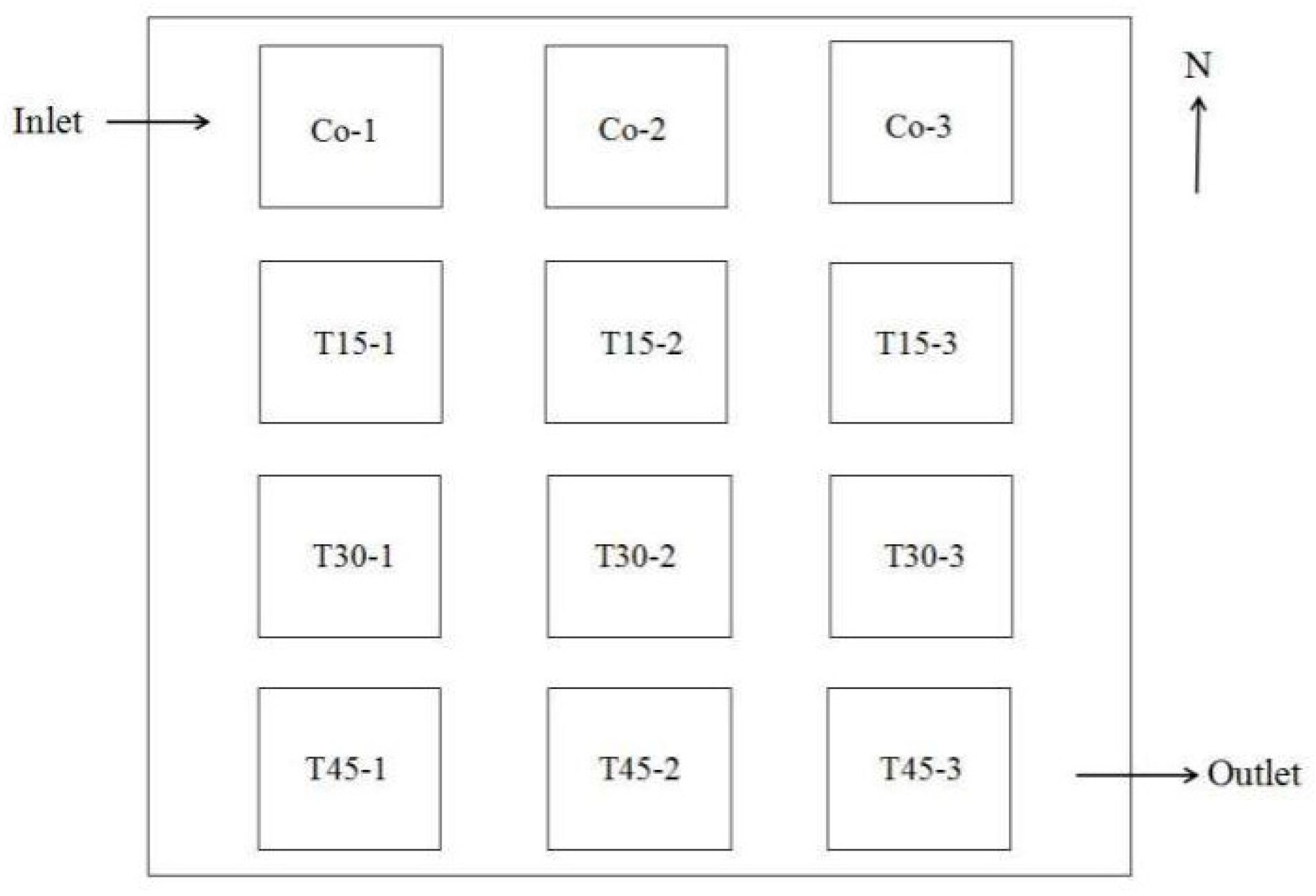
2.1. Experimental Animals and Experimental Paddy Field Management
2.2. Experimental Design
2.3. Test Methods
2.3.1. Monitoring Water Quality in the Paddy Fields
2.3.2. Growth Performance and Yield of Crabs
2.3.3. Qualitative and Quantitative Analysis of Phytoplankton
2.3.4. Qualitative and Quantitative Analyses of Zooplankton
2.3.5. Qualitative and Quantitative Analyses of Aquatic Vascular Plants
2.3.6. Qualitative and Quantitative Analysis of Benthic Animals
2.4. Statistical Analysis
3. Results
3.1. Growth Performance and Yield of Crabs
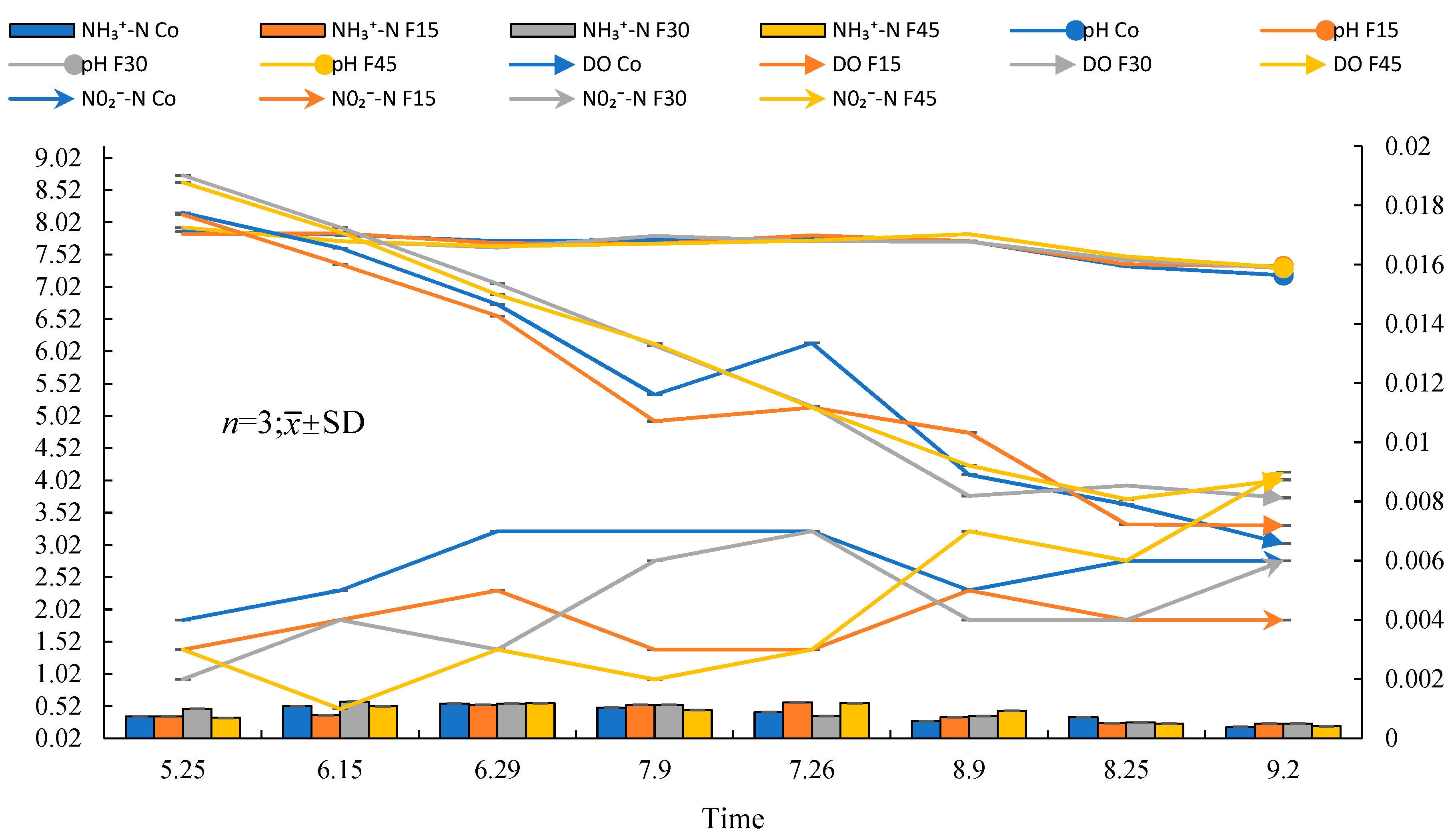
3.2. Water Quality
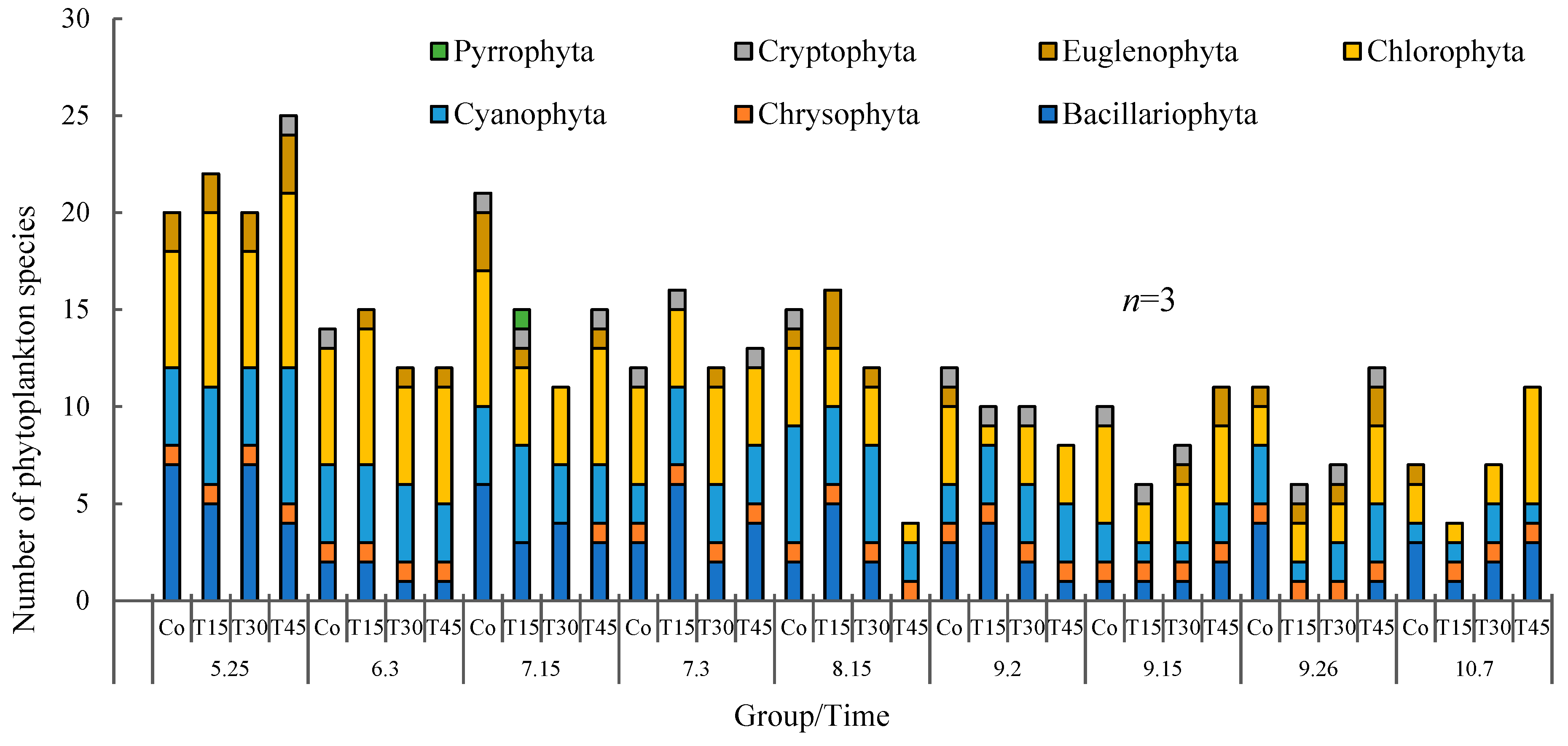
3.3. Phytoplankton in the Paddy Fields and Variations with Time
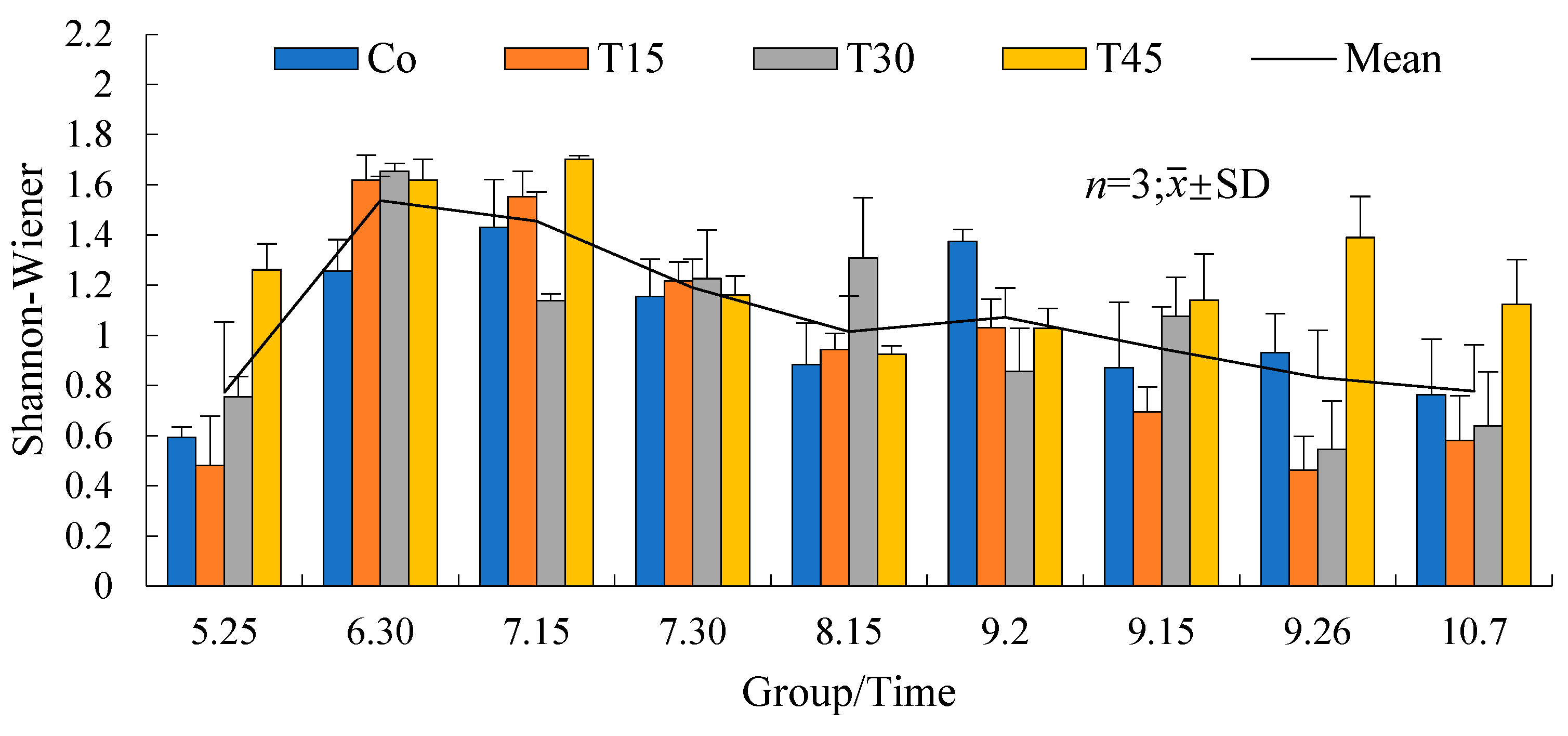
3.3.1. Phytoplankton Biodiversity
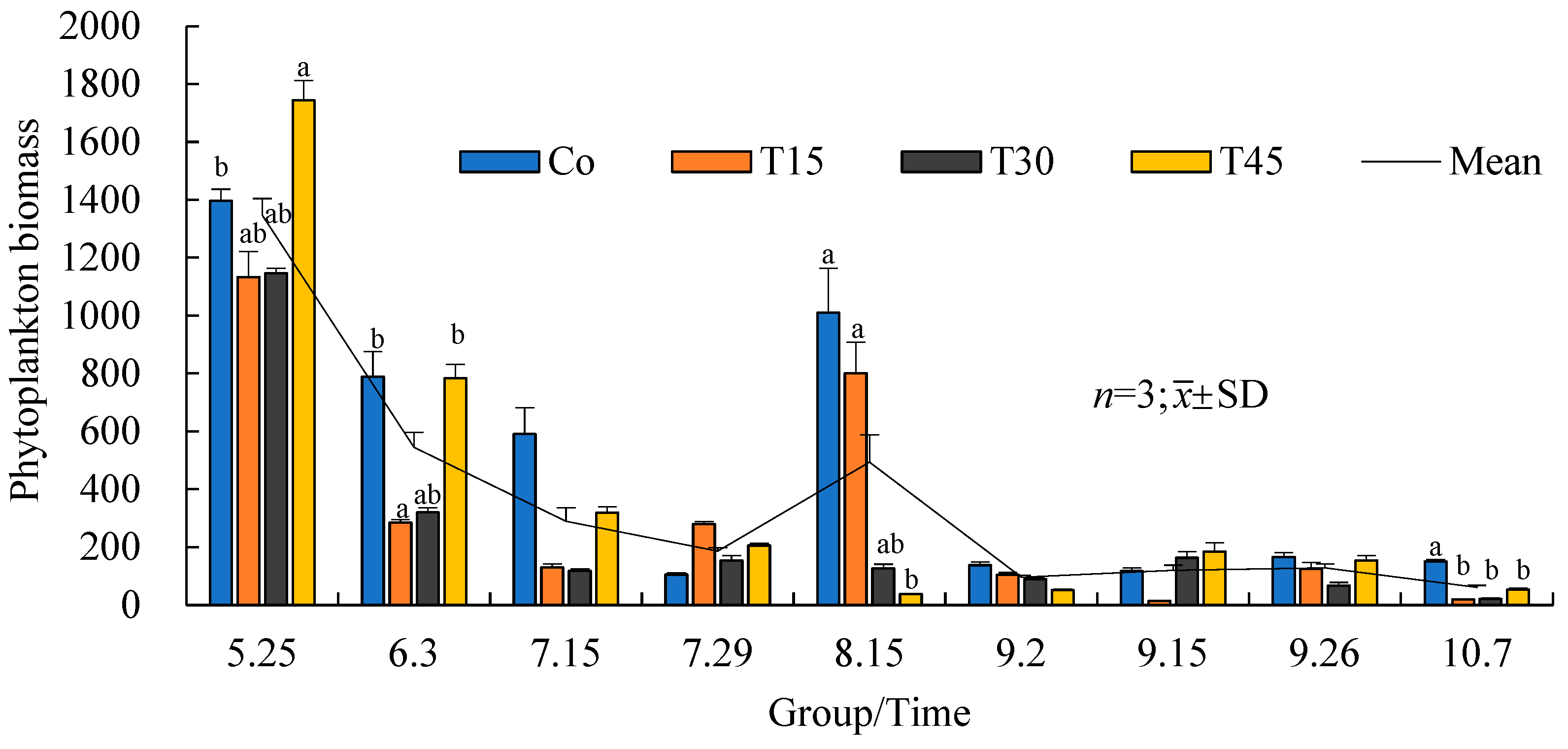
3.3.2. Variation Trends in Phytoplankton Biomass over Time
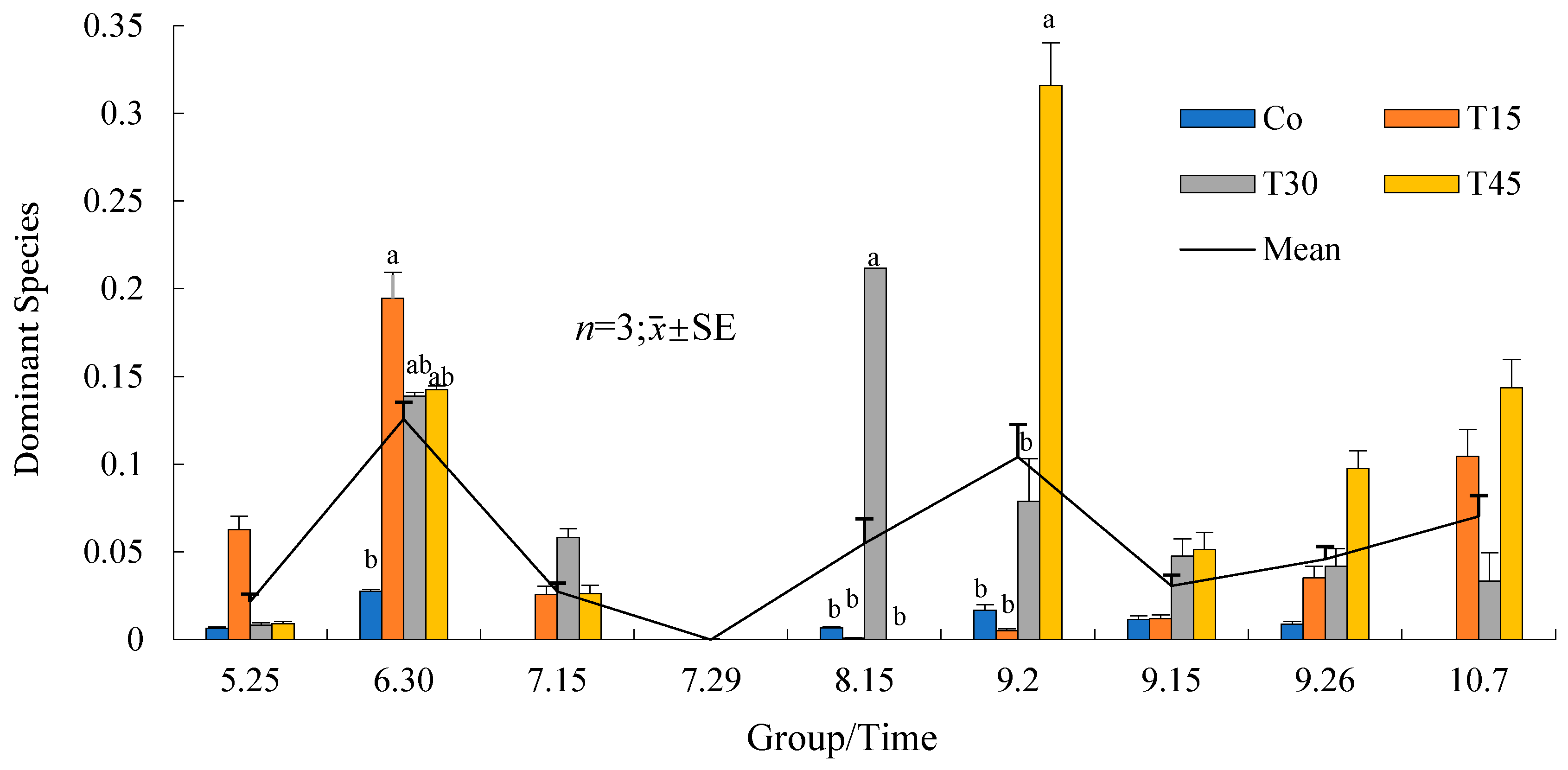
3.3.3. Succession and Population Changes in the Dominant Phytoplankton Species
3.4. Species, Quantities, and Changes in Zooplankton
3.4.1. Zooplankton Species Diversity
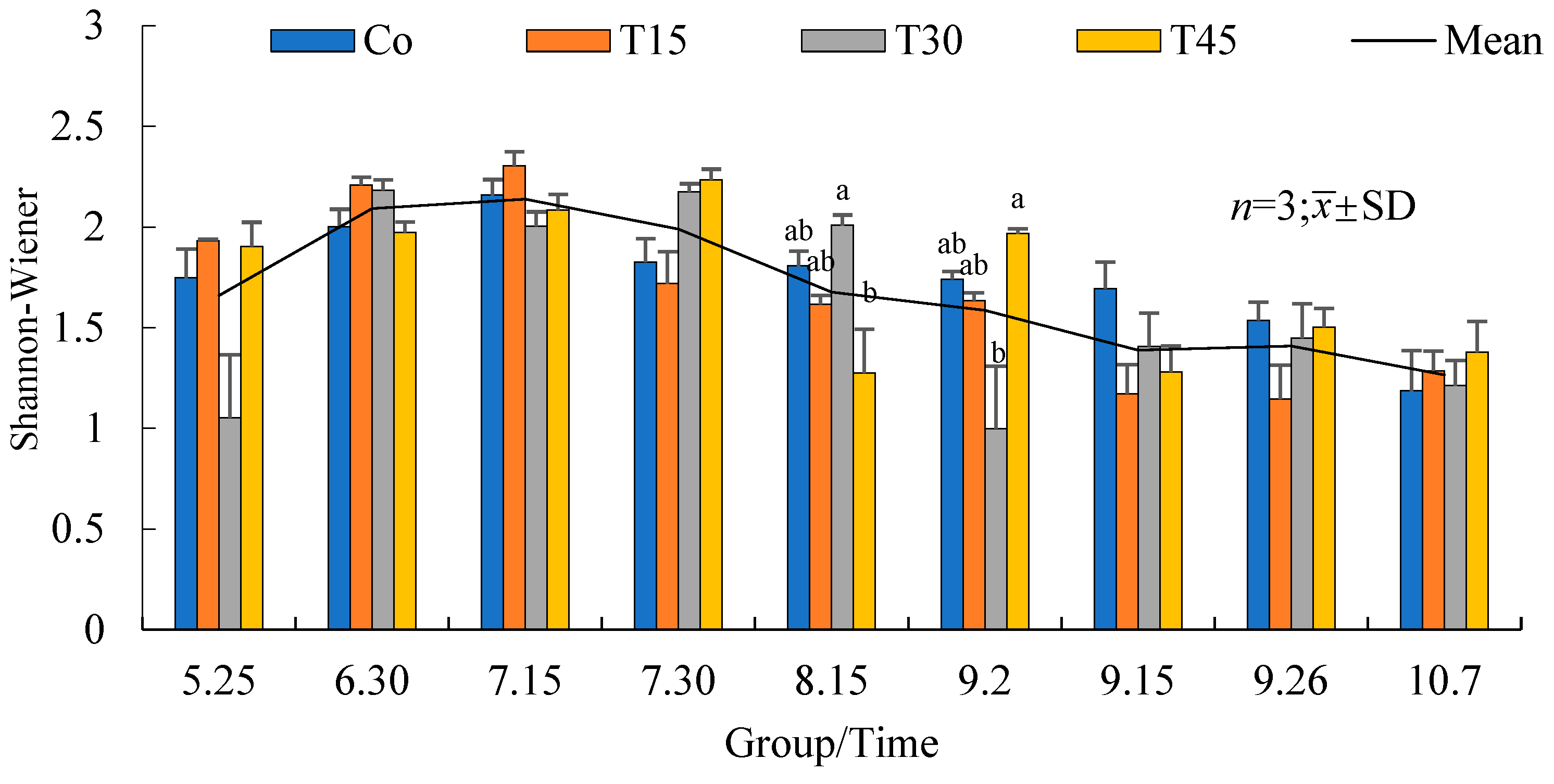
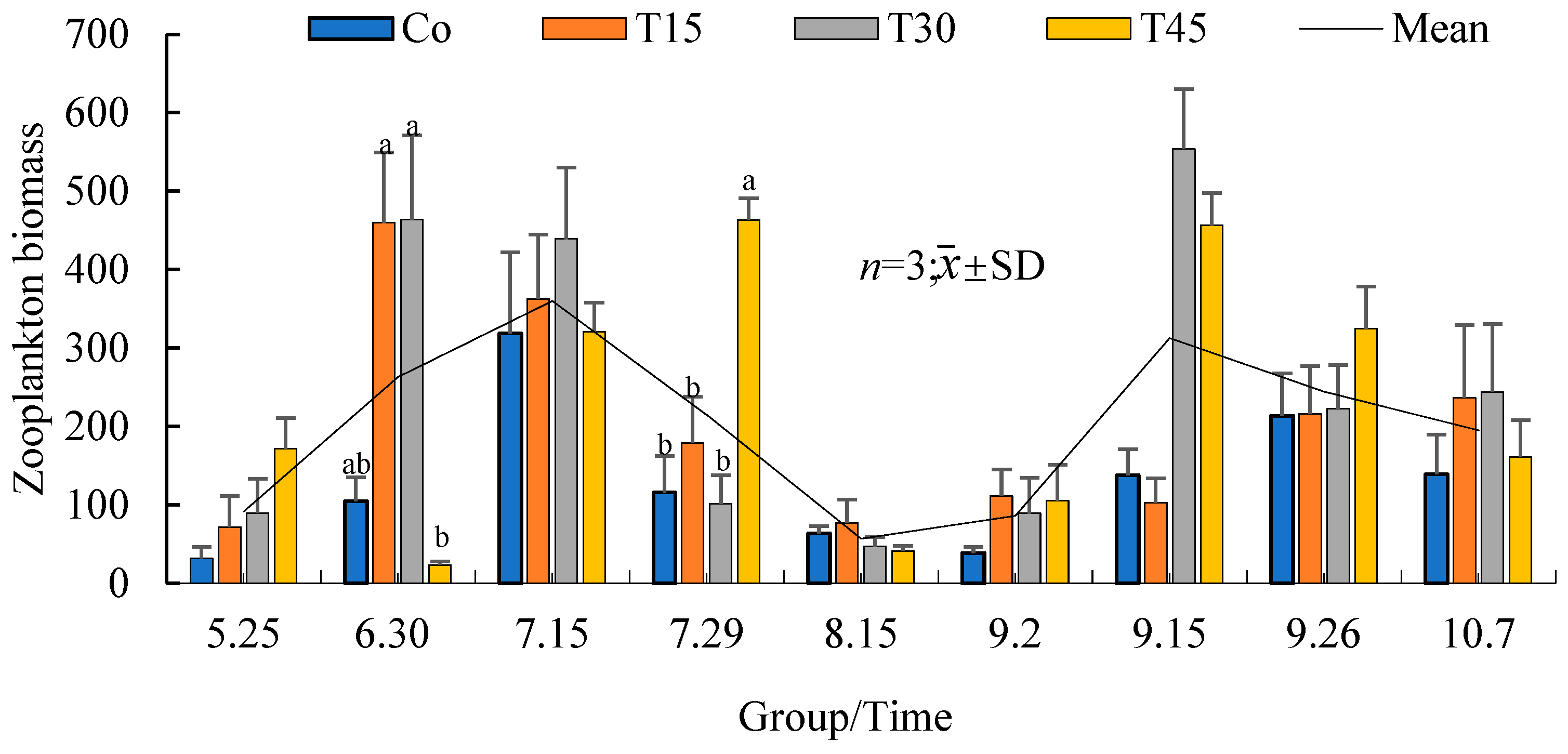
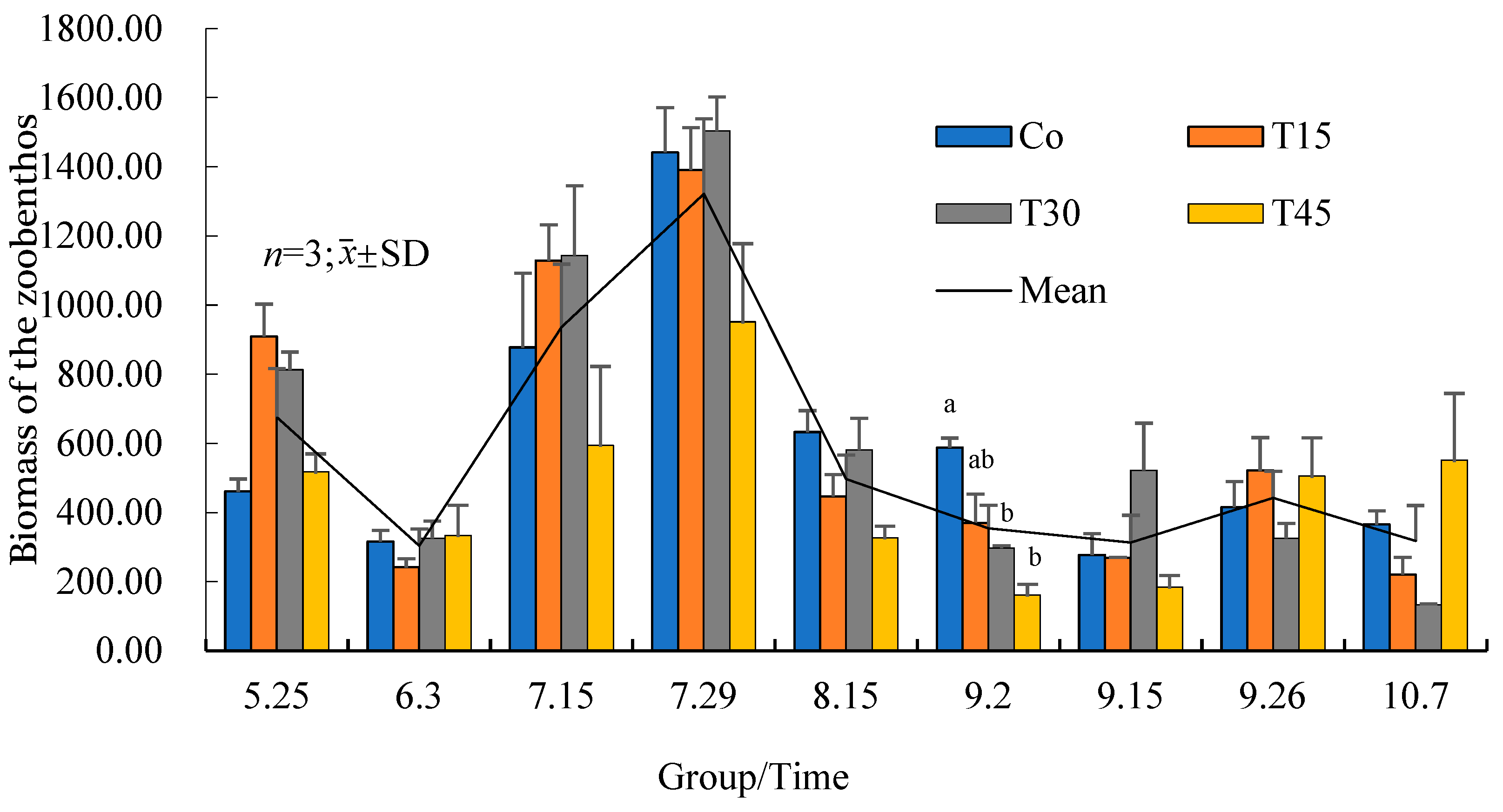
3.4.2. Variation Trends of the Zooplankton Biomass over Time
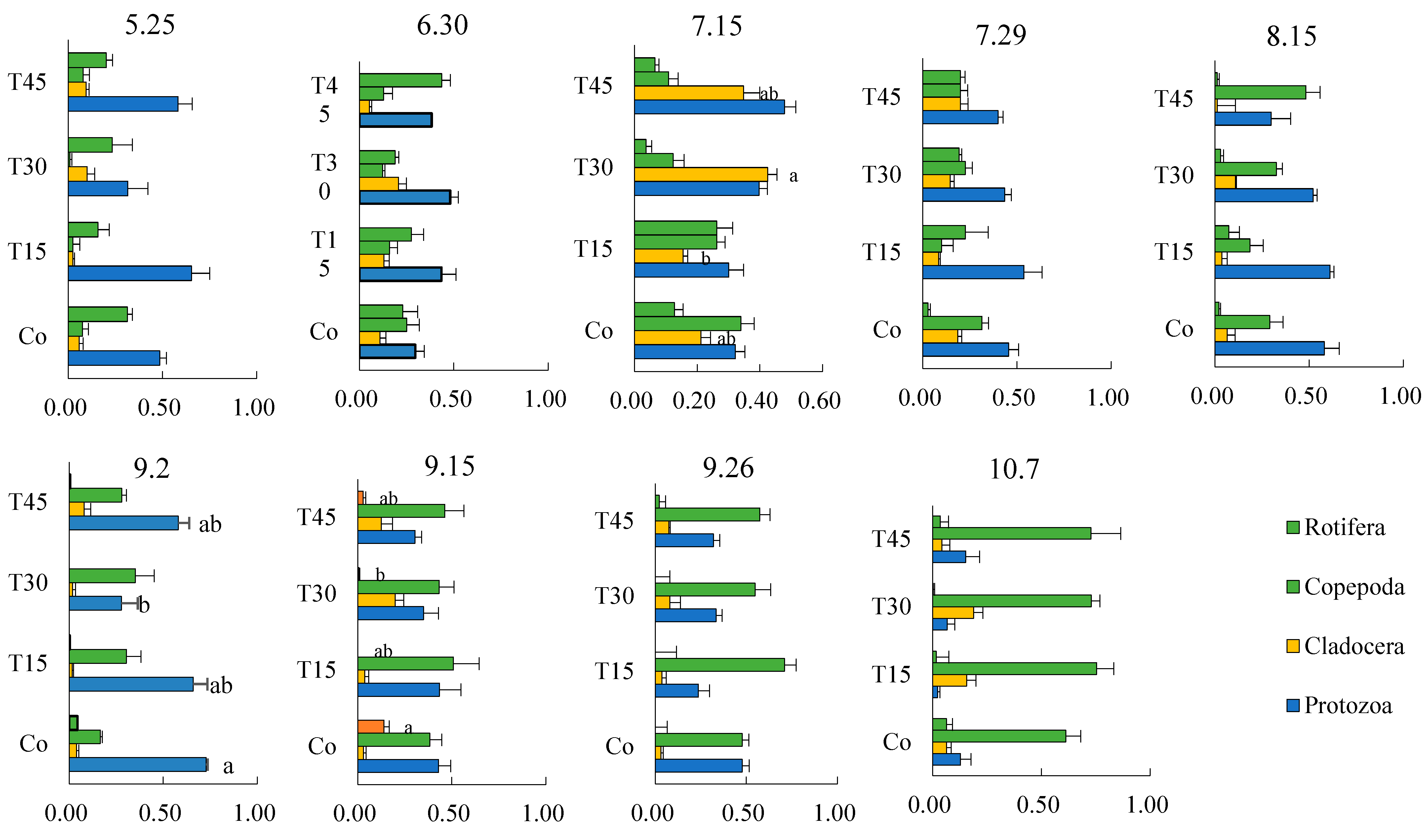
3.4.3. Succession of Dominant Zooplankton Species and Community Changes
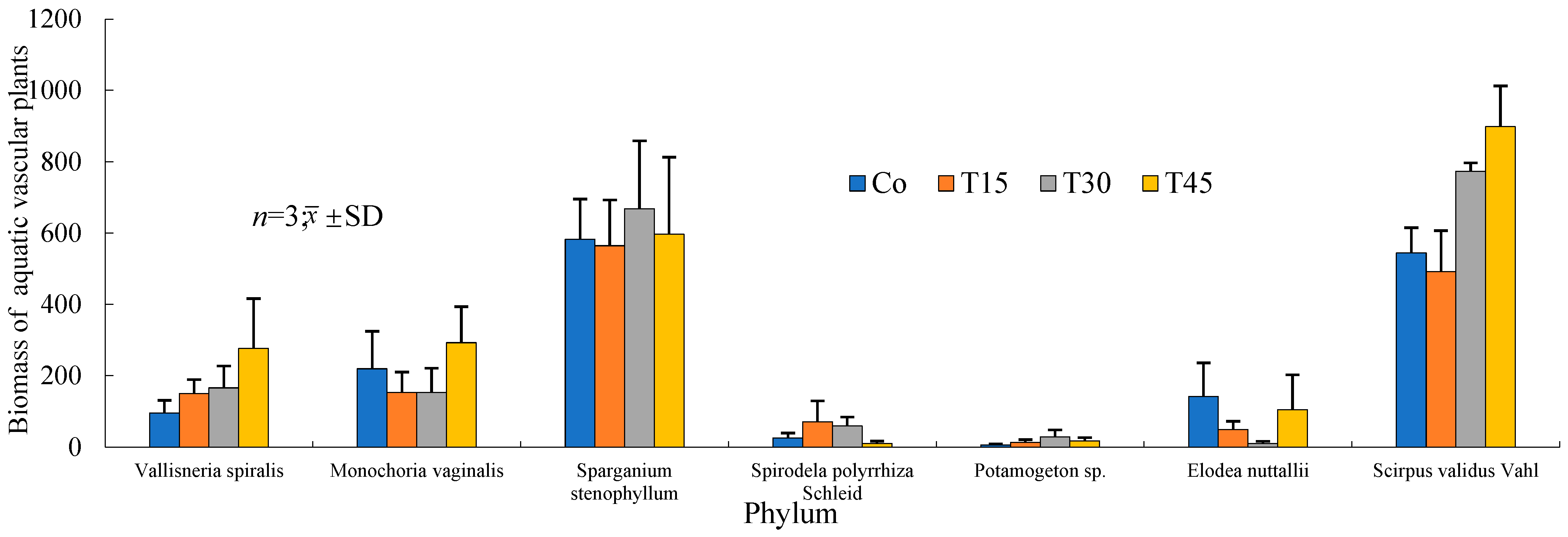
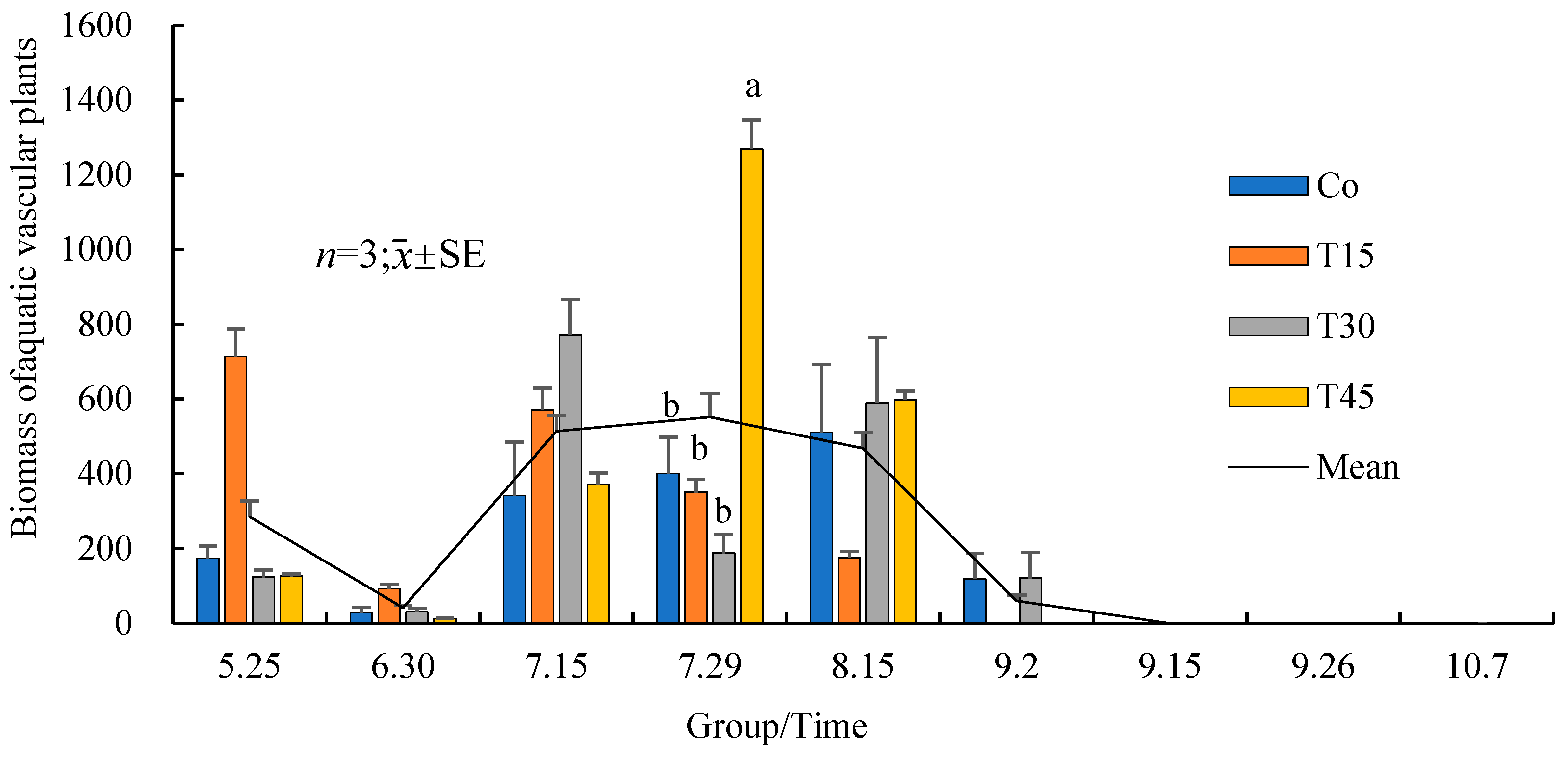
3.5. Species, Quantities, and Changes in Aquatic Vascular Plants
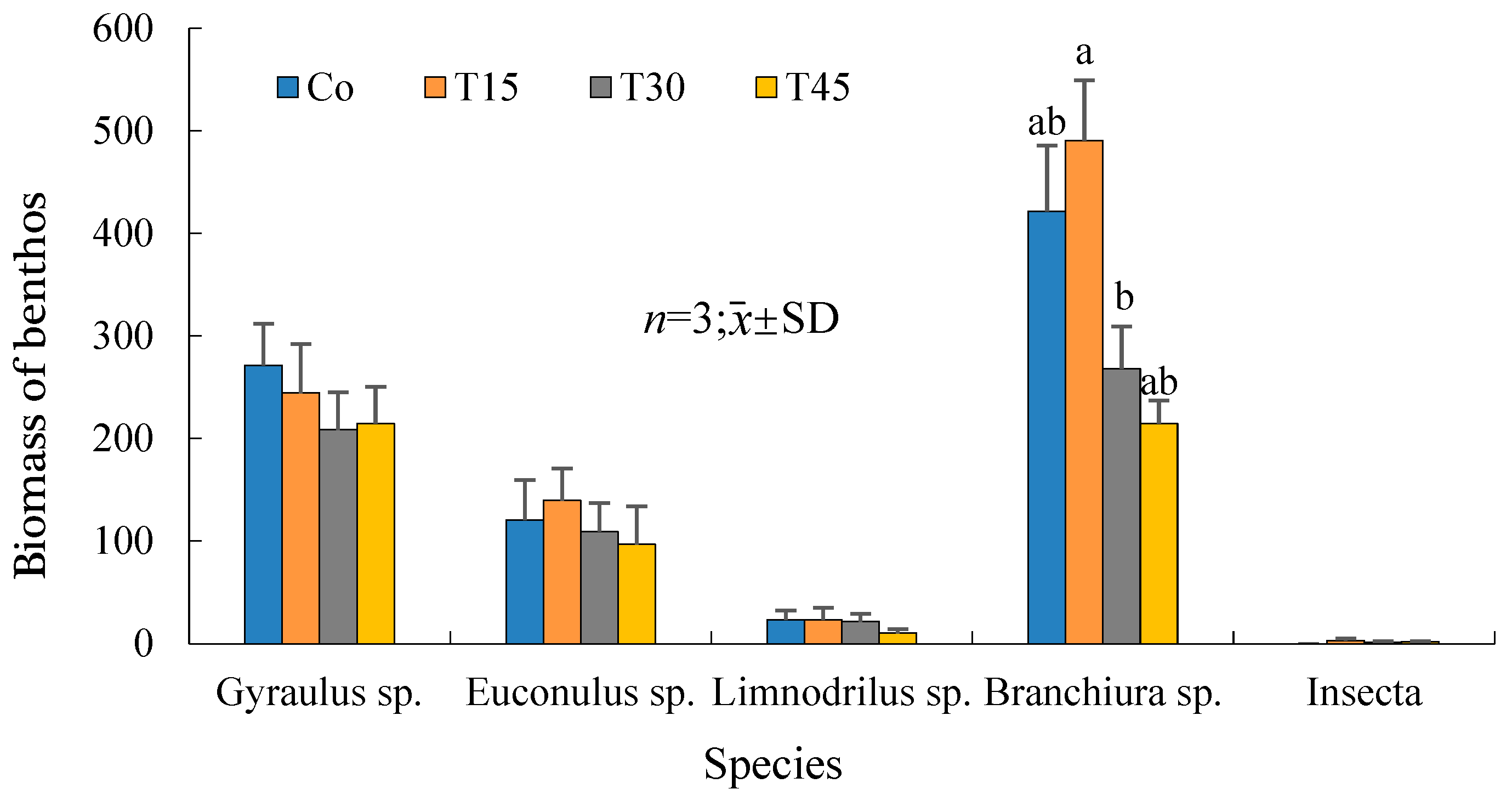
3.6. Variations in the Benthic Animal Species and Quantities with Time
4. Discussion
4.1. Effects of Diets with Different Protein Levels on the Growth Performance and Yield of Juvenile Crabs
4.2. Changes in Physical and Chemical Properties of Paddy Water Environment
4.3. Effects of Diets with Different Protein Levels on the Phytoplankton in Paddy Fields
4.4. Effects of Different Protein Levels in the Crab Diet on Zooplankton in Paddy Fields
4.5. Effects of Crabs on Aquatic Vascular Plants in Paddy Fields
4.6. Changes in Benthic Animals in Rice–Crab Co-Culture
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
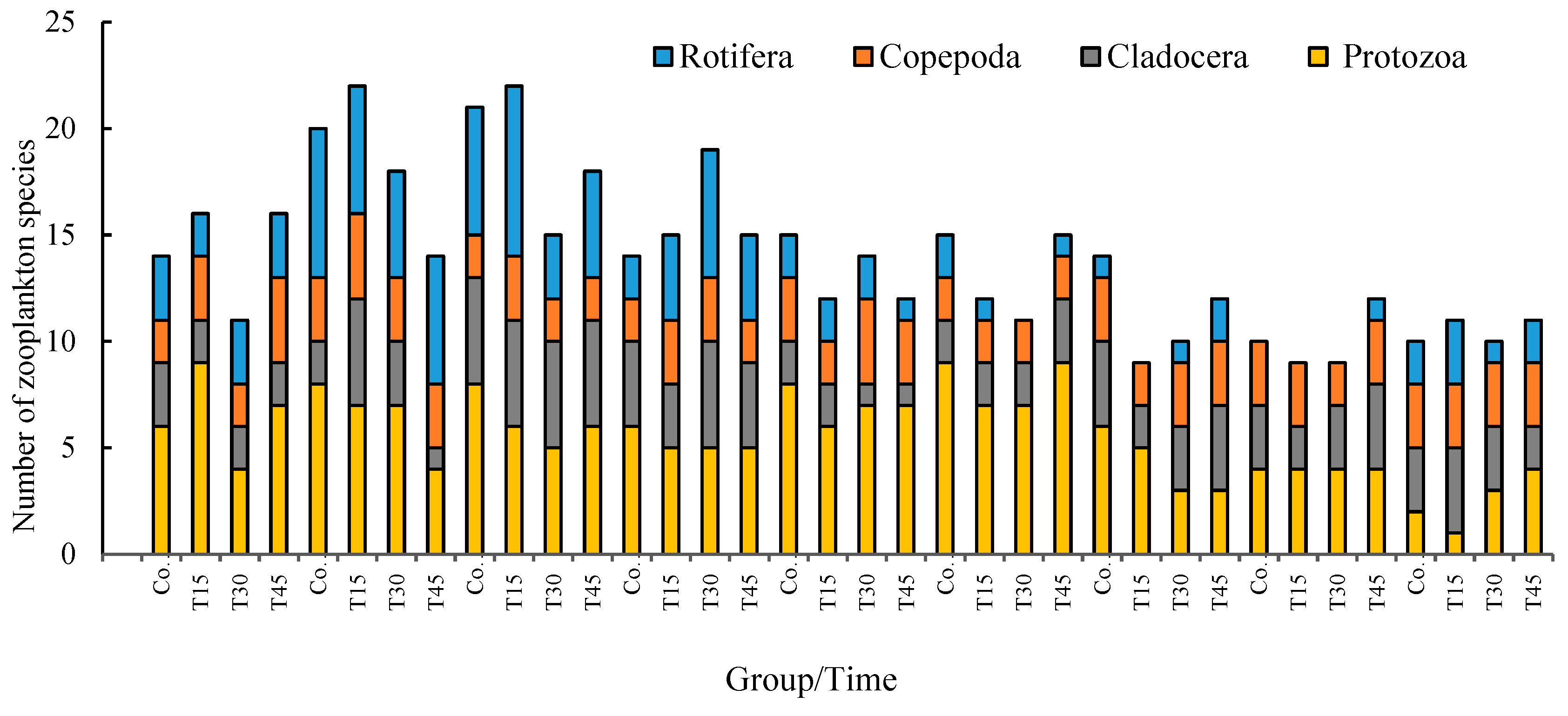{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unit Price/RMB yuan/ton | Cost/RMB yuan/ton | ||
|---|---|---|---|
| T15 | T30 | T45 | |
| 7360 | 442 | 1987 | 3459 |
| 3380 | 101 | 456 | 794 |
| 3920 | 118 | 118 | 118 |
| 2623 | 1904 | 1130 | 396 |
| 15,000 | 1125 | 825 | 525 |
| 11,000 | 55 | 55 | 55 |
| 19,600 | 392 | 392 | 392 |
| 133,000 | 2660 | 2660 | 2660 |
| 30,000 | 300 | 300 | 300 |
| 44,000 | 220 | 220 | 220 |
| 65,600 | 131 | 131 | 131 |
| 7200 | 108 | 108 | 108 |
| 178,000 | 178 | 178 | 178 |
| 1,000,000 | 100 | 100 | 100 |
| 22,900 | 23 | 23 | 23 |
| 7857 | 8684 | 9459 | |
| Species | Average Wet Weight of Cells/mg |
|---|---|
| Bacillariophyta | |
| Nitzschia sp. | 0.003 |
| Cyclotella meneghiniana | 0.00125 |
| Nitzschia frustulum | 0.006 |
| Nitzschia acicularis | 0.005 |
| Synedra sp. | 0.005 |
| Chaetoceros sp. | 0.0014 |
| Navicula amphibola | 0.0017 |
| Navicula placentula | 0.006 |
| Melosira sp. | 0.006 |
| Coscinodisous sp. | 0.02 |
| Fragilaria sp. | 0.001 |
| Gyrosigma sp. | 0.03 |
| Mastogloia sp. | 0.00325 |
| Navicula directa | 0.03 |
| Amphora exigua | 0.0017 |
| Pleurosigma sp. | 0.047 |
| Eunotogramma sp. | 0.001 |
| Navicula sp. | 0.001 |
| Chtysophyta | |
| Chromulina pygmaea Nygaard | 0.00065 |
| Cyanophyta | |
| Chroococcus sp. | 0.0001 |
| Spirulina sp. | 0.0077 |
| Merismopedia sinica | 0.00025 |
| Oscillatoria sp. | 0.01 |
| Microcystis sp. | 0.0016 |
| Nostoc sp. | 0.00025 |
| Anabaena sp. | 0.0005 |
| Merismopedia sp. | 0.00003 |
| Phormidium sp. | 0.002 |
| Chlorophyta | |
| Oocystis borgei | 0.004 |
| Schroederi krosch | 0.003 |
| Closterium sp. | 0.08 |
| Actinastrum lag sp. | 0.001 |
| Chlorella pyrenoidesa | 0.00015 |
| Selenastrum bibraianum | 0.001 |
| Chlamydomonas sp. | 0.01 |
| Pandorina morum | 0.04 |
| Dictyosphaerium sp. | 0.001 |
| Eudorina elegans | 0.02 |
| Tetraedron trilobulatum | 0.003 |
| Crucigenia sp. | 0.001 |
| Kirchneriellalunaris lunatis | 0.001 |
| Platymonas sp. | 0.012 |
| Spirogyra sp. | 0.02 |
| Ankistrodesmus convolutus | 0.002 |
| Pediastrum sp. | 0.01 |
| Sceneclesmus sp. | 0.002 |
| Euglenophyta | |
| Euglena sp. | 0.04 |
| Euglena oxyuris | 0.15 |
| Phacus sp. | 0.06 |
| Euglena viridis | 0.04 |
| Euglena pisciformis | 0.15 |
| Cryptophyta | |
| Cryptomonas sp. | 0.01 |
| Phrrophyta | |
| Gymmodinium sp. | 0.008 |
| Species | Average Wet Weight of Cells/mg |
|---|---|
| Protozoa | |
| Sarcomastigophora | |
| Saccamoeba sp. | 0.00003 |
| Difflugia sp. | 0.00003 |
| Difflugia oblonga | 0.00024 |
| Tintinnidium fliuviatile | 0.00024 |
| Arcella vulgaris | 0.00003 |
| Globigerinoides sp. | 0.00002 |
| Ciliophora | |
| Coleps sp. | 0.00003 |
| Strobilidium sp. | 0.00003 |
| Litonotus sp. | 0.00003 |
| Tintinnopsis sp. | 0.00003 |
| Lembadion sp. | 0.000016 |
| Zoothamnium sp. | 0.0000017 |
| Euplotes sp. | 0.000016 |
| Pseudoprorodon sp. | 0.00005 |
| Didinium nasufum | 0.00045 |
| Actinobolina sp. | 0.00003 |
| Prorodon ovum | 0.00005 |
| Pleuronema sp. | 0.0000017 |
| Stentor polymorphrus | 0.000002 |
| Vorticella sp. | 0.000014 |
| Colpoda sp. | 0.0000017 |
| Trachelius sp. | 0.000007 |
| Rhabdostyla sp. | 0.00005 |
| Cladocera | |
| Daphnia carinata | 0.2 |
| Daphnia magna | 0.01 |
| Moinidae brachiata | 0.1 |
| Moinidae rectirostris | 0.01 |
| Moinidae macrocopa | 0.05 |
| Chydoroidea quadrangula | 0.01 |
| Chydoroidea sphaericus | 0.03 |
| Chydoroidea longirostris | 0.03 |
| Copepoda | |
| Cyclops sp. | 0.03 |
| Copepodid | 0.003 |
| Calanoida sp. | 0.312 |
| Copepod nauplius | 0.003 |
| Rotifera | |
| Brachionus calyciflorus | 0.0025 |
| Brachionus ureus | 0.00024 |
| Brachionus quadridentatus | 0.00055 |
| Polyarthra trigla | 0.000331 |
| Polyarthra sp. | 0.0025 |
| Lecanidae inermis | 0.026 |
| Asplachna brightwelli | 0.0005 |
| Brachionus diversicornis | 0.0005 |
| Rotaria citrine | 0.00028 |
| Filinia sp. | 0.0003 |
| Lepadella ovalis | 0.0003 |
| Pedalia mira | 0.000027 |
| Keratella cochlearis | 0.0003 |
| Keratella valga | 0.00024 |
| Euchlanis pellucida | 0.0025 |
| Species | Biomass/mg·L−1 | |||
|---|---|---|---|---|
| Co | T15 | T30 | T45 | |
| Bacillariophyta | 16.64 ± 6.83 | 15.29 ± 6.16 | 13.46 ± 5.92 | 9.85 ± 5.59 |
| Nitzschia sp. | 1.33 ± 0.79 | 0.78 ± 0.44 | 0.78 ± 0.44 | 0.44 ± 0.36 |
| Cyclotella meneghiniana | 0.14 ± 0.24 | 0.23 ± 0.25 | 0.05 ± 0.08 | 0.00 ± 0.00 |
| Nitzschia frustulum | 1.11 ± 1.09 | 1.56 ± 1.5 | 2.00 ± 1.54 | 1.11 ± 1.09 |
| Nitzschia acicularis | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Synedra sp. | 4.07 ± 3.90 | 1.67 ± 1.01 | 1.11 ± 0.69 | 0.19 ± 0.31 |
| Chaetoceros sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.05 ± 0.09 | 0.05 ± 0.09 |
| Navicula amphibola | 0.25 ± 0.25 | 0.06 ± 0.11 | 0.06 ± 0.11 | 0.00 ± 0.00 |
| Navicula placentula | 0.67 ± 0.83 | 0.22 ± 0.38 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Melosira sp. | 0.44 ± 0.52 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.89 ± 1.51 |
| Coscinodisous sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Fragilaria sp. | 0.11 ± 0.14 | 0.04 ± 0.06 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Gyrosigma sp. | 1.11 ± 1.89 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Mastogloia sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.24 ± 0.41 |
| Navicula directa | 3.89 ± 3.22 | 4.44 ± 3.55 | 7.78 ± 5.83 | 5.56 ± 5.47 |
| Amphora exigua | 0.06 ± 0.11 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Pleurosigma sp. | 0.00 ± 0.00 | 5.22 ± 4.92 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Eunotogramma sp. | 0.78 ± 1.26 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Navicula sp. | 1.63 ± 1.1 | 1.07 ± 0.77 | 1.63 ± 0.93 | 2.00 ± 0.96 |
| Chrysophyta | 47.47 ± 35.67 | 24.46 ± 19.63 | 15.74 ± 11.73 | 19.72 ± 9.42 |
| Chromulina pygmaea Nygaard | 47.47 ± 35.67 | 24.46 ± 19.63 | 15.74 ± 11.73 | 19.72 ± 9.42 |
| Cyanophyta | 16.1 ± 5.87 | 15.23 ± 5.37 | 11.37 ± 4.13 | 17.87 ± 6.72 |
| Chroococcus sp. | 0.25 ± 0.16 | 0.28 ± 0.14 | 0.28 ± 0.15 | 0.76 ± 0.55 |
| Spirulina sp. | 1.03 ± 1.33 | 0.57 ± 0.67 | 0.57 ± 0.67 | 0.29 ± 0.48 |
| Merismopedia sinica | 0.05 ± 0.06 | 0.02 ± 0.03 | 0.01 ± 0.02 | 0.12 ± 0.19 |
| Oscillatoria sp. | 10.00 ± 3.85 | 10.00 ± 4.06 | 7.78 ± 2.92 | 11.11 ± 4.66 |
| Microcystis sp. | 0.53 ± 0.48 | 1.13 ± 0.87 | 0.12 ± 0.14 | 0.47 ± 0.41 |
| Nostoc sp. | 0.17 ± 0.25 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Anabaena sp. | 0.15 ± 0.17 | 0.06 ± 0.09 | 0.02 ± 0.03 | 0.00 ± 0.00 |
| Merismopedia sp. | 0.00 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.00 |
| Phormidium sp. | 3.93 ± 2.11 | 3.19 ± 1.34 | 2.59 ± 1.28 | 5.11 ± 3.35 |
| Chlorophyta | 42.28 ± 23.32 | 24.23 ± 12.36 | 21.59 ± 10.26 | 41.79 ± 23.33 |
| Oocystis borgei | 0.15 ± 0.25 | 0.3 ± 0.35 | 0.59 ± 0.79 | 0.3 ± 0.35 |
| Schroederi krosch | 0.32 ± 0.41 | 0.56 ± 0.47 | 0.00 ± 0.00 | 0.22 ± 0.26 |
| Closterium sp. | 2.96 ± 5.04 | 5.93 ± 6.98 | 2.96 ± 5.04 | 5.93 ± 6.98 |
| Actinastrum lag sp. | 0.00 ± 0.00 | 0.04 ± 0.06 | 0.04 ± 0.06 | 0.04 ± 0.06 |
| Chlorella pyrenoidesa | 7.25 ± 5.09 | 4.71 ± 2.34 | 5.56 ± 3.48 | 5.12 ± 2.42 |
| Selenastrum bibraianum | 0.63 ± 0.42 | 0.48 ± 0.26 | 0.78 ± 0.62 | 0.89 ± 0.83 |
| Chlamydomonas sp. | 14 ± 19.46 | 8.15 ± 7.70 | 1.11 ± 1.39 | 19.20 ± 2.62 |
| Pandorina morum | 4.44 ± 5.54 | 1.48 ± 2.52 | 7.41 ± 5.18 | 3.07 ± 0.15 |
| Dictyosphaerium sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.04 ± 0.06 | 0.04 ± 0.06 |
| Eudorina elegans | 8.89 ± 8.57 | 1.48 ± 1.75 | 1.48 ± 1.75 | 2.59 ± 2.81 |
| Tetraedron trilobulatum | 0.22 ± 0.26 | 0.22 ± 0.26 | 0.00 ± 0.00 | 0.33 ± 0.42 |
| Crucigenia sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.37 ± 0.63 | 0.37 ± 0.33 |
| Kirchneriellalunaris lunatis | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.04 ± 0.06 |
| Platymonas sp. | 0.89 ± 1.05 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Spirogyra sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.74 ± 1.26 |
| Ankistrodesmus convolutus | 0.67 ± 0.68 | 0.22 ± 0.28 | 0.15 ± 0.25 | 0.33 ± 0.35 |
| Pediastrum sp. | 0.37 ± 0.63 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Sceneclesmus sp. | 1.41 ± 0.72 | 0.67 ± 0.51 | 1.11 ± 0.61 | 2.52 ± 1.87 |
| Euglenophyta | 136.60 ± 22.87 | 104.40 ± 1.94 | 77.78 ± 20.02 | 183.40 ± 30.75 |
| Euglena sp. | 33.33 ± 20.79 | 33.33 ± 24.85 | 33.33 ± 24.85 | 61.11 ± 12.33 |
| Euglena oxyuris | 16.67 ± 15.71 | 22.22 ± 22.38 | 22.22 ± 22.38 | 22.30 ± 17.75 |
| Phacus sp. | 6.67 ± 11.33 | 4.44 ± 5.24 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Euglena viridis | 68.89 ± 29.37 | 44.44 ± 19.04 | 22.22 ± 4.44 | 88.89 ± 22.84 |
| Euglena pisciformis | 11.11 ± 18.89 | 0.00 ± 0.00 | 0.00 ± 0.00 | 11.11 ± 18.89 |
| Cryptophyta | 5.19 ± 3.19 | 2.22 ± 1.39 | 1.11 ± 1.05 | 3.70 ± 3.03 |
| Cryptomonas sp. | 5.19 ± 3.19 | 2.22 ± 1.39 | 1.11 ± 1.05 | 3.7 ± 3.03 |
| Pyrrophyta | 0.00 ± 0.00 | 0.30 ± 0.00.5 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Gymmodinium sp. | 0.00 ± 0.00 | 0.30 ± 0.5 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Total biomass | 160.7 | 108.4 | 85.49 | 127.1 |
| Date | Group | Dominant Species (Degree of Dominance) |
|---|---|---|
| 05-25 | Co | Chromulina pygmaea Nygaard (0.34) Chlorella pyrenoidesa (0.26) |
| T15 | Navicula sp. (0.03) Chromulina pygmaea Nygaard (0.21) Chlorella pyrenoidesa (0.17) | |
| T30 | Chromulina pygmaea Nygaard (0.27) Chlorella pyrenoidesa (0.41) | |
| T45 | Navicula sp. (0.02) Chromulina pygmaea Nygaard (0.29) Chlorella pyrenoidesa (0.33) | |
| 06-30 | Co | Chromulina pygmaea Nygaard (0.31) Chlorella pyrenoidesa (0.32) |
| T15 | Chromulina pygmaea Nygaard (0.17) Chroococcus sp. (0.14) Phormidium sp. (0.03) Chlorella pyrenoidesa (0.13) | |
| T30 | Navicula sp. (0.02) Chromulina pygmaea Nygaard (0.27) Chroococcus sp. (0.14) Phormidium sp. (0.03) Chlorella pyrenoidesa (0.17) Selenastrum bibraianum (0.02) | |
| T45 | Chromulina pygmaea Nygaard (0.26) Chroococcus sp. (0.15) Phormidium sp. (0.05) Chlorella pyrenoidesa (0.19) Chlamydomonas sp. (0.02) Sceneclesmus sp. (0.02) | |
| 07-15 | Co | Navicula sp. (0.04) Oscillatoria sp. (0.05) Microcystis sp. (0.02) Phormidium sp. (0.06) Chlorella pyrenoidesa (0.53) |
| T15 | Oscillatoria sp. (0.11) Microcystis sp. (0.09) Phormidium sp. (0.07) Chlorella pyrenoidesa (0.30) | |
| T30 | Navicula sp. (0.03) Chroococcus sp. (0.05) Phormidium sp. (0.04) Chlorella pyrenoidesa (0.69) | |
| T45 | Navicula sp. (0.07) Chromulina pygmaea Nygaard (0.12) Oscillatoria sp. (0.05) Phormidium sp. (0.06) Chlorella pyrenoidesa (0.35) | |
| 07-29 | Co | Chromulina pygmaea Nygaard (0.17) Oscillatoria sp. (0.03) Chlorella pyrenoidesa (0.06) Ankistrodesmus convolutus (0.08) |
| T15 | Chromulina pygmaea Nygaard (0.38) Oscillatoria sp. (0.03) Phormidium sp. (0.05) Chlorella pyrenoidesa (0.03) | |
| T30 | Navicula directa (0.04) Navicula sp. (0.03) Chromulina pygmaea Nygaard (0.19) Oscillatoria sp. (0.08) | |
| T45 | Chromulina pygmaea Nygaard (0.30) Chlorella pyrenoidesa (0.31) | |
| 08-15 | Co | Chromulina pygmaea Nygaard (0.30) Chlorella pyrenoidesa (0.07) |
| T15 | Chromulina pygmaea Nygaard (0.38) Chlorella pyrenoidesa (0.22) | |
| T30 | Chromulina pygmaea Nygaard (0.11) Chroococcus sp. (0.05) Oscillatoria sp. (0.03) Phormidium sp. (0.04) Chlorella pyrenoidesa (0.04) | |
| T45 | Chromulina pygmaea Nygaard (0.31) Oscillatoria sp. (0.04) Chlorella pyrenoidesa (0.33) | |
| 09-02 | Co | Nitzschia sp. (0.04) Chromulina pygmaea Nygaard (0.05) Chroococcus sp. (0.15) Chlorella pyrenoidesa (0.11) Sceneclesmus sp. (0.06) |
| T15 | Chromulina pygmaea Nygaard (0.25) Chlorella pyrenoidesa (0.25) | |
| T30 | Chromulina pygmaea Nygaard (0.14) Chlorella pyrenoidesa (0.48) | |
| T45 | Chromulina pygmaea Nygaard (0.03) Chroococcus sp. (0.04) Oscillatoria sp. (0.03) Chlorella pyrenoidesa (0.64) | |
| 09-15 | Co | Chromulina pygmaea Nygaard (0.17) Schroederi krosch (0.02) Pandorina morum (0.02) Eudorina elegans (0.03) Cryptomonas sp. (0.13) |
| T15 | Chromulina pygmaea Nygaard (0.21) Chlorella pyrenoidesa (0.61) | |
| T30 | Synedra sp. (0.03) Chromulina pygmaea Nygaard (0.35) Chlorella pyrenoidesa (0.19) | |
| T45 | Chromulina pygmaea Nygaard (0.24) Chlorella pyrenoidesa (0.53) Sceneclesmus sp. (0.04) | |
| 09-26 | Co | Chromulina pygmaea Nygaard (0.17) Chlorella pyrenoidesa (0.68) |
| T15 | Chromulina pygmaea Nygaard (0.05) Chlorella pyrenoidesa (0.87) | |
| T30 | Chlorella pyrenoidesa (0.51) Crucigenia sp. (0.04) | |
| T45 | Chromulina pygmaea Nygaard (0.18) Chroococcus sp. (0.07) Chlorella pyrenoidesa (0.31) Crucigenia sp. (0.06) | |
| 10-07 | Co | Synedra sp. (0.02) Navicula amphibola (0.02) Anabaena sp. (0.10) Selenastrum bibraianum (0.02) Eudorina elegans (0.10) |
| T15 | Chromulina pygmaea Nygaard (0.15) Chroococcus sp. (0.10) Chlorella pyrenoidesa (0.41) | |
| T30 | Chlorella pyrenoidesa (0.56) | |
| T45 | Navicula sp. (0.02) Chromulina pygmaea Nygaard (0.11) Chroococcus sp. (0.08) Chlorella pyrenoidesa (0.55) |
| Species | Biomass/mg/L | |||
|---|---|---|---|---|
| Co | T15 | T30 | T45 | |
| Protozoa | 1.75 ± 0.47 | 2.34 ± 1.02 | 1.36 ± 0.48 | 1.63 ± 0.47 |
| Sarcomastigophora | 1.52 ± 0.47 | 2.09 ± 1.01 | 1.25 ± 0.47 | 1.47 ± 0.48 |
| Saccamoeba sp. | 0.04 ± 0.03 | 0.05 ± 0.06 | 0.03 ± 0.02 | 0.02 ± 0.01 |
| Difflugia sp. | 0.13 ± 0.05 | 0.15 ± 0.08 | 0.13 ± 0.05 | 0.11 ± 0.05 |
| Difflugia oblonga | 0.37 ± 0.23 | 0.82 ± 0.61 | 0.41 ± 0.30 | 0.44 ± 0.23 |
| Tintinnidium fliuviatile | 0.97 ± 0.39 | 1.07 ± 0.49 | 0.68 ± 0.38 | 0.00 ± 0.37 |
| Arcella vulgaris | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Globigerinoides sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Ciliophora | 0.24 ± 0.1 | 0.25 ± 0.21 | 0.11 ± 0.04 | 0.15 ± 0.05 |
| Coleps sp. | 0.08 ± 0.06 | 0.07 ± 0.06 | 0.04 ± 0.02 | 0.05 ± 0.03 |
| Strobilidium sp. | 0.03 ± 0.02 | 0.08 ± 0.09 | 0.04 ± 0.02 | 0.06 ± 0.03 |
| Litonotus sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.01 | 0.01 ± 0.01 |
| Tintinnopsis sp. | 0.01 ± 0.01 | 0.05 ± 0.06 | 0.01 ± 0.02 | 0.00 ± 0.00 |
| Lembadion sp. | 0.06 ± 0.05 | 0.03 ± 0.02 | 0.01 ± 0.01 | 0.02 ± 0.01 |
| Zoothamnium sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Euplotes sp. | 0.00 ± 0.00 | 0.01 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Pseudoprorodon sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.01 | 0.01 ± 0.01 |
| Didinium nasufum | 0.03 ± 0.06 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Actinobolina sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Prorodon ovum | 0.01 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Pleuronema sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Stentor polymorphrus | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Vorticella sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Colpoda sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Trachelius sp. | 0.00 ± 0.00 | 0.01 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.01 |
| Rhabdostyla sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Cladocera | 22.16 ± 4.57 | 25.30 ± 5.77 | 29.19 ± 5.35 | 40.52 ± 6.71 |
| Daphnia carinata | 1.48 ± 2.52 | 8.89 ± 2.74 | 2.30 ± 0.44 | 0.74 ± 1.26 |
| Daphnia magna | 6.67 ± 1.33 | 0.00 ± 0.00 | 6.67 ± 1.33 | 16.67 ± 18.32 |
| Moinidae brachiata | 2.30 ± 0.78 | 1.52 ± 0.66 | 3.56 ± 1.73 | 2.52 ± 1.32 |
| Moinidae rectirostris | 5.93 ± 1.4 | 5.93 ± 0.21 | 4.44 ± 3.31 | 9.63 ± 3.05 |
| Moinidae macrocopa | 1.19 ± 0.42 | 0.52 ± 0.29 | 0.96 ± 0.12 | 0.44 ± 0.19 |
| Chydoroidea quadrangula | 2.59 ± 0.62 | 7.04 ± 0.46 | 9.26 ± 0.84 | 7.78 ± 0.61 |
| Chydoroidea sphaericus | 0.00 ± 0.00 | 0.07 ± 0.13 | 0.00 ± 0.00 | 0.07 ± 0.13 |
| Chydoroidea longirostris | 2.00 ± 1.89 | 1.33 ± 1.37 | 2.00 ± 2.31 | 2.67 ± 2.39 |
| Copepoda | 27.00 ± 5.19 | 29.65 ± 2.88 | 39.10 ± 3.27 | 30.70 ± 1.25 |
| Cyclops sp. | 8.44 ± 2.51 | 9.67 ± 0.22 | 10.89 ± 1.8 | 13.11 ± 2.83 |
| Copepodid | 0.60 ± 0.50 | 0.30 ± 0.17 | 0.30 ± 0.17 | 0.22 ± 0.14 |
| Calanoida sp. | 13.87 ± 1.78 | 16.18 ± 12.13 | 25.42 ± 0.86 | 13.87 ± 1.78 |
| Copepod nauplius | 4.09 ± 0.02 | 3.50 ± 0.04 | 2.49 ± 0.29 | 3.50 ± 0.87 |
| Rotifera | 0.70 ± 0.70 | 0.48 ± 0.29 | 0.21 ± 0.09 | 0.86 ± 0.72 |
| Brachionus calyciflorus | 0.07 ± 0.10 | 0.15 ± 0.16 | 0.04 ± 0.04 | 0.05 ± 0.06 |
| Brachionus ureus | 0.03 ± 0.02 | 0.10 ± 0.09 | 0.03 ± 0.03 | 0.05 ± 0.03 |
| Brachionus quadridentatus | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Polyarthra trigla | 0.16 ± 0.09 | 0.16 ± 0.14 | 0.06 ± 0.04 | 0.11 ± 0.08 |
| Polyarthra sp. | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Lecanidae inermis | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.02 ± 0.03 | 0.00 ± 0.00 |
| Asplachna brightwelli | 0.39 ± 0.65 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.58 ± 0.72 |
| Brachionus diversicornis | 0.01 ± 0.01 | 0.01 ± 0.02 | 0.00 ± 0.00 | 0.02 ± 0.04 |
| Rotaria citrine | 0.01 ± 0.01 | 0.03 ± 0.02 | 0.02 ± 0.02 | 0.04 ± 0.03 |
| Filinia sp. | 0.01 ± 0.01 | 0.00 ± 0.00 | 0.01 ± 0.01 | 0.00 ± 0.00 |
| Lepadella ovalis | 0.01 ± 0.01 | 0.02 ± 0.02 | 0.01 ± 0.01 | 0.01 ± 0.01 |
| Pedalia mira | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Keratella cochlearis | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Keratella valga | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Euchlanis pellucida | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.02 ± 0.03 | 0.00 ± 0.00 |
| Total biomass | 51.59 | 57.77 | 69.85 | 73.71 |
| Date | Group | Dominant Species (Degree of Dominance) |
|---|---|---|
| 05-25 | Co | Strobilidium sp. (0.26) Tintinnopsis sp. (0.03) Difflugia sp. (0.06) Saccamoeba sp. (0.05) Copepod nauplius (0.06) Brachionus calyciflorus (0.05) Brachionus ureus (0.23) |
| T15 | Coleps sp. (0.09) Strobilidium sp. (0.25) Tintinnopsis sp. (0.15) Saccamoeba sp. (0.05) Brachionus calyciflorus (0.04) Brachionus ureus (0.12) | |
| T30 | Strobilidium sp. (0.18) Daphnia carinata (0.13) Brachionus ureus (0.18) | |
| T45 | Coleps sp. (0.07) Strobilidium sp. (0.25) Tintinnidium fliuviatile (0.04) Daphnia magna (0.06) Brachionus ureus (0.10) | |
| 06-30 | Co | Tintinnidium fliuviatile (0.13) Moinidae brachiata (0.06) Cyclops sp. (0.02) Copepod nauplius (0.22) Polyarthra trigla (0.13) Brachionus diversicornis (0.02) Rotaria citrine (0.02) |
| T15 | Strobilidium sp. (0.02) Difflugia sp. (0.18) Tintinnidium fliuviatile (0.12) Zoothamnium sp. (0.02) Stentor polymorphrus (0.02) Moinidae brachiata (0.06) Chydoroidea quadrangula (0.04) Copepod nauplius (0.12) Brachionus ureus (0.02) Polyarthra trigla (0.17) Rotaria citrine (0.03) | |
| T30 | Tintinnopsis sp. (0.03) Difflugia sp. (0.20) Tintinnidium fliuviatile (0.17) Moinidae brachiata (0.07) Moinidae macrocopa (0.03) Chydoroidea quadrangula (0.04) Copepod nauplius (0.08) Brachionus ureus (0.08) Polyarthra trigla (0.09) | |
| T45 | Strobilidium sp. (0.12) Difflugia sp. (0.12) Tintinnidium fliuviatile (0.13) Moinidae brachiata (0.05) Copepod nauplius (0.11) Brachionus ureus (0.09) Polyarthra trigla (0.21) Brachionus diversicornis (0.02) | |
| 07-15 | Co | Strobilidium sp. (0.03) Difflugia sp. (0.09) Tintinnidium fliuviatile (0.13) Moinidae brachiata (0.08) Moinidae macrocopa (0.05) Cyclops sp. (0.03) Copepod nauplius (0.30) Polyarthra trigla (0.03) |
| T15 | Tintinnopsis sp. (0.03) Difflugia sp. (0.10) Tintinnidium fliuviatile (0.09) Moinidae brachiata (0.06) Cyclops sp. (0.03) Chydoroidea quadrangula (0.03) Copepod nauplius (0.19) Polyarthra trigla (0.13) Lepadella ovalis (0.08) | |
| T30 | Difflugia sp. (0.21) Tintinnidium fliuviatile (0.15) Moinidae brachiata (0.11) Cyclops sp. (0.04) Moinidae macrocopa (0.07) Chydoroidea quadrangula (0.08) Chydoroidea longirostris (0.06) Copepod nauplius (0.05) | |
| T45 | Difflugia sp. (0.15) Difflugia oblonga (0.04) Tintinnidium fliuviatile (0.20) Cyclops sp. (0.09) Moinidae brachiata (0.17) Moinidae macrocopa (0.03) Chydoroidea quadrangula (0.04) Chydoroidea longirostris (0.05) | |
| 07-29 | Co | Difflugia sp. (0.29) Tintinnidium fliuviatile (0.07) Moinidae brachiata (0.09) Cyclops sp. (0.06) Chydoroidea longirostris (0.05) Copepod nauplius (0.23) |
| T15 | Difflugia sp. (0.25) Difflugia oblonga (0.08) Tintinnidium fliuviatile (0.14) Cyclops sp. (0.04) Moinidae rectirostris (0.03) Chydoroidea longirostris (0.03) Copepod nauplius (0.05) Rotaria citrine (0.03) | |
| T30 | Difflugia sp. (0.19) Tintinnidium fliuviatile (0.05) Saccamoeba sp. (0.10) Moinidae brachiata (0.08) Cyclops sp. (0.04) Copepod nauplius (0.15) Polyarthra trigla (0.02) Filinia sp. (0.02) | |
| T45 | Rotaria citrine (0.11) Polyarthra trigla (0.04) Copepod nauplius (0.11) Chydoroidea quadrangula (0.09) Cyclops sp. (0.09) Moinidae brachiata (0.05) Tintinnidium fliuviatile (0.03) Difflugia oblonga (0.09) Difflugia sp. (0.24) | |
| 08-15 | Co | Coleps sp. (0.05) Difflugia sp. (0.05) Tintinnidium fliuviatile (0.03) Pleuronema sp. (0.03) Trachelius sp. (0.04) Saccamoeba sp. (0.04) Moinidae brachiata (0.04) Moinidae macrocopa (0.02) Cyclops sp. (0.02) Copepod nauplius (0.21) |
| T15 | Copepod nauplius (0.13) Trachelius sp. (0.22) Difflugia sp. (0.03) Coleps sp. (0.13) | |
| T30 | Coleps sp. (0.15) Strobilidium sp. (0.03) Difflugia sp. (0.09) Trachelius sp. (0.06) Rhabdostyla sp. (0.03) Moinidae brachiata (0.11) Cyclops sp. (0.09) Copepod nauplius (0.15) | |
| T45 | Coleps sp. (0.02) Difflugia sp. (0.05) Trachelius sp. (0.05) Saccamoeba sp. (0.02) Cyclops sp. (0.07) Copepod nauplius (0.23) | |
| 09-02 | Co | Coleps sp. (0.03) Difflugia sp. (0.03) Tintinnidium fliuviatile (0.08) Lembadion sp. (0.07) Zoothamnium sp. (0.02) Saccamoeba sp. (0.02) Cyclops sp. (0.05) Copepod nauplius (0.12) Polyarthra trigla (0.03) |
| T15 | Copepod nauplius (0.20) Cyclops sp. (0.07) Lembadion sp. (0.20) Difflugia oblonga (0.15) | |
| T30 | Difflugia sp. (0.03) Difflugia oblonga (0.07) Cyclops sp. (0.03) Copepod nauplius (0.30) | |
| T45 | Copepod nauplius (0.12) Moinidae rectirostris (0.04) Lembadion sp. (0.08) Tintinnidium fliuviatile (0.05) Difflugia oblonga (0.03) Strobilidium sp. (0.08) Coleps sp. (0.05) | |
| 09-15 | Co | Coleps sp. (0.08) Difflugia oblonga (0.05) Tintinnidium fliuviatile (0.05) Zoothamnium sp. (0.04) Cyclops sp. (0.02) Copepod nauplius (0.34) Polyarthra trigla (0.14) |
| T15 | Copepod nauplius (0.42) Cyclops sp. (0.06) Moinidae brachiata (0.03) Difflugia oblonga (0.07) Coleps sp. (0.06) | |
| T30 | Coleps sp. (0.12) Difflugia oblonga (0.13) Moinidae rectirostris (0.02) Cyclops sp. (0.04) Chydoroidea quadrangula (0.09) Copepod nauplius (0.35) | |
| T45 | Copepod nauplius (0.42) Chydoroidea quadrangula (0.06) Zoothamnium sp. (0.05) Tintinnidium fliuviatile (0.03) | |
| 09-26 | Co | Difflugia sp. (0.07) Difflugia oblonga (0.03) Lembadion sp. (0.32) Cyclops sp. (0.07) Calanoida sp. (0.03) Copepod nauplius (0.36) |
| T15 | Copepod nauplius (0.58) Calanoida sp. (0.02) Chydoroidea quadrangula (0.02) Cyclops sp. (0.06) Lembadion sp. (0.08) | |
| T30 | Difflugia sp. (0.03) Difflugia oblonga (0.06) Lembadion sp. (0.10) Cyclops sp. (0.08) Chydoroidea quadrangula (0.05) Copepod nauplius (0.43) | |
| T45 | Brachionus ureus (0.02) Copepod nauplius (0.39) Chydoroidea quadrangula (0.03) Cyclops sp. (0.11) Lembadion sp. (0.03) Difflugia oblonga (0.09) | |
| 10-07 | Co | Coleps sp. (0.06) Moinidae rectirostris (0.04) Cyclops sp. (0.05) Copepod nauplius (0.47) Copepodid (0.02) Polyarthra trigla (0.03) |
| T15 | Moinidae brachiata (0.02) Moinidae rectirostris (0.04) Cyclops sp. (0.08) Copepod nauplius (0.61) | |
| T30 | Coleps sp. (0.03) Cyclops sp. (0.17) Calanoida sp. (0.06) Copepod nauplius (0.38) | |
| T45 | Coleps sp. (0.05) Moinidae brachiata (0.03) Cyclops sp. (0.12) Copepod nauplius (0.52) Asplachna brightwelli (0.02) |
| Species | Biomass/g·m−2 | |||
|---|---|---|---|---|
| Co | T15 | T30 | T45 | |
| Annelida | 205.54 ± 438.17 | 290.72 ± 465.51 | 71.53 ± 208.79 | 124.92 ± 184.82 |
| Oligochaeta | 205.54 ± 438.17 | 290.72 ± 465.51 | 71.53 ± 208.79 | 124.92 ± 184.82 |
| Limnodrilus sp. | 18.14 ± 26.03 | 18.14 ± 32.55 | 11.91 ± 19.69 | 5.67 ± 10.33 |
| Branchiura sp. | 187.4 ± 424.71 | 272.58 ± 451.54 | 59.63 ± 193.87 | 119.25 ± 181.1 |
| Mollusca | 391.63 ± 174.88 | 383.92 ± 338.97 | 277.43 ± 153.15 | 300.6 ± 134.79 |
| Gastropoda | 391.63 ± 174.88 | 383.92 ± 338.97 | 277.43 ± 153.15 | 300.6 ± 134.79 |
| Gyraulus sp. | 271.18 ± 119.68 | 244.36 ± 141.38 | 208.6 ± 107.14 | 214.56 ± 106.47 |
| Euconulus sp. | 120.45 ± 115 | 139.57 ± 274.88 | 68.83 ± 83.12 | 86.04 ± 107 |
| Arthropoda | 0.00 ± 0.00 | 0.7 ± 3.55 | 0.35 ± 1.78 | 0.37 ± 1.78 |
| Insecta | 0.00 ± 0.00 | 0.7 ± 3.55 | 0.35 ± 1.78 | 0.37 ± 1.78 |
| Ephydra sp. | 0.00 ± 0.00 | 0.7 ± 3.55 | 0.35 ± 1.78 | 0.35 ± 1.78 |
| Corixa substriata | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.02 ± 0.1 |
References
- Bao, J.; Jiang, H.; Li, X. Thirty years of rice-crab coculture in China—Research progress and prospects. Rev. Aquac. 2022, 14, 1597–1612. [Google Scholar] [CrossRef]
- Jia, E.T.; Zheng, X.; Cheng, H.; Jie, L.; Zhang, D. Dietary fructooligosaccharide can mitigate the negative effects of immunity on Chinese mitten crab fed a high level of plant protein diet. Fish Shellfish. Immunol. 2019, 84, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhang, Z. Ecological economic analysis of a rice-crab model. Chin. J. Appl. Ecol. 2002, 13, 323–326. (In Chinese) [Google Scholar]
- Bashir, M.A.; Liu, J.; Geng, Y.; Wang, H.; Pan, J.; Zhang, D.; Rehim, A.; Aon, M.; Liu, H. Co-culture of rice and aquatic animals: An integrated system to achieve production and environmental sustainability. J. Clean. Prod. 2020, 249, 119310. [Google Scholar] [CrossRef]
- Zhang, Q.; Lv, D.; Ma, X.; Wang, A.; Wang, W. The effects of different transplanting ways on the phytoplankton community in rice-crab culture system. J. Food Agric. Environ. 2014, 12, 199–204. [Google Scholar]
- Zhang, C. In Partial Fulfillment of the Requirement for the Degree of Master in Hydrobiology. Master’s Thesis, Dalian Ocean University, Dalian, China, 2015. (In Chinese). [Google Scholar]
- Lunger, A.N.; Mclean, E.; Craig, S.R. The effects of organic protein supplementation upon growth, feed conversion and texture quality parameters of juvenile cobia (Rachycentron canadum). Aquaculture 2007, 264, 342–352. [Google Scholar] [CrossRef]
- Lin, S.; Luo, L.; Ye, Y. Effects of dietary protein level on growth, feed utilization and digestive enzyme activity of the Chinese mitten crab, Eriocheir sinensis. Aquac. Nutr. 2010, 16, 290–298. [Google Scholar] [CrossRef]
- Xu, C.; Wen, L.; Zhang, D.; Liu, J.; Zheng, X.; Zhang, C.; Yao, J.; Zhu, C.; Chi, C. Effects of partial fish meal replacement with two fermented soybean meals on the growth of and protein metabolism in the Chinese mitten crab (Eriocheir sinensis). Aquac. Rep. 2020, 17, 100328. [Google Scholar] [CrossRef]
- Ministry of Agriculture and Rural Development of China. Development report of China’s rice and fishery comprehensive planting and breeding industry. China Fish. 2020, 10, 12–19. (In Chinese) [Google Scholar]
- Lei, Y. Environmental Chemistry of Aquaculture Water (for Aquaculture Specialty), 1st ed.; China Agricultural Publishing House: Beijing, China, 2004; pp. 67–108. (In Chinese) [Google Scholar]
- Fu, L.; Zhou, G.; Li, Y.; Lu, Q.; Pan, J. Research progress on precocious puberty of Chinese mitten crab. Jiangsu Agric. Sci. 2017, 45, 5. (In Chinese) [Google Scholar]
- Li, H.; Sun, C.; Zhang, Q.; Zhang, P.; Bao, H.; Ren, H.; Yao, X.; Zhong, W.; Cui, K.; You, H.; et al. Study on the Composition of Plankton Species in Tianjin Area; Tianjin Aquatic Products Technology Promotion Statio: Tianjin, China, 2017. (In Chinese) [Google Scholar]
- Zhao, W. Aquatic Biology, 1st ed.; China Agricultural Publishing House: Beijing, China, 2005; pp. 62–365. (In Chinese) [Google Scholar]
- Li, Y.; Ding, J.; Xu, F. On the distribution and hatching of resting eggs of rotifer in fishponds. Acta Hydrobiol. Sin. 1985, 9, 20–31. (In Chinese) [Google Scholar]
- Cheng, Y.X.; Du, N.S.; Lai, W. Lipid composition in hepatopancreas of Chinese mitten crab Eriocheir sinensis at different stages. Acta Hydrobiol. Sin. 1998, 44, 420–429. [Google Scholar]
- Bao, J.; Jiang, H.; Shen, H.; Xing, Y.; Chen, Q. First description of milky disease in the Chinese mitten crab Eriocheir sinensis caused by the yeast Metschnikowia bicuspidata. Aquaculture 2021, 532, 735984. [Google Scholar] [CrossRef]
- Prein, M. Integration of aquaculture into crop–animal systems in Asia. Agric. Syst. 2002, 71, 127–146. [Google Scholar] [CrossRef]
- Hu, L.; Guo, L.; Zhao, L.; Shi, X.; Ren, W.; Zhang, J.; Tang, J.; Chen, X. Productivity and the complementary use of nitrogen in the coupled rice-crab system. Agric. Syst. 2020, 178, 102742. [Google Scholar] [CrossRef]
- Chen, X.; Wang, J.; Xin, H.; Yue, W.; Shu, H. Tissue-expression profiles unveils the gene interaction of hepatopancreas, eyestalk, and ovary in precocious female Chinese mitten crab, Eriocheir sinensis. BMC Genet. 2018, 20, 12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Li, Z.; Cui, Y. Survival, Growth, Sex Ratio, and Maturity of the Chinese Mitten Crab (Eriocheir sinensis) Reared in a Chinese Pond. J. Freshw. Ecol. 2001, 16, 633–640. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Li, W.; Liu, D.; Wu, W.; Liu, Y.; Wen, X.; Liao, Y. Seasonal characteristics of CO2 fluxes in a rain-fed wheat field ecosystem at the Loess Plateau. Span. J. Agric. Res. 2013, 11, 980–988. [Google Scholar] [CrossRef] [Green Version]
- Mb, A.; Mma, B.; Mk, C.; Ph, A.; Bta, B.; Lpa, B. Dissolved oxygen deficits in a shallow eutrophic aquatic ecosystem (fishpond)-sediment oxygen demand and water column respiration alternately drive the oxygen regime. Sci. Total Environ. 2020, 766, 142647. [Google Scholar]
- Zhang, Y.J.; Wang, A.; Ma, X.; Wang, W.; Li, Y. Preliminary study on level changes of water quality in rice-crab culture. Guangdong Agric. Sci. 2013, 40, 16–19. [Google Scholar]
- Wang, A.; Wang, W.; Ma, X.; Wang, Q.; Yu, Y.; Chen, W. Study on the changes of water nnvironmental factors in rice-crab culture system. Hubei Agric. Sci. 2011, 50, 6. (In Chinese) [Google Scholar]
- Lobo-Araújo, L.W.; Toledo, M.; Efe, M.; Malhado, A.; Vital, M.; Toledo-lima, G.; Macario, P.; Santos, J.; Ladle, R. Bird communities in three forest types in the Pernambuco Centre of Endemism, Alagoas, Brazil. Iheringia Série Zool. 2013, 103, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Jian, X.; Hu, L.; Tang, J.; Wu, X.; Li, N. Ecological mechanisms underlying the sustainability of the agricultural heritage rice-fish coculture system. Proc. Natl. Acad. Sci. USA 2011, 108, E1381–E1387. [Google Scholar]
- Frei, M.; Khan, M.; Razzak, M.; Hossain, M.; Dewan, S.; Becker, K. Effects of a mixed culture of common carp, Cyprinus carpio L.; Nile tilapia, Oreochromis niloticus (L.), on terrestrial arthropod population, benthic fauna, and weed biomass in rice fields in Bangladesh. Biol. Control. 2007, 41, 207–213. [Google Scholar] [CrossRef]
- Altieri, M.A. Linking ecologists and traditional farmers in the search for sustainable agriculture. Front. Ecol. Environ. 2004, 2, 35–42. [Google Scholar] [CrossRef]
- Ren, W.; Hu, L.; Guo, L.; Zhang, J.; Tang, L. Preservation of the genetic diversity of a local common carp in the agricultural heritage rice-fish system. Proc. Natl. Acad. Sci. USA 2018, 115, E546–E554. [Google Scholar] [CrossRef] [Green Version]
- Lassen, M.F.; Bramm, M.; Richardson, K.; Yusoff, F.; Shariff, M. Phytoplankton community composition and size distribution in the Langat River estuary, Malaysia. Estuaries 2004, 27, 716–727. [Google Scholar] [CrossRef]
- Sterner, R.W.; Chrzanowski, T.; Elser, J.; George, N. Sources of nitrogen and phosphorus supporting the growth of bacteria and phytoplankton in an oligotrophic Canadian shield lake. Limnol. Oceanogr. 1995, 40, 242–249. [Google Scholar] [CrossRef]
- Odum, W.E. Ecological Guidelines for Tropical Coastal Development; IUCN Publications New Series. No. 42; IUCN: Gland, Switzerland, 1976. [Google Scholar]
- Cabecinha, E.; Cortes, R.; Cabral, J.; Ferreira, T.; Lourenço, M.; Pardal, M. Multi-scale approach using phytoplankton as a first step towards the definition of the ecological status of reservoirs. Ecol. Indic. 2009, 9, 240–255. [Google Scholar] [CrossRef]
- Sun, W.T.; Zhang, Q.; Xu-Zhou, M.; Wang, W.; Wang, A. A study on effects of different crab stocking density on water environment and rice yield. J. Shanghai Ocean. Univ. 2014, 23, 366–373. [Google Scholar]
- Qingtian, L.I.; Wang, H.; Zheng, L.I. The biodiversity and dynamics of oribatid mites in rice paddies. Syst. Appl. Acarol. 2000, 5, 9. [Google Scholar]
- Horn, W. Long-term development of the crustacean plankton in the Saidenbach Reservoir (Germany)–changes, causes, consequences. Hydrobiologia 2003, 504, 185–192. [Google Scholar] [CrossRef]
- Gilbert, K. Quantitative comparison of food niches in some freshwater zooplankton. Oecologia 1987, 72, 331–340. [Google Scholar]
- Perrow, M.R.; Jowitt, A.; Stansfield, J.; Phillips, G. The Practical Importance of the Interactions between Fish, Zooplankton and Macrophytes in Shallow Lake Restoration; Springer: Dordrecht, The Netherlands, 1999. [Google Scholar]
- Wen, X.F.; Zhang, Y.; Zhen, L.; Jin, J.; Duan, T. Research on effection of the comprehensive cultivation mode of the shrimp, crab, submerged plant polyculture and the ecological floating bed. Guangdong Agric. Sci. 2012, 24, 138–142. [Google Scholar]
- Ueji, M.; Inao, K. Rice paddy field herbicides and their effects on the environment and ecosystems. Weed Biol. Manag. 2001, 1, 71–79. [Google Scholar] [CrossRef]
- Guo, H.; Tang, D.; Shi, X.; Wu, Q.; Wang, Z. Comparative transcriptome analysis reveals the expression and characterization of digestive enzyme genes in the hepatopancreas of the Chinese mitten crab. Fish. Sci. 2019, 85, 979–989. [Google Scholar] [CrossRef]
- Sun, L.P.; Song, X.; Zhu, J.; Meng, X.; Zhang, L. Effects of submerged plants on growth performance and non-specific immunity of Chinese mitten crab (Eriocheir sinensis). Freshw. Fish. 2012, 42, 35–40. [Google Scholar]
- Zeng, Q.; Jeppesen, E.; Gu, X.; Mao, Z.; Chen, H. Cannibalism and Habitat Selection of Cultured Chinese Mitten Crab: Effects of Submerged Aquatic Vegetation with Different Nutritional and Refuge Values. Water 2018, 10, 1542. [Google Scholar] [CrossRef] [Green Version]
- Lv, D.; Wang, W.; Xu-Zhou, M.; Wang, Q.; Wang, A.; Chen, Z.; Tian, S. Ecological Prevention and Control of Weeds in Rice-crab Polycultured Field. Hubei Agric. Sci. 2011, 50, 1574–1578. [Google Scholar]
- Li, X.; Dong, S.; Lei, Y.; Li, Y. The effect of stocking density of Chinese mitten crab Eriocheir sinensis on rice and crab seed yields in rice–crab culture systems. Aquaculture 2007, 273, 487–493. [Google Scholar] [CrossRef]
- Hong, Y.; Jiang, C. Effects of stocking Chinese mitten crab on the zoobenthos and aquatic vascular plant in the east lake reservoir, Heilongjiang, China. Acta Hydrobiol. Sin. 2005, 29, 430–434, (In Chinese with English Abstract). [Google Scholar]
- Xu, Q.; Wang, H.; Zhang, S. The impact of overstocking of mitten crab, Eriocheir sinensis, on lacustrine zoobentic community. Acta Hydrobiol. Sin. 2003, 27, 41–46, (In Chinese with English Abstract). [Google Scholar]

















| Ingredients/% | Content | ||
|---|---|---|---|
| T15 | T30 | T45 | |
| Fish meal | 6 | 27 | 47 |
| Soybean meal | 3 | 13.5 | 23.5 |
| Beer yeast | 3 | 3 | 3 |
| Wheatmeal | 72.59 | 43.09 | 15.09 |
| Fish oil | 7.5 | 5.5 | 3.5 |
| Lecitin | 0.5 | 0.5 | 0.5 |
| Mineral premix | 2 | 2 | 2 |
| Vitamin premix | 2 | 2 | 2 |
| Squid paste | 1 | 1 | 1 |
| Glycine betaine | 0.5 | 0.5 | 0.5 |
| Choline chloride | 0.2 | 0.2 | 0.2 |
| Calcium dihydrogen phosphate | 1.5 | 1.5 | 1.5 |
| Chromium oxide | 0.1 | 0.1 | 0.1 |
| Ethoxyquin | 0.01 | 0.01 | 0.01 |
| Calcium propionate | 0.1 | 0.1 | 0.1 |
| Total | 100 | 100 | 100 |
| Crude protein | 15.00 | 30.49 | 45.25 |
| Crude lipid | 9.13 | 9.27 | 9.31 |
| Total energy/KJ·g−1 | 18.09 | 16.57 | 17.17 |
| Growth Performance and Yield | Groups | |||
|---|---|---|---|---|
| Co | T15 | T30 | T45 | |
| Initial body weight/g·ind−1 | 0.09 ± 0.02 | 0.09 ± 0.01 | 0.09 ± 0.01 | 0.09 ± 0.02 |
| Final body weight/g·ind−1 | 8.30 ± 3.19 a | 13.10 ± 1.59 ab | 14.35 ± 1.87 ab | 17.28 ± 6.23 b |
| Survival rate/% | 17.93 ± 8.25 | 12.51 ± 3.38 | 8.46 ± 2.76 | 10.38 ± 3.43 |
| Weight gain rate/% | 9641.86 ± 3738.90 a | 15,277.93 ± 1860.99 ab | 16,737.02 ± 2194.882 ab | 20,181.73 ± 7316.62 b |
| Specific growth rate/%·d−1 | 3.30 ± 0.33 a | 3.67 ± 0.09 ab | 3.74 ± 0.09 b | 3.85 ± 0.26 b |
| Total output/g·m−2 | 48.38 ± 10.14 | 58.62 ± 10.05 | 43.86 ± 12.27 | 60.41 ± 1.18 |
| Net output/g·m−2 | 42.62 ± 10.14 | 52.86 ± 10.05 | 38.10 ± 12.27 | 54.64 ± 1.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Wan, J.; Liang, X.; Wang, Y.; Liu, X.; Mei, J.; Sun, N.; Li, X. Effects of Protein Level on the Production and Growth Performance of Juvenile Chinese Mitten Crab (Eriocheir sinensis) and Environmental Parameters in Paddy Fields. Water 2022, 14, 1941. https://doi.org/10.3390/w14121941
Yu Y, Wan J, Liang X, Wang Y, Liu X, Mei J, Sun N, Li X. Effects of Protein Level on the Production and Growth Performance of Juvenile Chinese Mitten Crab (Eriocheir sinensis) and Environmental Parameters in Paddy Fields. Water. 2022; 14(12):1941. https://doi.org/10.3390/w14121941
Chicago/Turabian StyleYu, Yilin, Jiwu Wan, Xiaochen Liang, Yuquan Wang, Xueshen Liu, Jie Mei, Na Sun, and Xiaodong Li. 2022. "Effects of Protein Level on the Production and Growth Performance of Juvenile Chinese Mitten Crab (Eriocheir sinensis) and Environmental Parameters in Paddy Fields" Water 14, no. 12: 1941. https://doi.org/10.3390/w14121941