
Whole-Plant Water Use and Hydraulics of Populus euphratica and Tamarix ramosissima Seedlings in Adaption to Groundwater Variation

1
State Key Laboratory of Desert and Oasis Ecology, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi 830011, China
2
College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, China
3
Akesu National Station of Observation and Research for Oasis Agro-Ecosystem, Akesu 843017, China
*
Author to whom correspondence should be addressed.
Water 2022, 14(12), 1869; https://doi.org/10.3390/w14121869
Submission received: 23 April 2022
/
Revised: 26 May 2022
/
Accepted: 7 June 2022
/
Published: 10 June 2022
(This article belongs to the Section Ecohydrology)
Abstract
:Riparian phreatophytes in hyperarid areas face selection pressure from limiting groundwater availability and high transpiration demand. We examined whole-plant water use and hydraulic traits in Populus euphratica and Tamarix ramosissima seedlings to understand how they adapt to groundwater variations. These species coexist in the Tarim River floodplain of western China. Measurements were performed on 3-year-old seedlings grown in lysimeters simulating various groundwater depths. P. euphratica had relatively greater leaf area-specific water use due to its comparatively higher sapwood area to leaf area ratio (Hv). A high Hv indicates that its sapwood has a limited capacity to support its leaf area. P. euphratica also showed significantly higher leaf-specific conductivity (ksl) than T. ramosissima but both had similar sapwood-specific conductivities (kss). Therefore, it was Hv rather than kss which accounted for the interspecific difference in ksl. When groundwater was not directly available, ksl and Hv in P. euphratica were increased. This response favors water loss control, but limits plant growth. In contrast, T. ramosissima is more capable of using deep groundwater. Stomatal sensitivity to increasing leaf-to-area vapor pressure deficit was also higher in P. euphratica. Overall, P. euphratica is less effective than T. ramosissima at compensating for transpirational water loss at a whole-plant level. For this reason, P. euphratica is restricted to riverbanks, whereas T. ramosissima occurs over a wide range of groundwater depths.
1. Introduction
Riparian phreatophytic plants in arid regions face two major water-related challenges. One is groundwater access which is determined by root growth rate and changes in groundwater depth. The other is the fact that the canopy water supply must contend with high transpirational demand caused by a high leaf-to-air vapor pressure deficit (LVPD), even when the plants have groundwater access [1]. High LVPD prevails in desert riparian zones inhabited by phreatophytes. However, the groundwater depth there is highly variable [2] because there are substantial groundwater flow variations in arid regions. Moreover, human activity has introduced widespread hydrological alterations [3]. These perturbations significantly affect the establishment, survival, growth, and community structure of riparian plants [4,5]. In the riparian ecosystems of the southwestern U.S., the native Populus and Salix species were replaced by the invasive Tamarix species. Flow regime alterations, including subsequent groundwater decline, were the major factors responsible for this species substitution [6,7]. Tamarix expansion has reduced river runoff due to its high demand for water. Nevertheless, it has also diminished the availability of water resources for human consumption [8]. This scenario highlights the importance of understanding water use and riparian plant hydraulics to elucidate ecological processes and to improve and sustain riparian ecosystem management.
Knowing plant responses to changes in water availability helps to predict plant adaptions to environmental variations and determine trends in vegetation dynamics as a result of a changing climate [9,10]. In application, this is also helpful for developing eco-friendly solutions in managing riverine ecosystems [11] and improving regional evapotranspiration monitoring [12,13]. However, our understanding of plant responses to water availability is very limited, particularly in arid ecosystems [14]. Certain desert riparian ecosystems are characterized by highly variable water availability. Learning the differential responses of coexisting plant species to varying water availability is necessary to understand plant water relations in these ecosystems [15]. In general, plant species which grow rapidly during high water availability and tolerate drought are relatively more competitive in riparian communities [16]. When groundwater descends below the root zones, plants must rely on precipitation. This response induces plant species composition changes [17]. The composition changes which followed the Tamarix invasion of the southwestern U.S. were the result of alterations in plant water use in response to groundwater variations. The predominant Tamarix stands had very high evapotranspiration rates, even as groundwater levels dropped [18].
There are species-specific differences in ecophysiological response to water availability. These provide some insight into plant adaptions [19]. Plants growing in extremely variable conditions have powerful regulatory mechanisms which modulate water transport according to water availability. These compensations help maintain plant physiological functions [20]. The stomata control water flux and loss during drought. The hydraulic transport system [21] also regulates water relations. The greatest hydraulic conductivity losses occur in the xylem during drought [22]. The sapwood-specific hydraulic conductivity (kss) characterizes xylem hydraulic efficiency. The leaf-specific hydraulic conductivity (ksl) defines the sapwood hydraulic capacity to meet the transpirational demand of a given leaf area [23]. A higher ksl may be driven by the xylem hydraulic capacity, the sapwood/leaf area ratio, or both. Plants in water-limited ecosystems may alter their biomass allocation according to water availability to optimize their survival and/or productivity [24]. Therefore, in order to understand water use strategies in response to water availability changes, it is necessary to address plant–water relations at the whole-plant scale [25,26,27] and across a wide range of water availabilities. Plant hydraulics and stomatal behavior are closely correlated with carbon gain [28,29]. Consequently, a comparative study of the hydraulic traits of different plant species may account for species distributions at varying water availabilities and explain the divergence of plant adaptions.
The vast inland Tarim Basin of central Eurasia has a hyperarid climate characterized by high evaporative demand and extremely low precipitation (annual mean ~50 mm). Desert riparian vegetation grows along the 1321 km length of the Tarim River and is predominated by Populus euphratica and Tamarix ramosissima [30]. These phreatophytes are highly dependent on groundwater that declines with increasing distance from the active channel [31]. As the Tarim river is mainly recharged by glacier melt from surrounding mountains [32], great hydrological fluctuations can result in an annual groundwater variation greater than 2 m, even in riparian zones close to the active channel [33]. Increased agricultural water use over the past 50 years has declined groundwater in the whole basin, leading to the intensive degeneration of the desert riparian forest, especially in the lower reaches of the river [34]. This triggered an ecological engineer in 2000 to artificially delivered water to rise groundwater levels in the lower reaches of the river, thus conserving the desert riparian vegetation [34]. However, the rise in groundwater only occurred hundreds of meters near the riverbank [35]. Groundwater availability is still a key problem for desert riparian plants in the Tarim Basin.
P. euphratica and T. ramosissima share the same establishment niche because they are both established in newly deposited sediments after flood recessions and form co-dominant seedling stands near the active channel [36]. Their dominances change with increasing distance from the channel due to alterations in groundwater level [31]. This can be related to their water uses and the underlying hydraulics, as inequalities in hydraulic response to water conditions cause niche differentiation among plant species within the same habitat [37]. Currently, comparative studies on water relations between these co-occurring species include stomatal response to LVPD [38], xylem resistance to embolism [39], leaf-level water use [40], sap flow and water resources [41], and so on. These works basically focus on hydraulics at an organ level, but studies on the whole-plant level water use, as well as in relation to plant hydraulics, are entirely lacking. However, hydraulic traits are coordinated [42] and their relationships depend on water availability [43].
Our objective was to compare the water use and hydraulics of the phreatophytes P. euphratica and T. ramosissima in adaption to the groundwater variations they encounter in their coexisting habitats along the Tarim River riparian zones. We used lysimeters to simulate varying groundwater depths and compare water use and underlying hydraulic traits at a whole-plant level. Measurements included daily whole-plant water use, plant water status, hydraulic functional traits (kss, ksl, and stomatal sensitivity to LVPD), and structural traits (Hv and wood density).
2. Materials and Methods
2.1. Plant Materials and Experiment Design
The experiment was carried out at the Aksu Water Balance Station, Chinese Academy of Sciences (40°27′ N, 80°45′ E; hereafter, Aksu Station) in 2009. Three-year-old Populus euphratica and Tamarix ramosissima seedlings were grown in homemade lysimeters 1.2 m in height and 0.3 m in inner diameter, as shown in Figure 1. Transplanted seedlings were ~50 cm tall. At this stage, neither species had yet established dense stands. There was one seedling per lysimeter. The seedlings were established then subjected to the equivalent of 20 cm, 60 cm, and 100 cm distance from the groundwater between early July and mid-September. Each treatment included five replicates per species. Therefore, there were 2 species × 3 treatments × 5 replicates = 30 lysimeters in total for the experiment. For experimental procedure details, see the work by Li et al. [36]. An automatic weather station (M520, Vaisala Co. Ltd., Vantaa, Finland) was set ~50 m from the experimental site to measure and record meteorological data including rainfall, temperature, pressure, wind direction and velocity, relative humidity, solar radiation, etc.
2.2. Measurements
Daily evapotranspiration (ET) rates were obtained for each lysimeter in August, one month after the seedlings were subjected to the groundwater treatments. The volume of water added to maintain the corresponding groundwater depth in each lysimeter was recorded daily. Plant water status was determined on 31 August by measuring predawn (ψpd) and midday (ψmd) xylem water potentials (ψ) with a PMS Model 1000 pressure chamber (PMS Instrument Co., Albany, OR, USA). ψpd and ψmd were measured ~30 min before sunrise and at solar noon, respectively. On 31 August, gravimetric soil water content was measured for three lysimeters per species per treatment. These plants were also selected for rooting depth observation and biomass harvest. Soil water content was determined by sampling at 10 cm increments from the soil surface to the corresponding groundwater depth then drying and weighing the samples.
Stomatal response to LVPD was evaluated with a portable photosynthesis system (Li6400, LI-COR Biosciences, Lincoln, NE, USA) consisting of a standard chamber fitted with a light source (6400-02B, LI-COR Biosciences, Lincoln, NE, USA). Measurements were conducted on clear days in August between 10h30 and 13h30, local time. To assess sensitivity to increasing LVPD, the relative humidity (RH) within the cuvette was first set to 60% then progressively decreased to the lowest possible level by absorbing all water vapor from the air entering the LI-COR system (HR = 0% in the reference chamber). The LVPD response assay was repeated on three individuals per species per treatment. The cuvette CO2 concentration was maintained at 400 ppm and the photosynthetic photon flux density (PPFD) was held at the light saturation points previously determined by light response measurements [36]. These were 2400 μmol m−2 s−1 for P. euphratica and 1800 μmol m−2 s−1 for T. ramosissima. Leaves within the cuvette were cut immediately after each measurement and their leaf areas were measured. Thence, the gas exchange parameters were recalculated in the LI-COR system.
At the end of the experiment, three randomly selected seedlings per species per treatment were harvested to measure specific leaf area (SLA) and total leaf area (TLA). On each sampled seedling, fresh leaves from three randomly selected branches were gently extended onto a white board, and then photographed for area calculation using Image Pro Plus software (Media Cybernetics Inc., Rockville, MD, USA). The average SLA of each sampled seedling was obtained from the ratio of leaf area to dry biomass. Therefore, the TLA of each sampled seedling was estimated as average SLA × total harvested leaf biomass. Because leaves of T. ramosissima are cylindrical, we calculated the actual leaf surface area as the photographic projected area ×π/2, as described by Li et al. [36]. To estimate the sapwood-specific conductivity at the whole-plant scale, the sapwood area was evaluated by measuring the cross-sectional areas of the main stem at the soil surface. Heartwood was excluded from the calculations and differentiated from the sapwood by its distinct color. Wood density was measured on a debarked ~10-cm-long stem segment excised from each seedling sample. Fresh volume was measured by water displacement at a constant 20 °C. Sample mass was measured after 48 h drying. Wood density was reported as dry mass per unit fresh volume.
2.3. Data Processing and Statistics
Potential evapotranspiration (PET) was estimated from daily weather data by the FAO Penman–Monteith method [44]. To partition soil evaporation (E) from evapotranspiration (ET), E was estimated by calculating the soil water flux (K(S)) in unsaturated soil using a combination of Van Genuchten’s equation [45] and Mualem’s model [46] to predict unsaturated hydraulic conductivity:
where Ks is the saturated hydraulic conductivity (cm d−1) and α, m, and n are empirical parameters shaping the retention curve.
For the soils used in this experiment, Ks = 3.67 cm d−1 was determined by the constant head method [47]. In addition, α = 0.0081, n = 1.3481, and m = 0.2582 were derived from the soil water retention curve plotted by the pressure chamber method. Whole-plant water use was estimated from the difference between the ET and the soil evaporation on August 30 at the same time that soil water content, plant water potential, and TLA were evaluated.
The Huber value (Hv, cm2 m−2) was calculated as the ratio of sapwood area to TLA. Whole-plant hydraulic conductance (Kplant, g d−1 MPa−1) was calculated as Kplant = (ET−E)/[(ψpd−ψmd)] [26,48]. Leaf area-specific whole-plant hydraulic conductance (ksl, g d−1 m−2 MPa−1) was calculated by dividing Kplant by TLA. Sapwood-specific whole-plant hydraulic conductance (kss, g d−1 cm−2 MPa−1) was calculated by dividing Kplant by sapwood area. Stomatal sensitivity was calculated by Δgs/ΔLVPD where Δgs and ΔLVPD are the differences in stomatal conductance and LVPD, respectively, when RH is decreased. Differences in hydraulic traits among species and treatments were tested by two-way ANOVA. Statistical analyses were run in SPSS v.11.0 (IBM Corp., Armonk, NY, USA).
3. Results
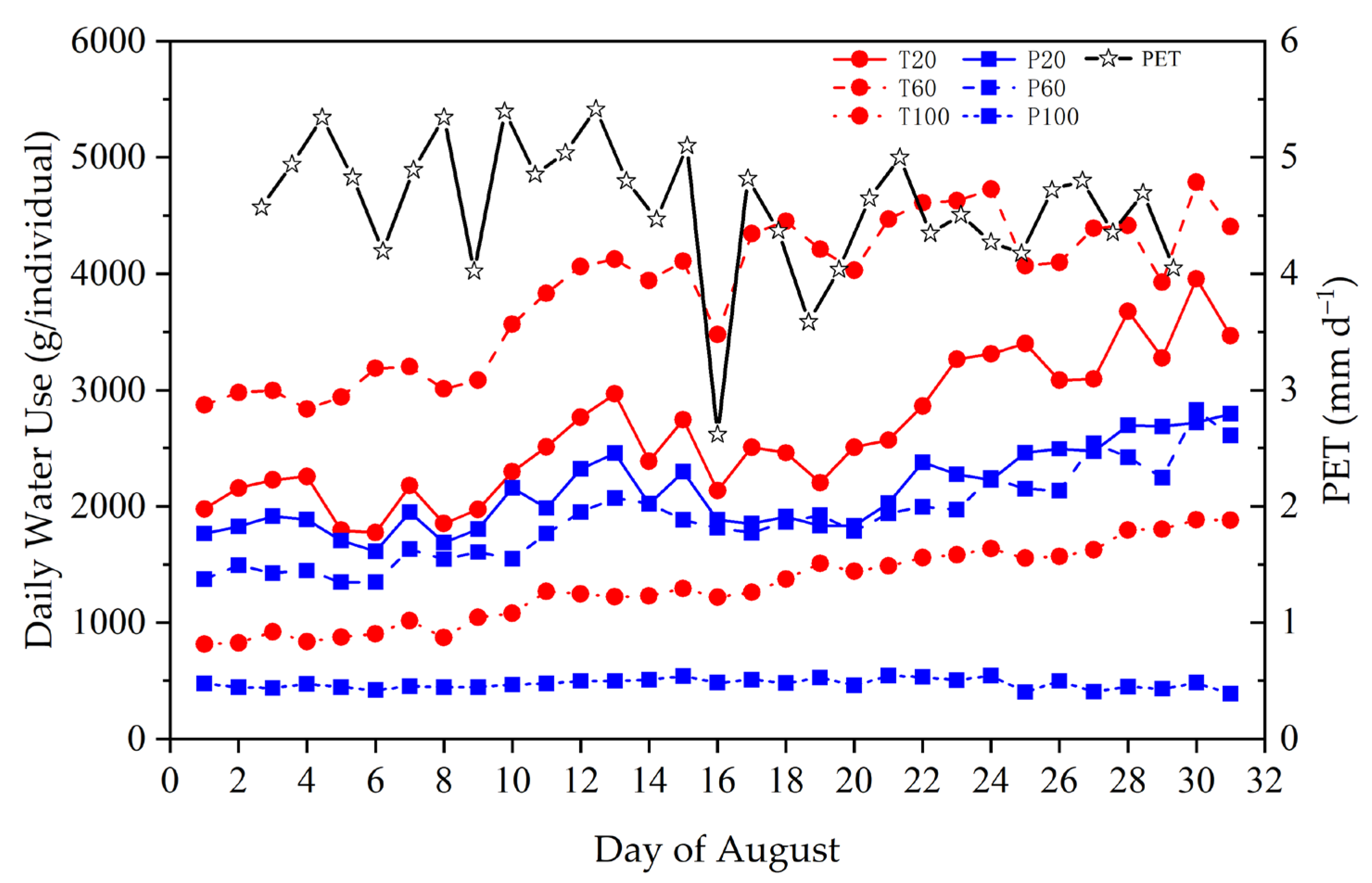
Daily PET fluctuated and trended to decrease with time in August, while daily water uses per individual, with the exception P. euphratica grown at 100 cm groundwater depth, trended to increase with time (Figure 2), indicating plant water use increasing with plant growth during the experiment. Here, the daily water use per individual included transpiration and soil evaporation, but the soil evaporation only occupied a slight portion of the total (<5% even under shallow groundwater condition), as estimated on the hydraulic conductivity in unsaturated soils sampled on 30 August. Thus, the daily water use per lysimeter could reflect the changes in whole-plant water use during the observation period.
As shown in Figure 2, the whole-plant water use differed between species and among treatments. For P. euphratica, seedlings grown 20 cm and 100 cm groundwater depths had similar daily water use, both >5 times higher than those at 100 cm groundwater depth at the end of the experiment. Differently, T. ramosissima seedlings grown at 60 cm groundwater depth had the highest water use, followed by those at 20 cm and 100 cm groundwater depth, respectively. T. ramosissima seedlings at both 20 cm and 60 cm groundwater depths had greater water use than P. euphratica ones. In comparison to the unchanged water use of P. euphratica seedlings grown at 100 cm groundwater depth, water use of T. ramosissima under the same condition underwent a clear increase, resulting in a greater difference in water use between species in the late stage of the experiment.
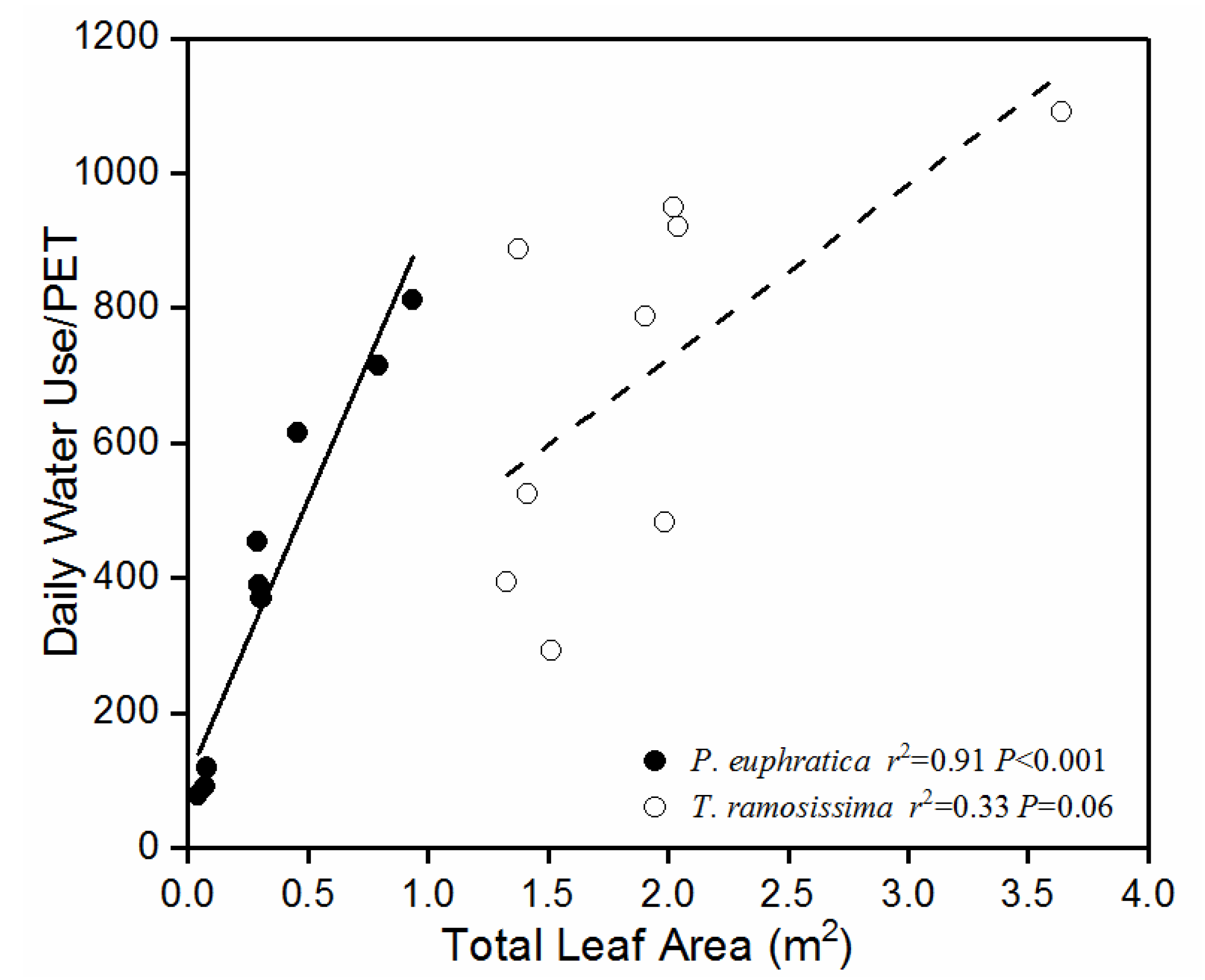
PET-normalized whole-plant daily water use increased linearly with TLA in P. euphratica (p < 0.001; Figure 3). However, the correlation between these two variables was weak in T. ramosissima (p = 0.06; Figure 3). The slope of the regression was greater for P. euphratica than T. ramosissima. Therefore, the water use per leaf area at each PET was higher in P. euphratica than T. ramosissima.
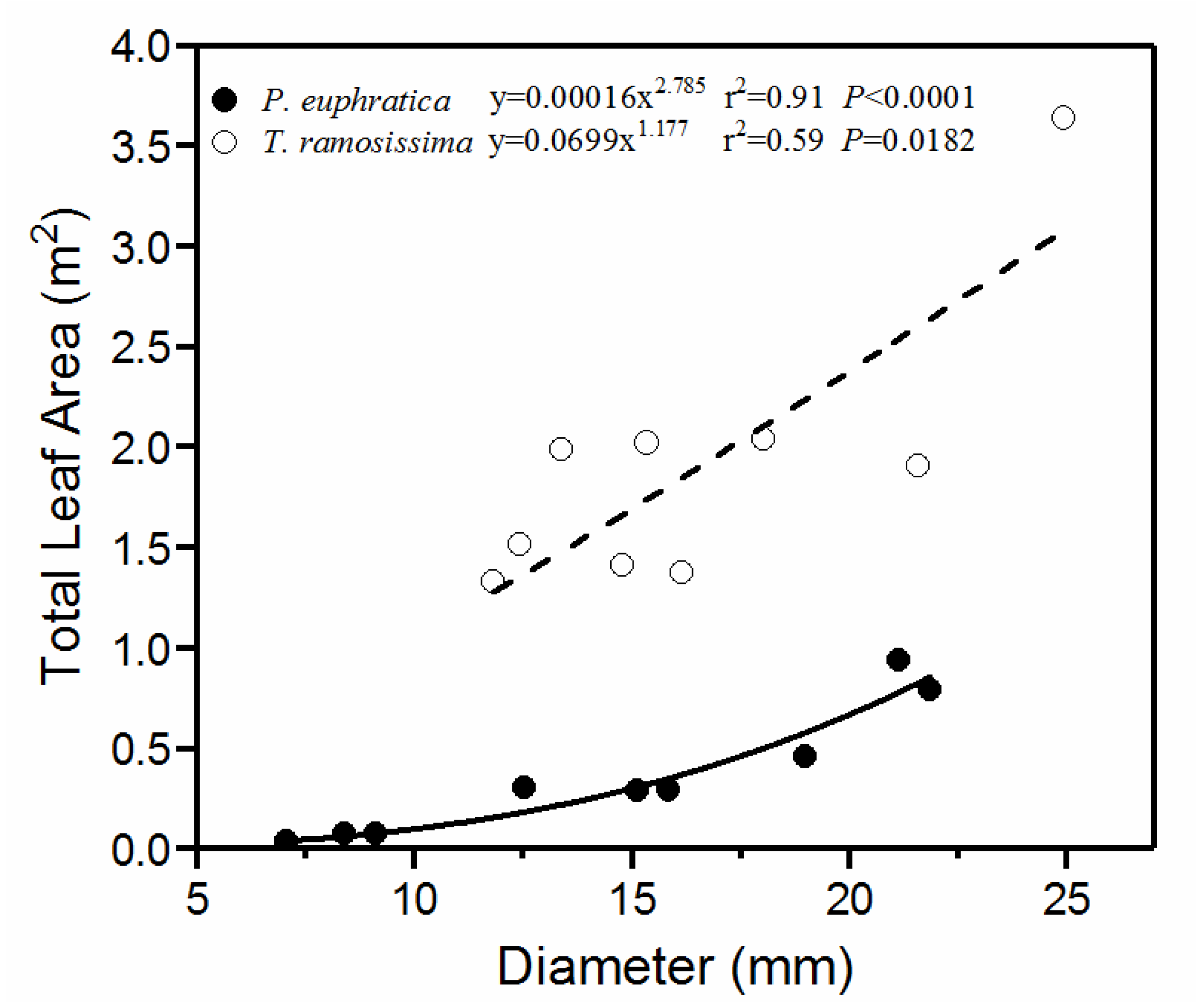
The relationship between TLA and basal stem diameter followed a power function model for both species (Figure 4). Within the observed diameter range, the TLA of T. ramosissima increased at a faster rate with diameter than that of P. euphratica. For each diameter, the TLA of T. ramosissima was several orders of magnitude higher than that of P. euphratica. Therefore, the stems of T. ramosissima supported higher leaf area than those of P. euphratica.
At the end of experiment, we found that both species, under 20 cm groundwater depth, had a shallow rooting depth less than 50 cm (Table 1), with abundant absorbing roots distributed surrounding the groundwater level [34]. P. euphratica root reached groundwater level under 60 cm groundwater depth, but not under 100 cm groundwater depth. In comparison, T. ramosissima roots under both 60 cm and 100 cm groundwater depths reached the bottom of lysimeters (Table 1), with abundant absorbing roots distributed in the cobble layer in the lysimeters (observations), which indicates T. ramosissima had a greater root growth rate to acclimate to groundwater alterations.
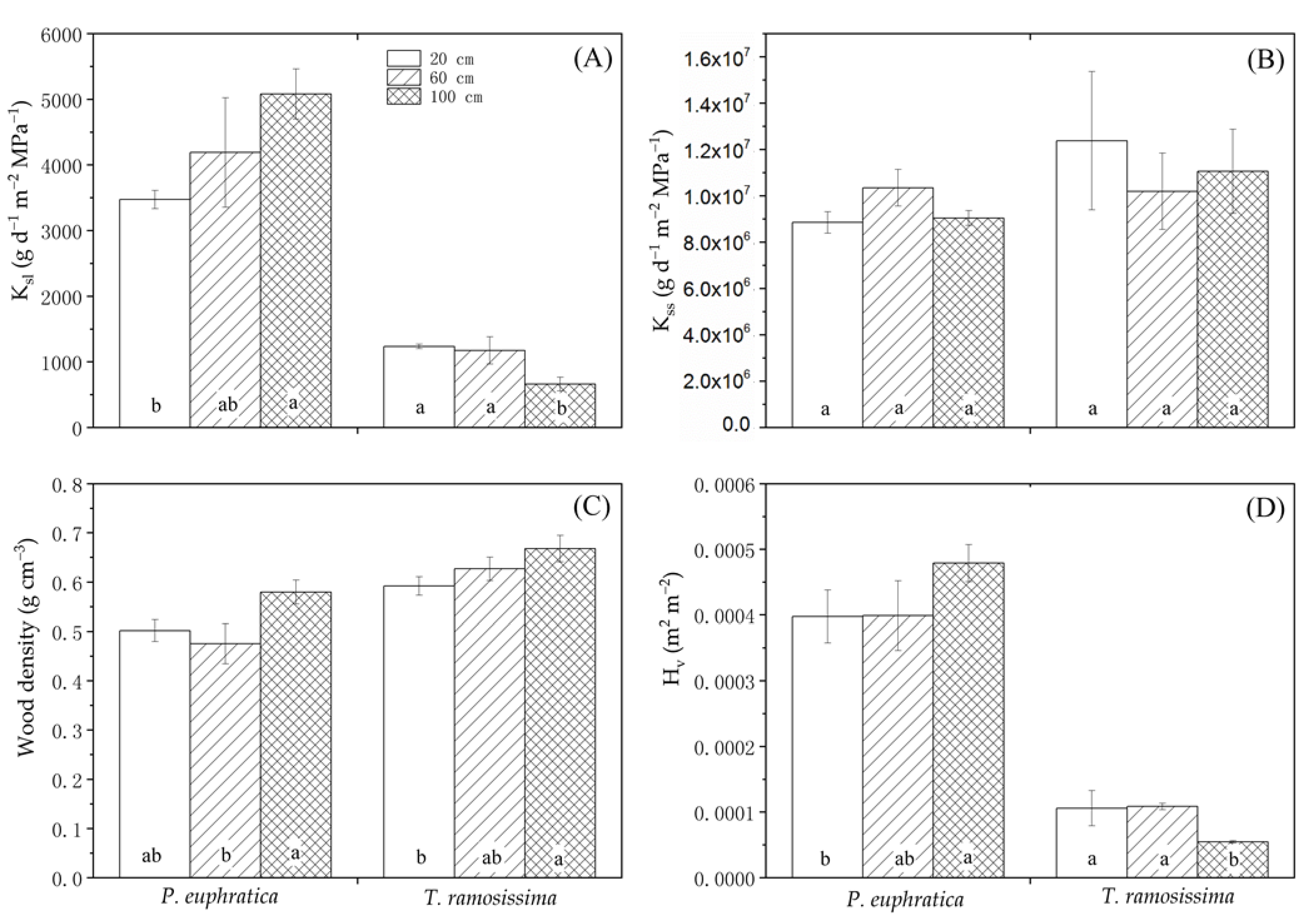
There were significant species effects on wood density, Hv, ksl, and stomatal sensitivity to LVPD (Table 2). P. euphratica had higher ksl and Hv and lower wood density than T. ramosissima (Figure 5). The treatment effect was significant only for wood density. There was a significant species × treatment interaction for Hv and ksl. This observation indicated different hydraulic architecture and functional responses of the species to groundwater variations.
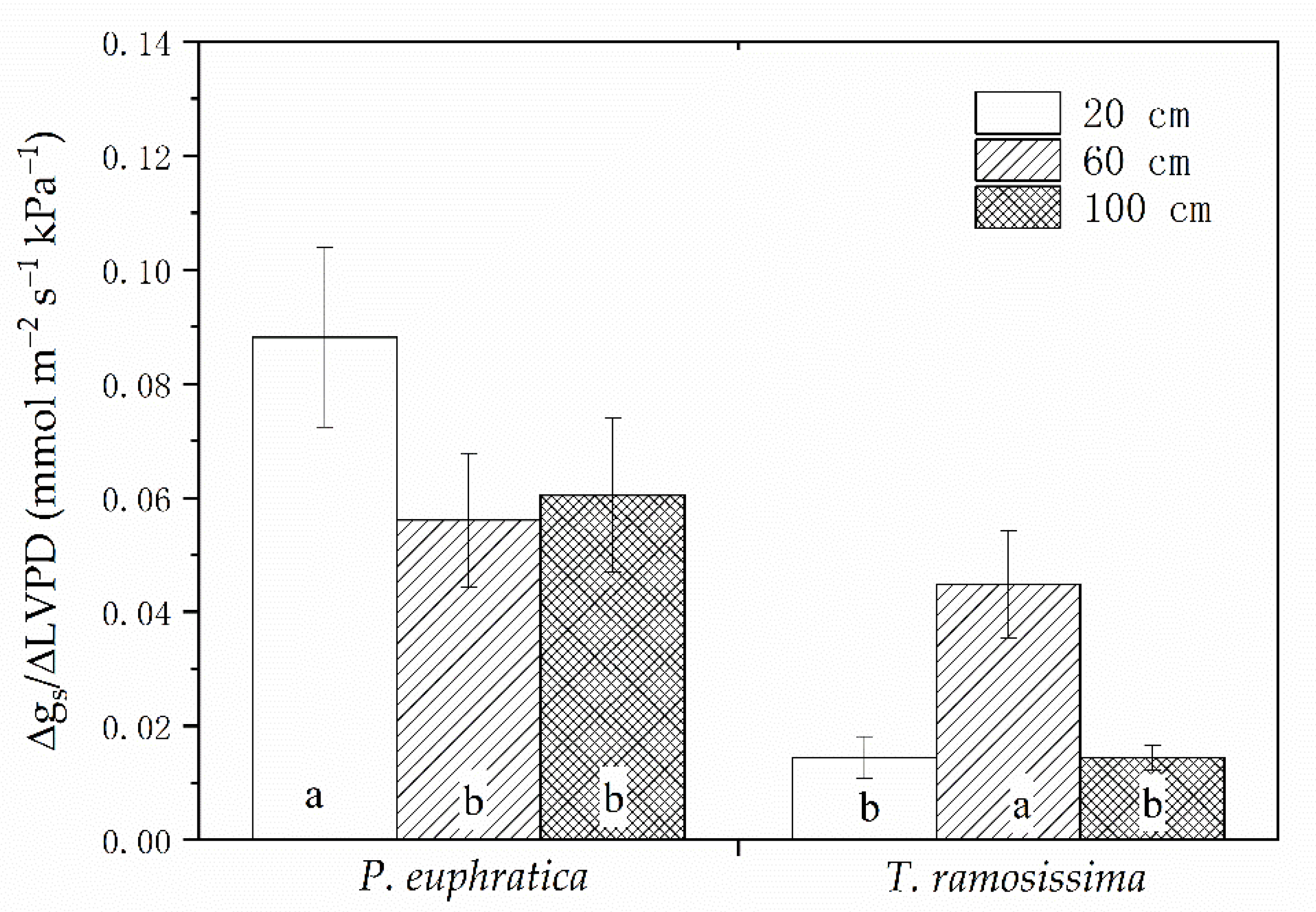
The variables had different change patterns between treatments for each species. The ksl was highest at 100 cm groundwater depth for P. euphratica but was still significantly (p < 0.05) lower than that that for T. ramosissima in shallow groundwater (Figure 5A). For each species, there were no significant differences among treatments in terms of kss (Figure 5B). There were similar patterns between treatments in both species with regard to wood density. It was significantly (p < 0.05) higher at 100 cm groundwater depth than it was for the shallower groundwater treatments (Figure 5C). The pattern of Hv was similar to that for ksl. P. euphratica seedlings at 100 cm groundwater depth had higher ksl than other treatments but the opposite was true for T. ramosissima (Figure 5D). At 20 cm groundwater depth, Δgs/ΔLVPD for P. euphratica was significantly (p < 0.05) greater than it was for the deeper groundwater treatments. In contrast, the highest Δgs/ΔLVPD for T. ramosissima was measured in the 60 cm groundwater depth treatment (Figure 6).
4. Discussion
Our study compared whole-plant water use and relevant hydraulic traits for Populus euphratica and Tamarix ramosissima at simulated groundwater depth conditions. T. ramosissima was found to have greater whole-plant water use than P. euphratica, even under ample groundwater conditions supply (Figure 2). A linear correlation between PET-normalized daily water use and TLA shows that the water use of both phreatophytes was highly dependent on TLA. Nevertheless, the total leaf area of T. ramosissima was only weakly correlated with PET-normalized water use (Figure 3). This species had a comparatively higher leaf-area maintenance per unit sapwood area (Figure 4), which could be promoted by its faster root growth to access declined groundwater (Table 1). Therefore, leaf area-based water use is relatively less constrained by the total leaf area at the individual level. This finding is consistent with the observation that T. ramosissima has a very high water demand due to its high leaf area index [8]. The whole-plant kss of P. euphratica was comparable to that of T. ramosissima but its ksl was relatively larger (Figure 5). For this reason, the difference in leaf area-based water use between species was explained by the inequalities in their leaf-specific and not sapwood-specific hydraulic efficiencies. The relatively higher Hv in P. euphratica (Figure 5D) suggests that comparatively less leaf area was maintained by each sapwood area. The higher Hv favored a higher ksl in P. euphratica (Figure 5A). The interspecific variations in ksl were driven by shoot leaf-sapwood allocation rather than by xylem hydraulic efficiency (kss) in both P. euphratica and T. ramosissima.
P. euphratica and T. ramosissima must efficiently transport water from the groundwater to the leaves despite the high transpirational demand of their hyperarid environment [49]. Rooting depth differed between species and treatments in this study (Table 1). Therefore, groundwater treatments substantially influence water transport resistance at the soil–root interface, where the most resistance occurs along the whole SPAC system [50]. Differences in plant height (0.9–1.5 m for P. euphratica and 1.1–1.4 m for T. ramosissima [36]) are comparatively minor sources of water transport resistance. The soil-to-leaf path length contributes little to the differences in hydraulic behavior because it has only a slight impact on the gravitational potential of water (0.01 MPa m−1) in plants. Moreover, vascular structure is not able to change much in a short growing season. Thus, our observations reflected the adaptions in hydraulic architecture (leaf area-sapwood area) and water transportation (stomatal and xylem conductance) in response to variations in groundwater availability.
In general, most of the observed parameters indicated a divergence in hydraulic behavior between coexisting species. Lower ksl was associated with lower Hv in T. ramosissima. This observation is consistent with the strong correlation between these two variables [51]. Since kss is unable to compensate for a lower ksl driven by Hv, T. ramosissima would present with relatively steeper xylem pressure gradients at each transpiration rate, supported by the observation that T. ramosissima has greater resistance to embolism than P. euphratica [39]. In fact, T. ramosissima reduced xylem water potential to more negative levels than P. euphratica [38]. In contrast, P. euphratica is extremely vulnerable to cavitation, as are certain other poplar species. It has a small hydraulic safety margin [52] and is therefore prone to a functional loss of xylem conductivity under high transpirational demand, even when the groundwater is shallow [53]. P. euphratica mitigates the risk of embolism by maintaining high ksl via elevated Hv. In this way, it reduces the water potential gradient generated by transpiration, whereas T. ramosissima avoids embolism by developing a safer xylem.
Stomatal behavior may have evolved from the coordination of stomatal control and soil-to-leaf water transport capacity [54]. P. euphratica stomata were relatively more sensitive to increases in LVPD than those of T. ramosissima (Figure 6). This observation was consistent with the fact that gs declines sharply with increasing LVPD [38]. In contrast, the stomata of T. ramosissima were relatively indifferent to increasing LVPD. Consequently, T. ramosissima maintains a comparatively higher gs under high transpirational demand than P. euphratica. Even with ample groundwater, P. euphratica cannot compensate for water loss under high transpirational demand as effectively as T. ramosissima because stomatal response to LVPD depends upon foliar water loss [55]. Stomatal regulation and other hydraulic traits influence carbon and water balances in plants [56] and interact with environmental water availability, ultimately influencing the distribution of plant species [57]. Though the sensitive stomatal response to LVPD in P. euphratica may control water loss, in the case of P. euphratica stands with groundwater hardly recharged by runoff, the consequent stomatal closure would ultimately lead to plant dieback due to carbon starvation. This can help us to understand the decline in P. euphratica stands far away from channel [58]. Correspondingly, the relatively greater hydraulic safety in T. ramosissima may account for its occurrence in a wide range of groundwater depths [39].
Within species, all factors except kss varied among treatments (Figure 5). Therefore, hydraulic plasticity is vital for adaption to fluctuations in groundwater depth. High wood density in trees growing where the groundwater is deep may ensure hydraulic safety when water availability is low. A negative correlation between wood density and xylem hydraulic efficiency has often been reported [59]. With increasing groundwater depth, P. euphratica maintains high ksl by increasing Hv. This response is consistent with observed increases in Hv to compensate for declining water availability [60]. In contrast, T. ramosissima decreased ksl in areas where the groundwater was deep. This response must have been the result of a decrease in Hv since kss did not change (Figure 5B). Tamarix invests in foliage growth [61], which disproportionately increases leaf area relative to sapwood area. This mechanism may explain the decreases in ksl and Hv in T. ramosissima growing where the groundwater was deep. It is unknown why P. euphratica has relatively higher stomatal sensitivity to LVPD when it grows over shallow groundwater, whereas stomatal sensitivity to LVPD is comparatively high for T. ramosissima growing at moderate groundwater depth. This discrepancy may be elucidated by a thorough understanding of the diurnal changes which occur in the balance between leaf water supply and transpiration [62].
5. Conclusions
In the present study, we investigated whole-plant water use and hydraulic traits in two phreatophytes on the Tarim River floodplain adapting to groundwater variations. T. ramosissima was found to have greater whole-plant water use regardless of groundwater conditions, which can be attributed to several hydraulic traits such as fast root growth, higher leaf area maintenance per sapwood area, indifferent stomatal response to LVPD, and greater hydraulic safety margin. While kss is similar between species, Hv is greatly lower in T. ramosissima, indicating that the significantly lower ksl was driven by Hv. Increased ksl and Hv in P. euphratica grown in deep groundwater favor water loss control, but limit plant growth, indicating P. euphratica is not as capable as T. ramosissima to use deep groundwater. Our findings provide an understanding of the expansion of T. ramosissima in areas with groundwater decline in the Tarim River Basin.
Author Contributions
Conceptualization, J.L.; Funding acquisition, J.L.; Investigation, Y.S.; Methodology, J.L. and S.H.; Writing—original draft, J.L.; Writing—review and editing, S.H., Y.S. and X.H. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by grant from the National Natural Science Foundation of China (No. 41171037).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data is available from the corresponding author upon reasonable request.
Acknowledgments
The authors thank Yong Zhang and Bin Huang for their valuable technical assistance and maintenance of the experimental facility.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thomas, F.M. Ecology of Phreatophytes; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Chen, Y.; Chen, Y.; Zhou, H.; Hao, X.; Zhu, C.; Fu, A.; Yang, Y.; Li, W. Research Advances in Plant Physiology and Ecology of Desert Riparian Forests under Drought Stress. Forests 2022, 13, 619. [Google Scholar] [CrossRef]
- Melissa, M.R.; John, C.S.; Dar, A.R.; Michael, B.S. Groundwater dependence of riparian woodlands and the disrupting effect of anthropogenically altered streamflow. Proc. Natl. Acad. Sci. USA 2021, 118, e2026453118. [Google Scholar]
- Stromberg, J.C.; Tiller, R.; Richter, B. Effects of groundwater decline on riparian vegetation of semiarid regions: The San Pedro, Arizona. Ecol. Appl. 1996, 6, 113–131. [Google Scholar] [CrossRef] [Green Version]
- Corenblit, D.; Steiger, J.; Gurnell, A.M.; Naiman, R.J. Plants intertwine fluvial landform dynamics with ecological succession and natural selection: A niche construction perspective for riparian systems. Glob. Ecol. Biogeogr. 2009, 18, 507–520. [Google Scholar] [CrossRef]
- Stromberg, J.C.; Lite, S.J.; Marler, R.; Paradzick, C.; Shafroth, P.B.; Shorrock, D.; White, J.M.; White, M.S. Altered stream-flow regimes and invasive plant species: The Tamarix case. Glob. Ecol. Biogeogr. 2007, 16, 381–393. [Google Scholar] [CrossRef]
- Merritt, D.M.; Poff, N.L. Shifting dominance of riparian Populus and Tamarix along gradients of flow alteration in western North American rivers. Ecol. Appl. 2010, 20, 135–152. [Google Scholar] [CrossRef] [Green Version]
- Sala, A.; Smith, S.D.; Devitt, D.A. Water use by Tamarix ramosissima and associated phreatophytes in a Mojave desert floodplain. Ecol. Appl. 1996, 6, 888–898. [Google Scholar] [CrossRef]
- Davis, M.A.; Pergl, J.; Robinson, A.M. Vegetation change: A reunifying concept in plant ecology. Perspect. Plant Ecol. Ecol. Syst. 2005, 7, 69–76. [Google Scholar] [CrossRef]
- Newman, B.D.; Wilcox, B.P.; Archer, S.R.; Breshears, D.D.; Dahm, C.N.; Duffy, C.J.; McDowell, N.G.; Phillips, F.M.; Scanlon, B.R.; Vivoni, E.R. Ecohydrology of water-limited environments: A scientific vision. Water Resour. Res. 2006, 42, W06302. [Google Scholar] [CrossRef]
- Lama, G.F.C.; Giovannini, M.R.M.; Errico, A.; Mirzaei, S.; Padulano, R.; Chirico, G.B.; Preti, F. Hydraulic Efficiency of Green-Blue Flood Control Scenarios for Vegetated Rivers: 1D and 2D Unsteady Simulations. Water 2021, 13, 2620. [Google Scholar] [CrossRef]
- Lu, Z.; Zhao, Y.; Wei, Y.; Feng, Q.; Xie, J. Differences among evapotranspiration products affect water resources and ecosystem management in an australian catchment. Remote Sens. 2019, 11, 958. [Google Scholar] [CrossRef] [Green Version]
- Carpintero, E.; Andreu, A.; Gómez-Giráldez, P.J.; Blázquez, Á.; González-Dugo, M.P. Remote-sensing-based water balance for monitoring of evapotranspiration and water stress of a Mediterranean Oak–Grass Savanna. Water 2020, 12, 1418. [Google Scholar] [CrossRef]
- McDowell, N.G.; Beerling, D.J.; Breashers, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.D.; Devitt, D.A.; Sala, A.; Cleverly, J.R.; Busch, D.E. Water relations of riparian plants from warm desert regions. Wetlands 1998, 18, 678–696. [Google Scholar] [CrossRef]
- Cleverly, J.; Smith, S.; Sala, A.; Devitt, D. Invasive capacity of Tamarix ramosissima in a Mohave Desert floodplain: The role of drought. Oecologia 1997, 111, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Elmore, A.J.; Manning, S.J.; Mustard, J.F.; Craine, J.M. Decline in alkali meadow vegetation cover in California: The effects of groundwater extraction and drought. J. Appl. Ecol. 2006, 43, 770–779. [Google Scholar] [CrossRef]
- Cleverly, J.R.; Dahm, C.N.; Thibault, J.R.; McDonnell, D.E.; Coonrod, J.E.A. Riparian ecohydrology: Regulation of water flux from the ground to the atmosphere in the Middle Rio Grande, New Mexico. Hydrol. Process. 2006, 20, 3207–3225. [Google Scholar] [CrossRef] [Green Version]
- O’Grady, A.P.; Cook, P.G.; Eamus, D.; Duguid, A.; Wischusen, J.D.H.; Fass, T.D. Worldege. Convergence of tree water use within and arid-zone woodland. Oecologia 2009, 160, 643–655. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Poyatos, R.; Aguadé, D.; Retana, J.; Mencuccini, M. A new look at water transport regulation in plants. New Phytol. 2014, 204, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Sperry, J.S.; Hacke, U.G.; Oren, R.; Comstock, J.P. Water deficits and hydraulic limits to leaf water supply. Plant Cell Environ. 2002, 25, 251–263. [Google Scholar] [CrossRef]
- Tyree, M.T.; Zimmermann, M.H. Xylem Structure and the Ascent of Sap; Springer: Berlin/Heidelber, Germany, 2002. [Google Scholar]
- Tyree, M.T.; Ewers, F.W. The hydraulic architecture of trees and other woody plants. New Phytol. 1991, 119, 345–360. [Google Scholar] [CrossRef]
- Reich, P.B.; Wright, I.J.; Cavender-Bares, J.; Craine, J.M.; Oleksyn, J.; Westoby, M.; Walters, M.B. The evolution of plant functional variation: Traits, spectra, and strategies. Int. J. Plant Sci. 2003, 164, S143–S164. [Google Scholar] [CrossRef]
- Meinzer, F.C. Functional convergence in plant responses to the environment. Oecologia 2003, 134, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bucci, S.J.; Goldstein, G.; Meinzer, F.C.; Scholz, F.G.; Rranco, A.C.; Bustamante, M. Functional convergence in hydraulic architecture and water relations of tropical savanna stress: From leaf to whole plant. Tree Physiol. 2004, 24, 891–899. [Google Scholar] [CrossRef] [Green Version]
- Gharum, M.; Turnbull, T.L.; Pfautsch, S.; Adams, M.A. Stomatal structure and physiology do not explain differences in water use among montane eucalypts. Oecologia 2015, 177, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.D.; Song, J.J.; Li, R.H.; Ye, Q. Plant hydraulics and photosynthesis of 34 woody species from different successional stages of subtropical forests. Plant Cell Environ. 2013, 36, 879–891. [Google Scholar] [CrossRef]
- Buckley, T.N.; Sack, L.; Farquhar, G.D. Optimal plant water economy. Plant Cell Environ. 2017, 40, 881–896. [Google Scholar] [CrossRef] [Green Version]
- Imin, B.; Dai, Y.; Shi, Q.; Guo, Y.; Li, H.; Nijat, M. Responses of two dominant desert plant species to the changes in groundwater depth in hinterland natural oasis, Tarim Basin. Ecol. Evol. 2020, 11, 9460–9471. [Google Scholar] [CrossRef]
- Chen, Y.N.; Pang, Z.H.; Chen, Y.P.; Li, W.H.; Xu, C.C.; Hao, X.M.; Huang, X.; Huang, T.M.; Ye, Z.X. Response of riparian vegetation to water-table changes in the lower reaches of Tarim River, Xinjiang Uygur, China. Hydrogeol. J. 2008, 16, 1371–1379. [Google Scholar] [CrossRef]
- Fang, G.; Yang, J.; Chen, Y.; Li, Z.; Ji, H.; De Maeyer, P. How hydrologic processes differ spatially in a large basin: Multisite and multi objective modeling in the Tarim River Basin. J. Geophys. Res. Atmos. 2018, 123, 7098–7113. [Google Scholar] [CrossRef]
- Zhang, X.W.; Cheng, T.F.; Chen, H.W.; Tian, X.M. Underground water monitoring and analysis on Tarim River Basin. J. Shihezi Univ. Natl. Sci. 2007, 25, 364–368. [Google Scholar]
- Zhu, Y.; Chen, Y.; Ren, L.; Lv, H.; Zhao, W.; Yuan, F.; Xu, M. Ecosystem restoration and conservation in the arid inland river basins of Northwest China: Problems and strategies. Ecol. Eng. 2016, 94, 629–637. [Google Scholar] [CrossRef]
- Huang, T.M.; Pang, Z.H. Changes in groundwater induced by water diversion in the Lower Tarim River, Xinjiang Uygur, NW China: Evidence from environmental isotopes and water chemistry. J. Hydrol. 2010, 387, 188–201. [Google Scholar] [CrossRef]
- Li, J.; Yu, B.; Zhao, C.; Nowak, R.S.; Zhao, Z.; Sheng, Y.; Li, J. Physiological and morphological responses of Tamarix ramosissima and Populus euphratica to altered groundwater availability. Tree Physiol. 2013, 33, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Aasamaa, K.; Sõber, A. Stomatal sensitivities to changes in leaf water potential, air humidity, CO2 concentration and light intensity, and the effect of abscisic acid on the sensitivities in six temperate deciduous tree species. Environ. Exp. Bot. 2011, 71, 72–78. [Google Scholar] [CrossRef]
- Thomas, F.M.; Foetzki, A.; Gries, D.; Bruelheide, H.; Li, X.; Zeng, F.; Zhang, X.M. Regulation of the water status in three co-occurring phreatophytes at the southern fringe of the Taklamakan Desert. J. Plant Ecol. 2008, 1, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Chen, Y.; Li, W.; Ayup, M. Xylem hydraulic conductivity and embolism in riparian plants and their responses to drought stress in desert of Northwest China. Ecohydrology 2013, 6, 984–993. [Google Scholar] [CrossRef]
- Wu, G.; Jiang, S.; Liu, W.; Zhao, C.; Li, J. Competition between Populus euphratica and Tamarix ramosissima seedlings under simulated high groundwater availability. J. Arid Land. 2016, 8, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Li, W.; Zhou, H.; Chen, Y.; Hao, X.; Fu, A.; Ma, J. Experimental study on water transport observations of desert riparian forests in the lower reaches of the Tarim River in China. Int. J. Biometeorol. 2017, 61, 1055–1062. [Google Scholar] [CrossRef]
- Pivovaroff, A.L.; Cook, V.M.W.; Santiago, L.S. Stomatal behaviour and stem xylem traits are coordinated for woody plant species under exceptional drought conditions. Plant Cell Environ. 2018, 41, 2617–2626. [Google Scholar] [CrossRef]
- Yin, Q.; Wang, L.; Lei, M.; Dang, H.; Quan, J.; Tian, T.; Chai, Y.; Yue, M. The relationships between leaf economics and hydraulic traits of woody plants depend on water availability. Sci. Total Environ. 2018, 621, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements-FAO Irrigation and Drainage Paper 56; FAO: Rome, Italy, 1998. [Google Scholar]
- Van Genuchten, M.T. A closed form equation for predicting the hydraulic conductivity of unsaturated soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef] [Green Version]
- Mualem, Y. A new model for predicting the hydraulic conductivity of unsaturated porous media. Water Resour. Res. 1976, 12, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Jury, W.A.; Gardner, W.R.; Gardner, W.H. Soil Physics; John Wiley: New York, NY, USA, 1991. [Google Scholar]
- Fichot, R.; Chamaillard, S.; Depardieu, C.; Thiec, D.L.; Cochard, H.; Barigah, T.S.; Brignolas, F. Hydraulic efficiency and coordination with xylem resistance to cavitation, leaf function, and growth performance among eight unrelated Populus deltoides×Populus nigra hybrids. J. Exp. Bot. 2011, 62, 2093–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rzepecki, A.; Zeng, F.; Thomas, F.M. Xylem anatomy and hydraulic conductivity of three co-occurring desert phreatophtes. J. Arid Environ. 2011, 75, 338–345. [Google Scholar] [CrossRef]
- Nardini, A.; Salleo, S. Limitation of stomatal conductance by hydraulic traits: Sensing or preventing xylem cavitation? Trees 2000, 15, 14–24. [Google Scholar] [CrossRef]
- Edwards, E.J. Correlated evolution of stem and leaf hydraulic traits in Pereskia (Cactaceae). New Phytol. 2006, 172, 479–489. [Google Scholar] [CrossRef]
- Hukin, D.; Cochard, H.; Dreyer, E.; Le Thiec, D.; Bogeat-Triboulot, M.B. Cavitation vulnerability in roots and shoots: Does Populus euphratica Oliv., a poplar from arid areas of Central Asia, differ from other poplar species? J. Exp. Bot. 2005, 56, 2003–2010. [Google Scholar] [CrossRef]
- Jiang, S.W.; Zhou, D.D.; Wu, G.L.; Li, J. Hydraulic conductivity and its seasonal variation of Populus euphratica shoot at the sites with varying groundwater depths. Arid Zone Res. 2017, 34, 648–654. [Google Scholar]
- Manzoni, S.; Vico, G.; Katul, G.; Palmroth, S.; Jackson, R.B.; Porporato, A. Hydraulic limits on maximum plant transpiration and the emergence of the safety-efficiency trade-off. New Phytol. 2013, 198, 169–178. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Hinckley, T.M.; Ceulemans, R. Apparent responses of stomata to transpiration and humidity in a hybrid poplar canopy. Plant Cell Environ. 1997, 20, 1301–1308. [Google Scholar] [CrossRef]
- Santiago, L.S.; Goldstein, G.; Meinzer, F.C.; Fisher, J.B.; Machado, K.; Woodruff, D.; Jones, T. Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees. Oecologia 2004, 140, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Pockman, W.T.; Sperry, J.S. Vulnerability to xylem cavitation and the distribution of Sonoran desert vegetation. Am. J. Bot. 2000, 87, 1287–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Q.; Liu, W.; Si, J.H.; Su, Y.H.; Zhang, Y.W.; Cang, Z.Q.; Xi, H.Y. Environmental effects of water resource development and use in the Tarim River basin of northwestern China. Environ. Geol. 2005, 48, 202–210. [Google Scholar]
- Hacke, U.G.; Sperry, J.S.; Pockman, W.T.; Davis, S.D.; McCulloh, K.A. Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 2001, 126, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Magnani, F.; Grace, J.; Borghetti, M. Adjustment of tree structure in response to the environment under hydraulic constraints. Funct. Ecol. 2002, 16, 385–393. [Google Scholar] [CrossRef]
- Glenn, E.P.; Nagler, P.L. Comparative ecophysilolgy of Tamarix ramosissima and native trees in western U.S. riparian zones. J. Arid Environ. 2005, 61, 419–446. [Google Scholar] [CrossRef]
- McCulloh, K.A.; Woodruff, D.R. Linking stomatal sensitivity and whole-tree hydraulic architecture. Tree Physiol. 2012, 32, 369–372. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Populus euphratica (a) and Tamarix ramosissima (b) growing in lysimeters.

Figure 2.
Daily PET (mm d−1) and water use (g d−1) per lysimeter with Populus euphratica and Tamarix ramosissima seedlings grown 20 cm, 60 cm, and 100 cm from the groundwater level.
Figure 2.
Daily PET (mm d−1) and water use (g d−1) per lysimeter with Populus euphratica and Tamarix ramosissima seedlings grown 20 cm, 60 cm, and 100 cm from the groundwater level.

Figure 3.
Relationships between PET (mm d-1) normalized daily water use (g d-1) and whole-plant leaf area in Populus euphratica and Tamarix ramosissima grown at a groundwater level gradient.
Figure 3.
Relationships between PET (mm d-1) normalized daily water use (g d-1) and whole-plant leaf area in Populus euphratica and Tamarix ramosissima grown at a groundwater level gradient.

Figure 4.
Relationship between leaf area and basal stem diameter in Populus euphratica and Tamarix ramosissima individuals.
Figure 4.
Relationship between leaf area and basal stem diameter in Populus euphratica and Tamarix ramosissima individuals.

Figure 5.
Whole-plant leaf area-specific hydraulic conductance (A), sapwood-area hydraulic conductance (B), wood densities (C), and Huber values (D) for Populus euphratica and Tamarix ramosissima seedlings grown 20 cm, 60 cm, and 100 cm from the groundwater level. Values are mean ± SE. Letters indicate significant differences among treatments for each species as determined by ANOVA (p < 0.05).
Figure 5.
Whole-plant leaf area-specific hydraulic conductance (A), sapwood-area hydraulic conductance (B), wood densities (C), and Huber values (D) for Populus euphratica and Tamarix ramosissima seedlings grown 20 cm, 60 cm, and 100 cm from the groundwater level. Values are mean ± SE. Letters indicate significant differences among treatments for each species as determined by ANOVA (p < 0.05).

Figure 6.
Stomatal conductance in response to increases in leaf-to-air vapor pressure deficit (LVPD) expressed as the ratio of Δgs to ΔLVPD. Values are mean ± SE. Letters indicate significant differences among treatments for each species as determined by ANOVA (p < 0.05).
Figure 6.
Stomatal conductance in response to increases in leaf-to-air vapor pressure deficit (LVPD) expressed as the ratio of Δgs to ΔLVPD. Values are mean ± SE. Letters indicate significant differences among treatments for each species as determined by ANOVA (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Rooting depths (cm) of P. euphratica and T. ramosissima grown at 20 cm, 60 cm, and 100 cm groundwater depths (DGW). Values are means ± SE (n = 3).
Table 1.
Rooting depths (cm) of P. euphratica and T. ramosissima grown at 20 cm, 60 cm, and 100 cm groundwater depths (DGW). Values are means ± SE (n = 3).
| Treatment | 20 DGW | 60 DGW | 100 DGW |
|---|---|---|---|
| P. euphratica | 48.2 ± 1.3 | 87.8 ± 7.8 | 73.9 ± 9.4 |
| T. ramosissima | 47.7 ± 4.6 | bottom reached | bottom reached |
Table 2.
Probabilities determined by ANOVA for species, treatment, and species × treatment effects on wood density, Huber value (Hv), whole-plant leaf area-specific hydraulic conductance (ksl), whole-plant sapwood-specific hydraulic conductance (kss), and stomatal sensitivity to LVPD (Δgs/ΔLVPD).
Table 2.
Probabilities determined by ANOVA for species, treatment, and species × treatment effects on wood density, Huber value (Hv), whole-plant leaf area-specific hydraulic conductance (ksl), whole-plant sapwood-specific hydraulic conductance (kss), and stomatal sensitivity to LVPD (Δgs/ΔLVPD).
| Factor | Species | Treatment | Species × Treatment |
|---|---|---|---|
| wood density | <0.001 | 0.017 | 0.395 |
| Hv | <0.001 | 0.173 | 0.007 |
| ksl | <0.001 | 0.43 | 0.045 |
| kss | 0.104 | 0.847 | 0.684 |
| Δgs/ΔLVPD | <0.001 | 0.238 | 0.101 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, J.; Hu, S.; Sheng, Y.; He, X. Whole-Plant Water Use and Hydraulics of Populus euphratica and Tamarix ramosissima Seedlings in Adaption to Groundwater Variation. Water 2022, 14, 1869. https://doi.org/10.3390/w14121869
AMA Style
Li J, Hu S, Sheng Y, He X. Whole-Plant Water Use and Hydraulics of Populus euphratica and Tamarix ramosissima Seedlings in Adaption to Groundwater Variation. Water. 2022; 14(12):1869. https://doi.org/10.3390/w14121869
Chicago/Turabian StyleLi, Jun, Shunjun Hu, Yu Sheng, and Xiran He. 2022. "Whole-Plant Water Use and Hydraulics of Populus euphratica and Tamarix ramosissima Seedlings in Adaption to Groundwater Variation" Water 14, no. 12: 1869. https://doi.org/10.3390/w14121869
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.