
Empirical Setting of the Water Stressed Baseline Increases the Uncertainty of the Crop Water Stress Index in a Humid Temperate Climate in Different Water Regimes



Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Treatments
2.2. Experimental Measurements
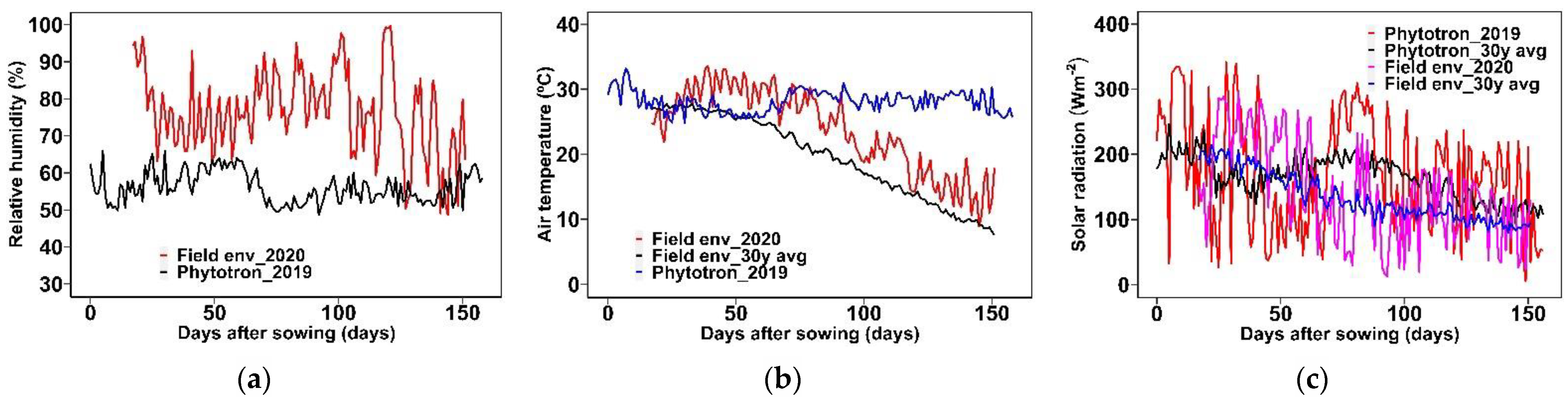
2.2.1. Phytotron
2.2.2. Field Environment
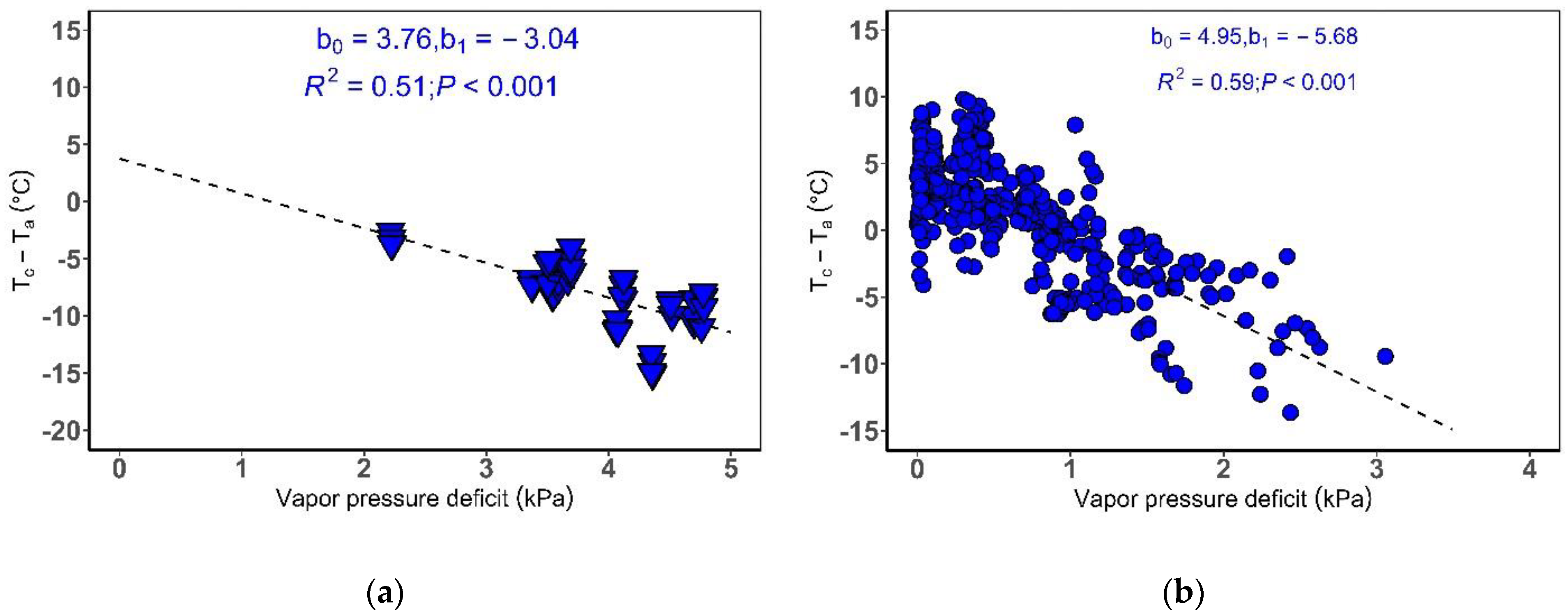
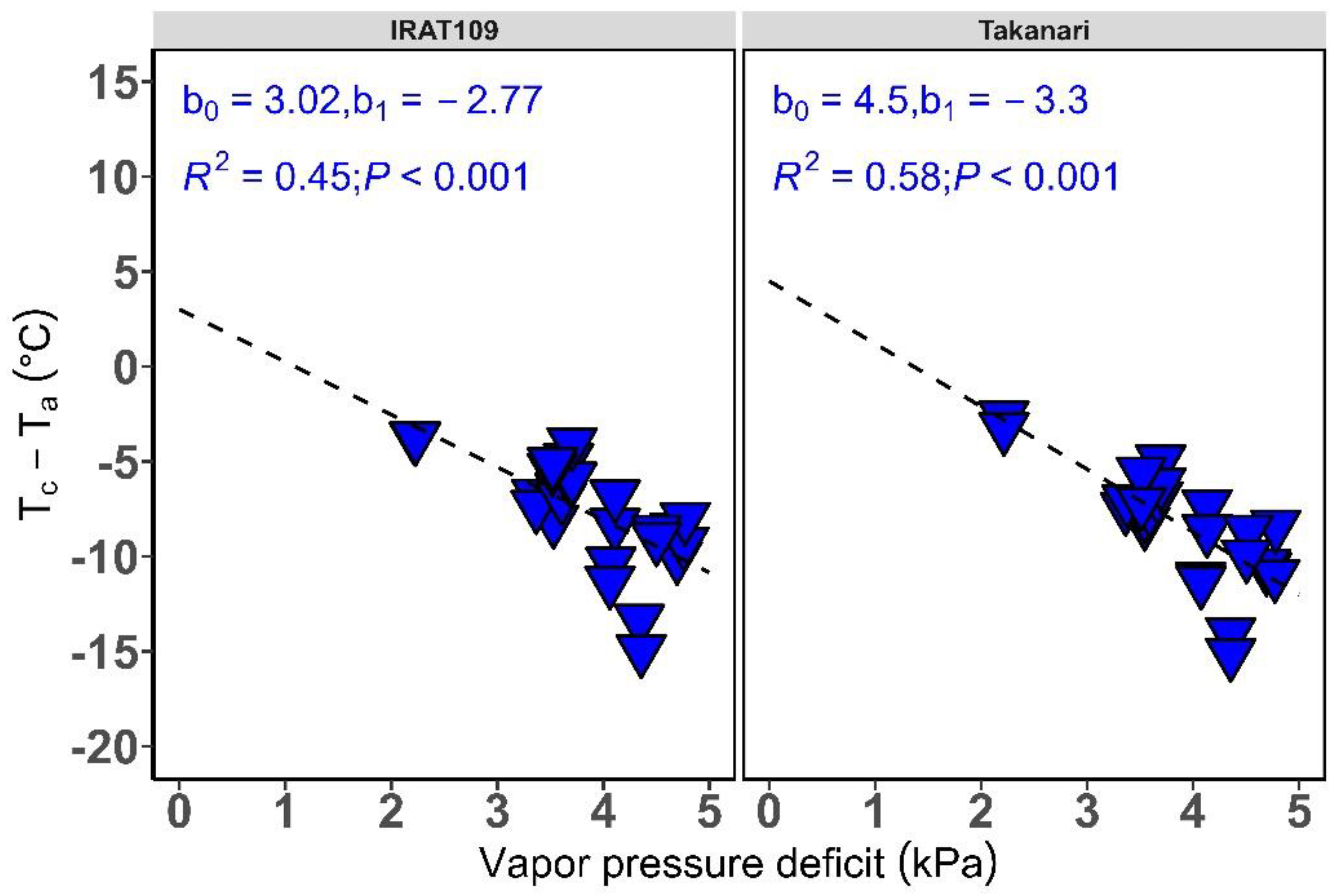
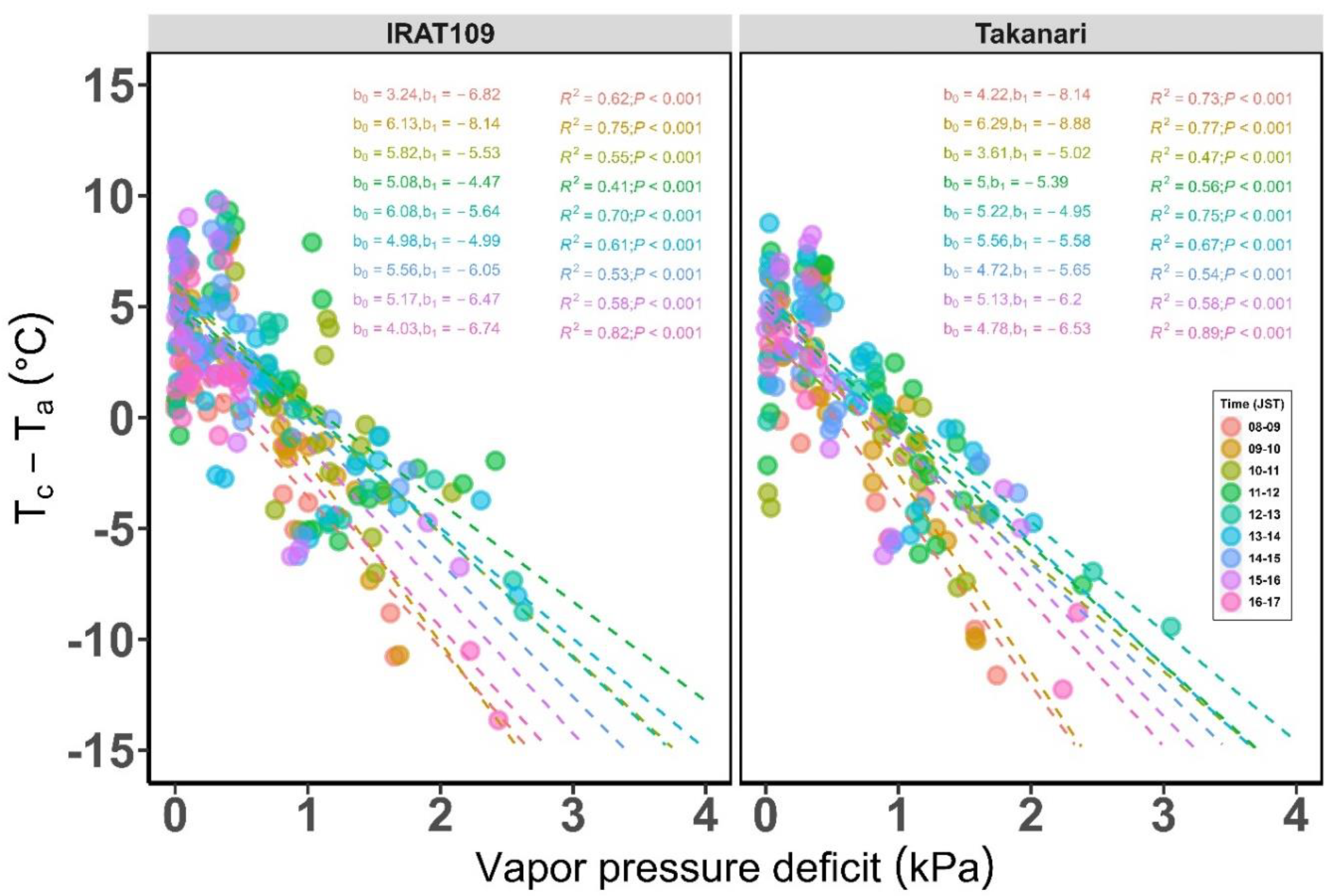
2.3. Determination of Reference Baseline for Rice in a Humid Temperate Climate
2.3.1. Vapor Pressure Deficit (VPD)
2.3.2. Establishing Reference Baseline Parameters
2.4. Determination of Crop Water Stress Index (CWSI)
2.5. Data Analysis
3. Results
3.1. Non-Water Stressed and Water Stressed Reference Baseline for Rice in a Humid Temperate Climate
Phytotron and Field Environment
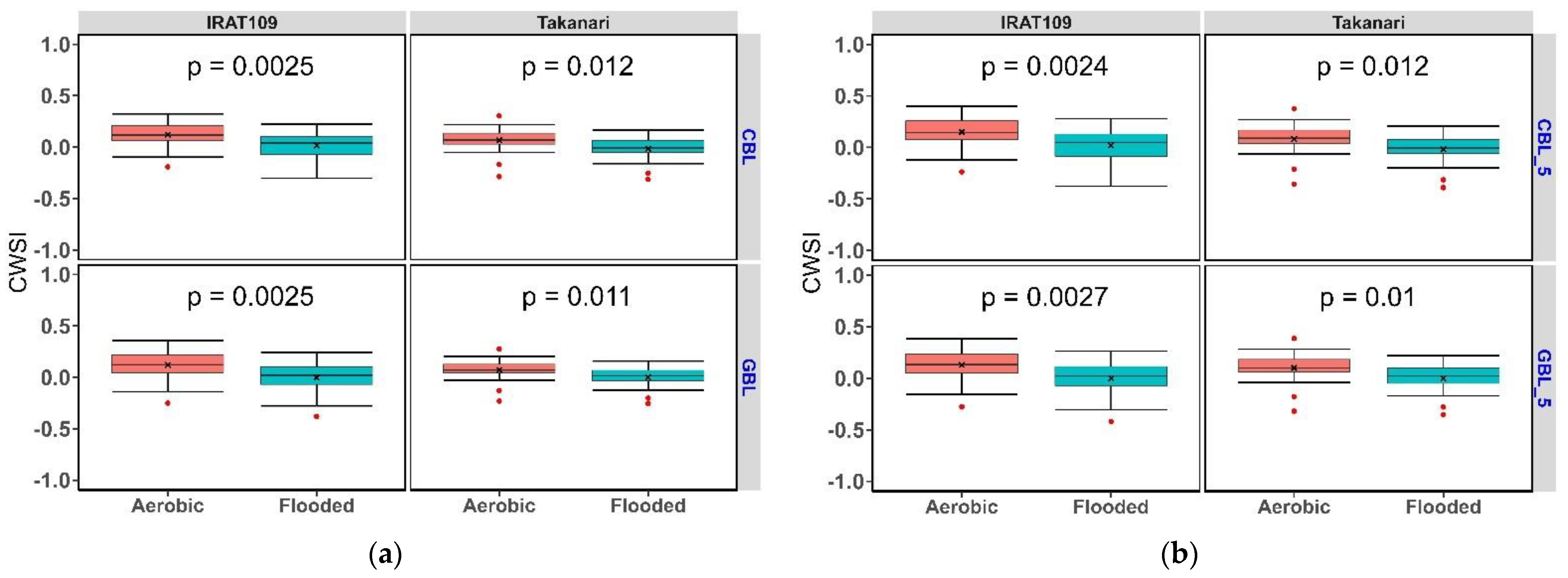
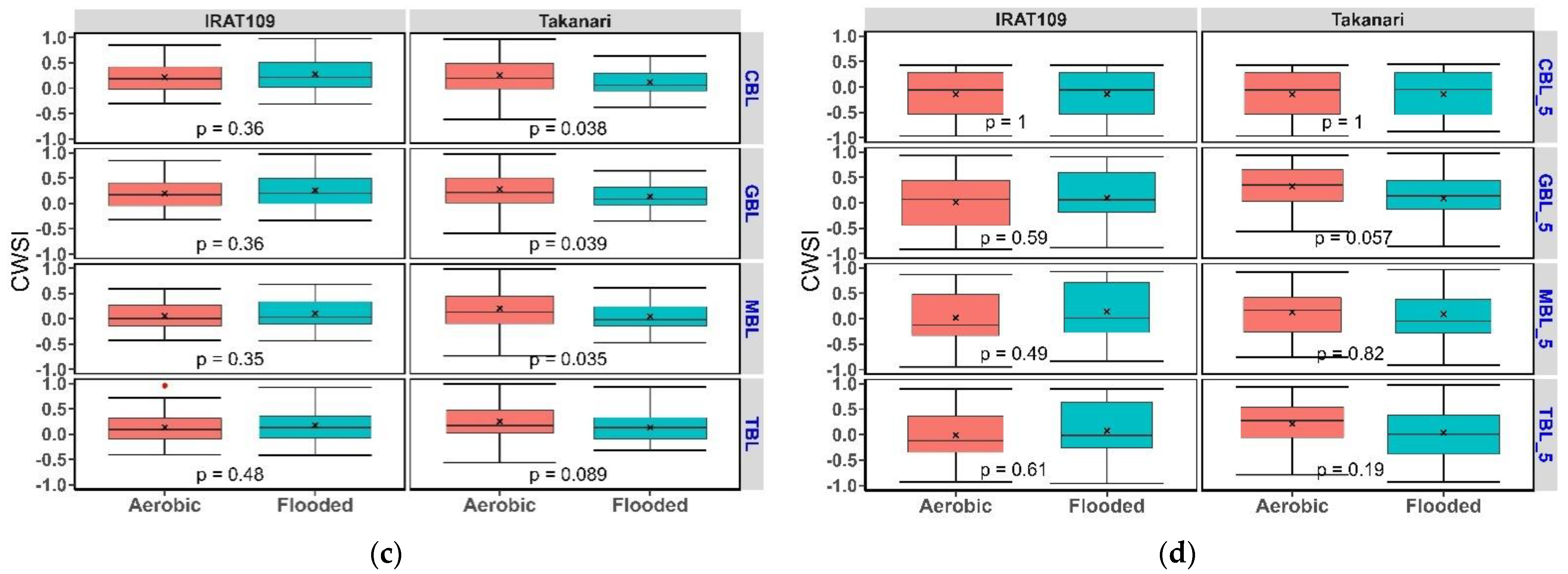
3.2. Crop Water Stress Index in Contrasting Water Regimes and Genotypes
Water Regimes and Genotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elert, E. Rice by the numbers: A good grain. Nature 2014, 514, S50–S51. [Google Scholar] [CrossRef] [Green Version]
- Bouman, B.A.M.; Lampayan, R.; Tuong, T. Water Management in Irrigated Rice: Coping with Water Scarcity; International Rice Research Institute: Los Banos, Philippines, 2007. [Google Scholar]
- Dubois, O. The State of the World’s Land and Water Resources for Food and Agriculture: Managing Systems at Risk; Earthscan: London, UK, 2011. [Google Scholar]
- Lampayan, R.M.; Rejesus, R.M.; Singleton, G.R.; Bouman, B.A. Adoption and economics of alternate wetting and drying water management for irrigated lowland rice. Field Crop. Res. 2015, 170, 95–108. [Google Scholar] [CrossRef]
- Oliver, V.; Cochrane, N.; Magnusson, J.; Brachi, E.; Monaco, S.; Volante, A.; Courtois, B.; Vale, G.; Price, A.; Teh, Y.A. Effects of water management and cultivar on carbon dynamics, plant productivity and biomass allocation in European rice systems. Sci. Total Environ. 2019, 685, 1139–1151. [Google Scholar] [CrossRef]
- Kato, Y.; Okami, M.; Katsura, K. Yield potential and water use efficiency of aerobic rice (Oryza sativa L.) in Japan. Field Crop. Res. 2009, 113, 328–334. [Google Scholar] [CrossRef]
- Peng, S.; Bouman, B.; Visperas, R.M.; Castañeda, A.; Nie, L.; Park, H.-K. Comparison between aerobic and flooded rice in the tropics: Agronomic performance in an eight-season experiment. Field Crop. Res. 2006, 96, 252–259. [Google Scholar] [CrossRef]
- Tsujimoto, Y.; Fuseini, A.; Inusah, B.I.; Dogbe, W.; Yoshimoto, M.; Fukuoka, M. Different effects of water-saving management on canopy microclimate, spikelet sterility, and rice yield in the dry and wet seasons of the sub-humid tropics in northern Ghana. Field Crop. Res. 2020, 260, 107978. [Google Scholar] [CrossRef]
- Khorsand, A.; Rezaverdinejad, V.; Asgarzadeh, H.; Majnooni-Heris, A.; Rahimi, A.; Besharat, S.; Sadraddini, A.A. Linking plant and soil indices for water stress management in black gram. Sci. Rep. 2021, 11, 869. [Google Scholar] [CrossRef]
- Shimono, H.; Okada, M.; Inoue, M.; Nakamura, H.; Kobayashi, K.; Hasegawa, T. Diurnal and seasonal variations in stomatal conductance of rice at elevated atmospheric CO2 under fully open-air conditions. Plant Cell Environ. 2009, 33, 322–331. [Google Scholar] [CrossRef]
- Kato, Y.; Okami, M. Root growth dynamics and stomatal behaviour of rice (Oryza sativa L.) grown under aerobic and flooded conditions. Field Crop. Res. 2010, 117, 9–17. [Google Scholar] [CrossRef]
- Bouman, B.A.M.; Humphreys, E.; Tuong, T.P.; Barker, R. Rice and Water. Adv. Agron. 2007, 92, 187–237. [Google Scholar]
- Agam, N.; Cohen, Y.; Alchanatis, V.; Ben-Gal, A. How sensitive is the CWSI to changes in solar radiation? Int. J. Remote Sens. 2013, 34, 6109–6120. [Google Scholar] [CrossRef]
- DeJonge, K.C.; Taghvaeian, S.; Trout, T.J.; Comas, L. Comparison of canopy temperature-based water stress indices for maize. Agric. Water Manag. 2015, 156, 51–62. [Google Scholar] [CrossRef]
- Taghvaeian, S.; Comas, L.; DeJonge, K.C.; Trout, T.J. Conventional and simplified canopy temperature indices predict water stress in sunflower. Agric. Water Manag. 2014, 144, 69–80. [Google Scholar] [CrossRef]
- Poirier-Pocovi, M.; Bailey, B. Sensitivity analysis of four crop water stress indices to ambient environmental conditions and stomatal conductance. Sci. Hortic. 2019, 259, 108825. [Google Scholar] [CrossRef]
- Idso, S.B.; Jackson, R.D.; Pinter, P.J., Jr.; Reginato, R.J.; Hatfield, J.L. Normalizing the stress-degree-day parameter for environmental variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Testi, L.; Goldhamer, D.A.; Iniesta, F.; Salinas, M. Crop water stress index is a sensitive water stress indicator in pistachio trees. Irrig. Sci. 2008, 26, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Idso, S.B. Non-water-stressed baselines: A key to measuring and interpreting plant water stress. Agric. Meteorol. 1982, 27, 59–70. [Google Scholar] [CrossRef]
- Maes, W.H.; Steppe, K. Estimating evapotranspiration and drought stress with ground-based thermal remote sensing in agriculture: A review. J. Exp. Bot. 2012, 63, 4671–4712. [Google Scholar] [CrossRef] [Green Version]
- Poirier-Pocovi, M.; Volder, A.; Bailey, B. Modeling of reference temperatures for calculating crop water stress indices from infrared thermography. Agric. Water Manag. 2020, 233, 106070. [Google Scholar] [CrossRef]
- Apolo-Apolo, O.E.; Martínez-Guanter, J.; Pérez-Ruiz, M.; Egea, G. Design and assessment of new artificial reference surfaces for real time monitoring of crop water stress index in maize. Agric. Water Manag. 2020, 240, 106304. [Google Scholar] [CrossRef]
- Jones, H.G.; Stoll, M.; Santos, T.; De Sousa, C.; Chaves, M.M.; Grant, O.M. Use of infrared thermography for monitoring stomatal closure in the field: Application to grapevine. J. Exp. Bot. 2002, 53, 2249–2260. [Google Scholar] [CrossRef]
- Xu, J.; Lv, Y.; Liu, X.; Dalson, T.; Yang, S.; Wu, J. Diagnosing Crop Water Stress of Rice using Infra-red Thermal Imager under Water Deficit Condition. Int. J. Agric. Biol. 2016, 18, 565–572. [Google Scholar] [CrossRef]
- Luan, Y.; Xu, J.; Lv, Y.; Liu, X.; Wang, H.; Liu, S. Improving the performance in crop water deficit diagnosis with canopy temperature spatial distribution information measured by thermal imaging. Agric. Water Manag. 2020, 246, 106699. [Google Scholar] [CrossRef]
- Ingram, K.T.; Real, J.G.; Maguling, M.A.; Obien, M.A.; Loresto, G.C. Comparison of selection indices to screen lowland rice for drought resistance. Euphytica 1990, 48, 253–260. [Google Scholar] [CrossRef]
- Fitriyah, A.; Fatikhunnada, A.; Okura, F.; Nugroho, B.D.A.; Kato, T. Analysis of the Drought Mitigated Mechanism in Terraced Paddy Fields Using CWSI and TVDI Indices and Hydrological Monitoring. Sustainability 2019, 11, 6897. [Google Scholar] [CrossRef] [Green Version]
- Prior, A.; Apolo-Apolo, O.E.; Castro-Valdecantos, P.; Pérez-Ruiz, M.; Egea, G. Long-Term Assessment of Reference Baselines for the Determination of the Crop Water Stress Index in Maize under Mediterranean Conditions. Water 2021, 13, 3119. [Google Scholar] [CrossRef]
- Bellvert, J.; Marsal, J.; Girona, J.; Zarco-Tejada, P.J. Seasonal evolution of crop water stress index in grapevine varieties determined with high-resolution remote sensing thermal imagery. Irrig. Sci. 2014, 33, 81–93. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Rubio, A.E.; Viñuela, I.; Hernández, A.; Gutiérrez-Gordillo, S.; Rodríguez-Pleguezuelo, C.R.; Durán-Zuazo, V.H. Thermal imaging at plant level to assess the crop-water status in almond trees (cv. Guara) under deficit irrigation strategies. Agric. Water Manag. 2018, 208, 176–186. [Google Scholar] [CrossRef]
- Abidin, M.S.B.Z.; Shibusawa, S.; Ohaba, M.; Li, Q.; Bin Khalid, M. Capillary flow responses in a soil–plant system for modified subsurface precision irrigation. Precis. Agric. 2013, 15, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Aschonitis, V.; Diamantopoulou, M.; Papamichail, D. Modeling plant density and ponding water effects flooded rice ET and crop coefficients: Critical discussion about the concepts used in current methods. Theor. Appl. Climatol. 2017, 132, 1165–1186. [Google Scholar] [CrossRef]
- Leinonen, I.; Jones, H.G. Combining thermal and visible imagery for estimating canopy temperature and identifying plant stress. J. Exp. Bot. 2004, 55, 1423–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, H.G.; Sirault, X.R.R. Scaling of Thermal Images at Different Spatial Resolution: The Mixed Pixel Problem. Agronomy 2014, 4, 380–396. [Google Scholar] [CrossRef] [Green Version]
- Fukuoka, M.; Yoshimoto, M.; Hasegawa, T. Varietal Range in Transpiration Conductance of Flowering Rice Panicle and Its Impact on Panicle Temperature. Plant Prod. Sci. 2012, 15, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Murray, F.W. On the Computation of Saturation Vapor Pressure. J. Appl. Meteorol. 1967, 6, 203–204. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Hernández, A.; Padilla-Díaz, C.M.; Diaz-Espejo, A.; Fernández, J.E. Assessing plant water status in a hedgerow olive orchard from thermography at plant level. Agric. Water Manag. 2017, 188, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Parent, B.; Suard, B.; Serraj, R.; Tardieu, F. Rice leaf growth and water potential are resilient to evaporative demand and soil water deficit once the effects of root system are neutralized. Plant Cell Environ. 2010, 33, 1256–1267. [Google Scholar] [CrossRef]
- Irmak, S.; Haman, D.Z.; Bastug, R. Determination of Crop Water Stress Index for Irrigation Timing and Yield Estimation of Corn. Agron. J. 2000, 92, 1221–1227. [Google Scholar] [CrossRef]
- Gutiérrez-Gordillo, S.; García-Tejero, I.F.; Zuazo, V.H.D.; Escalera, A.G.; Gil, F.F.; Amores-Agüera, J.J.; Rodríguez, B.C.; Hernández-Santana, V. Assessing the Water-Stress Baselines by Thermal Imaging for Irrigation Management in Almond Plantations under Water Scarcity Conditions. Water 2020, 12, 1298. [Google Scholar] [CrossRef]
- King, B.; Tarkalson, D.; Sharma, V.; Bjorneberg, D. Thermal Crop Water Stress Index Base Line Temperatures for Sugarbeet in Arid Western U.S. Agric. Water Manag. 2020, 243, 106459. [Google Scholar] [CrossRef]
- Agam, N.; Cohen, Y.; Jimenez-Berni, J.A.; Alchanatis, V.; Kool, D.; Dag, A.; Yermiyahu, U.; Ben-Gal, A. An insight to the performance of crop water stress index for olive trees. Agric. Water Manag. 2013, 118, 79–86. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (Tc −Ta)WSB (°C) a | |||||
|---|---|---|---|---|---|
| Genotype/Water Regime | Condition | CBL | GBL | MBL b | TBL |
| IRAT109 | |||||
| Flooded | 8.08 ± 0.42 | 6.13 ± 0.31 | n/a | n/a | |
| Aerobic | Phytotron | 8.08 ± 0.42 | 6.13 ± 0.31 | n/a | n/a |
| Takanari | |||||
| Flooded | 8.08 ± 0.42 | 10.21 ± 0.56 | n/a | n/a | |
| Aerobic | 8.08 ± 0.42 | 10.21 ± 0.56 | n/a | n/a | |
| IRAT109 | |||||
| Flooded | 10.52 ± 0.82 | 10.54 ± 0.81 | 13.08 ± 1.03 | 11.45 ± 1.69 | |
| Aerobic | Field environment | 10.52 ± 0.82 | 10.54 ± 0.81 | 13.08 ± 1.03 | 11.45 ± 1.69 |
| Takanari | |||||
| Flooded | 10.53 ± 0.83 | 10.46 ± 0.84 | 10.39 ± 0.77 | 9.91 ± 1.68 | |
| Aerobic | 10.52 ± 0.82 | 10.45 ± 0.83 | 10.38 ± 0.76 | 9.90 ± 1.67 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godson-Amamoo, S.; Kato, T.; Katsura, K. Empirical Setting of the Water Stressed Baseline Increases the Uncertainty of the Crop Water Stress Index in a Humid Temperate Climate in Different Water Regimes. Water 2022, 14, 1833. https://doi.org/10.3390/w14121833
Godson-Amamoo S, Kato T, Katsura K. Empirical Setting of the Water Stressed Baseline Increases the Uncertainty of the Crop Water Stress Index in a Humid Temperate Climate in Different Water Regimes. Water. 2022; 14(12):1833. https://doi.org/10.3390/w14121833
Chicago/Turabian StyleGodson-Amamoo, Samuel, Tasuku Kato, and Keisuke Katsura. 2022. "Empirical Setting of the Water Stressed Baseline Increases the Uncertainty of the Crop Water Stress Index in a Humid Temperate Climate in Different Water Regimes" Water 14, no. 12: 1833. https://doi.org/10.3390/w14121833