
Chronic Toxic Effects of Waterborne Mercury on Silver Carp (Hypophthalmichthys molitrix) Larvae


1
Engineering Research Center of Ecology and Agricultural Use of Wetland, Ministry of Education, College of Agriculture, Yangtze University, Jingzhou 434025, China
2
Marine College, Shandong University, Weihai 264209, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Water 2022, 14(11), 1774; https://doi.org/10.3390/w14111774
Submission received: 1 April 2022
/
Revised: 10 May 2022
/
Accepted: 27 May 2022
/
Published: 31 May 2022
(This article belongs to the Special Issue Effects of Emerging Pollutants on Ecological Health in Aquatic Environment)
Abstract
:Mercury (Hg) is a kind of heavy metal pollutant widely existing in the aquatic environment, and it is also recognized to have a highly toxic effect on fish. In this study, silver carp (Hypophthalmichthys molitrix) larvae were exposed to 0 (control), 1, 5, and 10 μg/L Hg2+ for 2 weeks. Antioxidant ability, neurotoxicity, and thyroid hormones (THs) content were evaluated. In comparison with the control, the superoxide dismutase (SOD) activity and the glutathione (GSH) activity were lower in silver carp exposed to 10 μg/L Hg2+. The lowest catalase (CAT) activity was found in the 10 μg/L Hg2+, while malondialdehyde (MDA) content was not significantly different among all groups. Compared with the control, monoamine oxidase (MAO) activity and nitric oxide (NO) content were significantly higher in the 10 μg/L Hg2+, while acetylcholinesterase (AChE) activity significantly decreased. Compared with the control, triiodothyronine (T3) content was significantly higher in the 1 μg/L Hg2+ and significantly lower in the 10 μg/L Hg2+; the 1 μg/L and 5 μg/L Hg2+ groups had significantly higher thyroxine (T4) content than the other groups. In the 1 μg/L Hg2+, the integrated biomarker response (IBR) index value was the highest. In summary, exposure to Hg could decrease the antioxidant ability, cause changes in neurotoxic parameters, and induce disorders of the thyroid hormone system in silver carp larvae. The results of this study may contribute to the understanding of the adverse effects of chronic mercury poisoning on fish.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Mercury (Hg) is a heavy metal contaminant widely present in aquatic ecosystems, exhibiting teratogenicity, neurological, nephrogenic, and immunotoxicity [1,2,3,4]. The impact of various heavy metal pollutants, including Hg, on aquatic environments, has become an increasingly serious problem due to increased human activities [5,6]. Hg has been found in various aquatic environments around the world. For example, the concentration of total Hg reached 16.8 ± 8.4 ng/L in Ya-Er Lake, Xinjiang Province, China [7]. Total Hg was found at 1.8 ± 1.00 ng/L at the bottom of Minamata Bay, Japan [8]. Hg is usually present in the aquatic environment mainly in three forms: elemental mercury (Hg0), inorganic mercury (IHg), and organic mercury (MeHg), with IHg being the most abundant form in freshwater environments and easily converted to methylmercury through continuous biogeochemical cycling [9]. Methylmercury is more likely to accumulate in sediment-feeding aquatic animals and persists in fish tissue via biomagnification in the food chain [10]. Trace amounts of Hg in aquatic environments accumulate in animals with high nutrient levels through the food chain. It endangers aquatic organisms as well as human health and safety [11]. Due to its high toxicity, it is capable of many adverse effects on reproduction and nervous systems [12]. At present, it has become one of the hot spots of ecotoxicological research.
Previous studies on the toxic effects of Hg in different aquatic animals have been extensively reported [13]. For example, Hg exposure leads to oxidative stress, and genes encoding antioxidant enzymes were found to be significantly upregulated in the spleen, head, and kidney of Pelteobagrus fulvidraco [14]. Hg exposure increases the accumulation of Hg in the liver of Danio rerio and damages the ultrastructure of the liver [15]. Hg exposure causes neurotoxicity in the marine medaka Oryzias melastigma through the induction of oxidative stress and the disruption of cellular metabolism [16]. Hg could cause damage to the immune system of fish, including cell necrosis or necrotizing apoptosis and changes in immune gene expression profiles in marine gilthead sea bream and European sea bass [17,18]. Exposure to Hg at even low concentrations could lead to physiological, biochemical, genetic, metabolic, and morphological abnormalities in fish [19,20]. Silver carp (Hypophthalmichthys molitrix) is one of the four major Chinese carp species, as well as one of the most essential freshwater commercial fish, with a global production of 4.79 million tons in 2018 [21,22]. Previous studies on silver carp have focused on growth and development [23], genetic reproduction [24], and ecological regulation [25,26]. However, relatively few studies on the toxicity of heavy metal Hg to silver carp have been reported, especially for nerve damage and related biomarkers such as thyroid hormones (THs).
Therefore, this study aimed to investigate the effect of chronic waterborne Hg exposure on antioxidant ability, neurotoxicity, and THs content in silver carp larvae. Furthermore, this study also provides a comprehensive overview of the physiological stress of chronic waterborne Hg on silver carp larvae via using the integrated biomarker response (IBR) index. This study may increase our understanding of the effects of Hg toxicity on silver carp larvae.
2. Materials and Methods
2.1. Experimental Animals and Test Chemicals
Silver carp larvae were purchased from the Yangtze River Fisheries Research Institute (Jingzhou, China). Silver carp were acclimated in the 120 L plastic box for 7 days, and the experimental water was tap water with full aeration for more than 24 h. Fish were fed hatched brine shrimp (Artemia nauplii) twice a day. The experimental conditions were a water temperature of 24.0 ± 1.0 °C, photoperiod 12L:12D, and dissolved oxygen >7.0 mg/L.
Mercuric chloride (HgCl2, purity > 99%) was purchased from Sigma-Aldrich (St. Louis, MO, USA). As a stock solution of 1 g/L HgCl2, deionized water was prepared. Then, the solution was stored in a brown bottle at room temperature, protected from light. Finally, it was diluted to the mass concentration required for the experimental design.
2.2. Experimental Design and Sample Collection
After acclimation, healthy silver carp larvae (average initial weight 0.027 ± 0.004 g; average body length 1.25 ± 0.06 cm) were selected and randomly assigned to 12 tanks (40 × 23 × 25 cm). In the experiment, silver carp larvae were set up in four groups, three parallel in each group. Each tank contained 20 L tap water and 60 silver carp larvae. The selection of test concentrations of Hg2+ was based on data from our previous acute toxicology experiments (not yet published). Four concentrations of Hg2+ were set at 0, 1, 5, and 10 μg/L in the experiment. All solutions samples were analyzed and the measured concentrations were within 20% of the nominal concentration following OECD guidelines. During the two-week experimental period, half of the water in the tank was replaced with tap water aerated for more than 24 h every day. The temperature was kept at 24.0 ± 1.0 °C, dissolved oxygen >7.0 mg/L, and photoperiod 12L:12D throughout the experimental period. All procedures and animal handling comply with the guidelines approved by the Chinese Society of Laboratory Animal Science. The research has been approved by the Animal Ethics Committee of Yangtze University.
At the end of the exposure experiment, the mucus on the surface of the fish was washed with sterile distilled water. After absorbing the water on the surface of the fish with absorbent tubes, the whole fish were collected in 2 mL Eppendorf tubes and transferred to the −80 °C refrigerator for the next analysis. Because the silver carp larvae were too small, the whole fish was used for biochemical analysis.
2.3. Biochemical Analysis
The 10% concentration of homogenate was prepared by three whole fish per tank with precooled sterile 0.9% saline solution at a ratio of 10% as m (tissue, g): v (saline solution, ml). The homogenate was centrifuged for 10 min at 4 °C in the centrifuge (SIGMA, Germany). The supernatant was collected and assayed for relevant biochemical parameters. The superoxide dismutase (SOD), catalase (CAT), glutathione (GSH), malondialdehyde (MDA) contents, monoamine oxidase (MAO), nitric oxide (NO), and acetylcholinesterase (AChE) were determined with commercial kits by following the manufacturer’s instructions (Jiancheng Bioengineering Research Institute Co., Ltd., Nanjing, China). Enzyme-linked immunosorbent assay (ELISA) kits were used to determine the content of triiodothyronine (T3) and thyroxine (T4) following the manufacturer’s instructions (Uscnlife, Ltd., Wuhan, China). The details of the method were as described in the previous study [27].
2.4. Integrated Biomarker Response
For several biomarkers (antioxidant enzymes, neurotoxicity enzymes, and THs content), IBR index values were calculated following our previous description [28].
2.5. Statistical Analysis
The statistical significance of data was analyzed using one-way analysis of variance (ANOVA) and Duncan’s multiple range test. Data were presented as the mean ± standard error (SE). A probability score of p < 0.05 had been used to determine significance. All statistical analysis was performed with SPSS (version 25). Besides, star plots were drawn using the Origin software (version 9.8).
3. Results
3.1. Antioxidant Ability
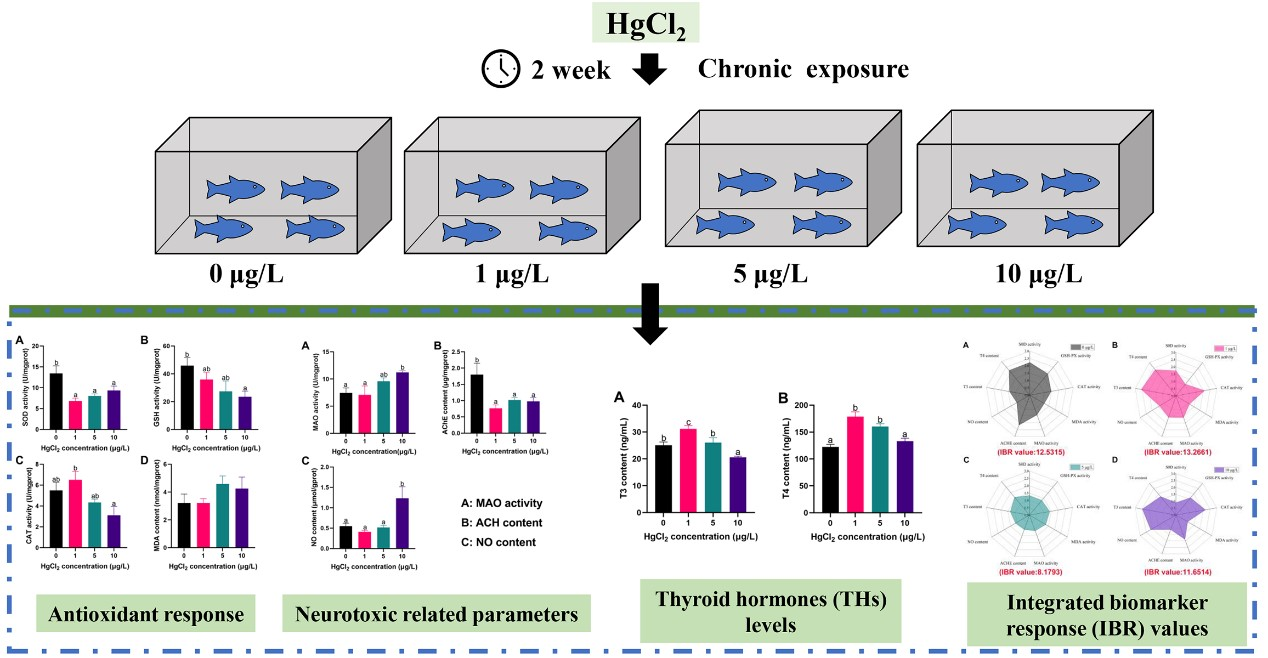
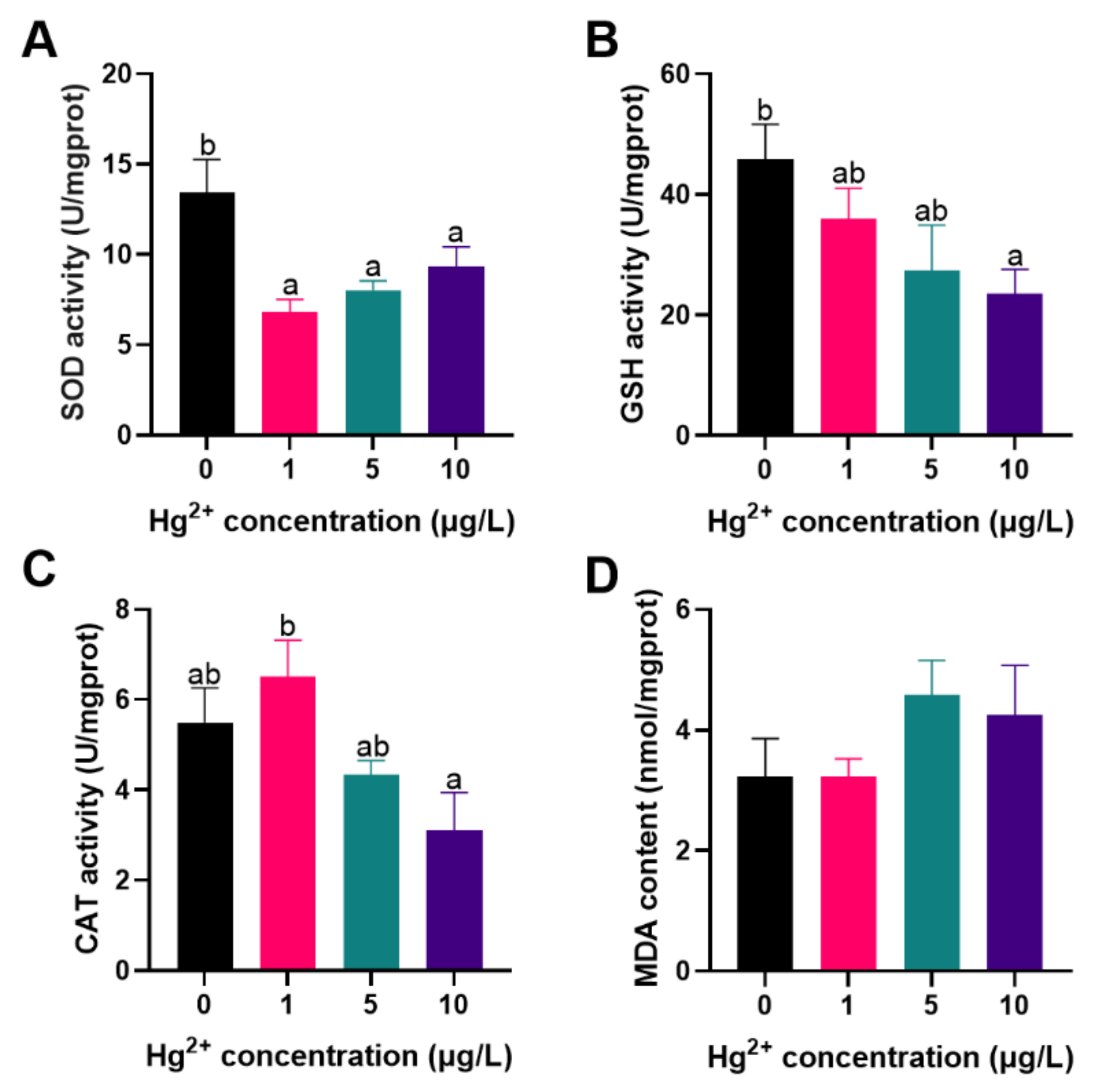
Antioxidant enzyme activities of fish exposed to different concentrations of Hg2+ are shown in Figure 1. The results showed that the SOD activity of fish was significantly lower than the control after exposure to Hg2+ (p < 0.05; Figure 1A); Significantly lower GSH activity was found in fish exposed to 10 μg/L Hg2+, relative to the control (p > 0.05; Figure 1B). The activity of CAT of fish exposed to 10 μg/L Hg2+ was significantly lower than that exposed to 1 μg/L Hg2+ (p < 0.05; Figure 1C). There was no significant change in MDA content among all groups (p > 0.05; Figure 1D). Antioxidant enzyme activities are often used as a biomarker to assess the damage caused by pollutants to aquatic organisms [29]. Under normal conditions, the production of reactive oxygen species (ROS) is in balance with an organism’s antioxidant defense system. When the organism is under external pressure, the organism inevitably produces excessive reactive oxygen species, leading to oxidative stress [30]. Hg exposure induces the production of intracellular ROS, which is a distinctive feature of heavy metal toxicity [31,32].
As the first line of antioxidant defense, SOD acts as a superoxide radical scavenger, converting superoxide anion radicals into O2 and H2O2, while CAT decomposes H2O2 into H2O and O2 to eliminate excess radicals [33,34,35]. GSH is a non-enzymatic antioxidant that directly eliminates reactive oxygen species under oxidative stress, while MDA is used as a lipid peroxidation metabolite in response to the lipid peroxidation status in the organism [36,37]. Several studies have recently reported that Hg toxicity inhibits the activity of antioxidant enzymes in aquatic animals such as Pelteobagrus fulvidraco [14], Pseudosciaena crocea [38], and Apostichopus japonicus [39]. Similarly, the study results suggest that the SOD, GSH, and CAT activities of silver carp larvae exposed to high concentrations of Hg2+ were significantly decreased. Therefore, it is assumed that Hg exposure disrupts the antioxidant defense system of fish, causing oxidative stress. However, more research on the molecular toxicity of Hg exposure for the antioxidant ability of silver carp is required.
3.2. Neurotoxic Related Parameters
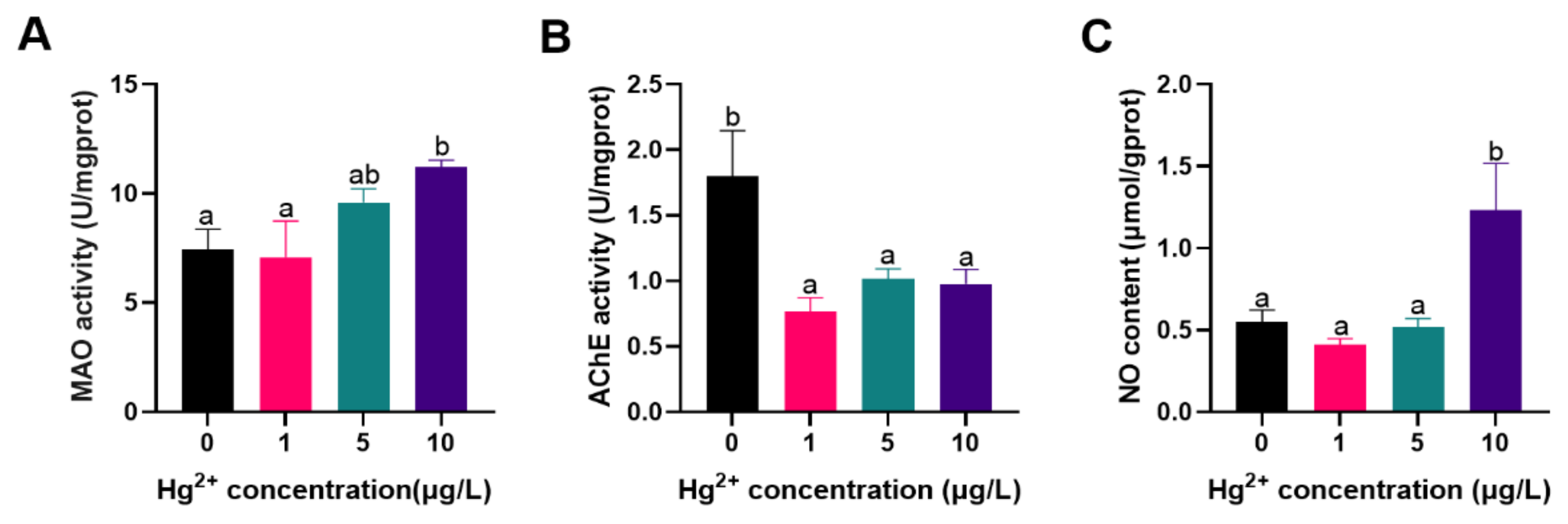
The effects of Hg2+ on the neurotoxic-related enzymes of silver carp larvae are shown in Figure 2. Compared to the control, MAO activity and NO content were significantly increased exposed to 10 μg/L Hg2+ (p < 0.05; Figure 2A,C). In contrast, the Hg2+ exposure groups showed a significant decrease in AChE activity in fish compared to the control (p < 0.05; Figure 2B). Chronic Hg exposure was associated with oxidative damage to the central nervous system in fish. Hg in the aquatic environment causes neurotoxicity in aquatic animals through several mechanisms, including oxidative stress, metabolic abnormalities, and cytoskeleton dysfunction [40,41]. MAO regulates many aspects of the central nervous system, including neurotransmission and neuronal electrical activity [42,43]. Different inhibitory effects on MAO activity after different concentrations of Hg stress have been previously reported [44,45]. In this study, 10 μg/L Hg2+ increased the MAO activity, which contradicted the findings of previous research. This may be partly due to the small concentration of current Hg or the insensitivity of MAO activity induced by Hg-toxicity, which instead produced excitotoxic effects on fish. NO is a neuroinformatic molecule with a wide range of physiological roles [46]. The current results found that 10 μg/L Hg2+ has NO content, which could have toxic effects on fish. AChE is used to degrade acetylcholine, which blocks neurotransmitter excitatory effects on the postsynaptic membrane and maintains normal nerve signal transmission in the organism [47]. The activity of AChE was significantly reduced in the 10 μg/L Hg2+ in this study. The possible reason for this speculation was that decreased AChE activity was associated with neuronal damage. Several recent studies have reported the effect of exposure to other heavy metals contamination on the inhibition of AChE in fish, with results similar to those of the present study [48,49].
3.3. Thyroid Hormones (THs) Content
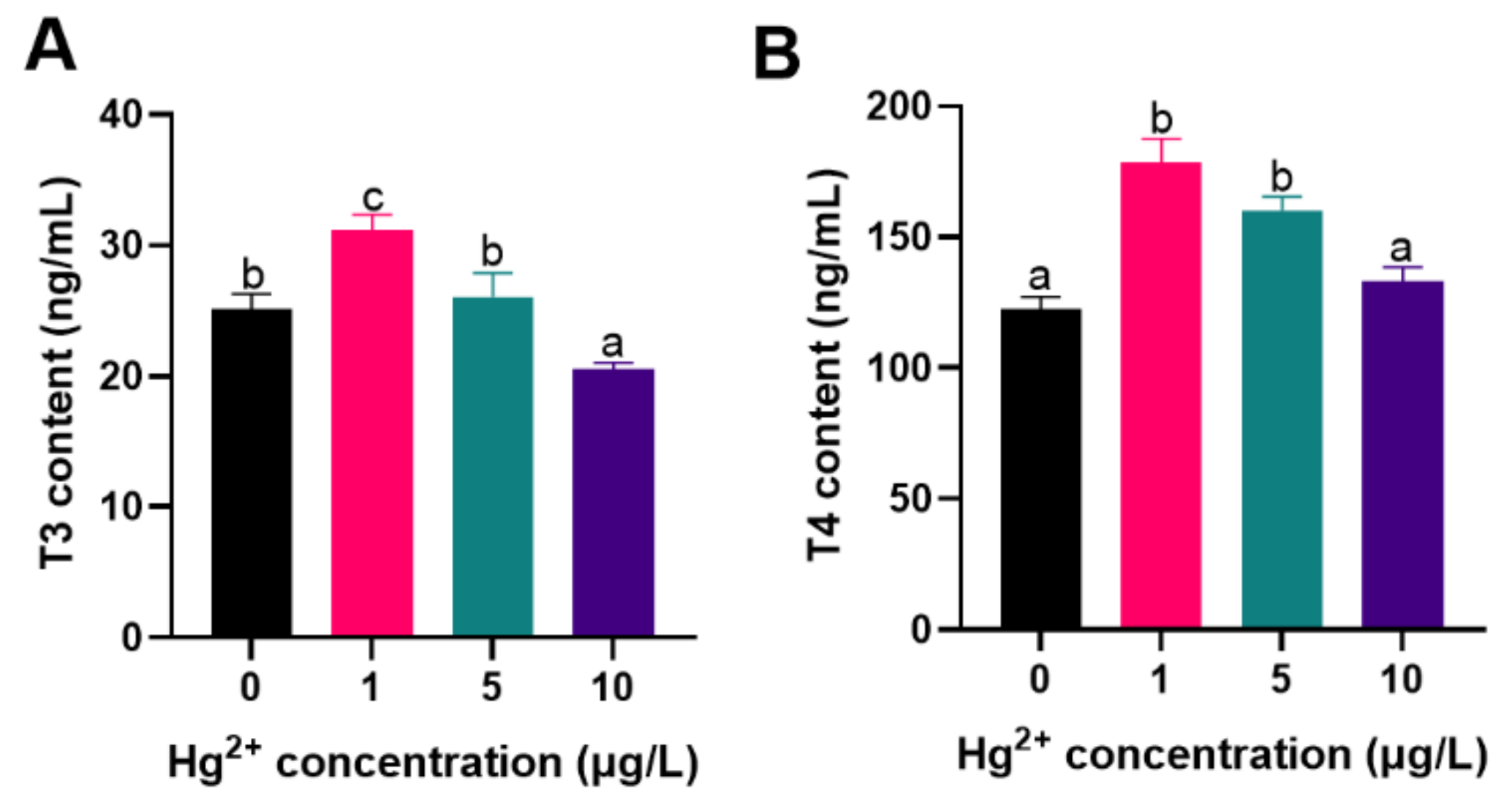
THs content was quantified as shown in Figure 3. Compared to the control, T3 content was significantly increased in the 1 μg/L Hg2+ and significantly decreased in the 10 μg/L Hg2+ (p < 0.05; Figure 3A); T4 content of fish in the 1 μg/L and 5 μg/L Hg2+ groups were significantly higher than that in other groups (p < 0.05; Figure 3B). THs are important regulators in the early growth and development of animals [50]. Environmental contaminants that interfere with THs synthesis have been the subject of interest in aquatic toxicology studies [50,51,52]. T3 and T4 are the main forms of thyroid production and are essential for fish growth and development, osmotic pressure regulation, and nutrient metabolism [53,54].
In this study, T3 content was significantly decreased in fish exposed to high concentrations of Hg2+ and T4 content was significantly increased after exposure to low concentrations of Hg2+, suggesting that the toxicity of Hg induces thyroid dysfunction. In a previous study, low concentrations of Hg exposure had increased the content of whole-body THs in zebrafish larvae [55]. This variation reflects the fact that Hg-induced thyroid dysfunction varies depending on fish species, developmental stage, and treatment dosage.
3.4. Integrated Biomarker Response
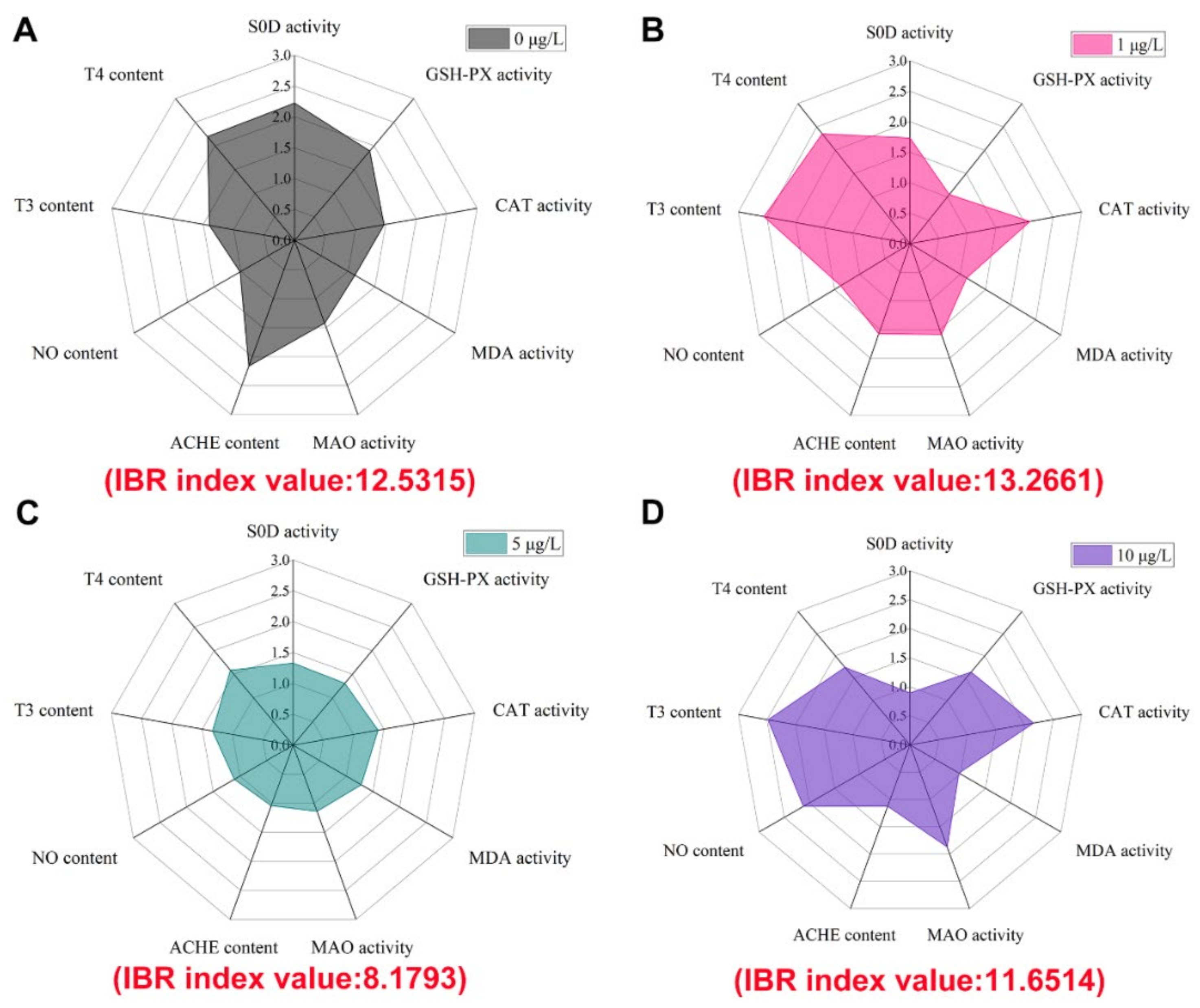
The IBR index values were calculated by integrating the measured nine biomarker responses, and the results for each experimental group were shown in Figure 4. The IBR index values showed that the highest pressure was caused in the 1 μg/L Hg2+ (13.2661) and the lowest pressure was caused in the 5 μg/L Hg2+ (8.1793). The IBR is a novel index that integrates biomarker responses. It is an effective tool for assessing pollutant sensitivity using multiple biomarker responses [56,57]. The IBR index values standardize biomarkers to represent environmental stresses, thus providing a concise and comprehensive assessment of the health of organisms under various stress situations [58]. The IBR index showed the highest value in the 5 μg/L Hg2+, indicating that the 5 μg/L Hg2+ is at the highest stress. In general, high concentrations of Hg2+ stress should have higher overall stress of toxicity. The IBR results of this study did not match the expected results. It is speculated that the reason for this may be the higher overall stress on fish due to excitotoxicity produced by a low concentration of Hg2+.
Although IBR index values can reflect the biotoxicity effects of mercury exposure and contribute to assessing the effects of toxic contaminants on aquatic animals, relatively few reports have used the IBR index to evaluate the toxicity of heavy metals. Furthermore, the type and number of biomarkers selected are also crucial for the calculation of IBR index values [59,60]. Therefore, the IBR index as a quantitative tool to assess the effects of heavy metal toxicity stress on fish still needs more research.
4. Conclusions
Overall, this study indicates that chronic waterborne Hg exposure could decrease the antioxidant capacity of silver carp larvae and cause changes in neurotoxic parameters. At the same time, waterborne Hg exposure could lead to abnormal levels of THs in silver carp larvae and may induce thyroid dysfunction. Furthermore, the IBR index value indicated that a low concentration of Hg2+ (1 μg/L) exposure generated stronger stress than Hg2+ exposure under the highest concentration (10 μg/L). This study provides current biochemical data on the toxicity of Hg, which will be essential for understanding the risk of Hg in aquatic environments.
Author Contributions
Y.-J.W.: Culture and Sampling and Indices Measurement; C.-Z.C.: Writing-Original draft preparation; P.L.: Writing- Reviewing and Editing; L.L.: Data analysis; Y.C.: Conceptualization; Z.-H.L.: Conceptualization and Overall guidance. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Engineering Research Center of Ecology and Agricultural Use of Wetland, Ministry of Education (Yangtze University) (KFT202006); National Key Research and Development Program of China (2018YFD0900902, 2018YFD0900905).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of Yangtze University.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data of this paper are from our experiment and based on the previous published papers cited in our paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mamdouh, A.-Z.; Zahran, E.; Mohamed, F.; Zaki, V. Nannochloropsis oculata feed additive alleviates mercuric chloride-induced toxicity in Nile tilapia (Oreochromis niloticus). Aquat. Toxicol. 2021, 238, 105936. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Chen, L.; Wu, Y.H.; Li, P.; Li, Y.F.; Ni, Z.H. Alteration of thyroid hormone levels and related gene expression in Chinese rare minnow larvae exposed to mercury chloride. Environ. Toxicol. Pharmacol. 2014, 38, 325–331. [Google Scholar] [CrossRef]
- Li, P.; Li, Z.H.; Wu, Y.H. Interactive effects of temperature and mercury exposure on the stress-related responses in the freshwater fish Ctenopharyngodon idella. Aquacult. Res. 2021, 52, 2070–2077. [Google Scholar] [CrossRef]
- Li, Z.-H.; Li, Z.-P.; Tang, X.; Hou, W.-H.; Li, P. Distribution and Risk Assessment of Toxic Pollutants in Surface Water of the Lower Yellow River, China. Water 2021, 13, 1582. [Google Scholar] [CrossRef]
- Islam, M.S.; Ahmed, M.K.; Raknuzzaman, M.; Habibullah-Al-Mamun, M.; Islam, M.K. Heavy metal pollution in surface water and sediment: A preliminary assessment of an urban river in a developing country. Ecol. Indic. 2015, 48, 282–291. [Google Scholar] [CrossRef]
- Li, Z.H.; Li, P.; Dzyuba, B.; Randak, T. Influence of environmental related concentrations of heavy metals on motility parameters and antioxidant responses in sturgeon sperm. Chem.-Biol. Interact. 2010, 188, 473–477. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, X.; Cao, M.; Pan, Y.; Xiao, C.; Wang, P.; Liang, Y.; Liu, G.; Cai, Y. Release of legacy mercury and effect of aquaculture on mercury biogeochemical cycling in highly polluted Ya-Er Lake, China. Chemosphere 2021, 275, 130011. [Google Scholar] [CrossRef]
- Tomiyasu, T.; Matsuyama, A.; Eguchi, T.; Marumoto, K.; Oki, K.; Akagi, H. Speciation of mercury in water at the bottom of Minamata Bay, Japan. Mar. Chem. 2008, 112, 102–106. [Google Scholar] [CrossRef]
- Bravo, A.G.; Cosio, C. Biotic formation of methylmercury: A bio–physico–chemical conundrum. Limnol. Oceanogr. 2020, 65, 1010–1027. [Google Scholar] [CrossRef]
- Donadt, C.; Cooke, C.A.; Graydon, J.A.; Poesch, M.S. Mercury bioaccumulation in stream fish from an agriculturally-dominated watershed. Chemosphere 2021, 262, 128059. [Google Scholar] [CrossRef]
- Da Silva, J.P.; Do S Costa, M.; Campina, F.F.; Bezerra, C.F.; De Freitas, T.S.; Sousa, A.K.; Sobral Souza, C.E.; De Matos, Y.M.; Pereira-Junior, F.N.; Menezes, I.R. Evaluation of chelating and cytoprotective activity of vanillin against the toxic action of mercuric chloride as an alternative for phytoremediation. Environ. Geochem. Health 2021, 43, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- Nowosad, J.; Kucharczyk, D.; Łuczyńska, J. Changes in mercury concentration in muscles, ovaries and eggs of European eel during maturation under controlled conditions. Ecotoxicol. Environ. Saf. 2018, 148, 857–861. [Google Scholar] [CrossRef]
- Li, Z.H.; Li, P.; Wu, Y. Regulation of glutathione-dependent antioxidant defense system of grass carp Ctenopharyngodon idella under the combined stress of mercury and temperature. Env. Sci Pollut Res Int 2021, 28, 1689–1696. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, Y.; Rao, J.; Liu, Z.; Chen, Q. Effects of inorganic mercury exposure on histological structure, antioxidant status and immune response of immune organs in yellow catfish (Pelteobagrus fulvidraco). J. Appl. Toxicol. 2018, 38, 843–854. [Google Scholar] [CrossRef]
- Chen, Q.-L.; Sun, Y.-L.; Liu, Z.-H.; Li, Y.-W. Sex-dependent effects of subacute mercuric chloride exposure on histology, antioxidant status and immune-related gene expression in the liver of adult zebrafish (Danio rerio). Chemosphere 2017, 188, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, D.; Lin, L.; Wang, M. Quantitative proteomic analysis reveals proteins involved in the neurotoxicity of marine medaka Oryzias melastigma chronically exposed to inorganic mercury. Chemosphere 2015, 119, 1126–1133. [Google Scholar] [CrossRef]
- Morcillo, P.; Esteban, M.Á.; Cuesta, A. Heavy metals produce toxicity, oxidative stress and apoptosis in the marine teleost fish SAF-1 cell line. Chemosphere 2016, 144, 225–233. [Google Scholar] [CrossRef]
- Morcillo, P.; Romero, D.; Meseguer, J.; Esteban, M.; Cuesta, A. Cytotoxicity and alterations at transcriptional level caused by metals on fish erythrocytes in vitro. Environ. Sci. Pollut. Res. 2016, 23, 12312–12322. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Wang, S.; Dong, W.; Hua, X.; Li, Y.; Song, X.; Chu, Q.; Hou, S.; Li, Y. The toxicological effects of mercury exposure in marine fish. Bull. Environ. Contam. Toxicol. 2019, 102, 714–720. [Google Scholar] [CrossRef]
- Huang, W.; Cao, L.; Shan, X.; Lin, L.; Dou, S. Toxicity testing of waterborne mercury with red sea bream (Pagrus major) embryos and larvae. Bull. Environ. Contam. Toxicol. 2011, 86, 398–405. [Google Scholar] [CrossRef]
- Sha, H.; Luo, X.-Z.; Wang, D.; Li, X.-h.; Zou, G.-W.; Liang, H.-W. New insights to protection and utilization of silver carp (Hypophthalmichthys molitrix) in Yangtze River based on microsatellite analysis. Fish. Res. 2021, 241, 105997. [Google Scholar] [CrossRef]
- FAO. FAO Yearbook: The State of World Fisheries and Aquaculture 2020: Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- George, A.E.; Chapman, D.C. Aspects of embryonic and larval development in bighead carp Hypophthalmichthys nobilis and silver carp Hypophthalmichthys molitrix. PLoS ONE 2013, 8, e73829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabó, T.; Urbányi, B.; Müller, T.; Szabó, R.; Horváth, L. Assessment of induced breeding of major Chinese carps at a large-scale hatchery in Hungary. Aquac. Rep. 2019, 14, 100193. [Google Scholar] [CrossRef]
- Houshmand, P.; Hedayati, A. Dietary Effect of Probiotic and Prebiotic on Some Mucus Indices of Silver Carp (Hypophthalmichthys molitrix) Exposed to Silver Nanoparticles. Biol. Trace Elem. Res. 2022, 1–9. [Google Scholar] [CrossRef]
- Meng, L.-J.; Zhang, Y.; Li, X.-X.; Liu, J.-H.; Wen, B.; Gao, J.-Z.; Chen, Z.-Z. Comparative analysis of bacterial communities of water and intestines of silver carp (Hypophthalmichthys molitrix) and bighead carp (H. nobilis) reared in aquaculture pond systems. Aquaculture 2021, 534, 736334. [Google Scholar] [CrossRef]
- Li, Z.-H.; Chen, L.; Wu, Y.-H.; Li, P.; Li, Y.-F.; Ni, Z.-H. Effects of waterborne cadmium on thyroid hormone levels and related gene expression in Chinese rare minnow larvae. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 161, 53–57. [Google Scholar] [CrossRef]
- Chen, C.-Z.; Li, P.; Wang, W.-B.; Li, Z.-H. Response of growth performance, serum biochemical parameters, antioxidant capacity, and digestive enzyme activity to different feeding strategies in common carp (Cyprinus carpio) under high-temperature stress. Aquaculture 2022, 548, 737636. [Google Scholar] [CrossRef]
- Gu, P.; Li, Q.; Zhang, W.; Gao, Y.; Sun, K.; Zhou, L.; Zheng, Z. Biological toxicity of fresh and rotten algae on freshwater fish: LC50, organ damage and antioxidant response. J. Hazard. Mater. 2021, 407, 124620. [Google Scholar] [CrossRef]
- Kong, Y.; Li, M.; Shan, X.; Wang, G.; Han, G. Effects of deltamethrin subacute exposure in snakehead fish, Channa argus: Biochemicals, antioxidants and immune responses. Ecotoxicol. Environ. Saf. 2021, 209, 111821. [Google Scholar] [CrossRef]
- Mieiro, C.; Pereira, M.; Duarte, A.; Pacheco, M. Brain as a critical target of mercury in environmentally exposed fish (Dicentrarchus labrax)—bioaccumulation and oxidative stress profiles. Aquat. Toxicol. 2011, 103, 233–240. [Google Scholar] [CrossRef]
- Roos, D.H.; Puntel, R.L.; Santos, M.M.; Souza, D.O.; Farina, M.; Nogueira, C.W.; Aschner, M.; Burger, M.E.; Barbosa, N.B.; Rocha, J.B. Guanosine and synthetic organoselenium compounds modulate methylmercury-induced oxidative stress in rat brain cortical slices: Involvement of oxidative stress and glutamatergic system. Toxicol. Vitr. 2009, 23, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Romeo, M.; Bennani, N.; Gnassia-Barelli, M.; Lafaurie, M.; Girard, J. Cadmium and copper display different responses towards oxidative stress in the kidney of the sea bass Dicentrarchus labrax. Aquat. Toxicol. 2000, 48, 185–194. [Google Scholar] [CrossRef]
- Li, Z.H.; Li, P.; Randak, T. Ecotoxocological effects of short-term exposure to a human pharmaceutical Verapamil in juvenile rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. C 2010, 152, 385–391. [Google Scholar] [CrossRef]
- Li, Z.H.; Chen, L.; Wu, Y.H.; Li, P.; Li, Y.F.; Ni, Z.H. Effects of mercury on oxidative stress and gene expression of potential biomarkers in larvae of the Chinese rare minnow Gobiocypris rarus. Arch. Environ. Contam. Toxicol. 2014, 67, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Gopi, N.; Vijayakumar, S.; Thaya, R.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Al-Anbr, M.N.; Vaseeharan, B. Chronic exposure of Oreochromis niloticus to sub-lethal copper concentrations: Effects on growth, antioxidant, non-enzymatic antioxidant, oxidative stress and non-specific immune responses. J. Trace Elem. Med. Biol. 2019, 55, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Zlabek, V.; Velisek, J.; Grabic, R.; Machova, J.; Kolarova, J.; Li, P.; Randak, T. Antioxidant responses and plasma biochemical characteristics in the freshwater rainbow trout, Oncorhynchus mykiss, after acute exposure to the fungicide propiconazole. Czech J. Anim. Sci. 2011, 56, 61–69. [Google Scholar] [CrossRef]
- Wu, F.; Huang, W.; Liu, Q.; Xu, X.; Zeng, J.; Cao, L.; Hu, J.; Xu, X.; Gao, Y.; Jia, S. Responses of antioxidant defense and immune gene expression in early life stages of large yellow croaker (Pseudosciaena crocea) under methyl mercury exposure. Front. Physiol. 2018, 9, 1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Ren, T.; Han, Y.; Jiang, Z.; Hu, Y.; Bai, Z.; Wang, L.; Ding, J. The Effects of Sub-lethal Dietary Mercury on Growth Performance, Bioaccumulation, and Activities of Antioxidant Enzymes in Sea Cucumber, Apostichopus japonicus. Bull. Environ. Contam. Toxicol. 2018, 100, 683–689. [Google Scholar] [CrossRef]
- Barboza, L.G.A.; Vieira, L.R.; Branco, V.; Figueiredo, N.; Carvalho, F.; Carvalho, C.; Guilhermino, L. Microplastics cause neurotoxicity, oxidative damage and energy-related changes and interact with the bioaccumulation of mercury in the European seabass, Dicentrarchus labrax (Linnaeus, 1758). Aquat. Toxicol. 2018, 195, 49–57. [Google Scholar] [CrossRef]
- Pereira, P.; Puga, S.; Cardoso, V.; Pinto-Ribeiro, F.; Raimundo, J.; Barata, M.; Pousão-Ferreira, P.; Pacheco, M.; Almeida, A. Inorganic mercury accumulation in brain following waterborne exposure elicits a deficit on the number of brain cells and impairs swimming behavior in fish (white seabream—Diplodus sargus). Aquat. Toxicol. 2016, 170, 400–412. [Google Scholar] [CrossRef]
- Mukherjee, A.; Bhowmick, A.R.; Mukherjee, J.; Moniruzzaman, M. Physiological response of fish under variable acidic conditions: A molecular approach through the assessment of an eco-physiological marker in the brain. Environ. Sci. Pollut. Res. 2019, 26, 23442–23452. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Li, P.; Randak, T. Effect of a human pharmaceutical carbamazepine on antioxidant responses in brain of a model teleost in vitro: An efficient approach to biomonitoring. J. Appl. Toxicol. 2010, 30, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Crump, K.L.; Trudeau, V.L. Mercury-induced reproductive impairment in fish. Environ. Toxicol. Chem. Int. J. 2009, 28, 895–907. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, Z.H.; Zhong, L.Q. Effects of low concentrations of triphenyltin on neurobehavior and the thyroid endocrine system in zebrafish. Ecotoxicol. Environ. Saf. 2019, 186, 109776. [Google Scholar] [CrossRef]
- Li, Z.-H.; Li, P.; Shi, Z.-C. Chronic exposure to tributyltin induces brain functional damage in juvenile common carp (Cyprinus carpio). PLoS ONE 2015, 10, e0123091. [Google Scholar] [CrossRef] [Green Version]
- Basha, D.C.; Rani, M.U.; Devi, C.B.; Kumar, M.R.; Reddy, G.R. Perinatal lead exposure alters postnatal cholinergic and aminergic system in rat brain: Reversal effect of calcium co-administration. Int. J. Dev. Neurosci. 2012, 30, 343–350. [Google Scholar] [CrossRef]
- Lee, J.-W.; Choi, H.; Hwang, U.-K.; Kang, J.-C.; Kang, Y.J.; Kim, K.I.; Kim, J.-H. Toxic effects of lead exposure on bioaccumulation, oxidative stress, neurotoxicity, and immune responses in fish: A review. Environ. Toxicol. Pharmacol. 2019, 68, 101–108. [Google Scholar] [CrossRef]
- Santos, D.; Luzio, A.; Matos, C.; Bellas, J.; Monteiro, S.M.; Félix, L. Microplastics alone or co-exposed with copper induce neurotoxicity and behavioral alterations on zebrafish larvae after a subchronic exposure. Aquat. Toxicol. 2021, 235, 105814. [Google Scholar] [CrossRef]
- Zhang, S.; Guo, X.; Lu, S.; Sang, N.; Li, G.; Xie, P.; Liu, C.; Zhang, L.; Xing, Y. Exposure to PFDoA causes disruption of the hypothalamus-pituitary-thyroid axis in zebrafish larvae. Environ. Pollut. 2018, 235, 974–982. [Google Scholar] [CrossRef]
- Liu, W.; Yang, J.; Li, J.; Zhang, J.; Zhao, J.; Yu, D.; Xu, Y.; He, X.; Zhang, X. Toxicokinetics and persistent thyroid hormone disrupting effects of chronic developmental exposure to chlorinated polyfluorinated ether sulfonate in Chinese rare minnow. Environ. Pollut. 2020, 263, 114491. [Google Scholar] [CrossRef]
- Li, P.; Li, Z.H. Environmental co-exposure to TBT and Cd caused neurotoxicity and thyroid endocrine disruption in zebrafish, a three-generation study in a simulated environment. Environ. Pollut. 2020, 259, 113868. [Google Scholar] [CrossRef] [PubMed]
- Deal, C.K.; Volkoff, H. The role of the thyroid axis in fish. Front. Endocrinol. 2020, 11, 861. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Li, P.; Wu, Y.H. Effects of temperature fluctuation on endocrine disturbance of grass carp Ctenopharyngodon idella under mercury chloride stress. Chemosphere 2021, 263, 128137. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, Y.; Liu, Z.; Chen, Q. Environmentally relevant concentrations of mercury exposure alter thyroid hormone levels and gene expression in the hypothalamic–pituitary–thyroid axis of zebrafish larvae. Fish Physiol. Biochem. 2018, 44, 1175–1183. [Google Scholar] [CrossRef]
- Kim, W.-K.; Lee, S.-K.; Park, J.-W.; Choi, K.; Cargo, J.; Schlenk, D.; Jung, J. Integration of multi-level biomarker responses to cadmium and benzo [k] fluoranthene in the pale chub (Zacco platypus). Ecotoxicol. Environ. Saf. 2014, 110, 121–128. [Google Scholar] [CrossRef]
- Li, Z.H.; Zlabek, V.; Turek, J.; Velisek, J.; Pulkrabova, J.; Kolarova, J.; Sudova, E.; Berankova, P.; Hradkova, P.; Hajslova, J.; et al. Evaluating environmental impact of STPs situated on streams in the Czech Republic: An integrated approach to biomonitoring the aquatic environment. Water Res. 2011, 45, 1403–1413. [Google Scholar] [CrossRef]
- Baudou, F.G.; Ossana, N.A.; Castañé, P.M.; Mastrángelo, M.M.; Núñez, A.A.G.; Palacio, M.J.; Ferrari, L. Use of integrated biomarker indexes for assessing the impact of receiving waters on a native neotropical teleost fish. Sci. Total Environ. 2019, 650, 1779–1786. [Google Scholar] [CrossRef]
- Biswal, A.; Srivastava, P.P.; Pal, P.; Gupta, S.; Varghese, T.; Jayant, M. A multi-biomarker approach to evaluate the effect of sodium chloride in alleviating the long-term transportation stress of Labeo rohita fingerlings. Aquaculture 2021, 531, 735979. [Google Scholar] [CrossRef]
- Devin, S.; Burgeot, T.; Giambérini, L.; Minguez, L.; Pain-Devin, S. The integrated biomarker response revisited: Optimization to avoid misuse. Environ. Sci. Pollut. Res. 2014, 21, 2448–2454. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Response to the antioxidant ability of silver carp larvae exposed to Hg2+. (A) SOD activity; (B) GSH activity; (C) CAT activity; (D) MDA content. Vertical bars represent the mean ± SE (n = 3). Different letters indicate significant differences between groups (p < 0.05).
Figure 1.
Response to the antioxidant ability of silver carp larvae exposed to Hg2+. (A) SOD activity; (B) GSH activity; (C) CAT activity; (D) MDA content. Vertical bars represent the mean ± SE (n = 3). Different letters indicate significant differences between groups (p < 0.05).

Figure 2.
Response to the neurotoxic related parameters of silver carp larvae exposed to Hg2+ (A) MAO activity; (B) AChE content; (C) NO content. Vertical bars represent the mean ± SE (n = 3). Different letters indicate significant differences between groups (p < 0.05).
Figure 2.
Response to the neurotoxic related parameters of silver carp larvae exposed to Hg2+ (A) MAO activity; (B) AChE content; (C) NO content. Vertical bars represent the mean ± SE (n = 3). Different letters indicate significant differences between groups (p < 0.05).

Figure 3.
Response to the thyroid hormones levels of silver carp larvae exposed to Hg2+. (A) T3 content; (B) T4 content. Vertical bars represent the mean ± SE (n = 3). Different letters indicate significant differences between groups (p < 0.05).
Figure 3.
Response to the thyroid hormones levels of silver carp larvae exposed to Hg2+. (A) T3 content; (B) T4 content. Vertical bars represent the mean ± SE (n = 3). Different letters indicate significant differences between groups (p < 0.05).

Figure 4.
Star plot and IBR index values of normalized biomarker data in different groups. (A) 0 μg/L; (B) 1 μg/L Hg2+; (C) 5 μg/L Hg2+; (D) 10 μg/L Hg2+.
Figure 4.
Star plot and IBR index values of normalized biomarker data in different groups. (A) 0 μg/L; (B) 1 μg/L Hg2+; (C) 5 μg/L Hg2+; (D) 10 μg/L Hg2+.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Y.-J.; Chen, C.-Z.; Li, P.; Liu, L.; Chai, Y.; Li, Z.-H. Chronic Toxic Effects of Waterborne Mercury on Silver Carp (Hypophthalmichthys molitrix) Larvae. Water 2022, 14, 1774. https://doi.org/10.3390/w14111774
AMA Style
Wang Y-J, Chen C-Z, Li P, Liu L, Chai Y, Li Z-H. Chronic Toxic Effects of Waterborne Mercury on Silver Carp (Hypophthalmichthys molitrix) Larvae. Water. 2022; 14(11):1774. https://doi.org/10.3390/w14111774
Chicago/Turabian StyleWang, Ya-Jun, Cheng-Zhuang Chen, Ping Li, Ling Liu, Yi Chai, and Zhi-Hua Li. 2022. "Chronic Toxic Effects of Waterborne Mercury on Silver Carp (Hypophthalmichthys molitrix) Larvae" Water 14, no. 11: 1774. https://doi.org/10.3390/w14111774
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.