
Potential Use of Aquatic Vascular Plants to Control Cyanobacterial Blooms: A Review
by
 , ,
, ,
Inna Nezbrytska
 ,
,
Oleg Usenko
,
Igor Konovets
,
Tetiana Leontieva
,
Igor Abramiuk
,
Mariia Goncharova
and
Olena Bilous
*
Institute of Hydrobiology of NAS of Ukraine, Geroiv Stalingrada 12, 04210 Kyiv, Ukraine
*
Author to whom correspondence should be addressed.
Water 2022, 14(11), 1727; https://doi.org/10.3390/w14111727
Submission received: 24 April 2022
/
Revised: 24 May 2022
/
Accepted: 26 May 2022
/
Published: 27 May 2022
(This article belongs to the Special Issue Wetland Response to Climate Change)
Abstract
:Intense “blooming” of cyanobacteria (blue-green algae) caused by eutrophication and climate change poses a serious threat to freshwater ecosystems and drinking water safety. Preventing the proliferation of cyanobacteria and reducing water nutrient load is a priority for the restoration of eutrophic water bodies. Aquatic plants play an important role in the function and structure of aquatic ecosystems, affecting the physiochemistry of the water and bottom sediments, primary production, and biotic interactions that support a balanced ecosystem. This review examines the inhibitory effect of aquatic vascular plants on harmful blooms of cyanobacteria. Aquatic plants are able to successfully inhibit the growth of cyanobacteria through various mechanisms, including by reducing nutrient and light availability, creating favorable conditions for the development of herbivorous zooplankton, and releasing allelopathic active substances (allelochemicals) with algicidal effect. Allelopathy is species-specific and therefore acts as one of the key mechanisms by which the development of cyanobacterial populations in aquatic ecosystems is regulated. However, allelopathic activity of aquatic vascular plants depends on various factors (species characteristics of aquatic plants, area, and density of overgrowth of water bodies, physiochemical properties of allelopathically active substances, hydrological and hydrochemical regimes, temperature, light intensity, etc.), which may regulate the impact of allelochemicals on algal communities. The paper also discusses some problematic aspects of using fast-growing species of aquatic vascular plants to control cyanobacterial blooms.
1. Introduction
The increase in frequency and duration of blue-green algal (cyanobacterial) blooms poses serious threats to the environment, including degradation of local and global water resources [1,2,3,4,5,6]. This is due to the production of a wide range of toxic secondary metabolites, namely cyanotoxins, by some species of planktonic microalgae (Microcystis aeruginosa (Kützing) Kützing; Dolichospermum flosaquae (Bornet & Flahault) P.Wacklin, L.Hoffmann & Komárek (=Anabaena flosaquae Brébisson ex Bornet and Flauhault); Aphanizomenon flosaquae Ralfs ex Bornet and Flahault; Planktothrix agardhii (Gomont) Anagnostidis and Komárek; Planktothrix rubescens (De Candolle ex Gomont) Anagnostidis and Komárek; Cylindrospermopsis raciborskii (Woloszynska) Seenaya and Sabbaraju (=Raphidiopsis raciborskii (Woloszynska) Aguilera, Berrendero Gómez, Kastovsky, Echenique and Salerno), etc.) [1,7,8,9]. Eutrophication and climate change are the two main factors contributing to the intensification and spreading of harmful blue-green algae blooms [1,2]. Many approaches that involve the use of synthetic chemicals to mitigate cyanobacterial blooms have been proposed [10,11,12]; however, recent studies demonstrate that aquatic vascular plants offer a promising solution through natural biological control of cyanobacterial blooms [5,12,13,14,15,16,17]. Aquatic vascular plants play an important role in the functioning and structure formation of aquatic ecosystems and ensure their sustainability [18,19,20]. They do so by affecting the physical and chemical properties of the water and bottom sediments, carbon fixation and biomass production, nutrient transformations, and biotic interactions that support a balanced ecosystem [21,22].
The purpose of this review is to examine the mechanisms by which aquatic vascular plants can control harmful blooms of blue-green algae (cyanobacteria) with a focus on the main benefits and some problematic aspects of their use.
2. Mechanisms of Aquatic Vascular Plants’ Influence on Cyanobacterial Blooms
Aquatic plants reduce phytoplankton blooms, including cyanobacterial blooms, by releasing allelopathic compounds into the environment, altering physical and chemical parameters of the aquatic environment (e.g., light regime, inorganic nutrient concentrations), or by facilitating the development of herbivorous zooplankton [18,21,23]. The mechanism and degree of inhibition of phytoplankton by aquatic macrophytes (benthic and floating aquatic plants) varies depending on the area that they occupy in a water body (Figure 1).
Aquatic vascular plants inhibit phytoplankton by reducing the solar radiation available for photosynthesis (shading effect) [26,27,28]. This is a particularly common mechanism for floating-leaved plants such as Trapa natans L., Nuphar lutea L. Smith, Pontederia crassipes Mart., Pistia stratiotes L. [23,28,29,30] (Figure 2). Large, dense thickets of Trapa natans may absorb more than 95% of incident sunlight and inhibit the growth of other plants and their associated microscopic flora and fauna to become the dominant species in aquatic environments [28,31,32,33]. A similar pattern of competitive exclusion was described for Pontederia crassipes or common water hyacinth (former name Eichhornia crassipes Mart.), water lettuce (Pistia stratiotes) and yellow waterlily (Nuphar lutea) [23,28,34].
Reductions of cyanobacterial biomass, which is often observed in thickets of aquatic plants, may be associated with competition for nutrients [18,35,36]. The competition for nutrients applies especially to the effects of non-rooted, floating leaf plants, because submerged rooted plants absorb essential nutrients from the bottom sediments, which supports their active growth even under very low dissolved nutrient concentrations in the water column [18,36,37,38]. Essien et al. [22] demonstrated that the distribution of Pistia stratiotes, Lemma sp., Nymphea lotus L. and Pontederia crassipes is associated with large quantities of organic matter in the water. These authors also noted that these plants play a significant role in improving water quality by taking up organic and inorganic substances from the aquatic environment. Such reduction in nutrient concentrations can contribute to decreasing the intensity of cyanobacterial blooms.
Aquatic vascular plants can indirectly affect the amount of nutrients in the water. The presence of submerged macrophytes increases the surface area available for periphyton and, as a consequence, competition for nutrients between phytoplankton and periphyton [18,39]. In addition, under anoxic conditions formed in the areas overgrown with macrophytes, heterotrophic denitrification increases [18,21,40]. This promotion of denitrification leads to a decrease in nitrate concentration and changes the ratio of nitrogen species, which can limit the growth of cyanobacteria [21].
Some authors report that the density of planktonic algae may decrease due to intensified sedimentation. However, this trend is usually observed in larger microalgae species or those lacking flagella or gas vacuoles [41,42]. Many species of planktonic blue-green algae (cyanobacteria), including Aphanizomenon flosaquae, Dolichospermum flosaquae, Microcystis aeruginosa, Planktothrix agardhii and Planktothrix rubescens, have gas vacuoles that ensure their buoyancy [11,43,44,45]. This mechanism of reducing the intensity of blue-green algae development by aquatic vascular plants appears to play a secondary role.
Aquatic vascular plants are known to serve as a refuge for zooplankton [18,46]. Shallow lakes support higher zooplankton densities within or close to the areas overgrown with aquatic vascular plants during the day [47]. Given that most species of zooplankton are herbivorous, increased densities of zooplankton will decrease cyanobacteria abundance. Many authors note that the high species richness and development of high densities of zooplankton in dense beds of submerged aquatic plants [48,49,50,51], and their exometabolites (phenolcarboxylic acids) stimulate Daphnia magna Straus fertility [52]. However, it should be noted that helophytes and floating leaf plants create poorer physicochemical conditions for the development of large populations of aquatic animals compared to submerged plants [48,53]. In addition, cyanotoxins produced by blue-green algae are released in large quantities and can adversely affect zooplankton. Thus, this indirect mechanism does not always play an important role.
In view of the above, allelopathy appears to be the most effective mechanism to control cyanobacterial development in aquatic communities. Allelopathic interactions occur due to the synthesis and release of specific allelochemicals by plants [21,24,54,55,56,57]. Many macrophyte species, as active producers of these substances, inhibit the development of cyanobacteria [21,24].
3. The Main Groups of Allelopathically Active Substances of Aquatic Vascular Plants with Algicidal Action
The allelopathically active compounds (allelochemicals) with algicidal effects can be categorized [58,59] into four main groups based on their biosynthesis pathways: polyphenols, nitrogen-containing compounds, fatty acids, and terpenoids.
Structural formulae of polyphenols include an aromatic ring with one or more hydroxyl groups. Depending on peculiarities of chemical structure, they are divided into simple phenols, phenolcarbonic acids, coumarins, flavonoids, quinones, and tannins [60]. Algicidal activity of exometabolites of aquatic vascular plants is determined by their physical and chemical properties [15,58,61,62]. Polyphenols containing hydroxyl groups in “ortho” and/or “para” position cause a stronger inhibitory effect on blue-green algae compared to polyphenols with a “meta” position of hydroxyl groups [12,63,64]. Polyphenols (including shikimate pathway-producing polyphenols) are one of the main classes of allelochemicals in Myriophyllum spicatum L., Phragmites communis Trin. and Trapa natans [65,66,67,68,69]. The algicidal activity of phenolic compounds of M. spicatum was first described in 1987 by Planas et al. [70]. It was later discovered that species of Myriophyllum produce various phenolic allelochemicals that inhibit the growth of cyanobacteria, especially cyanobacteria that cause harmful algal blooms [5,71]. The total content of phenolic compounds in M. spicatum can reach up to 12% (dry weight), and the content of algaecide telimagrandin II—up to 2% [72]. The influence of phenolcarboxylic acids on epiphytic algae has also been demonstrated [73,74]. In this case, gallic, salicylic, and caffeic acids show the highest allelopathic activity, changing the dominant epiphytic algae when the ecological state of water bodies changes.
Nitrogen-containing compounds include alkaloids, amines, amino acids, and their derivatives. Among these allelopathically active compounds, alkaloids are highly toxic to algae and include a large group of secondary metabolites containing one or more nitrogen atoms, often in the heterocyclic ring, and have alkaline properties [75,76]. Some aquatic vascular plants, such as Nuphar lutea, produce acaloids (resorcin, etc.) and are highly active against not only phytoplankton species but also zooplankton, including crustaceans [57,77].
Fatty acids are carboxylic acids possessing long aliphatic carbon chains (saturated or unsaturated). The shorter the carbon chain and the more unsaturated the bonds in the fatty acid, the higher is the toxic effect on algae [12,61]. Plants such as Potamogeton natans L., Nuphar lutea, Nymphaea alba and Myriophyllum spicatum are considered to be the most active producers of allelochemical fatty acids. The proportion of fatty acids among volatile organic compounds in these plants may exceed 60–70% [5]. Although fatty acids are common compounds in plant metabolism, release of large quantities by vascular plants can inhibit microalgal growth.
Terpenoids is the common name for a class of hydrocarbons and their oxygen-containing derivatives (alcohols, aldehydes, carboxylic acids, and esters, etc.) [59,78]. Only a limited number of substances in this class have algicidal effects compared to polyphenols, alkaloids, and fatty acids [58]. In addition, their ability to inhibit algal growth is considerably weaker compared to other allelopathic compounds. A typical example of a substance belonging to this class with toxic effects on blue-green algae is terpenoid β-ionone [58,79].
The most promising species for biological control of cyanobacteria due to their production of allelopathic compounds are Nuphar lutea, Myriophyllum spicatum, Ceratophyllum demersum L., Phragmites communis and Elodea canadensis Michx., as well as plants of genera Eichhornia, Pistia, Potamogeton, Typha and Trapa [5,16,80,81]. Table 1 represents the main allelochemicals of aquatic vascular plants that possess algicidal properties.
4. Physiological and Biochemical Mechanisms of Allelopathic Effect of Aquatic Vascular Plants on Cyanobacteria
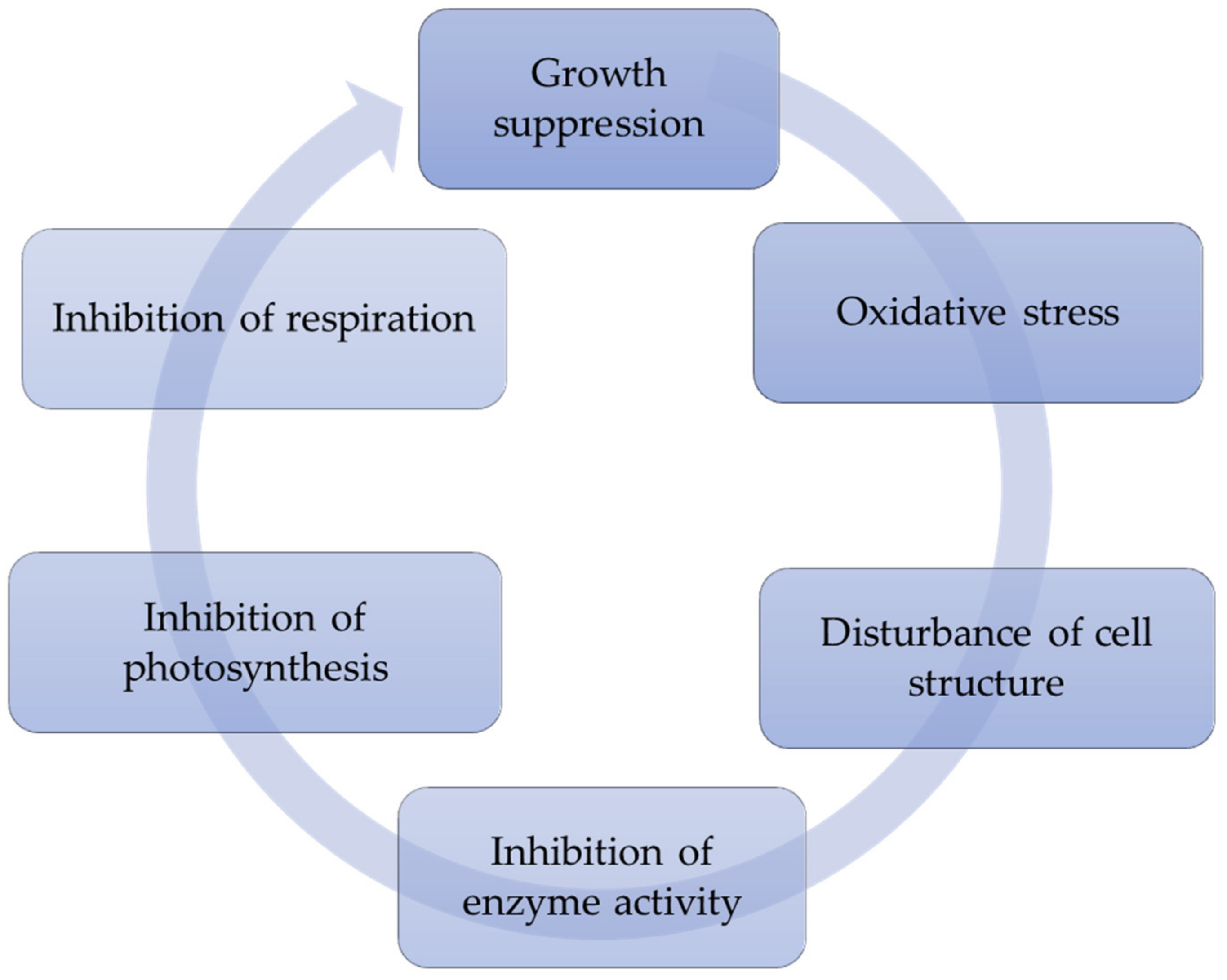
Allelopathically active compounds, as well as other stressors, affect a number of physiological and biochemical processes in phytoplankton species (Table 2), including water bloom agents, and thereby limit their growth and development (Figure 3) [58,100,101,102].
4.1. Disturbance of Cell Structure
Acting as natural barriers for cells and organelles, biomembranes are the first structures to be affected by allelochemicals [56]. Disturbance of the membrane integrity leads to disruption of the organelles’ structure and function. In some algal species, the degree of unsaturation of fatty acids increases under the influence of macrophytic allelochemicals and, as a consequence, there is an increase in fluidity and a decrease in selective permeability of cell membranes [12,16,109]. According to Li et al. [101], ethyl 2-methylacetoacetate causes changes in cell membranes of Microcystis aeruginosa. Moreover, the shape of microalgal cells changes at some concentrations of allelochemical substances that is manifested in the protoplast shrinking and its separation from the cell wall, shrinking of the nucleus volume, and impairment of the chloroplast structure [12].
4.2. Oxidative Stress
Oxidative stress is considered one of the most important physiological and biochemical effects of allelopathically active substances of aquatic vascular plants on phytoplankton [16,110,111]. Under the influence of ethyl 2-methylacetoacetate, the concentration of reactive oxygen species (ROS) and products of lipid peroxidation in algal cells increases, and the activity of some antioxidant enzymes is inhibited [16]. Allelochemicals may increase the activity of some enzymes and inhibit others due to various functional differences in enzymes [79]. Lourenção et al. [102] recorded a sharp decrease in superoxide dismutase (SOD) activity in Microcystis aeruginosa with addition of Pistia stratiotes extracts, while the activity of other enzymes that offer antioxidant protection was not inhibited. Lourenção et al. [102] demonstrated that superoxide dismutase (SOD) in Microcystis aeruginosa is a main target for bioactive substances, which are present in aqueous and ethanolic extracts of P. stratiotes.
Phenolic compounds dissolved in the waters of continental reservoirs, under certain conditions, affect the photosynthetic activity of planktonic algae, especially those that cause water blooms. The main sources of the formation of the pool of extracellular phenols are intravital and postmortem secretions of aquatic vascular plants. During biogenic leaching, easily oxidized phenols are transformed into the corresponding quinones, which, penetrating into algae cells, change the normal course of their photosynthesis. In this case, the processes of non-cyclic and cyclic phosphorylation are inhibited. Depending on the amount of quinone entering the cells of algae, their photosynthetic activity slows down or may stop completely [13,112].
4.3. Inhibition of Extracellular Enzyme Activity
Many phytoplankton species produce extracellular enzymes that enable them to use complex substrates to sustain life or colonize habitats [113]. A decrease in the activity of extracellular alkaline phosphatase is one of the main inhibitory effects of polyphenols on phytoplankton [79,80]. This enzyme is synthesized by many microalgal species experiencing inorganic phosphorus deficiency. Inactivation of alkaline phosphatase in conditions of phosphorus deficiency leads to the death of algal cells. This enzyme is especially important for homeostasis of blue-green algae [113].
4.4. Inhibition of Photosynthesis and Respiration
Allelopathically active substances of aquatic vascular plants inhibit the growth of cyanobacteria by reducing the rates of photosynthesis and respiration [12,79,114]. A decrease in microalgal respiratory rate occurred after exposure to α-asarone and allelochemicals isolated from Phragmites communis [79]. The effect of allelopathy on algal photosynthesis can be manifested in disruption of electron flow in the photosynthetic electron transport chain, a reduction in pigment content, and a change in chlorophyll-a fluorescence [17,81,83]. Many allelopathic compounds produced by aquatic vascular plants (telimagrandin II, gallic acid, pyrogallic acid etc.) inhibit photosystem II (PS II) [81,100,103]. Damage to PS II can be mitigated by de novo synthesis of the D1 protein, which is a key subunit of PS II [81,115]. Nevertheless, allelochemicals such as pyrogalic acid inhibit expression of the psbA gene that encodes the D1 protein in Microcystis Lemmermann and Cylindrospermopsis G.Seenayya and N.Subba Raju, and thus prevents protein synthesis, which promotes adaptation to stress [111]. Zhu et al. [83] showed that polyphenols strongly inhibit the photosynthetic transport of electrons in cyanobacteria but do not have a significant effect on photosynthesis of green algae, in particular because of differences in localization and structure of the photosynthetic apparatus. In addition, it has been shown that under the influence of certain allelochemical substances, in particular linoleic acid, ferulic acid, and pyrogallol of Microcystis aeruginosa, the concentration of the photosynthetic pigment chlorophyll-a is significantly reduced [58,105]. Inhibition of photosynthesis, the main physiological process in competing autotrophs, is an effective strategy for survival and colonization of aquatic habitats and is typical for many macrophyte species [100,113]. Given that aquatic vascular plants have a selective ability to inhibit the development of certain phytoplankton species, they can serve as an effective means of biocontrol against cyanobacterial agents of harmful blooms in water bodies.
5. Factors Affecting the Allelopathic Activity of Aquatic Vascular Plants
Allelopathic activity of aquatic vascular plants depends on various biotic and abiotic factors [12,16,79,81,113,116]. The complex of biotic factors that determine the impact of aquatic vascular plants on phytoplankton includes the density and benthic or surface areal cover of aquatic vegetation, species-specific features, and the growth stage of aquatic plants. Among the abiotic factors, the hydrological regime (flow rate, water exchange intensity, etc.) and physicochemical conditions (temperature, irradiance, nutrient concentration, pH, dissolved oxygen content, etc.) in water bodies are crucial.
5.1. Biotic Factors
5.1.1. Area and Density of Overgrown Areas of Aquatic Vascular Plants
A number of studies demonstrated an inhibitory effect of aquatic vascular plants on phytoplankton growth in a wide range of macrophyte areal cover (3–100%) [21,25,74,117]. The potential impact of allelopathy is particularly notable in water bodies with a high level of macrophyte coverage, 20% or more [21,24,118]. In this case, the density of macrophyte thickets plays an important role. Studies show a negative allelopathic effect of higher aquatic plants on phytoplankton at a biomass density of 1 to 10 g/dm3 (wet weight) [82,119].
5.1.2. Species-Specific Features and Growth Stage of Plants
Allelopathy is a species-specific phenomenon, that is, the same plant species or allelochemical may cause different effects on different species of algae [21,59,82,120]. For example, Ceratophyllum demersum significantly suppresses the growth of Anabaena species but does not inhibit the growth of Chlorella and Scenedesmus species [121]. The allelochemical ethyl 2-methylacetoacetate, isolated from Phragmites communis, shows high inhibitory activity against Microcystis aeruginosa, but has almost no influence on Chlorella vulgaris Beijer. [65]. Allelopathically active compounds of some species, for example, Myriophyllum spicatum, Nuphar lutea, Pistia stratiotes, Typha latifolia L., inhibit both prokaryotic and eukaryotic microalgae [79,82].
Aquatic vascular plants produce various allelochemical compounds, which have unequal inhibitory effect on phytoplankton species [16,57]. However, a number of studies have shown that prokaryotic algae are more sensitive to exometabolites of aquatic vascular plants compared to eukaryotic algae [21,58]. Allelopathically active compounds produced by plants belonging to the same phylum, or even genus may have diverse effects on phytoplankton [79,82,120]. Among Cyanobacteria, species of Dolichospermum (Ralfs ex Bornet and Flahault) P.Wacklin, L.Hoffmann and J.Komárek (=Anabaena Bory ex Bornet and Flahault), Microcystis Lemmermann and Oscillatoria Vaucher ex Gomont are highly sensitive to a wide range of allelochemicals [82,120]. In addition, allelopathic influence of aquatic vascular plants on toxic Microcystis strains is more pronounced than on non-toxic strains [120,122,123]. Ceratophyllum demersum causes a higher inhibitory effect on the microcystin-producing strain of Microcystis aeruginosa compared to Microcystis panniformis Komárek, Komárková-Legnerová, Sant’Anna, M.T.P.Azevedo, and P.A.C.Senna, which does not produce toxins [123]. This effect can be explained by the fact that under conditions of stress caused by microcystins, aquatic vascular plants produce and release allelochemicals more intensively [122,123].
The sensitivity of algae to allelopathy from aquatic vascular plants also varies depending on the growth phase and initial cell density of microalgae. Microcystis aeruginosa is inhibited by allelochemical compounds of Phragmites communis to a greater extent at the initial growth phase (lag phase) than at the rapid growth phase (logarithmic phase) [65]. The growth stage of aquatic vascular plants can also affect their allelopathic activity. Some authors report that younger and actively vegetating aquatic vascular plants demonstrate higher allelopathic activity [124,125].
5.1.3. Concentration of Allelopathically Active Compounds and Exposure Duration
Allelochemicals of aquatic vascular plants demonstrate both inhibitory and stimulating effects on cyanobacteria depending on the concentration of these compounds in water [59,81]. Wu et al. [90] studied the effect of allelochemicals isolated from Pistia stratiotes on the growth of Microcystis aeruginosa, as well as its release of microcystin-LR. Low concentrations (20–60 mg/dm3) stimulated growth of algal cultures, while high concentrations (100–200 mg/dm3) caused an inhibitory effect on growth and decreased release of microcystin-LR. Along with the concentration of allelochemical compounds, the duration of their effect is also important. The inhibitory effect of N-phenyl-1-naphthylamine on Microcystis aeruginosa was more substantial at lower doses and longer exposures [107].
5.2. Abiotic Factors
5.2.1. Hydrological Regime
Hydrological conditions, particularly the flow rate, water exchange rate, and water level regime, are important abiotic factors influencing the distribution, species composition, and metabolic activity of aquatic vegetation [126,127,128,129]. Freshwater aquatic vascular plants and planktonic microalgae develop intensively in both lentic and lotic ecosystems [113]. However, high density and species diversity of plants are usually observed at low and moderate flow rates, while the fast flow is limiting to their growth [79,113,129]. Water flow rate causes an indirect effect on plant photosynthesis, in particular by regulating the availability of dissolved nutrients [126,130]. Photosynthesis and rate of nutrient uptake by freshwater vascular plants positively correlate with lower flow rates such as 0–0.1 m/s [127,129,130]. Under these conditions, greater production and release of allelochemicals by aquatic vascular plants are observed, as well as greater accumulation of these compounds in the water [79]. Alternatively, there is intensive leaching and dilution of allelochemicals in faster water flows [79,131]. In this regard, the allelopathic influence of aquatic vascular plants on algae is considered to be more pronounced in lake systems than in river systems [79].
It should also be noted that the most promising potential for controlling blue-green algae, which cause water blooms, pertains to shallow areas of water bodies, where the content of phenolic compounds reaches 2 mg/dm3. In addition, 0.2–0.4 mg/dm3 was found to be sufficient to control water bloom, depending on the pool of these compounds [63,132] present in thickets of aquatic vascular plants.
5.2.2. Physicochemical Conditions
Among physicochemical factors that influence the growth, distribution and allelopathic activity of aquatic vascular plants, the nutrient concentration, temperature regime and light intensity should be primarily mentioned [80,113].
Low availability of nutrients increases biotic competition and enhances the allelopathic activity of aquatic vascular plants [54,133,134,135]. Phosphorus limitation causes Myriophyllum spicatum to intensify the release of telimagrandin II [81,113].
Fitzgerald [136] noted that aquatic vascular plants inhibit the growth of microalgae in aquatic environments with limited nitrogen [79]. A similar trend was observed by Mjelde and Faafeng [137], who found that aquatic vascular plants inhibit the development of phytoplankton in lakes with low content of this nutrient. Therefore, the release of allelochemicals by aquatic vascular plants is an effective protection and survival strategy in various stressful conditions.
Aquatic plants demonstrate optimal rates of photosynthesis at relatively high temperatures (between 20 and 35 °C [23,138,139]), and an increase in their growth and local distribution follows seasonal increases in water temperature [23,140]. An increase in water temperature contributes to the accumulation of aquatic vascular plant biomass and increased release of allelopathically active substances [59,85]. For example, the intensive development of Myriophyllum verticillatum is observed in August, and during this period it causes the strongest allelopathic inhibition of phytoplankton species [85]. The temperature largely determines the rate of synthesis and release of allelopathically active compounds by aquatic vascular plants, affecting the activity of enzymes involved in the biosynthesis of allelochemicals and the diffusion rate of these compounds in water.
Increased temperature may contribute to the development and spread of species such as Pontederia crassipes, Pistia stratiotes and Nuphar lutea [23,30]. Pontederia crassipes (common water hyacinth) inhibits the growth and activity of enzymes in Microcytis aeruginosa at temperatures less than 25 °C [141,142]. The optimal temperature range for the growth of water hyacinth is 25–30 °C, maximal–33–35°C [139]. Plant species Pontederia crassipes and Pistia stratiotes are known to be among the most common species in regions with warm climates [34,143].
The stability of allelochemical compounds in water depends on temperature and other factors such as light intensity, pH level and dissolved oxygen content [59,103,144,145]. Nakai et al. [103] showed that increasing temperature (from 20 to 30 °C) and light intensity (from 25 to 75 μmol m−2 s−1) reduce inhibition of Microcystis aeruginosa growth, and that this process is mediated by the impact of polyphenols ((+) catechin, eugeniin, ellagic acid, gallic acid and pyrogallic acid) and fatty acids (nonanoic acid, cis-6-octadecenoic acid and cis-9-octadecenoic acid), which are also produced by Myriophyllum spicatum [6]. Other researchers report the effect of light intensity on the stability of allelopathically active phenolic compounds in water [144,146]. Light can cause chemical changes in allelopathically active substances due to stimulation of their oxidation, polymerization or degradation [144].
Different conditions of pH and dissolved oxygen content also affect the degradation of allelochemicals, thus modifying allelopathic effects [59,62,147]. Under high values of pH (≥10) polyphenols transfer from molecular to ionic state and easily undergo oxidized polymerization, which reduces their bioavailability and toxicity [62]. Alkaloids, in contrast, exist in the molecular state at low pH [59]. For these reasons, when using aquatic vascular plants as producers of allelopathically active substances to control blue-green algal blooms, the low stability of these compounds under the influence of various abiotic factors should be considered. High pH and dissolved oxygen concentrations are the consequence of intensive growth processes and high metabolic activity of aquatic vascular plants. The low level of these indicators is associated with the intensification of accumulated phytomass decomposition. Therefore, selective extraction of biomass of fast-growing species of aquatic vascular plants should be conducted to prevent these stressful conditions.
6. The Advantages and Considerations of Using Aquatic Vascular Plants to Control Harmful Cyanobacterial Blooms
Analysis of the literature demonstrates that many species of aquatic vascular plants are characterized by significant potential for restoring water quality, and their introduction into areas of harmful “blooms” is an effective biological method to control these processes. Aquatic vascular plants are able to inhibit the growth of cyanobacteria both directly, releasing allelopathically active substances with algicidal effect, and indirectly, reducing the concentration of nutrients in water, access to light, etc.
In practical terms, allelopathy is a promising approach to fight against cyanobacterial blooms. Allelopathically active compounds produced by aquatic vascular plants are characterized by high selectivity and show a pronounced inhibitory effect against cyanobacteria, which cause harmful blooms. Their impact on other algae and aquatic organisms may be minimal or non-existent, implying that these natural mechanisms of action on cyanobacteria are environmentally safe.
Direct application of synthetic algicidal compounds into water bodies to control the growth of water-blooming pathogens may disrupt the aquatic ecosystem due to their high concentration in local areas. In addition, it may lead to fast inactivation of toxic agents and eliminate the effect of inhibitory concentrations [12,148,149]. In this regard, the use of slow-release algicides is a more effective and environmentally safer way to tackle harmful algal blooms. The combination of this approach with the allelopathic inhibiting effect of aquatic vascular plants is of particular interest.
Aquatic vascular plants affect the nutrient cycle in aquatic ecosystems. Along with associated periphyton algae, they act as “traps” for nutrients, helping to restore water-quality status and preventing biomass increase of planktonic cyanobacteria species. Different mechanisms of the growth inhibition of harmful algal taxa may act simultaneously and synergistically (for example, reduction in access to bioavailable nutrients and development of the allelopathic effects).
Despite apparent advantages, some problematic aspects should be considered in the use of aquatic vascular plants to fight against harmful algal blooms. Increasing anthropogenic eutrophication leads to excessive reproduction of aquatic vascular plants, and more competitive species may completely displace less competitive ones. For example, the increased invasive potential is typical for Pontederia crassipes, Pistia stratiotes, Trapa natans тa Nuphar lutea [23,28,34,150,151,152]. Rapidly growing aquatic vascular plants change physicochemical conditions and affect the structure and function of all hydrobionts, including fish. Abundance of fish populations as well as their diversity decrease in aquatic environments where the native structural complexity is reduced by invasive, non-indigenous species (NIS). One of the main reasons for this phenomenon might be a change in the “predator–prey” interaction pattern [153,154] and blockage of waterways and fish movement [22]. Excessive growth of aquatic vascular plants can cause anoxic conditions that lead to increased mortality of fishes and other aquatic fauna by reducing light penetration in the water column to support photosynthesis [29,34]. Therefore, mass reproduction of aquatic vascular plants can negatively impact productivity of fish populations. Selection of certain macrophyte species to prevent harmful cyanobacterial blooms should take into consideration their effects on aquatic animals such as fishes. It should be noted as well that the decomposition of their biomass may be accompanied by the release of organic and inorganic substances into the water and cause secondary pollution of water bodies.
Therefore, the use of fast-growing species of aquatic vascular plants to prevent harmful cyanobacterial blooms is an environmentally safe and effective method if accompanied by further selective or complete removal of their phytomass, as well as control of allochthonous nutrients, which contribute to their intensive reproduction.
7. Conclusions
The use of aquatic vascular plants to limit harmful algal blooms caused by cyanobacteria has a number of valuable advantages. First, they are able to limit cyanobacterial blooms by reducing dissolved nutrient concentrations in aquatic ecosystems. However, their main advantage is synthesis and release of compounds into the aquatic environment that are allelopathically specific against physiological processes of some groups of algae such as cyanobacteria. The use of these natural mechanisms to control the blue-green algae abundance is much safer for aquatic ecosystems compared to synthetic chemical compounds that can lead to secondary pollution of water bodies. However, literature analysis shows that allelopathic activity of aquatic plants depends on many biotic and abiotic factors such as the species characteristics of aquatic vascular plants, pool of biologically active compounds of exometabolites of aquatic vascular plants, area and density of their thickets in water bodies, physicochemical properties of allelopathically active substances, hydrochemical and hydrological regime, and temperature and irradiance levels. In addition, some rapidly growing macrophyte species (e.g., Pontederia crassipes, Pistia stratiotes, Trapa natans, Nuphar lutea) are able to inhibit the growth of other plants, as well as related microscopic flora and fauna, and monopolize aquatic habitats. Large dense thickets of such aquatic plants can change the “predator–prey” system, clog waterways and obstruct fish migrations, and reduce oxygenation of water bodies, which is one of the most common causes of fish productivity decline. Therefore, these species-specific effects and other characteristics should be considered in the selection of the species of aquatic vascular plants for harmful algal bloom control.
Author Contributions
Conceptualization, I.N. and O.B.; validation, I.K., M.G. and O.U.; formal analysis, T.L.; resources, I.N. and T.L.; data curation, O.B.; writing—original draft preparation, I.N.; writing—review and editing, O.B., I.N., I.A., I.K., O.U., T.L. and M.G.; visualization, O.B. and I.N.; supervision, O.B.; project administration, O.B.; funding acquisition, O.B., I.N, I.A. and T.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by the grant of the National Academy of Sciences of Ukraine to research laboratories/groups of young scientists to conduct research in priority areas of science and technology (code 6541230, Project State Reg. No. 0121U110756, grant number 26/02-2022(3),) and a Joint Excellence in Science and Humanities (JESH) Ukraine Emergency Call 2022 of the Austrian Academy of Sciences to Bilous Olena (Fund. Number 80539).
Data Availability Statement
Data are available upon request from the authors.
Acknowledgments
The authors express their appreciation for the critical review by anonymous reviewers that improved the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Stroom, J.M.; Kardinaal, W.E.A. How to combat cyanobacterial blooms: Strategy toward preventive lake restoration and reactive control measures. Aquat. Ecol. 2016, 50, 541–576. [Google Scholar] [CrossRef] [Green Version]
- Kirpenko, N.I.; Krot, Y.G.; Usenko, O.M. Surface waters “Blooms”—Fundamental and applied aspects. Hydrobiol. J. 2019, 55, 18–30. [Google Scholar] [CrossRef]
- Kirpenko, N.I.; Krot, Y.G.; Usenko, O.M. Toxicological aspects of the surface water: “Blooms” (a Review). Hydrobiol. J. 2020, 56, 3–16. [Google Scholar] [CrossRef]
- Kurashov, E.; Krylova, J.; Protopopova, E. The Use of Allelochemicals of Aquatic Macrophytes to Suppress the Development of Cyanobacterial “Blooms”. In Plankton Communities; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Tazart, Z.; Manganelli, M.; Scardala, S.; Buratti, F.M.; Di Gregorio, F.N.; Douma, M.; Mouhri, K.; Testai, E.; Loudiki, M. Remediation strategies to control toxic cyanobacterial blooms: Effects of macrophyte aqueous extracts on Microcystis aeruginosa (growth, toxin production and oxidative stress response) and on bacterial ectoenzymatic activities. Microorganisms 2021, 9, 1782. [Google Scholar] [CrossRef]
- Burford, M.A.; Beardall, J.; Willis, A.; Orr, P.T.; Magalhaes, V.F.; Rangel, L.M.; Azevedo, S.M.F.O.E.; Neilan, B.A. Understanding the winning strategies used by the bloom-forming cyanobacterium Cylindrospermopsis raciborskii. Harmful Algae 2016, 54, 44–53. [Google Scholar] [CrossRef]
- Kurmayer, R.; Deng, L.; Entfellner, E. Role of toxic and bioactive secondary metabolites in colonization and bloom formation by filamentous cyanobacteria Planktothrix. Harmful Algae 2016, 54, 69–86. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Dreher, T.W.; Li, R. An overview of diversity, occurrence, genetics and toxin production of bloom-forming Dolichospermum (Anabaena) species. Harmful Algae 2016, 54, 54–68. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, D.I.; Duquette, A.; Goodson, A.; Keppler, C.J.; Williams, S.H.; Brock, L.M.; Stackley, K.D.; White, D.; Wilde, S.B. The effects of three chemical algaecides on cell numbers and toxin content of the cyanobacteria Microcystis aeruginosa and anabaenopsis sp. J. Environ. Manag. 2014, 54, 1110–1120. [Google Scholar] [CrossRef]
- Abeynayaka, H.D.L.; Asaeda, T.; Tanaka, K.; Atsuzawa, K.; Kaneko, Y.; Nishda, H.; Inada, S. An alternative method to improve the settleability of gas-vacuolated cyanobacteria by collapsing gas vesicles. Water Supply 2016, 16, 1552–1560. [Google Scholar] [CrossRef]
- Tan, K.; Huang, Z.; Ji, R.; Qiu, Y.; Wang, Z.; Liu, J. A review of allelopathy on microalgae. Microbiology 2019, 165, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, V.D.; Sakevich, A.I.; Usenko, O.M. Higher aquatic plants as the factor limiting water bloom caused by Cyanobacteria. Rate NASU 2005, 8, 174–177. [Google Scholar]
- Romanenko, V.D.; Sakevich, A.I.; Usenko, O.M. On the mechanism of action of easily oxidizable phenols on photosynthetic activity of algae. Hydrobiol. J. 2006, 42, 82–92. [Google Scholar] [CrossRef]
- Usenko, O.M.; Guseynova, V.P.; Sakevich, A.I. Peculiarities of the influence of polyphenols on algae under conditions of changes in pH of the environment. Hydrobiol. J. 2008, 44, 37–44. [Google Scholar] [CrossRef]
- Kirpenko, N.I.; Usenko, O.M. Influence of higher aquatic plants on microalgae (a Review). Hydrobiol. J. 2013, 49, 57–74. [Google Scholar] [CrossRef]
- Tazart, Z.; Douma, M.; Tebaa, L.; Loudiki, M. Use of macrophytes allelopathy in the biocontrol of harmful Microcystis aeruginosa blooms. Water Supply 2018, 19, 245–253. [Google Scholar] [CrossRef]
- Van Donk, E.; van de Bund, W.J. Impact of submerged macrophytes including charophytes on phyto- and zooplankton communities: Allelopathy versus other mechanisms. Aquat. Bot. 2002, 72, 261–274. [Google Scholar] [CrossRef]
- Scheffer, M. The story of some shallow lakes. In Ecology of Shallow Lakes; Springer: Dordrecht, The Netherlands, 2004; pp. 1–19. [Google Scholar]
- Thomaz, S.M.; Cunha, E.R. The role of macrophytes in habitat structuring in aquatic ecosystems: Methods of measurement, causes and consequences on animal assemblages’ composition and biodiversity. Acta Limnol. Bras. 2010, 22, 218–236. [Google Scholar] [CrossRef]
- Hilt, S.; Gross, E.M. Can allelopathically active submerged macrophytes stabilise clear-water states in shallow lakes? Basic Appl. Ecol. 2008, 9, 422–432. [Google Scholar] [CrossRef] [Green Version]
- Essien, A.I.; Udoinim, M.M.; Akpan, G.D.; Effiong, J.O. Aquatic macrophytes and Limnological implications on fisheries resources: A case study of Obio Akpa Stream in Akwa Ibom State, Nigeria. Int. J. Water Soil Res. 2012, 3, 15–21. [Google Scholar]
- Henriot, C.P.; Cuenot, Q.; Levrey, L.H.; Loup, C.; Chiarello, L.; Masclaux, H.; Bornette, G. Relationships between key functional traits of the waterlily Nuphar lutea and wetland nutrient content. Peer J. 2019, 7, e7861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blindow, I.; Hargeby, A.; Andersson, G. Seasonal changes of mechanisms maintaining clear water in a shallow lake with abundant Chara vegetation. Aquat. Bot. 2002, 72, 315–334. [Google Scholar] [CrossRef]
- Meijer, M.L.; Jeppesen, E.; Van Donk, E.; Moss, B.; Scheffer, M.; Lammens, E.; van Nes, E.; Van Berkum, J.A.; De Jong, G.J.; Faafeng, B.A.; et al. Long-term responses to fish-stock reduction in small shallow lakes: Interpretation of five-year results of four biomanipulation cases in The Netherlands and Denmark. Hydrobiologia 1994, 275–276, 457–466. [Google Scholar] [CrossRef]
- Pokorný, J.; Květ, J.; Ondok, J.; Toul, Z.; Ostrý, I. Production-ecological analysis of a plant community dominated by Elodea canadensis michx. Aquat. Bot. 1984, 19, 263–292. [Google Scholar] [CrossRef]
- Ozimek, T.; Gulati, R.D.; van Donk, E. Can macrophytes be useful in biomanipulation of lakes? The Lake Zwemlust example. Hydrobiologia 1990, 200–201, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Soloviy, K.; Malovanyy, M. Freshwater ecosystem macrophytes and microphytes: Development, environmental problems, usage as raw material. Review. Environ. Probl. 2019, 4, 115–124. [Google Scholar] [CrossRef]
- Wu, M.Y.; Wu, J. In-vitro investigations on ultrasonic control of water chestnut. J. Aquat. Plant Manag. 2007, 45, 76–83. [Google Scholar]
- Cunha, D.G.F.; Bottino, F.; do Carmo Calijuri, M. Can free-floating and emerged macrophytes influence the density and diversity of phytoplankton in subtropical reservoirs? Lake Reserv. Manag. 2012, 28, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Kiviat, E. Water chestnut (Trapa natans) In Exotic Plants with Identified Detrimental Impacts on Wildlife Habitats; New York State Cornell University: New York, NY, USA, 1987; pp. 31–38. [Google Scholar]
- Kiviat, E. Under the spreading water-chestnut. News Hudsonia 1993, 9, 1–6. [Google Scholar]
- Groth, A.T.; Lovett-Doust, L.; Lovett-Doust, J. Population density and module demography in Trapa natans (Trapaceae), an annual, clonal aquatic macrophyte. Am. J. Bot. 1996, 83, 1406–1415. [Google Scholar] [CrossRef]
- Mironga, J.M.; Mathooko, J.M.; Onywere, S.M. The effect of water Hyacinth (Eichhornia Crassipes) infestation on phytoplankton productivity in Lake Naivasha and the status of control. J. Environ. Sci. Eng. 2011, 5, 1252–1260. [Google Scholar]
- Van Donk, E.; Gulati, R.D.; Iedema, A.; Meulemans, J.T. Macrophyte-related shifts in the nitrogen and phosphorus contents of the different trophic levels in a biomanipulated shallow lake. Hydrobiologia 1993, 251, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Barko, J.W.; James, W.F. Effects of submerged aquatic macrophytes on nutrient dynamics, sedimentation, and resuspension. In The Structuring Role of Submerged Macrophytes in Lakes; Springer: New York, NY, USA, 1998; pp. 197–214. [Google Scholar] [CrossRef]
- Seto, M.; Takamura, N.; Iwasa, Y. Individual and combined suppressive effects of submerged and floating-leaved macrophytes on algal blooms. J. Theor. Biol. 2013, 319, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Samal, K.; Kar, S.; Trivedi, S.; Upadhyay, S. Assessing the impact of vegetation coverage ratio in a floating water treatment bed of Pistia stratiotes. SN Appl. Sci. 2021, 3, 120. [Google Scholar] [CrossRef]
- Blindow, I. The composition and density of epiphyton on several species of submerged macrophytes—The neutral substrate hypothesis tested. Aquat. Bot. 1987, 29, 157–168. [Google Scholar] [CrossRef]
- Weisner, S.; Eriksson, G.; Granéli, W.; Leonardson, L. Influence of macrophytes on nitrate removal in wetlands. Ambio 1994, 23, 363–366. [Google Scholar]
- Koschel, R.; Benndorf, J.; Proft, G.; Recknagel, F. Calcite precipitation as a natural control mechanism of eutrophication. Arch. Hydrobiol. 1983, 98, 380–408. [Google Scholar]
- Reynolds, C.S. The Ecology of Freshwater Phytoplankton; Cambridge University Press: Cambridge, UK, 1984; p. 384. [Google Scholar]
- Smith, R.V.; Peat, A. Comparative structure of the gas-vacuoles of blue-green algae. Archiv. Mikrobiol. 1967, 57, 111–122. [Google Scholar] [CrossRef]
- Walsby, A.E. The gas vacuoles of blue-green algae. Sci. Am. 1977, 237, 90–97. [Google Scholar] [CrossRef]
- Walsby, A.E.; Avery, A.; Schanz, F. The critical pressures of gas vesicles in Planktorhrix rubescens in relation tothe depth of winter mixing in Lake Zürich, Switzerland. J. Plankton Res. 1998, 20, 1357–1375. [Google Scholar] [CrossRef] [Green Version]
- Timms, R.M.; Moss, B. Prevention of growth of potentially dense phytoplankton populations by zooplankton grazing, in the presence of zooplanktivorous fish, in a shallow wetland ecosystem. Limnol. Oceanogr. 1984, 29, 472–486. [Google Scholar] [CrossRef]
- Lauridsen, T.; Pedersen, L.J.; Jeppesen, E.; Sønergaard, M. The importance of macrophyte bed size for cladoceran composition and horizontal migration in a shallow lake. J. Plankton Res. 1996, 18, 2283–2294. [Google Scholar] [CrossRef] [Green Version]
- Basińska, A.; Kuczyńska-Kippen, N. Differentiated macrophyte types as a habitat for rotifers in small mid-forest water bodies. Biologia 2009, 64, 1100–1107. [Google Scholar] [CrossRef] [Green Version]
- Basu, B.K.; Kalff, J.; Pinel-Alloul, B. The influence of macrophyte beds on plankton communities and their export from fluvial lakes in the St Lawrence River. Freshw. Biol. 2000, 45, 373–382. [Google Scholar] [CrossRef]
- Manatunge, J.; Asaeda, T.; Priyadarshana, T. The influence of structural complexity on fish-zooplankton interactions: A study using artificial submerged macrophytes. Environ. Biol. Fish. 2000, 58, 425–438. [Google Scholar] [CrossRef]
- Kuczynska-Kippen, N. Zooplankton structure in architecturally differentiated macrophyte habitats of shallow lakes in the Wielkopolska Region, Poland. Oceanol. Hydrobiol. Stud. 2006, 35, 179–191. [Google Scholar]
- Konovets, I.M.; Usenko, O.M.; Mardarevich, M.G.; Kudryavtseva, D.O. Influence of phenolcarbonic acids on Daphnia magna Straus fertility. Hydrobiol. J. 2021, 57, 80–88. [Google Scholar] [CrossRef]
- Kuczynska-Kippen, N.; Nagengast, B. The impact of the architecture of macrophytes on the spatial structure of zooplankton of the Wielkowiejskie lake. Rocz. AR Pozn. CCCLIV. 2003, 6, 121–129. [Google Scholar]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984; p. 422. [Google Scholar]
- Usenko, O.M.; Sakevich, A.I. Allelopathic influence of higher aquatic plants on the functional activity of plankton algae. Hydrobiol. J. 2005, 41, 54–66. [Google Scholar] [CrossRef]
- Fink, P. Ecological functions of volatile organic compounds in aquatic systems. Mar. Freshw. Behav. Physiol. 2007, 40, 155–168. [Google Scholar] [CrossRef]
- Sakevych, O.I.; Usenko, O.M. Allelopathy in Hydroecosystems; Logos: Kiev, Ukraine, 2008; p. 342. (In Ukrainian) [Google Scholar]
- Zhu, X.; Dao, G.; Tao, Y.; Zhan, X.; Hu, H. A review on control of harmful algal blooms by plant-derived allelochemicals. J. Hazard. Mater. 2021, 401, 123403. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Yin, Y.; Kang, L.; Feng, L.; Liu, Y.; Du, Z.; Tian, Y.; Zhang, L. A review: Application of allelochemicals in water ecological restoration—Algal inhibition. Chemosphere 2021, 267, 128869. [Google Scholar] [CrossRef] [PubMed]
- Ren, S. Allelopathy in Sustainable Agriculture and Forestry; Springer: New York, NY, USA, 2008; p. 426. [Google Scholar]
- Zhang, T.T.; Zheng, C.Y.; He, M.; Wu, A.P.; Nie, L.W. Inhibition on algae of fatty acids and the structure-effect relationship. China Environ. Sci. 2009, 29, 274–279. [Google Scholar]
- Gao, Y.N.; Dong, J.; Fu, Q.Q.; Wang, Y.P.; Chen, C.; Li, J.H.; Li, R.; Zhou, C.J. Allelopathic effects of submerged macrophytes on phytoplankton. Allelopath. J. 2017, 40, 01–22. [Google Scholar] [CrossRef] [Green Version]
- Usenko, O.M.; Sakevych, O.I. Resistance of Algae to Biologically Active Substances; Logos: Kiev, Ukraine, 2010; p. 192. (In Ukrainian) [Google Scholar]
- Nakai, S.; Inoue, Y.; Hosomi, M. Algal growth inhibition effects and inducement modes by plant-producing phenols. Water Res. 2001, 35, 1855–1859. [Google Scholar] [CrossRef]
- Li, F.M.; Hu, H.Y. Isolation and characterization of a novel antialgal allelochemical from Phragmites communis. Appl. Environ. Microbiol. 2005, 71, 6545–6553. [Google Scholar] [CrossRef] [Green Version]
- Nakai, S.; Zou, G.; Okuda, T.; Nishijima, W.; Hosomi, M.; Okada, M. Polyphenols and fatty acids responsible for anti-cyanobacterial allelopathic effects of submerged macrophyte Myriophyllum spicatum. Water Sci. Technol. 2012, 66, 993–999. [Google Scholar] [CrossRef]
- Nakai, S.; Inoue, Y.; Hosomi, M.; Murakami, A. Myriophyllum spicatum-released allelopathic polyphenols inhibiting growth of blue-green algae Microcystis aeruginosa. Water Res. 2000, 34, 3026–3032. [Google Scholar] [CrossRef]
- Usenko, O.M. Comparison studies on the content of phenols and quinones in the phytomass of higher aquatic plants under natural conditions. Hydrobiol. J. 2012, 48, 73–80. [Google Scholar] [CrossRef]
- Usenko, O.M. Endo- and exogenous phenolcarboxylic acids Trapa natans L. and Nuphar lutea L. TNPU Ser. Biol. 2015, 3–4, 675–678. (In Ukrainian) [Google Scholar]
- Planas, D.; Sarhan, F.; Dube, L.; Godmaire, H.; Cadieux, C. Ecological significance of phenolic compounds of Myriophyllum spicatum. Verh. Internat. Verein. Limnol. 1981, 21, 1492–1496. [Google Scholar] [CrossRef]
- Saito, K.; Matsumoto, M.; Sekine, T.; Murakoshi, I.; Morisaki, N.; Iwasaki, S. Inhibitory substances from Myriophyllum brasiliense on growth of blue-green algae. J. Nat. Prod. 1989, 52, 1221–1226. [Google Scholar] [CrossRef]
- Gross, E.M. Seasonal and spatial dynamics of allelochemicals in the submersed macrophyte Myriophyllum spicatum L. SIL Proc. 2000, 27, 2116–2119. [Google Scholar] [CrossRef] [Green Version]
- Usenko, O.M.; Konovets, I.M. Analysis of pheolcarbonic acids content in phytomass of higher aquatic plants. Hydrobiol. J. 2014, 50, 47–60. [Google Scholar] [CrossRef]
- Usenko, O.M.; Konovets, I.N.; Tarashchuk, O.S.; Gorbunova, Z.N. Phenolcarbonic acids of the submerged aquatic plants and their effect on phytoepiphyton structure. Hydrobiol. J. 2019, 55, 55–64. [Google Scholar] [CrossRef]
- Bi, X.; Dai, W.; Zhang, S.; Pang, Y.; Xing, K.Z.; Zhang, D.J. Inhibition of photosynthesis related gene expression by berberine in Microcystis aeruginosa. Allelopath. J. 2014, 34, 335–343. [Google Scholar]
- Canton, M.C.; Holguin, F.O.; Boeing, W.J. Alkaloid gramine to control algal invaders: Algae inhibition and gramine persistence. Bioresour. Technol. Rep. 2019, 7, 100304. [Google Scholar] [CrossRef]
- Balanda, O.V.; Medved, V.A.; Sakevich, A.I. Alkaloids of Nuphar lutea (L.) Smith. and their influence on the vital activity of cyanobacteria and algae. Hydrobiol. J. 2004, 40, 108–120. [Google Scholar] [CrossRef]
- Usenko, O.M.; Balanda, O.V. Terpene compounds of water plants and their impact on the functional activity of hydrobionts. Visn. Lviv Univ. Ser. Biol. 2013, 62, 216–226. (In Ukrainian) [Google Scholar]
- Hu, H.; Hong, Y. Algal-bloom control by allelopathy of aquatic macrophytes—A review. Front. Environ. Sci. Eng. China 2008, 2, 421–438. [Google Scholar] [CrossRef]
- Palamarchuk, V.D. Influence of phenol acids of hydrophytes on the development of plankton algae. Int. J. Algae 2002, 4, 20–28. [Google Scholar] [CrossRef]
- Mohamed, Z.A. Macrophytes-cyanobacteria allelopathic interactions and their implications for water resources management—A review. Limnologica 2017, 63, 122–132. [Google Scholar] [CrossRef]
- Körner, S.; Nicklisch, A. Allelopathic growth inhibition of selected phytoplankton species by submerged macrophytes. J. Phycol. 2002, 38, 862–871. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, B.; Wang, J.; Gao, Y.; Wu, Z. Study on the mechanism of allelopathic influence on cyanobacteria and chlorophytes by submerged macrophyte (Myriophyllum spicatum) and its secretion. Aquat. Toxicol. 2010, 98, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Aliotta, G.; Molinaro, A.; Monaco, P.; Pinto, G.; Previtera, L. Three biologically active phenylpropanoid glucosides from Myriophyllum verticillatum. Phytochemistry 1992, 31, 109–111. [Google Scholar] [CrossRef]
- Hilt, S.; Ghobrial, M.; Gross, E.M. In situ allelopathic potential of Myriophyllum verticillatum (Haloragaceae) against selected phytoplankton species. J. Phycol. 2006, 42, 1189–1198. [Google Scholar] [CrossRef] [Green Version]
- Erhard, D.; Gross, E.M. Allelopathic activity of Elodea canadensis and Elodea nuttallii against epiphytes and phytoplankton. Aquat. Bot. 2006, 85, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.H.; Yu, S.W.; Yang, S.Y.; Zhao, B.W.; Yu, Z.W.; Wu, H.L.; Huang, S.Y.; Tang, C.S. Allelochemicals from root exudates of water hyacinth (Eichhornis crassipes). Acta Photophysiol. Sin. 1993, 19, 92–96. [Google Scholar]
- Shanab, S.M.M.; Shalaby, E.A.; Lightfoot, D.A.; El-Shemy, H.A. Allelopathic effects of water hyacinth Eichhornia crassipes. PLoS ONE 2010, 5, e13200. [Google Scholar] [CrossRef]
- Gross, E.M.; Erhard, D.; Iványi, E. Allelopathic activity of Ceratophyllum demersum L. and Najas marina ssp. intermedia (Wolfgang) Casper. Hydrobiologia 2003, 506, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wu, H.; Chen, J.; Ye, J. Effects of allelochemical extracted from water lettuce (Pistia stratiotes Linn.) on the growth, microcystin production and release of Microcystis aeruginosa. Environ. Sci. Pollut. Res. 2013, 20, 8192–8201. [Google Scholar] [CrossRef] [PubMed]
- Aliotta, G.; Monaco, P.; Pinto, G.; Pollio, A.; Previtera, L. Potential allelochemicals from Pistia stratiotes L. J. Chem. Ecol. 1991, 17, 2223–2234. [Google Scholar] [CrossRef] [PubMed]
- Waridel, P.; Wolfender, J.L.; Lachavanne, J.B.; Hostettmann, K. ent-Labdane diterpenes from the aquatic plant Potamogeton pectinatus. Phytochemistry 2003, 64, 1309–1317. [Google Scholar] [CrossRef]
- Cangiano, T.; DellaGreca, M.; Fiorentino, A.; Isidori, M.; Monaco, P.; Zarrelli, A. Lactone diterpenes from the aquatic plant Potamogeton natans. Phytochemistry 2001, 56, 469–473. [Google Scholar] [CrossRef]
- Zhang, S.; Xia, W.; Yang, X.; Zhang, T. Inhibition effect on Microcystis aeruginosa PCC7806 as well as separation and identification of algicidal substances isolated from Salvinia natans (L.). All. J. Hyg. Res. 2016, 45, 442–447. [Google Scholar]
- Zhou, S.; Nakai, S.; Hosomi, M.; Sezaki, Y.; Tominaga, M. Allelopathic growth inhibition of cyanobacteria by reed. Allelopath. J. 2006, 18, 277–285. [Google Scholar]
- Zhang, T.T.; Hu, W.; Zhang, D. Allelopathic effect of Typha Angustifolia L. on phytoplankton. Adv. Mat. Res. 2011, 383–390, 3724–3728. [Google Scholar] [CrossRef]
- Kang, P.G.; Hong, J.; Kim, E.; Kim, B. Effects of extracts of reed and cattail on the growth of a cyanobacterium, Microcystis aeruginosa. J. Freshw. Ecol. 2020, 35, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Aliotta, G.; Greca, M.D.; Monaco, P.; Pinto, G.; Pollio, A.; Previtera, L. In vitro algal growth inhibition by phytotoxins of Typha latifolia L. J. Chem. Ecol. 1990, 16, 2637–2646. [Google Scholar] [CrossRef]
- Wang, H.Q.; Liang, F.; Qiao, N.; Dong, J.X.; Zhang, L.Y.; Guo, Y.F. Chemical composition of volatile oil from two emergent plants and their algae inhibition activity. Pol. J. Environ. Stud. 2014, 6, 2371–2374. [Google Scholar]
- Leu, E.; Krieger-Liszkay, A.; Goussias, C.; Gross, E.M. Polyphenolic allelochemicals from the aquatic angiosperm Myriophyllum spicatum L. inhibit photosystem II. Plant Physiol. 2002, 130, 2011–2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.M.; Hu, H.Y.; Chong, Y.X.; Men, Y.J.; Guo, M.T. Effects of allelochemical EMA isolated from Phragmites communis on algal cell membrane lipid and ultrastructure. Huan Jing Ke Xue 2007, 28, 1534–1538. [Google Scholar] [PubMed]
- Lourenção, A.; Mecina, G.F.; Cordeiro-Araújo, M.K.; Bittencourt-Oliveira, M.C.; Chia, M.A.; Bronzel-Júnior, J.L.; Granero, F.O.; Silva, L.P.; da Silva, R.M.G. Characterization of allelochemicals from Pistia stratiotes extracts and their effects on the growth and physiology of Microcystis aeruginosa. Environ. Sci. Pollut. Res. 2021, 28, 57248–57259. [Google Scholar] [CrossRef] [PubMed]
- Nakai, S.; Asaoka, S.; Okuda, T.; Nishijima, W. Growth inhibition of Microcystis aeruginosa by allelopathic compounds originally isolated from Myriophyllum spicatum: Temperature and light efects and evidence of possible major mechanisms. J. Chem. Eng. Jpn. 2014, 47, 488–493. [Google Scholar] [CrossRef]
- Nakai, S.; Yamada, S.; Hosomi, M. Anti-cyanobacterial fatty acids released from Myriophyllum spicatum. Hydrobiologia 2005, 543, 71–78. [Google Scholar] [CrossRef]
- Ni, L.; Rong, S.; Gu, G.; Hu, L.; Wang, P.; Li, D.; Yue, F.; Wang, N.; Wu, H.; Li, S. Inhibitory effect and mechanism of linoleic acid sustained-release microspheres on Microcystis aeruginosa at different growth phases. Chemosphere 2018, 212, 654–661. [Google Scholar] [CrossRef]
- Li, F.M.; Hu, H.Y.; Chong, Y.X.; Men, Y.J.; Guo, M.T.I. Influence of EMA isolated from Phragmites communis on physiological characters of Microcystis aeruginosa. J. Environ. Sci. 2007, 27, 377–381. [Google Scholar]
- Gao, Y.N.; Ge, F.J.; Zhang, L.P.; He, Y.; Lu, Z.Y.; Zhang, Y.Y.; Liu, B.Y.; Zhou, Q.H.; Wu, Z.B. Enhanced toxicity to the cyanobacterium Microcystis aeruginosa by low-dosage repeated exposure to the allelochemical N-phenyl-1-naphthylamine. Chemosphere 2017, 174, 732–738. [Google Scholar] [CrossRef]
- Wang, H.Q.; Zhu, H.J.; Zhang, L.Y.; Xue, W.J.; Yuan, B. Identification of antialgal compounds from the aquatic plant Elodea nuttallii. Allelopath. J. 2014, 34, 207–213. [Google Scholar]
- Xin, L.; Hong-Ying, H.; Jia, Y.; Yin-Hu, W. Enhancement effect of ethyl-2-methyl acetoacetate on triacylglycerols production by a freshwater microalga, Scenedesmus sp. LX1. Bioresour. Technol. 2010, 101, 9819–9821. [Google Scholar] [CrossRef]
- Shao, J.; Wu, Z.; Yu, G.; Peng, X.; Li, R. Allelopathic mechanism of pyrogallol to Microcystis aeruginosa PCC7806 (Cyanobacteria): From views of gene expression and antioxidant system. Chemosphere 2009, 75, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Shi, J.; Yang, S. The effect of pyrogallic acid on growth, oxidative stress, and gene expression in Cylindrospermopsis raciborskii (Cyanobacteria). Ecotoxicology 2013, 22, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, V.D.; Usenko, O.M.; Sakevich, A.I. Metabolic mechanisms of relationship between higher aquatic plants and water bloom forming blue-green algae. Hydrobiol. J. 2005, 41, 43–55. [Google Scholar] [CrossRef]
- Gross, E.M. Allelopathy of aquatic autotrophs. Crit. Rev. Plant Sci. 2003, 22, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.T.; Zheng, C.Y.; Hu, W.; Xu, W.W.; Wang, H.F. The allelopathy and allelopathic mechanism of phenolic acids on toxic Microcystis aeruginosa. J. Appl. Phycol. 2010, 22, 71–77. [Google Scholar] [CrossRef]
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. Biochemistry and Molecular Biology of Plants; John Wiley and Sons: Hoboken, NJ, USA, 2015; p. 1280. [Google Scholar]
- Gopal, B.; Goel, U. Competition and allelopathy in aquatic plant communities. Bot. Rev. 1993, 59, 155–210. [Google Scholar] [CrossRef]
- Ruggiero, A.; Solimini, A.G.; Carchini, G. Nutrient and chlorophyll a temporal patterns in eutrophic mountain ponds with contrasting macrophyte coverage. Hydrobiologia 2003, 506–509, 657–663. [Google Scholar] [CrossRef]
- Canfield, D.E., Jr.; Shireman, J.V.; Colle, D.E.; Haller, W.T.; Watkins, C.E., II; Maceina, M.J. Prediction of chlorophyll a concentrations in florida lakes: Importance of aquatic macrophytes. Can. J. Fish. Aquat. Sci. 1984, 41, 497–501. [Google Scholar] [CrossRef]
- Nakai, S.; Inoue, Y.; Hosomi, M.; Murakami, A. Growth inhibition of blue-green algae by allelopathic effects of macrophytes. Water Sci. Technol. 1999, 39, 47–53. [Google Scholar] [CrossRef]
- Mulderij, G.; Mooij, W.M.; Smolders, A.J.P.; Donk, E.V. Allelopathic inhibition of phytoplankton by exudates from Stratiotes aloides. Aquat. Bot. 2005, 82, 284–296. [Google Scholar] [CrossRef]
- Kogan, S.I.; Chinnova, G.A. Relations between Ceratophyllum demersum (L.) and some blue-green algae. Hydrobiol. J. 1972, 8, 21–27. [Google Scholar]
- Amorim, C.A.; Ulisses, C.; Moura, A.N. Biometric and physiological responses of Egeria densa Planch. cultivated with toxic and non-toxic strains of Microcystis. Aquat. Toxicol. 2017, 191, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Amorim, C.A.; Moura-Falcão, R.H.D.; Valença, C.R.; Souza, V.R.D.; Moura, A.D.N. Allelopathic effects of the aquatic macrophyte Ceratophyllum demersum L. on phytoplankton species: Contrasting effects between cyanobacteria and chlorophytes. Acta Limnol. Bras. 2019, 31, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mulderij, G.; Van Donk, E.; Roelofs, J.G.M. Differential sensitivity of green algae to allelopathic substances from Chara. Hydrobiologia 2003, 491, 261–271. [Google Scholar] [CrossRef]
- Kurashov, E.A.; Krylov, Y.V.; Chernova, A.M.; Mytrukova, G.G. Component composition of volatile low-molecular organic substances Nuphar lutea (Nymphaeaceae) at the beginning of the growing season. Water Chem. Ecol. 2013, 5, 67–80. (In Ukrainian) [Google Scholar]
- Westlake, D.F. Some effects of low-velocity currents on the metabolism of aquatic macrophytes. J. Exp. Bot. 1967, 18, 187–205. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Westlake, D.F. Water velocities around water plants in chalk streams. Folia Geobot. 1990, 25, 279–289. [Google Scholar] [CrossRef]
- Riis, T.; Biggs, B.J.F. Hydrologic and hydraulic control of macrophyte establishment and performance in streams. Limnol. Oceanogr. 2003, 48, 1488–1497. [Google Scholar] [CrossRef] [Green Version]
- Franklin, P.; Dunbar, M.; Whitehead, P. Flow controls on lowland river macrophytes: A review. Sci. Total Environ. 2008, 400, 369–378. [Google Scholar] [CrossRef]
- Madsen, J.D.; Chambers, P.A.; James, W.F.; Koch, E.W.; Westlake, D.F. The interaction between water movement, sediment dynamics and submersed macrophytes. Hydrobiologia 2001, 444, 71–84. [Google Scholar] [CrossRef]
- Wetzel, R.G. Limnology: Lake and River Ecosystems, 3rd ed.; Academic Press: San Diego, CA, USA, 2001; p. 1006. [Google Scholar] [CrossRef]
- Sakevich, A.I.; Usenko, O.M. Phenol compounds in the water of the Dnieper reservoirs. Hydrobiol. J. 2003, 39, 37–47. [Google Scholar] [CrossRef]
- Reigosa, M.J.; Sánchez-Moreiras, A.; González, L. Ecophysiological approach in allelopathy. Crit. Rev. Plant. Sci. 1999, 18, 577–608. [Google Scholar] [CrossRef]
- Maberly, S.C.; Spence, D.H.N. Photosynthesis and photorespiration in freshwater organisms: Amphibious plants. Aquat. Bot. 1989, 34, 267–286. [Google Scholar] [CrossRef]
- Arthaud, F.; Vallod, D.; Robin, J.; Bornette, G. Eutrophication and drought disturbance shape functional diversity and life-history traits of aquatic plants in shallow lakes. Aquat. Sci. 2011, 74, 471–481. [Google Scholar] [CrossRef]
- Fitzgerald, G.P. Some factors in the competition or antagonism among bacteria, algae and aquatie weeds. J. Phycol. 1969, 5, 351. [Google Scholar] [CrossRef] [PubMed]
- Mjelde, M.; Faafeng, B. Ceratophyllum demersum hampers phytoplankton development in some small Norwegian lakes over a wide range of phosphorus concentrations and geographical latitude. Freshw. Biol. 1997, 37, 355–365. [Google Scholar] [CrossRef]
- Bornette, G.; Puijalon, S. Macrophytes: Ecology of aquatic plants. In Encyclopedia of Life Sciences; John Wiley and Sons Ltd.: Chichester, UK, 2009; pp. 1–9. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Brunel, S. Assessing and managing the current and future pest risk from water hyacinth, (Eichhornia crassipes), an invasive aquatic plant threatening the environment and water security. PLoS ONE 2016, 11, e0120054. [Google Scholar] [CrossRef]
- Santamaría, L.; van Vierssen, W. Photosynthetic temperature responses of fresh- and brackish-water macrophytes: A review. Aquat. Bot. 1997, 58, 135–150. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, Z.; Chen, D.; Zhang, J.; Yang, W.; Jin, Y. Allelopathic effects of Eichhornia crassipes on the growth of Microcystis aeruginosa. J. Agric. Sci. Technol. 2012, 2, 1400–1406. [Google Scholar]
- Wu, X.X.; Zhang, Z.Y.; Jin, Y.G. Physiological mechanism of Eichhornia crassipes in inhibiting the growth of Microcytis aeruginosa. Russ. J. Plant Physiol. 2019, 66, 433–439. [Google Scholar] [CrossRef]
- Meerhoff, M.; Mazzeo, N.; Moss, B.; Rodríguez-Gallego, L. The structuring role of free-floating versus submerged plants in a subtropical shallow lake. Aquat Ecol. 2003, 37, 377–391. [Google Scholar] [CrossRef]
- Gross, E.M.; Hilt, S.; Lombardo, P.; Mulderij, G. Searching for allelopathic effects of submerged macrophytes on phytoplankton—state of the art and open questions. Hydrobiologia 2007, 584, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.B.; Zhang, Y.Y.; Liu, B.Y. Allelopathy of Aquatic Macrophytes on Phytoplankton; Science Press: Beijing, China, 2016; pp. 44–80. [Google Scholar]
- Erhard, D.; Gross, E.M. Do environmental factors influence composition of potential allelochemicals in the submersed freshwater macrophyte Elodea nuttallii (Hydrocharitaceae)? Verh. Int. Verein. Limnol. 2005, 29, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Marchiosi, R.; dos Santos, W.D.; Constantin, R.P.; de Lima, R.B.; Soares, A.R.; Finger-Teixeira, A.; Mota, T.R.; de Oliveira, D.M.; Foletto-Felipe, M.d.P.; Abrahão, J. Biosynthesis and metabolic actions of simple phenolic acids in plants. Phytochem. Rev. 2020, 19, 865–906. [Google Scholar] [CrossRef]
- Huang, H.; Xiao, X.; Lin, F.; Grossart, H.P.; Nie, Z.; Sun, L.; Xu, C.; Shi, J. Continuous-release beads of natural allelochemicals for the long-term control of cyanobacterial growth: Preparation, release dynamics and inhibitory effects. Water Res. 2016, 95, 113–123. [Google Scholar] [CrossRef]
- Ni, L.; Acharya, K.; Ren, G.; Li, S.; Li, Y.; Li, Y. Preparation and characterization of anti-algal sustained-release granules and their inhibitory effects on algae. Chemosphere 2013, 91, 608–615. [Google Scholar] [CrossRef]
- Brundu, G.; Stinca, A.; Angius, L.; Bonanomi, G.; Celesti-Grapow, L.; D’Auria, G.; Griffo, R.; Migliozzi, A.; Motti, R.; Spigno, P. Pistia stratiotes L. and Eichhornia crassipes (Mart.) Solms.: Emerging invasive alien hydrophytes in Campania and Sardinia (Italy). EPPO Bull. 2012, 42, 568–579. [Google Scholar] [CrossRef]
- Yang, X.; Chen, S.; Zhang, R. Utilization of two invasive free-floating aquatic plants (Pistia stratiotes and Eichhornia crassipes) as sorbents for oil removal. Environ. Sci. Pollut. Res. 2014, 21, 781–786. [Google Scholar] [CrossRef]
- Ayanda, O.I.; Ajayi, T.; Asuwaju, F.P. Eichhornia crassipes (Mart.) Solms: Uses, Challenges, Threats, and Prospects. Sci. World J. 2020, 2020, 3452172. [Google Scholar] [CrossRef]
- Petr, T. Interactions between Fish and Aquatic Macrophytes in Inland Waters—A Review; FAO Fisheries Technical Paper; FAO: Rome, Italy, 2000; p. 185. [Google Scholar]
- Egertson, C.J.; Downing, J.A. Relationship of fish catch and composition to water quality in a suite of agriculturally eutrophic lakes. Can. J. Fish. Aquat. Sci. 2004, 61, 1784–1796. [Google Scholar] [CrossRef]
Figure 1.
The mechanism of inhibition of cyanobacteria and other phytoplankton growth depends on the relative area of aquatic macrophyte thickets [21,24,25].

Figure 2.
Floating aquatic plants (Trapa natans L., Nuphar lutea (L.) Smith) in the Kyiv Reservoir (Dnipro River, Ukraine).
Figure 2.
Floating aquatic plants (Trapa natans L., Nuphar lutea (L.) Smith) in the Kyiv Reservoir (Dnipro River, Ukraine).

Figure 3.
Physiological and biochemical mechanisms of influence of allelopathically active compounds of aquatic vascular plants on target phytoplankton species.
Figure 3.
Physiological and biochemical mechanisms of influence of allelopathically active compounds of aquatic vascular plants on target phytoplankton species.


{kind=link}
{kind=link}
{kind=link}
Table 1.
The main allelopathically active compounds of some aquatic vascular plant species, which have inhibitory effects on phytoplankton.
Table 1.
The main allelopathically active compounds of some aquatic vascular plant species, which have inhibitory effects on phytoplankton.
| Macrophyte Species | Allelopathically Active Compounds | Phytoplankton Species, Subjected to Inhibitory Effects | Literature Source |
| Hydrophytes | |||
| Spiked water-milfoil (Myriophyllum spicatum L.) | Tannins (telimagrandin II); Phenolic acids (gallic, ellagic, pyrogallic); Flavonoids ((+)-catechin); Fatty acids (nonanoic, oleic, petroselinic) | Limnothrix redekei (Goor) Meffert, Microcystis aeruginosa (Kützing) Kützing, Planktothrix agardhii (Gomont) Anagnostidis and Komárek, | [66,82,83] |
| Whorled water-milfoil (Myriophyllum verticillatum L.) | Phenylpropanoids (α-asarone) | Limnothrix redekei, Microcystis aeruginosa | [84,85] |
| Water caltrop (Trapa natans L.) | Phenolic acids (benzoic, p-hydroxybenzoic, salicylic, cinnamic, α-resorcylic, protocatechuic, coumaric, vanillic, gallic, caffeic, ferulic, syringic, ß-resorcylic and sinapinic) | Microcystis aeruginosa, phytoplankton in general | [5,69] |
| Yellow waterlily (Nuphar lutea (L.) Sm.) | Fatty acids (hexadecanoic acid, tetradecanoic acid); Esters of phthalic acid (dibutyl phthalate, diisobutyl phthalate) | Microcystis aeruginosa | [5] |
| Canadian waterweed (Elodea canadensis Michx.) | Flavonoids (luteolin 7-O-diglucuronide, apigenin 7-O-diglucuronide, chrysoeriol 7-O-diglucuronide) | Microcystis aeruginosa | [81,86] |
| Common water hyacinth (Pontederia crassipes Mart. = Eichhornia crassipes Mart.) | Amines (N-phenyl-1-naphthylamine, N-phenyl-2-naphthylamine); Fatty acids (linoleic acid) | Microcystis aeruginosa, Microcystis sp., green microalgae | [87,88] |
| Coontail (Ceratophyllum demersum L.) | Esters of carboxylic acids (3-hydroxy-2,2,4-trimethylpentyl ester of 2-methylpropanoic acid); Ethers of phthalic acid (dibutyl phthalate) | Aphanizomenon flosaquae Ralfs ex Bornet and Flahault, Microcystis aeruginosa, Pseudanabaena limnetica (Lemmermann) Komarek, Oscillatoria tenuis C. Agardh ex Gomont, green microalgae | [5,82,89] |
| Water lettuce (Pistia stratiotes L.) | Phenylpropanoids (α-asarone); Fatty acids (linoleic acid, γ-linolenic acid) | Microcystis aeruginosa, Synechococcus leopoliensis (Raciborski) Komárek, phytoplankton in general | [90,91] |
| Sago pondweed (Potamogeton pectinatus L.) | Terpenoids (lactone diterpenes) | Microcystis aeruginosa, Oscillatoria tenuis, green microalgae | [5,79,92] |
| Floating pondweed (Potamogeton natans L.) | Terpenoids (lactone diterpenes, furan diterpenes) | Microcystis aeruginosa, green microalgae | [93] |
| Floating fern (Salvinia natans L.) | Ethers of phthalic acid (dibutyl phthalate) | Microcystis aeruginosa | [94] |
| Helophytes (partly submerged plants) | |||
| Common reed (Phragmites communis Trin. (=Phragmites australis (Cav.) Trin. ex Steud)) | Phenolic acids (p-coumaric acid, ferulic acid, vanillic acid, syringic acid, caffeic acid, gallic acid); Fatty acids (tetradecanoic acid, palmitic acid, nonanoic acid and stearic acid) | Microcystis aeruginosa, Chlorella pyrenoidosa H.Chick | [65,95] |
| Narrowleaf cattail (Typha angustifolia L.) | Phenolic acids and their derivatives (syringic acid, isoferulic acid) | Dolichospermum flosaquae, Microcystis aeruginosa, | [96,97] |
| Broadleaf cattail (Typha latifolia L.) | Fatty acids (linoleic acid, α-linolenic acid) | Dolichospermum flosaquae, Microcystis aeruginosa, Chlorella vulgaris Beijerinck, Chlorella pyrenoidosa | [79,98,99] |
Table 2.
The mechanism of inhibitory action of some allelopathically active substances of aquatic vascular plants on cyanobacteria.
Table 2.
The mechanism of inhibitory action of some allelopathically active substances of aquatic vascular plants on cyanobacteria.
| Allelopathically Active Compounds | Structural | Mode of Action |
|---|---|---|
| Polyphenols | ||
| α-Asarone |  | Inhibition of respiration, inhibition of growth [79] |
| Gallic acid |  | Inhibition of photosystem II (PSII), reduction of chlorophyll content, oxidative stress [12,58,103] |
| Pyrogallic acid |  | |
| Ferulic acid |  | Reduction of membrane potential, reduction of chlorophyll content [58] |
| Telimagrandin II |  | Inhibition of photosystem II (PSII); inhibition of alkaline phosphatase activity [79,103] |
| Fatty acids/esters | ||
| Nonanoic acid |  | Inhibition of oxygen evolution, inhibition of growth [103,104] |
| Linoleic acid |  | Oxidative stress, reduction of chlorophyll content, blocking the transport of electrons [58,105] |
| α-Linolenic acid |  | Inhibition of growth [58] |
| Ethyl 2-methylacetoacetate |  | Oxidative stress, changes in the structure of cell membranes, inhibition of growth [12,16,106] |
| Nitrogen-containing compounds | ||
| N-phenyl-1-naphthylamine |  | Oxidative stress, inhibition of growth [107] |
| Terpenoids | ||
| β-Ionone |  | Distortion of thylakoids [108] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nezbrytska, I.; Usenko, O.; Konovets, I.; Leontieva, T.; Abramiuk, I.; Goncharova, M.; Bilous, O. Potential Use of Aquatic Vascular Plants to Control Cyanobacterial Blooms: A Review. Water 2022, 14, 1727. https://doi.org/10.3390/w14111727
AMA Style
Nezbrytska I, Usenko O, Konovets I, Leontieva T, Abramiuk I, Goncharova M, Bilous O. Potential Use of Aquatic Vascular Plants to Control Cyanobacterial Blooms: A Review. Water. 2022; 14(11):1727. https://doi.org/10.3390/w14111727
Chicago/Turabian StyleNezbrytska, Inna, Oleg Usenko, Igor Konovets, Tetiana Leontieva, Igor Abramiuk, Mariia Goncharova, and Olena Bilous. 2022. "Potential Use of Aquatic Vascular Plants to Control Cyanobacterial Blooms: A Review" Water 14, no. 11: 1727. https://doi.org/10.3390/w14111727
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.