
Moroccan Lagoon Microbiomes
by
 , , and add
Show full author list
, , and add
Show full author list
Bouchra Chaouni
1,2,3, 
Abdellah Idrissi Azami
3, 
Soumaya Essayeh
4,
El Houcine Arrafiqui
4,
Abdelhakim Bailal
4,
Sanae Raoui
5,
Saaïd Amzazi
5,
Alan Twaddle
6,
Chahrazade El Hamouti
7,
Noureddine Boukhatem
7,
Mohammed Timinouni
8,
Fatima El Otmani
9,
Rajaa Chahboune
10,
Said Barrijal
11,
Abdellatif El Homani
12,
Chakib Nejjari
3,
El Houssine Zaid
1,
Noureddine Hamamouch
1,
Fadil Bakkali
3,
Linda Amaral-Zettler
2,13,† and
and
Hassan Ghazal
3,14,*add
Show full author list
1
Laboratory of Plant and Microbial Biotechnology, Biodiversity and Environment, Faculty of Sciences, Mohammed V University in Rabat, Rabat 10000, Morocco
2
Josephine Bay Paul Center for Comparative Molecular Biology and Evolution, Marine Biological Laboratory, Woods Hole, MA 02543, USA
3
Laboratory of Genomics and Bioinformatics, Mohammed VI University of Health Sciences, Casablanca 82403, Morocco
4
Polydisciplinary Faculty of Nador, Mohammed Premier University, Nador 62702, Morocco
5
Laboratory of Human Pathologies Biology, Faculty of Sciences, Mohammed V University in Rabat, Rabat 10000, Morocco
6
Department of Biology, Center for Genomics and Systems Biology, New York University, New York, NY 10012, USA
7
Laboratory of Bioresources, Biotechnology, Ethnopharmacology and Health, Faculty of Sciences, Mohammed First University, Oujda 60000, Morocco
8
Laboratory of Microbiology, Institut Pasteur du Maroc, Casablanca 20250, Morocco
9
Microbiology, Health and Environment Team Research, Faculty of Sciences, University Chouaib Doukali, El Jadida 24000, Morocco
10
Laboratory of Life and Health Sciences, Faculty of Medicine and Pharmacy of Tangier, Abdelmalek Essaâdi University, Tétouan 93002, Morocco
11
Faculty of Sciences and Technologies, Abdelmalek Essaâdi University, Tangier 93000, Morocco
12
School of Medicine, Avalon University, Youngstown, OH 44505, USA
13
Department of Ecology and Evolutionary Biology, Brown University, Providence, RI 02912, USA
14
National Center for Scientific and Technical Research (CNRST), Angle Avenues des FAR et Allal El Fassi, Hay Ryad, Rabat 10102, Morocco
*
Author to whom correspondence should be addressed.
†
Current address: Department of Marine Microbiology and Biogeochemistry, NIOZ Royal Netherlands Institute for Sea Research, 1790 AB Den Burg, The Netherlands.
Water 2022, 14(11), 1715; https://doi.org/10.3390/w14111715
Submission received: 28 March 2022
/
Revised: 29 April 2022
/
Accepted: 3 May 2022
/
Published: 27 May 2022
(This article belongs to the Section Biodiversity and Functionality of Aquatic Ecosystems)
Abstract
:Lagoons are fragile marine ecosystems that are considerably affected by anthropogenic pollutants. We performed a spatiotemporal characterization of the microbiome of two Moroccan lagoons, Marchica and Oualidia, both classified as Ramsar sites, the former on the Mediterranean coast and the latter on the Atlantic coast. We investigated their microbial diversity and abundance using 16S rRNA amplicon- and shotgun-based metagenomics approaches during the summers of 2014 and 2015. The bacterial microbiome was composed primarily of Proteobacteria (25–53%, 29–29%), Cyanobacteria (34–12%, 11–0.53%), Bacteroidetes (24–16%, 23–43%), Actinobacteria (7–11%, 13–7%), and Verrucomicrobia (4–1%, 15–14%) in Marchica and Oualidia in 2014 and 2015, respectively. Interestingly, 48 strains were newly reported in lagoon ecosystems, while eight unknown viruses were detected in Mediterranean Marchica only. Statistical analysis showed higher microbial diversity in the Atlantic lagoon than in the Mediterranean lagoon and a robust relationship between alpha diversity and geographic sampling locations. This first-ever metagenomics study on Moroccan aquatic ecosystems enriched the national catalog of marine microorganisms. They will be investigated as candidates for bioindication properties, biomonitoring potential, biotechnology valorization, biodiversity protection, and lagoon health assessment.
1. Introduction
In Morocco, an important population density inhabits coastal lagoons, which makes these lagoons vulnerable to profound physical, ecological, and associated societal disturbance. They are highly susceptible to anthropogenic influences, are usually impacted by mining, tourism, agriculture, livestock, and residential sewage, and have limited water exchange with seas. These factors influence microbial diversity and community structure, which is expected to differ from the adjacent marine environment. Marine ecosystems have been the target of numerous metagenomics studies to explore microbial diversity and develop biotechnological applications. The metagenomics approach used to study the microbiome gave an insight into the different microorganisms inhabiting this environment, ranging from Cyanobacteria, a major player in global carbon cycling [1], to Pelagibacterales, classified as the most abundant microbes in the world’s oceans. It helps to identify microbial community diversity and evolution patterns and examine specific microbial group vertical distributions (ICoMM, [2], Tara oceans [3]). Studies show that the microbial diversity, proportion, and distribution in the marine environment are affected by different factors, such as temperature [4], organic matter [5], pH [6], depth, and salinity [7]. The latter is the most important factor shaping microbial diversity and structure [8]. High-throughput sequencing (HTS) has helped assess lagoon microbial diversity through metagenomics [9]. However, the marine microbiome and shaping environmental factors are still understudied worldwide, including in Morocco. We chose two Moroccan coastal lagoon ecosystems for their importance, namely, Marchica and Oualidia, to describe their extant microbiome using metagenomics. Both lagoons are classified as Ecological and Biological Sites of Interest (SBEI) and Ramsar sites (Convention on Wetlands of International Importance). Oualidia is the site of intensive aquaculture production of the Pacific oyster and a migratory stopover for birds between Europe and Africa, whereas Marchica is the largest, most attractive lagoon of the south shore of the Mediterranean Sea in Morocco and a model of sustainable development. Both lagoons have suffered profound anthropogenic pressures from economic and agricultural actions in adjacent zones. Ostracods are the most famous bioindicators of pollution in Marchica [10]. Most conducted studies have only discussed phytoplankton composition. Phytoplankton blooms were contributed to by Chaetoceros Ehrenberg, Pseudo-nitzschia Peragallo, and Nitzschia longissima (Brébisson) Ralfs. Their abundance and seasonality characterized Marchica as a highly eutrophicated lagoon [11]. The waters of the Oualidia Lagoon were dominated by diatoms, ranging from 70% to 98% of the algal community. However, Dinoflagellate’s relative abundance increased after small diatom cells diminished [12]. Lagoon–ocean exchanges and ecological factors directly influenced the richness and diversity of planktonic algae. In addition, the distribution of different species in the lagoon was directly impacted by the hydrodynamics of the lagoon and its shallow water conditions [12]. Moreover, in Oualidia, harmful species such as Prorocentrum Ehrenberg, Dinophysis Ehrenberg, and Alexandrium Halim were detected at low concentrations and could cause toxicity accumulation in oysters [12].
As part of the Ocean Sampling Day project (OSD) to sample the world’s marine environment [13], we followed both 16S-amplicon and WGS metagenomic sequencing approaches to characterize the microbial diversity of the Marchica and Oualidia Moroccan lagoons and compared them with neighboring open sea marine environments. The sampling in Marchica and Oualidia was performed on 21 June 2014 and 2015; the boreal summer solstice. This is the first time such a metagenomics approach has been applied in Morocco to study a marine ecosystem and initiate Morocco’s marine microbial inventory. We hypothesize that the operational taxon unit (OTU) distribution differs between the 2014 and 2015 sampling campaigns; it is influenced by geography, and taxa diversity correlates with environmental parameters.
2. Materials and Methods
2.1. Water Sampling
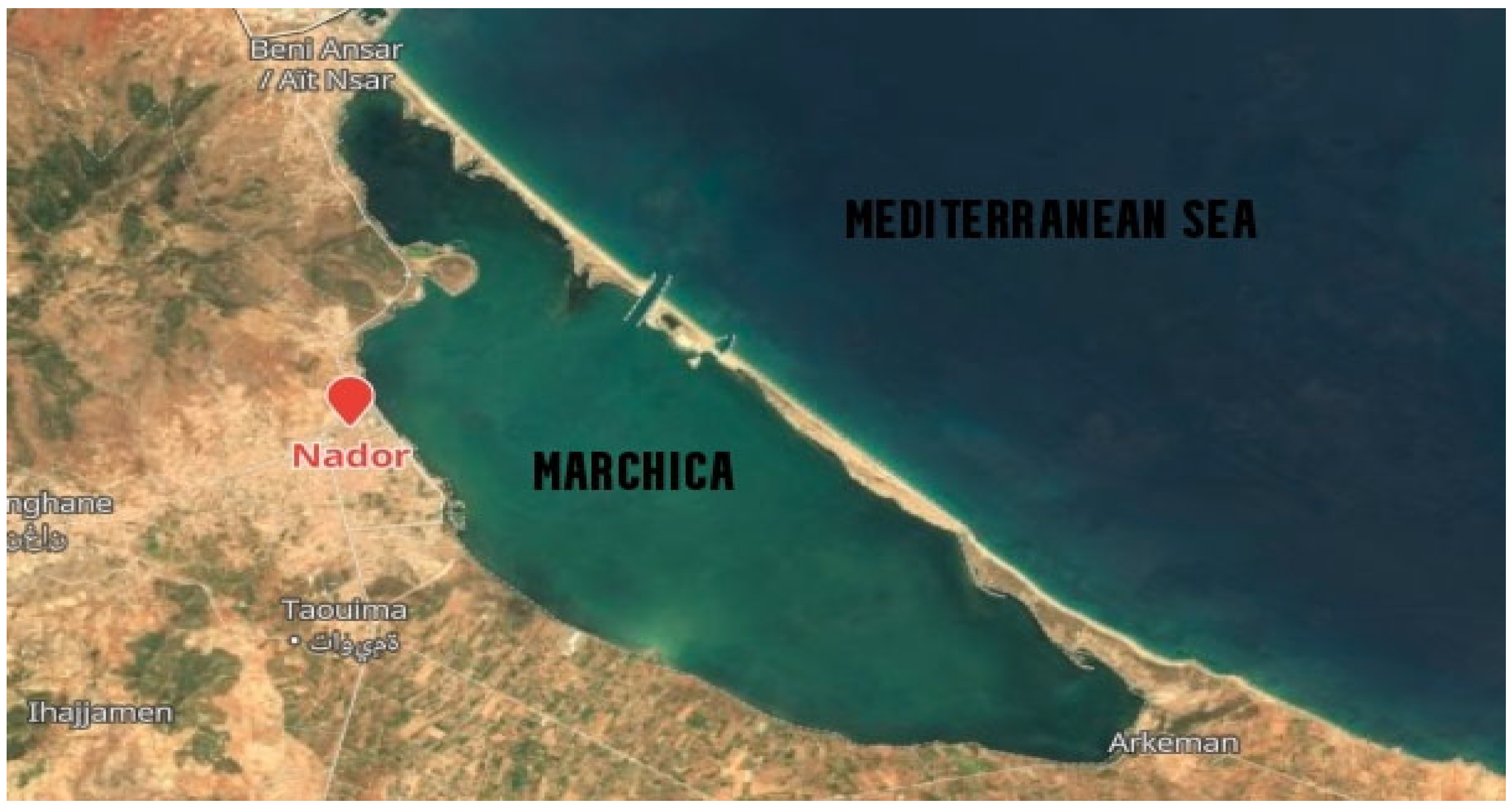
Samples were collected from Marchica (N 35.11562, W 2.52803) and Oualidia (N 32.74675, W 9.036667) on 21 June 2014 and 2015, the boreal summer solstice (Figure 1 and Figure 2). Approximately 20 L was collected using a 10% acid-washed bucket, then sequentially filtered on five Sterivex filter units with 0.22 μm pore size using a hand pump and stored at −80 °C. Metadata such as sample volume, depth, salinity, temperature, pH, latitude/longitude, dissolved oxygen (DO), turbidity, and conductivity (EC) were measured in situ using multivariate calibrated probes. Nitrate, nitrite (NO3−, NO2−), and phosphate concentrations were determined using the molecular UV/Visible spectrophotometric method. The biochemical oxygen demand (BOD) for each sample was recorded for five days at 20 °C using an incubation flask (OxiTop ® Control measuring system, Munich, Germany). We measured chemical oxygen demand (COD) by applying a robust oxidizing reagent (potassium dichromate K2Cr2O7) under acidic conditions (Table 1).
2.2. 16S/18S Amplicon Sequencing
DNA extraction was performed using the isolation kit Power Water (MoBio, USA) as instructed by the manufacturer. Amplification of the 16S rRNA gene was performed using the primer pair 515F-Y (5′-GTGYCAGCMGCCGCGGTAA-3′) and 926R (5′-CCGYCAATTYMTTTRAGTTT-3′) [14]. The forward primer TAReuk454FWD1 (5′-CCAGCASCYGCGGTAATTCC-3′) and the reverse primer TAReukREV3_modified (5′-ACTTTCGTTCTTGATYRATGA-3′) were used for 18S rRNA gene amplification. Ovation Rapid DR Multiplex System 1–96 (NuGEN) was used to construct Illumina libraries, pooled and size selected by preparative gel electrophoresis. The detailed protocol can be found under this link (https://github.com/MicroB3-IS/osd-analysis/wiki/Guide-to-OSD-2014-data accessed on 1 December 2020). Sequencing was performed on an Illumina MiSeq using V3 Chemistry. Samples were sequenced in one MiSeq run (2 × 300 bp), generating 2 × 40,000 reads per sample.
2.3. Preparation of Shotgun Libraries for Metagenomic Samples and Sequencing
One half of the supplied DNA material per sample was sheared to approximately 500 bp fragments using a Covaris S220 sonicator. DNA was purified and concentrated by clean-up using MinElute columns (Qiagen). DNA concentrations were measured, and 100 ng (or less, if the sample contained insufficient amounts, successful libraries could be obtained from as little as 5 ng) was used to prepare Illumina libraries. Libraries were generated with the Ovation Rapid DR multiplex 1–96 system (NuGEN). Libraries were amplified using standard Illumina primers for 8 to 15 cycles with MyTaq (Bioline). Eighteen cycles were necessary to generate the library. Sequencing was performed on an Illumina MiSeq using V3 Chemistry. Samples were sequenced in eight MiSeq runs (2 × 300 bp) generating 2 × 1,000,000 reads per sample.
2.4. Data Processing
Taxonomic analysis: Sequence data were preprocessed following the OSD workflow (github.com/MicroB3-IS/osd-analysis/wiki/Sequence-Data-Preprocessing accessed on 1 December 2020), which generated “workable” metagenome and amplicon fasta files. Raw files were stored at EBI (www.ebi.ac.uk/ena/data/view/ERX947554 accessed on 1 December 2020). Table 2 summarizes number of raw 16S rRNA sequences collected from sterivex filters from both lagoons.
Amplicon sequences were processed with the VAMPS web service [15], where taxonomy classification for 16S and 18S rRNA gene sequences was assigned in a Global Alignment for Sequence Taxonomy (GAST) proceeding [16] and SILVA rRNA gene reference database [17]. To crosscheck the identified genera, we used the DADA2 microbiome pipeline [18]. Filtering parameters used the following settings: 200 bp for the forward (R1) and 190 bp for the reverse (R2) reads. All identical sequencing reads were combined into unique sequences, denoised, merged if the forward and reverse reads overlapped by at least 12 bases, and set as chimera-free. Similar sequences were grouped into distinct OTUs, and taxonomy was assigned to sequence variants using the SILVA reference database. Table 3 summarizes the number of reads that made it through each step in the DADA2 pipeline.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 2.
Number of raw DNA sequence reads (R1 Forward and R2 Reverse) obtained from each Sterivex filter for both amplicon from Marchica and Oualidia water lagoons.
Table 2.
Number of raw DNA sequence reads (R1 Forward and R2 Reverse) obtained from each Sterivex filter for both amplicon from Marchica and Oualidia water lagoons.
| Filter Size | # Reads | Total Length (Mb) | Average Read Length (bp) | ||
|---|---|---|---|---|---|
| Marchica | 2014 | 0.22 μm | 132,322 | 80.1 | 269 |
| 2015 | 0.22 μm | 76,482 | 46.6 | 271 | |
| Oualidia | 2014 | 0.22 μm | 179,448 | 108.2 | 268 |
| 2015 | 0.22 μm | 146,886 | 89.2 | 270 | |
| Total | 535,138 | 324.1 | |||
Table 3.
Number of DNA sequence reads that made it through each step in the DADA2 tool pipeline.
| Samples | Input | Filtered | DenoisedF | DenoisedR | Merged | Nonchim |
|---|---|---|---|---|---|---|
| OSD2414 (Marchica) | 66,161 | 28,157 | 27,383 | 27,412 | 23,687 | 19,662 |
| OSD2415 (Marchica) | 38,241 | 18,570 | 18,124 | 18,198 | 16,957 | 16,355 |
| OSD4714 (Venice) | 44,664 | 21,137 | 20,199 | 20,354 | 17,763 | 15,331 |
| OSD4715 (Venice) | 60,996 | 31,535 | 30,156 | 30,205 | 26,166 | 23,088 |
| OSD8114 (Ria Formosa) | 53,394 | 22,538 | 22,059 | 21,935 | 18,895 | 16,205 |
| OSD8115 (Ria Formosa) | 90,023 | 45,355 | 44,652 | 44,584 | 41,038 | 37,323 |
| OSD9114 (Oualidia) | 89,724 | 43,757 | 41,329 | 41,707 | 34,329 | 29,064 |
| OSD9115 (Oualidia) | 73,443 | 33,878 | 32,946 | 32,968 | 28,967 | 23,401 |
| OSD9314 (ElJadida) | 41,323 | 20,930 | 19,889 | 20,052 | 16,612 | 13,680 |
| OSD9315 (ElJadida) | 62,923 | 31,790 | 31,088 | 31,164 | 27,840 | 25,242 |
| OSD9414 (Saidia Marina) | 61,932 | 28,805 | 27,557 | 27,544 | 22,009 | 18,190 |
| OSD9415 (Saidia Marina) | 48,501 | 20,674 | 20,169 | 20,156 | 18,152 | 16,940 |
Metagenome analysis: We used MetaSpades [19] to assemble trimmed forward (R1) and reverse (R2) reads and then MetaQuast [20] to control assembly quality. A total of 196,153 contigs were produced for Marchica, labeled (OSD24, https://www.ebi.ac.uk/ena/browser/view/ERS667644 accessed on 1 December 2020), and 110,719 for Oualidia, labeled (OSD91, https://www.ebi.ac.uk/ena/browser/view/ERS667576 accessed on 1 December 2020). A total of 54,965 reads in OSD24 passed the Quality Control (QC), while only 19,536 reads in OSD91 did. We used Prodigal [21] for the microbial genus, species, and strain level detection for sequence annotation, while we used MetaPhlAn2 [22] to obtain the relative abundance. We used MG-RAST [23] for protein-coding gene prediction for the whole microbiome (Figure 3). Finally, we used Prokka [24] to annotate the sequences that belong to the most abundant species of the five phyla Proteobacteria, Bacteroides, Cyanobacteria, Verrucomicrobia, and Actinobacteria.
Viral signal detection: Reads were mapped to a viral database, including 1575 complete viral genomes from RefSeq and 6322 contigs from the TOV_43 dataset [25]. Read recruitment values were calculated as part of the bowtie2 [26] output. The number of reads and aligned nucleotides were computed using a custom Python script employing Biopython. Coverage values for all sequences were calculated using a Python script modified to normalize for metagenome sizes. All other calculations were performed using the Python modules pandas and numpy, and images were generated using a combination of Seaborn, matplotlib, and palettable python modules. The virus-sample heatmap was produced with the package pheatmap for drawing heatmaps in R using default hierarchical clustering for the viral sequences (rows) and a manual clustering of samples by geographical zone (columns). Geo-coordinates were confirmed using OSD GitHub (https://github.com/MicroB3-IS/osd-analysis/wiki/Guide-to-OSD-2014-data accessed on 30 October 2020).
Statistical analyses: Graphics and ecological statistical analyses for the relative abundance of specific phyla were performed using the R Statistical package R v. 3.0.2 (www.R-project.org/ accessed on 15 November 2020). Visualization and analysis of inter- and intracommunity comparisons comprise the realization of community heatmaps, skyline plots, pie charts, dendrograms, refraction, and diversity indexes using R packages (phyloseq, ggplot2, ape, pheatmap, vegan, jsonlite, Rcolorbrewer) and python3 modules scipy, numpy, and cogent.
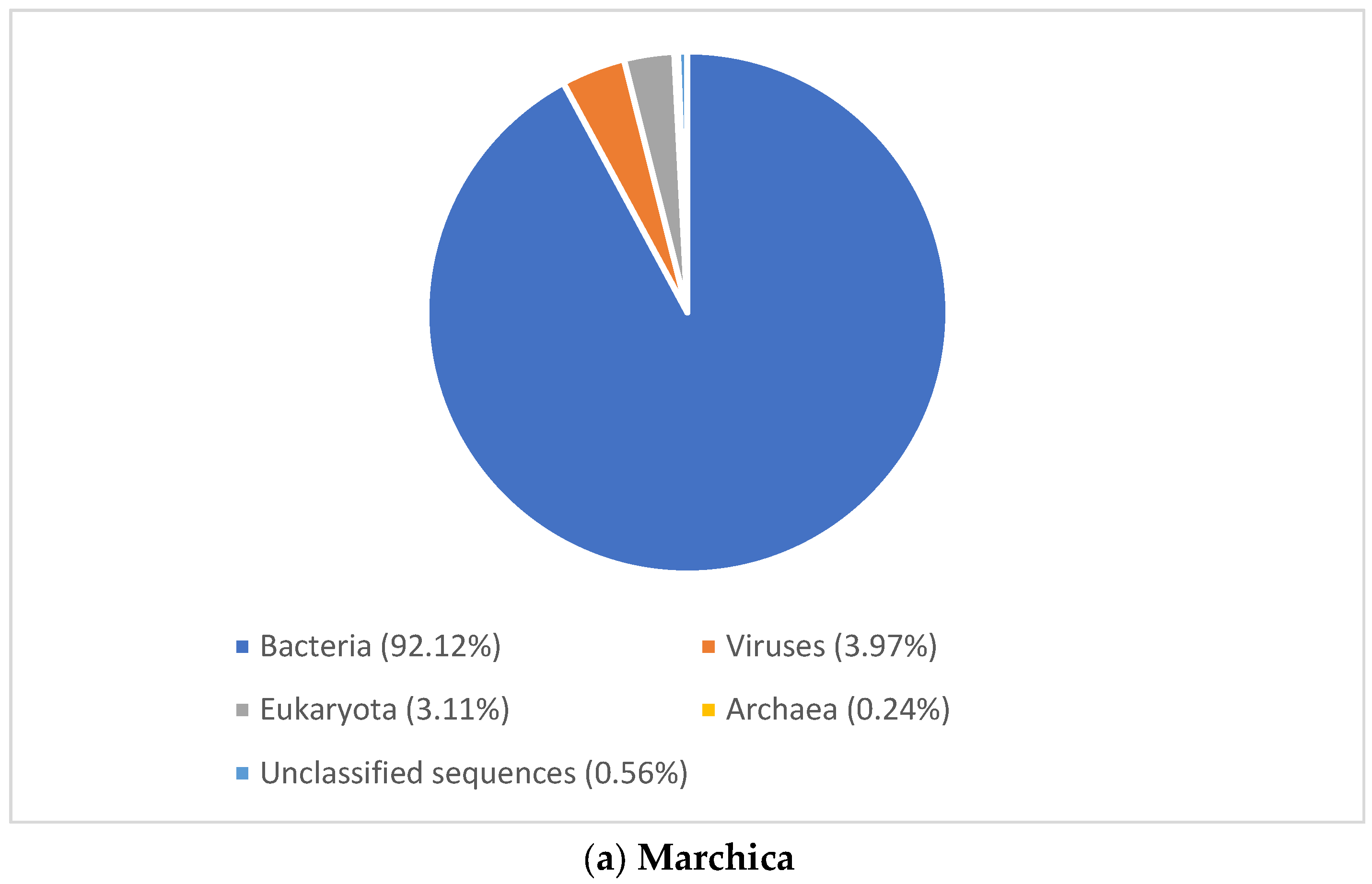
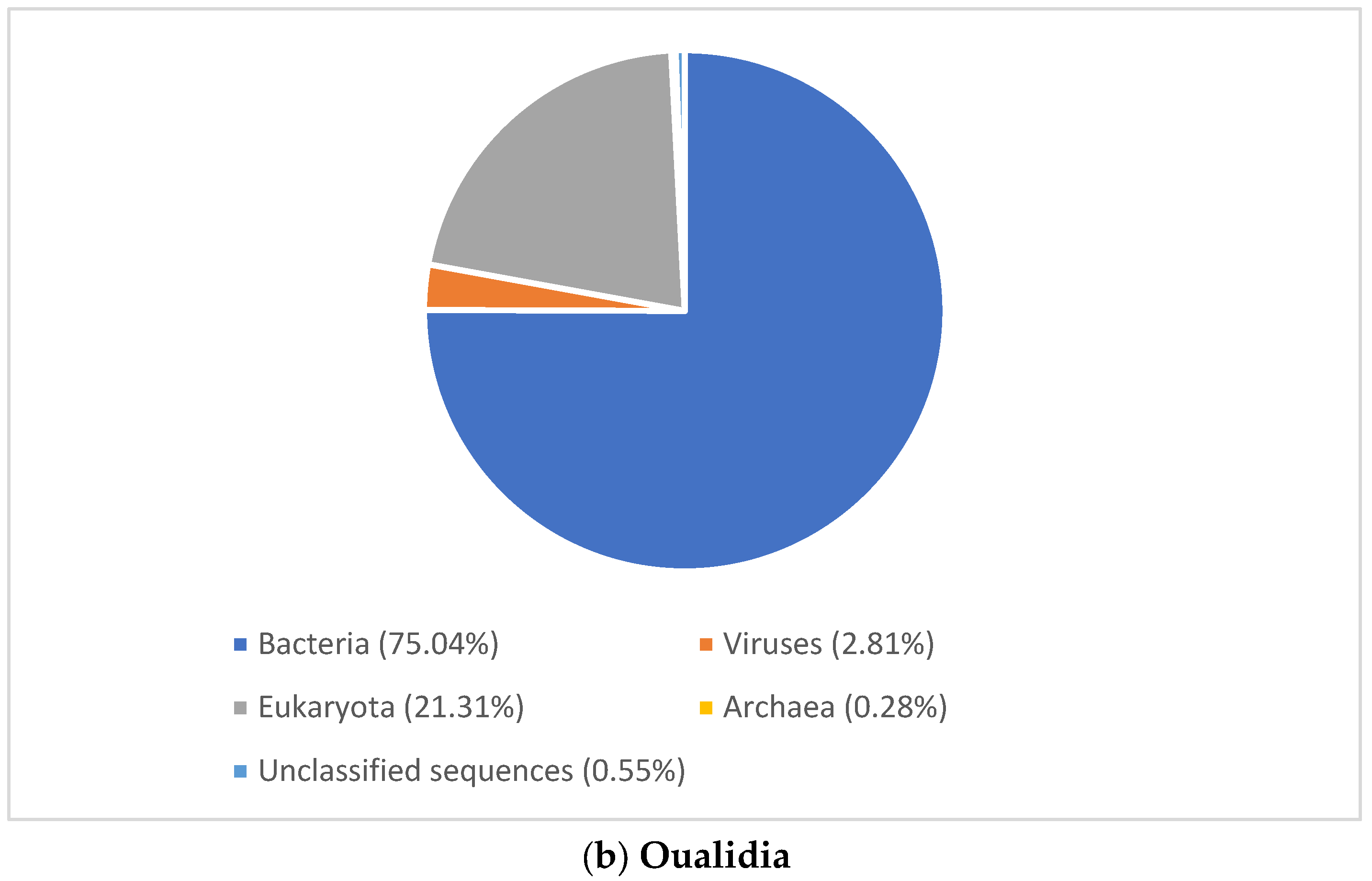
Figure 3.
Domain level classification of all metagenomic reads using MG-RAST [23]. (a) Marchica lagoon (b) Oualidia lagoon.
Figure 3.
Domain level classification of all metagenomic reads using MG-RAST [23]. (a) Marchica lagoon (b) Oualidia lagoon.


3. Results
3.1. Physicochemical Characteristics of the Lagoon Sampling Sites
The measurements of physicochemical parameters are shown in Table 1. The water temperature of the Marchica (~27 °C) and Oualidia (~20.5 °C) lagoons varied significantly with the sampling locations. The electrical conductivity (EC) shows a clear evolution, with a maximum in the summer of 2015 in Oualidia (45.1 ms/cm) and a minimum in 2014 (39.6 ms/cm). In Marchica, it varied between 53 ms/cm and 54 ms/mc, which was similarly observed in 2015 by [27]. Salinity, as represented by EC, was much higher in Marchica (36–40 ppt) than in the Mediterranean Sea (24–32 ppt), whereas Oualidia waters showed a lower salinity value (27–29 ppt) regardless of their connection to the Atlantic where salinity can reach above 36 ppt. Waters of Marchica and Oualidia were mildly alkaline (pH~8.3), which is the optimum for marine bacterial growth [28]. The recorded DO concentrations in Marchica (8.5 mg/L) and Oualidia (7.5 mg/L) indicated that surface waters were tolerably oxygenated (6.15 to 9.02 mg/L) [27]. In addition, both lagoons were slightly turbid (3 Nephelometric Turbidity Units (NTU)). In Marchica, nitrate concentrations reached 12.5 mg/L in 2014 and dropped to 4.8 mg/L in 2015, while in Oualidia, they followed the same pattern, varying between 10.79 mg/L in 2014 and 2.68 mg/L in 2015. The concentrations of phosphate in the surface waters of Marchica and Oualidia lagoons were 0.03–0.04 mg/L and 0–0 mg/L, respectively, in 2014 and 2015.
3.2. Microbial Community Structure
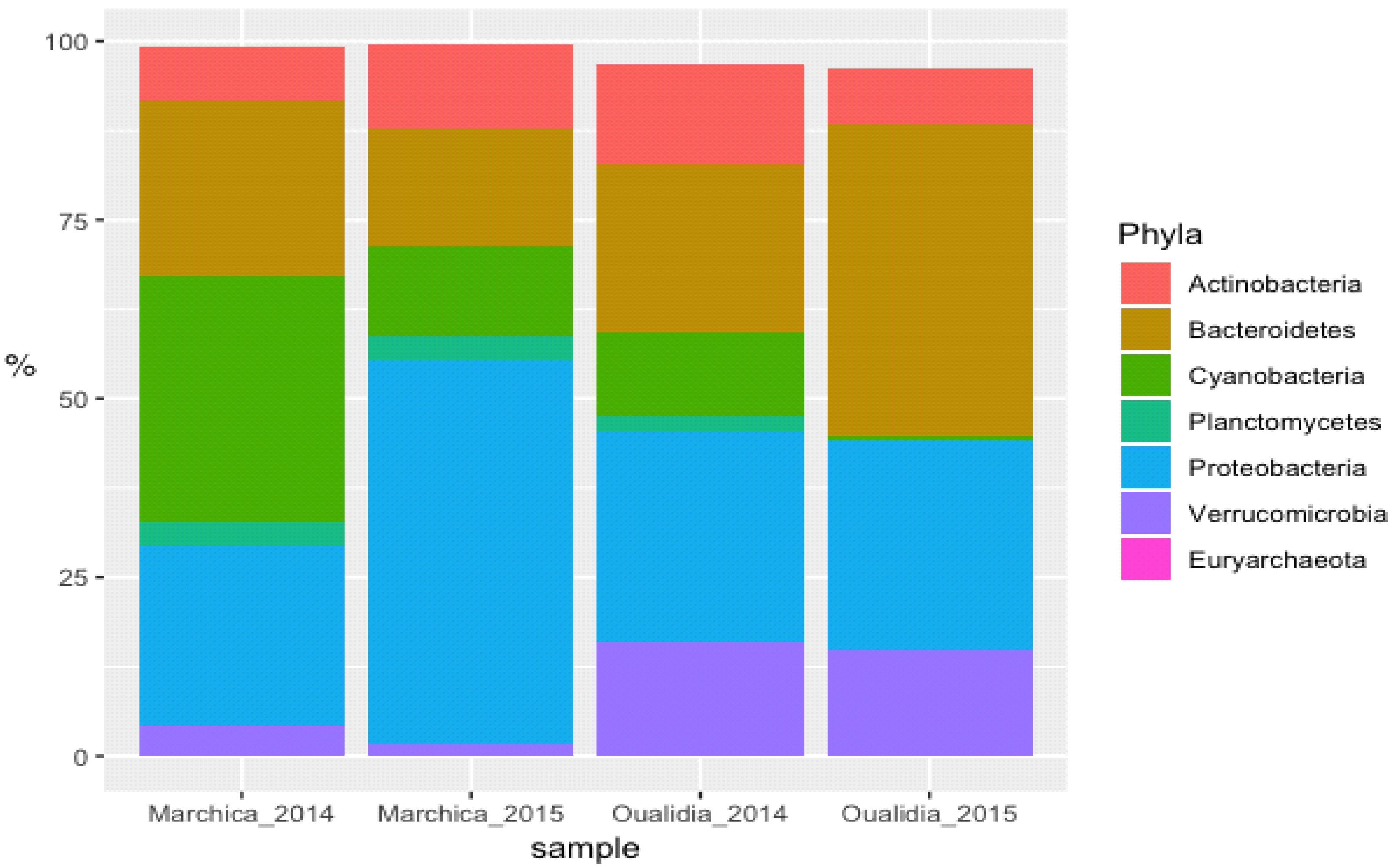
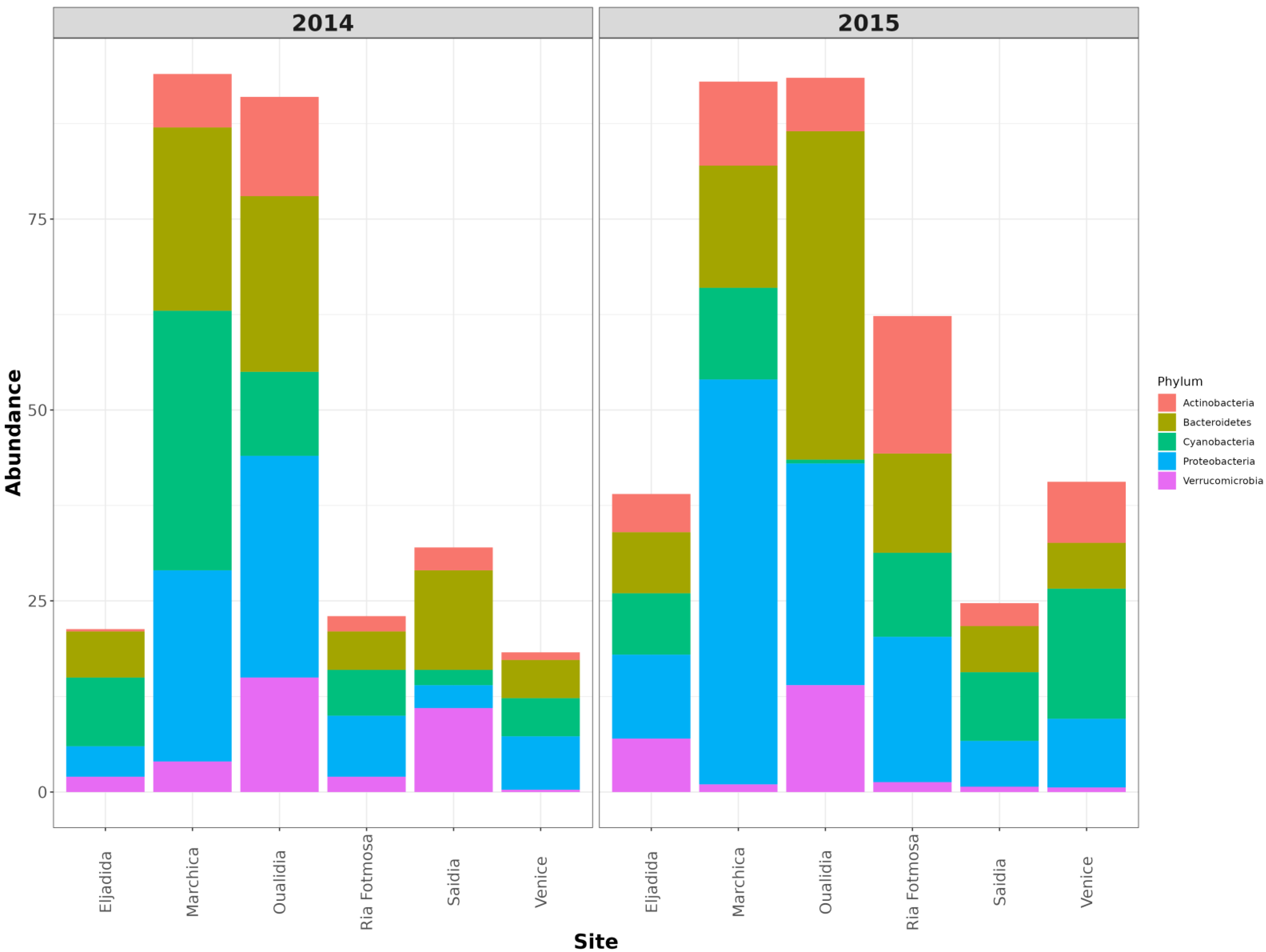
To gain an insight into microbial diversity in Moroccan lagoons, we used bioinformatics tools to analyze the metagenome and the amplicon sequence data of the Marchica and Oualidia sites produced from water samples after the sampling campaigns of the 21st of June 2014 and 2015 (Figure 4 and Figure 5, Table 4). 16S rRNA gene classification using the Silva reference database revealed a high percentage of bacteria in both lagoons. On average, bacteria accounted for 90% of the total prokaryotes in Marchica and ~70% in Oualidia, outnumbering Archea. We only found <1% archaeal gene sequences in both sites assigned to Euryarchaeotes. In total, ten phyla were identified using MG-RAST, not including the unknown class, while VAMPS and DADA2 identified 27 phyla, which formed 62 classes, 129 orders, 260 families, 799 genera, and 1379 candidate species for the 2014 and 2015 sampling in Marchica and Oualidia lagoons. Based on VAMPS and DADA2, the five phyla that were the most abundant in both lagoons, Marchica and Oualidia, respectively, were Proteobacteria (53.62%, 29.18%), Bacteroidetes (16.46%, 43.49%), Cyanobacteria (0.53%, 34.35%), Verrucomicrobia (1.75%, 15.82%), and Actinobacteria (7.42%, 13.98%).
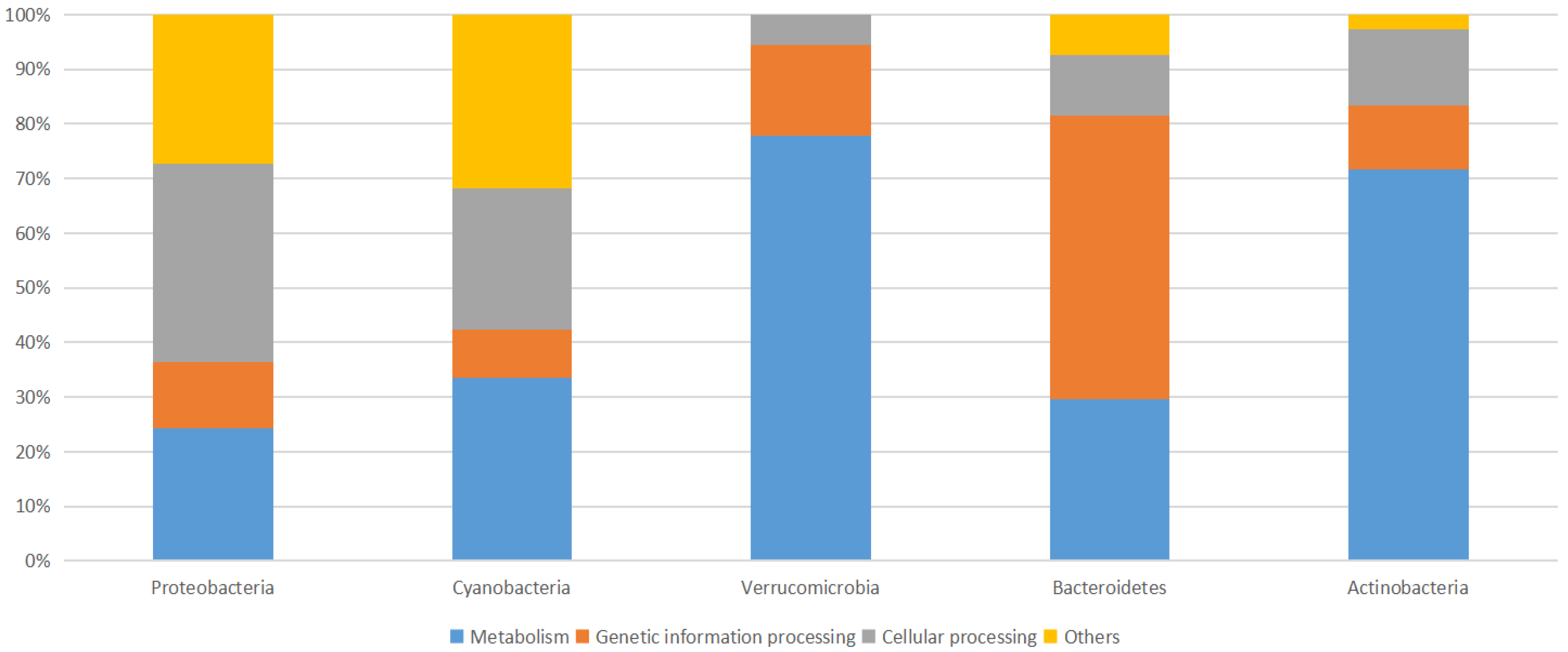
Figure 4.
Cross comparison of 16S rRNA amplicon sequences from selected abundant high-level bacterial taxa from Marchica and Oualidia metagenomes.
Figure 4.
Cross comparison of 16S rRNA amplicon sequences from selected abundant high-level bacterial taxa from Marchica and Oualidia metagenomes.

Table 4.
16s rRNA amplicon sequences from selected high abundant sequences assigned to a phylum for both sampling campaigns 2014 and 2015 in both lagoons.
Table 4.
16s rRNA amplicon sequences from selected high abundant sequences assigned to a phylum for both sampling campaigns 2014 and 2015 in both lagoons.
| Marchica 2014 | Oualidia 2014 | ||||
| Taxa | #Hits | % | Taxa | #Hits | % |
| Actinobacteria | 4793 | 7.42 | Actinobacteria | 11,670 | 13.98 |
| Bacteroidetes | 15,896 | 24.62 | Bacteroidetes | 19,660 | 23.55 |
| Cyanobacteria | 22,178 | 34.35 | Cyanobacteria | 9721 | 11.65 |
| Planctomycetes | 2190 | 3.39 | Planctomycetes | 1904 | 2.28 |
| Proteobacteria | 16,310 | 25.26 | Proteobacteria | 24,559 | 29.42 |
| Verrucomicrobia | 2712 | 4.20 | Verrucomicrobia | 13,202 | 15.82 |
| Euryarchaeota | 3 | 0 | Euryarchaeota | 19 | 0.02 |
| Marchica 2015 | Oualidia 2015 | ||||
| Taxa | #Hits | % | Taxa | #Hits | % |
| Actinobacteria | 4408 | 11.75 | Actinobacteria | 5613 | 7.80 |
| Bacteroidetes | 6173 | 16.46 | Bacteroidetes | 31,312 | 43.49 |
| Cyanobacteria | 4751 | 12.67 | Cyanobacteria | 384 | 0.53 |
| Planctomycetes | 1217 | 3.25 | Planctomycetes | 142 | 0.20 |
| Proteobacteria | 20,109 | 53.62 | Proteobacteria | 21,011 | 29.18 |
| Verrucomicrobia | 656 | 1.75 | Verrucomicrobia | 10,740 | 14.92 |
| Euryarchaeota | 10 | 0.03 | Euryarchaeota | 1 | 0 |
Following OTU clustering, we calculated the Shannon and Simpson diversity indexes from 88,482 bacterial sequences to determine species relative abundances at site and time. By definition, species richness is related to the count of the different species found in a sample. If the number of species present in “Sample A” is greater than that in “Sample B”, then “Sample A” is richer than “Sample B”. The diversity increases when species richness also increases [29]. The Shannon value likewise increases as the species number increases [30]. However, the Simpson index rises when the diversity drops [31]. On the other hand, both ACE and Chao1 were conceived to infer richness based on abundance data (importance of rare OTUs). Chao1 gives more weight to the low-abundance species, while ACE informs whether abundant species are present or absent [32]. Based on Vamps, calculating alpha diversity assessed by observed richness, Shannon and Simpson indexes, ACE, and Chao1 revealed higher bacterial diversity in Oualidia in comparison to Marchica in both summer 2014 and 2015 (Table 5).
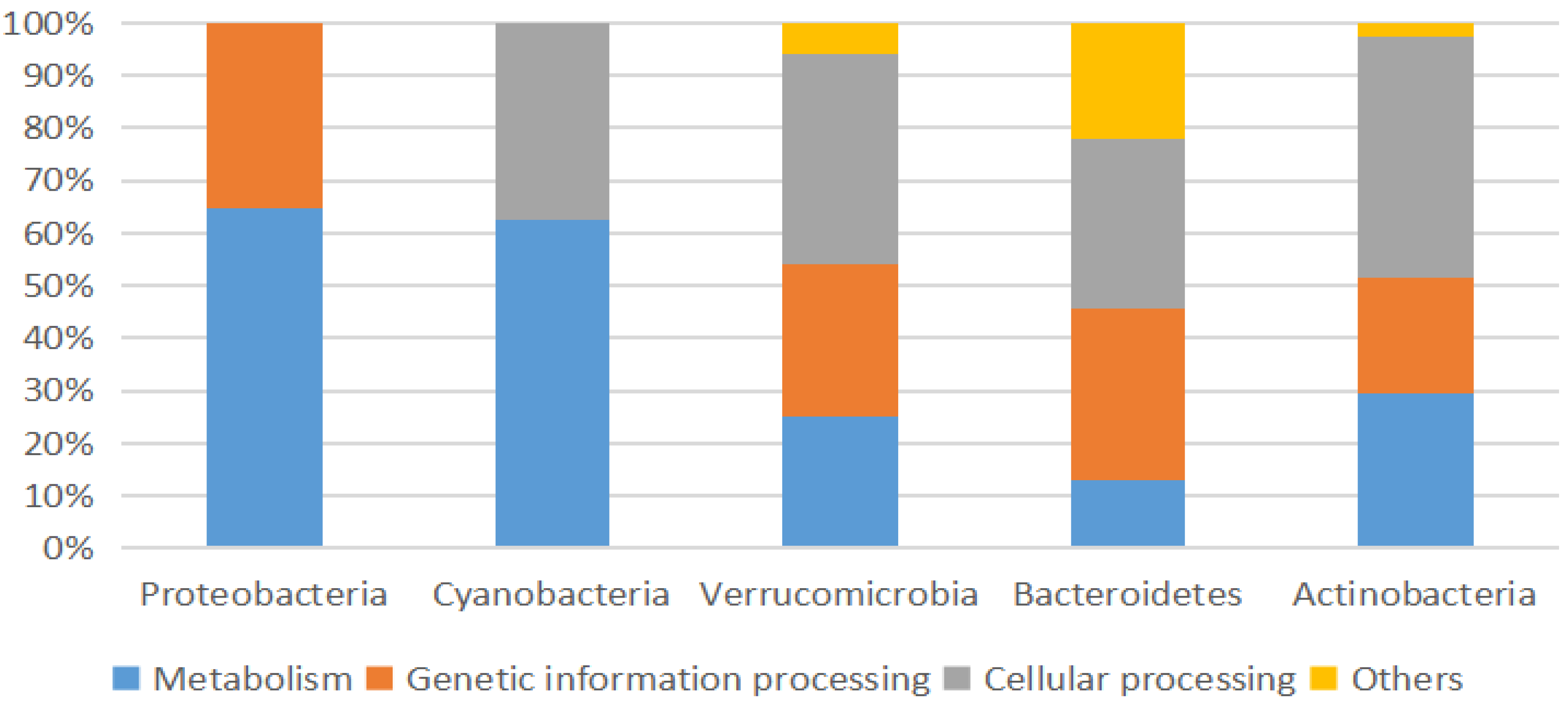
Figure 5.
Cross comparison of 16S rRNA amplicon sequences from selected 5 abundant high-level bacterial taxa from Marchica and Oualidia metagenomes to several saline metagenomes using R.
Figure 5.
Cross comparison of 16S rRNA amplicon sequences from selected 5 abundant high-level bacterial taxa from Marchica and Oualidia metagenomes to several saline metagenomes using R.

Table 5.
Microbial Alpha diversity indices; normalization: none; counts min/max: 0–100%.
| Dataset | Observed Richness | ACE | Chao1 | Shannon | Simpson |
|---|---|---|---|---|---|
| Marchica Bv4v5 (OSD24, 2014) | 462 | 677.66 | 656.16 | 4.18 | 0.86 |
| Marchica Bv4v5 (OSD24, 2015) | 316 | 490.39 | 497.5 | 4.13 | 0.87 |
| Oualidia Bv4v5 (OSD91, 2014) | 1144 | 1591.09 | 1605.32 | 6.13 | 0.94 |
| Oualidia Bv4v5 (OSD91, 2015) | 704 | 1053.14 | 1022.78 | 4.59 | 0.89 |
3.2.1. Viral Signature
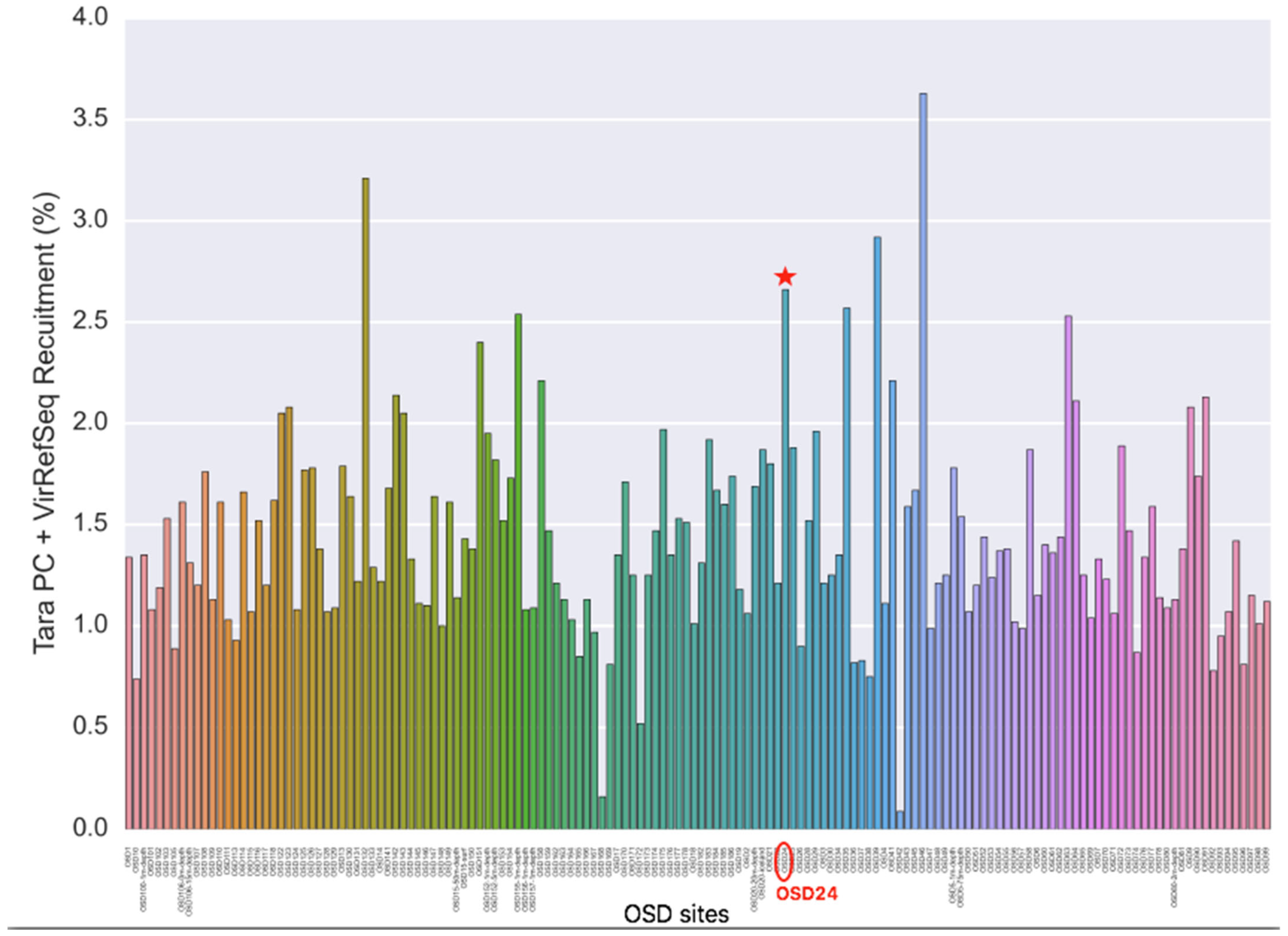
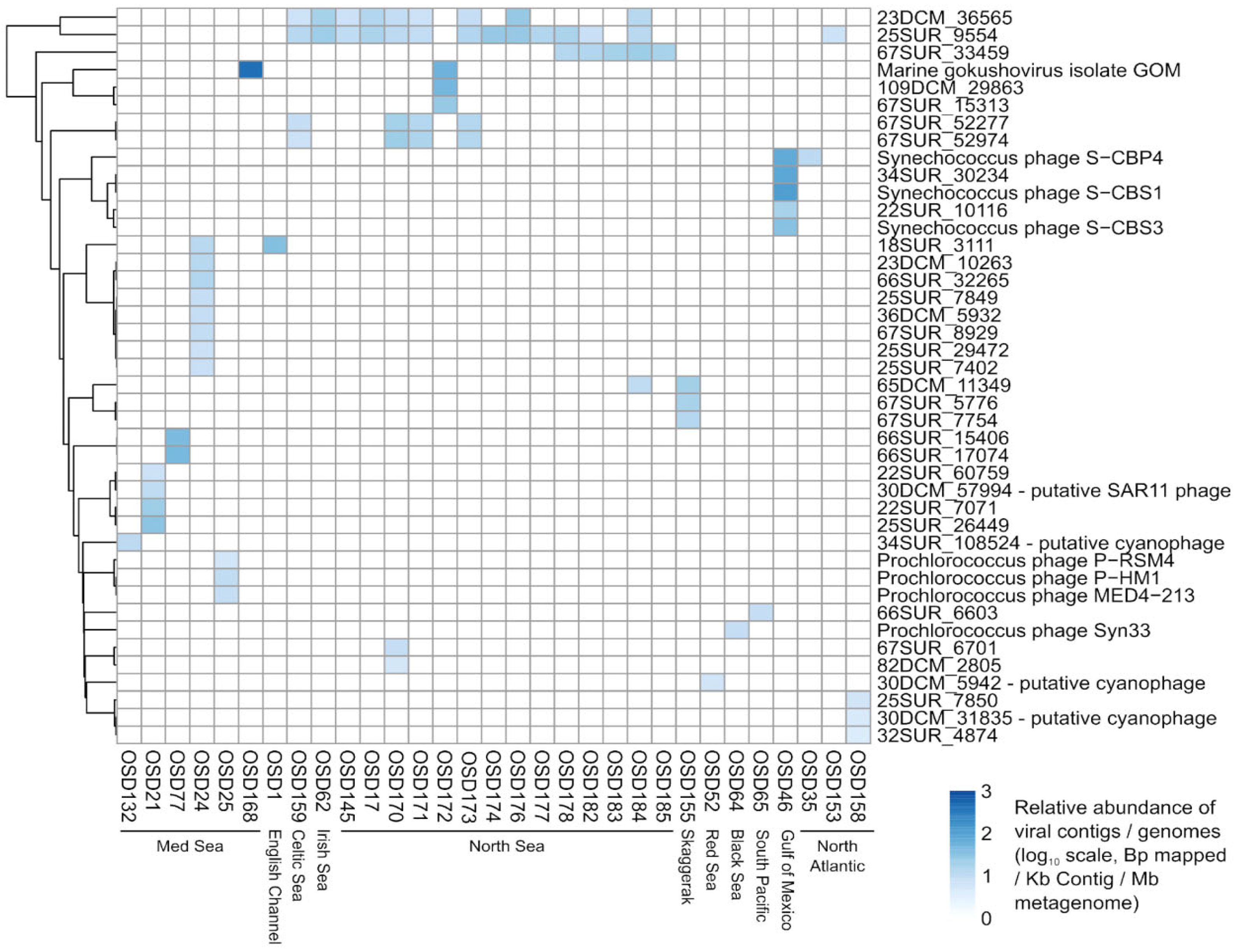
Looking at the OSD metagenomics dataset in 2014 as a whole revealed 42 viral genomes/contigs that were confidently identified (more than 75% of the genome/contig covered) across 31 OSD samples. Most of these viruses corresponded to known cyanophages from NCBI or “unknown Caudovirales” assembled from the Tara Oceans samples corresponding to coastal sites and/or the Mediterranean Sea. Here, we are interested in Moroccan lagoons. A coverage filter was applied, with a viral contig/genome being considered “detected” only if at least 75% of its bases were covered, which narrowed the number of viral sequences found. Reads in Marchica mapped at 2.7% against the combined ViralRefSeq and TOV_43 contigs (https://www.ivirus.us/data, accessed on 15 November 2020) (Figure 6). A “zoom-in” of these data revealed eight viruses in only Marchica Lagoon, none of which are known (Figure 7), suggesting the specificity of the Marchica virome.
3.2.2. Proteobacteria
This group is considered the largest and most diverse [33]. The functional attributes of Proteobacteria are linked to mechanisms such as denitrification and amino acid biosynthesis [34,35]. They formed the largest part of the bacterial group in both Marchica (25.2%, 2014; 53.6%, 2015) and Oualidia (29.4%, 2014; 29.1%, 2015) (Figure 4 and Figure 5, Table 4). α-Proteobacteria was the most predominant group at both sites, with a significant increase in 2015 in Marchica and a small increase in Oualidia (46% in Marchica; 21.59% in Oualidia) compared to 2014 (14.53% in Marchica; 18.5% in Oualidia). Marine Alphaproteobacteria are known to be more abundant in the epipelagic zone of coastal waters [36]. Rhodobacteraceae was the largest represented family in α-proteobacteria in Marchica (10.8%, 2014; 9.8%, 2015) and Oualidia (10.5%, 2014; 19.3%, 2015). It is dominant in different marine regions, namely, the Mediterranean, Atlantic, and Pacific Oceans [37], including areas low in nutrients but with a high rate of primary production [38]. Both α-Proteobacteria and Cyanobacteria, known for nitrate elimination, were cosmopolitan, which is consistent with former reports [39]. In Gammaproteobacteria, the relative abundance of Alteromonadaceae was higher in Marchica during 2014 (7.81%) than in 2015 (2.04%) but stable in Oualidia (2.31%, 2014; 2.44%, 2015). Shotgun metagenomic analysis of 2014 datasets showed the dominance of Candidatus Pelagibacter Rappé in Marchica (11%) and Oualidia (7%). This genus is known amid the broadly represented bacteria in open oceans [40]. In Mediterranean Saidia Marina (OSD94), the majority of alphaproteobacterial reads were ascribed to Candidatus Pelagibacter (10%), as well as in Venice Lagoon (OSD47) (7%). This was similarly observed in Mar Menor, a lagoon on the Mediterranean Spanish coast, where the dominance of Candidatus Pelagibacter was reported in 2012 (43%) [7]. In contrast, at the Atlantic site El Jadida (OSD93), we observed only a minority of reads attributed to Candidatus Pelagibacter (1%), while none were present in Ria Formosa Lagoon (OSD81), located south of Portugal on the Atlantic coast (ERS667579, https://www.ebi.ac.uk/ena/browser/view/ERS667579_, accessed on 1 December 2020). Furthermore, analysis of both lagoon metagenomes in 2014 showed remarkable differences at the strain level (Table 6). For instance, in the alphaproteobacteria group, Rickettsia conorii 7 Ogata—causative agent of Mediterranean spotted fever [41]—was the most dominant in both sites, but with a slightly higher proportion in Marchica (10.49%) than Oualidia (7.42%), followed by Orientia tsutsugamushi Boryong Cho (Marchica, 3.75%; Oualidia, 1.95%). A small proportion of reads were assigned to Anaplasma marginale Maries Brayton in Marchica (2.21%) compared to Oualidia (8.28%) and Anaplasma phagocytophilum HZ Hotopp (6% in Oualidia, 3% in Marchica), in contrast to the gammaproteobacteria strain Xylella fastidiosa Temecula1 Sluys, which was more abundant in Marchica (4.19%) compared to Oualidia (2.62%) (Table 6). Interestingly, we only observed the strain Anaplasma phagocytophilum HZ in our comparative Moroccan Mediterranean site Saidia Marina (ERS667573, https://www.ebi.ac.uk/ena/browser/view/ERS667573 accessed on 1 December 2020) (5%) and Atlantic site Eljadida (ERS667574, https://www.ebi.ac.uk/ena/browser/view/ERS667574 accessed on 1 December 2020) (5%) as well as in the Mediterranean lagoon Venice (ERS667621, https://www.ebi.ac.uk/ena/browser/view/ERS667621 accessed on 1 December 2020) (4%) and Atlantic Portuguese lagoon Ria Formosa (4%) (Table 6).
3.2.3. Cyanobacteria
Cyanobacteria are naturally occurring photosynthetic prokaryotes that can be found in almost all bodies, both fresh and saltwater [42]. These organisms are considered the dominant primary producers on Earth, contributing more than 25% of photosynthesis worldwide [43]. Cyanobacteria can grow into large colonies visible to the naked eye, often referred to as a Cyanobacteria harmful algal bloom (HAB) [44]. The most striking characteristic of the Marchica Lagoon was a large number of Cyanobacterial sequences (comprising nearly 34.5% of all 16S rRNA sequences in 2014, (see Figure 4 and Table 4) compared to Oualidia Lagoon (11.65%). This frequency significantly decreased in 2015 (12.6% in Marchica, 0.53% in Oualidia). The classification of the 16S rRNA metagenomic sequences indicated the presence almost exclusively of the Synechococcus Nägeli genus, which was highly abundant in Marchica (32%) compared to Oualidia (0.07%) in 2014. In 2015, the frequency dropped to 22% in Marchica and 0.04% in Oualidia. The top organisms identified as Cyanobacterial candidates in the 2014 WGS metagenome were Synechococcus strains (e.g., sp. CC9605 Salasar and sp. JA213). The sp. CC9605 strain occupies a crucial spot at the food chain base (https://www.UniProt.org/proteomes/UP000002711 accessed on 15 December 2020), whereas the sp. JA213 is not described. In addition, we were able to identify in Marchica a Cyanobacterial strain, namely, Nostoc azollae 0708 Ran (5%), but not in Oualidia. This strain was also identified in the Saidia Marina Mediterranean coast, Venice Lagoon, Eljadida Atlantic coast, and Ria Formosa Lagoon (Table 6). The Prochlorococcus Penny genus was absent in Oualidia and presented only at ~0.1% in Marchica. Differences emerged at the organismal levels as the analysis of the whole metagenome allowed; at the strain level, the identification of Prochlorococcus marinus 9313 Rocap species were the same in both lagoons with the same proportion of 3%.
3.2.4. Bacteroidetes
This group is more widespread in marine environments together with coastal waters, sediments, hydrothermal vents, and polar regions [45,46]. Most members of the Bacteroidetes phylum picked from the lagoon water samples belonged to the Flavobacteriales order (Marchica: 22% of bacterial reads in 2014; 15% in 2015; Oualidia: 20% in 2014; 20% in 2015), which is mainly composed of Cryomorphaceae and Flavobacteriaceae. Marine Flavobacteria clades can adopt different strategies to coexist [47]. Most hits (Marchica: 11% in 2014 and 8% in 2015, Oualidia: 5% in 2014 and 22% in 2015) belonged to the unassigned flavobacterial genus, according to 16S rRNA analysis, except for the genus Formosa Ivanova, which was more abundant in Marchica (5% in 2014 and 2.5% in 2015) than in Oualidia (0.36% in 2014 and 2.14% in 2015). This genus occurs in marine territories with high organic matter levels, for instance, associated with algae, fecal pellets, and invertebrates [48]. The 2014 WGS metagenomic analysis revealed two strains: Candidatus Amoebophilus asiaticus 5a2 Schmitz-Esser (10.34% in Marchica; 6.46% in Oualidia) and Bacteroides fragilis 9343 Coyne (2.87% and 2.25%, respectively).
3.2.5. Verrucomicrobia
Verrucomicrobia appears most regularly in polar and temperate zones [49]. They are also found in marine animals and plants as symbionts/parasites [50]. They feature interesting characteristics, such as the presence of genes homologous to eukaryotic tubulins [51] and methane oxidation capacity in low pH environments [52]. In the 2014 and 2015 datasets, they are largely represented by the genus Roseibacillus Yoon. In Oualidia, they showed a significant abundance (15.18%, 2014; 14.5%, 2015) compared to Marchica (3.46%, 2014; 0.86%, 2015). The OTU fraction assigned to Roseibacillus varied considerably across the Mediterranean and Atlantic sites we selected from the OSD database for this study. In Saidia Marina (OSD94) (7%, 2014; 0.5%, 2015) and Venice Lagoon (OSD47) (0.08%, 2014; 0.3%, 2015), abundances followed the same pattern as Marchica. Differences were more pronounced in the Atlantic sites, as the abundance of Roseibacillus dropped in Eljadida (OSD93) to (1%, 2014; 3%, 2015). The same was observed in Ria Formosa (OSD81) (1.5%, 2014; 0.3%, 2015). Analyses of the strain level allowed the identification of two strains, Methylacidiphilum infernorum V4 Dunfield and Akkermansia muciniphila 835 Van Passel (Table 6). Methylacidiphilum infernorum holds most of the key metabolic pathways of amino acid biosynthesis [53].
3.2.6. Actinobacteria
Actinobacteria was the least abundant phylum in both lagoons, but the proportions varied across the datasets (Marchica: 7.42%, 2014; 11.75%, 2015; Oualidia: 13.98%, 2014, 7.80%, 2015) (Table 4). In our study, it was represented by the order Micrococcales (Marchica 3% of total Actinobacteria, 2014; 0.8%, 2015; Oualidia: 15%, 2014; 7%, 2015). The marine Actinobacteria group was first reported on the southern California coast and Bermuda island [54]. Currently, they are described as omnipresent taxa of marine bacterioplankton [55] and may form up to 10–60% of the bacterioplankton group [56]. Two major strains belonging to the Micrococcales order were identified in the two studied lagoons, Rothia dentocariosa 17,931 Eisenberg and Tropheryma whipplei 27 Bentley (Table 6), both known as causative human disease strains [55]. Another bacterium belonging to the Corynebacteriales order was identified, namely, Mycobacterium leprae TN Cole, a pathogenic microorganism causing leprosy [57].
3.2.7. Eukaryotes
The 18S rRNA sequences accounted for 11,220 (14%) in Marchica and 33,442 (28%) in Oualidia during the 2014 sampling campaign, whereas 36,243 and 31,511 eukaryotic reads were obtained in 2015 (49% and 30% of total small subunit ribosomal RNA (SSUs), respectively). The main identified eukaryotic phyla were Dinophyta, Ochrophyta, and Chlorophyta (Table 7 and Table 8). Gyrodinium Hulbert (25%, 2014; 33%, 2015), Pseudo-Nitzschia Peragallo (15%, 2014; 2%, 2015), and Tetraselmis Stein (4%, 2014; 0.1%, 2015) were detected in Marchica samples in important relative abundances compared to other genera. In Oualidia, the most abundant organisms revealed by 18S rRNA were the Ostreococcus Courties genus (0%, 2014; 16%, 2015), Pelagodinium HJ Spero (12%, 2014; 0%, 2015), and Thalassiosira Cleve (8%, 2014; 25%, 2015).
3.3. Microbial Communities Functional Analysis
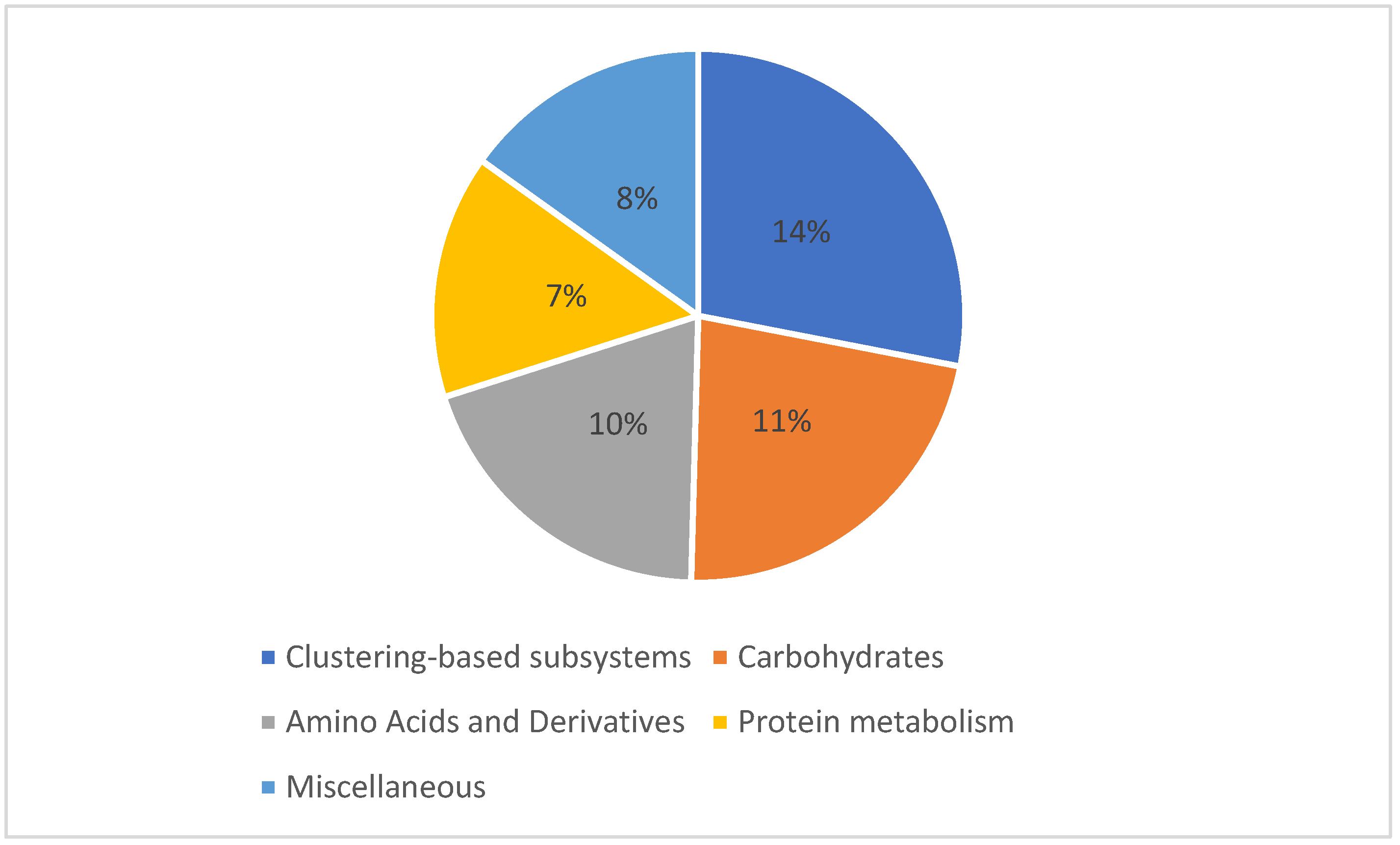
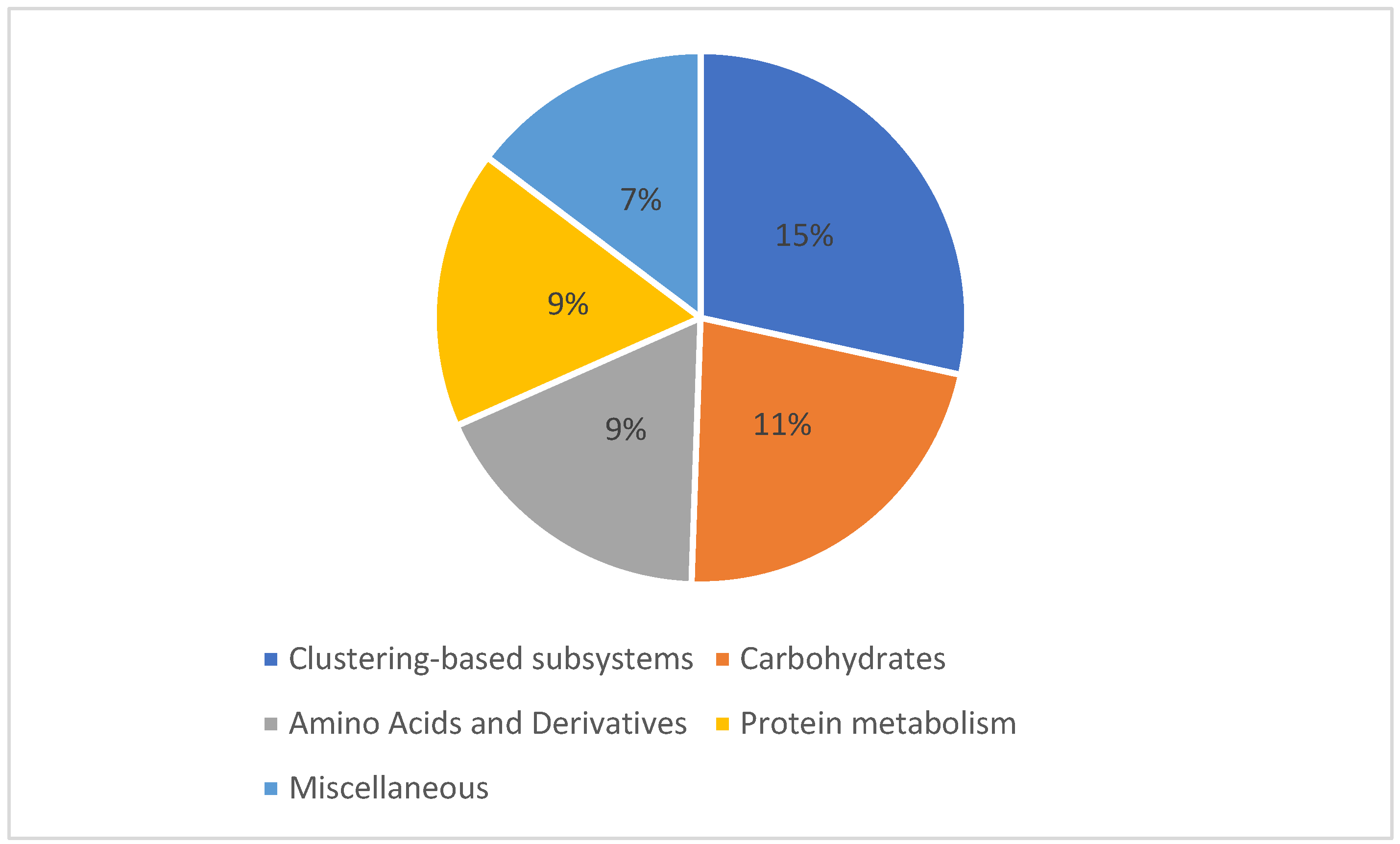
A shotgun metagenomics approach was applied to the collected and filtered samples on 21 June 2014 from Marchica and Oualidia lagoons to examine the functional diversity of the microbial communities. In Marchica, of the sequences that passed the quality control test (QC), 603 sequences possessed rRNA genes, 120,555 sequences (61.46%) had predicted proteins with known functions, and 74,995 sequences (38.23%) had predicted proteins of unknown functions. In Oualidia, 1452 sequences contained rRNA genes, 31,957 sequences (30.01%) had predicted proteins with known functions, and 73,081 sequences (68.63%) had predicted proteins with unknown functions. The top genes encoded functions were similar in both locations, mainly comprising genes required for amino acid, carbohydrate, and protein metabolism. This shows that microorganisms dominating surface water environments could share essential genes for their survival and adaptation in these ecosystems [58]. The metabolic distribution appeared to follow a similar pattern throughout all sample sets regarding composition and fraction size. In Marchica, the reads were dominated by features responsible for carbohydrate metabolism, which made up 11.9% of the total reads, followed by amino acid and derivative metabolism (10%), miscellaneous (8%), and protein metabolism reads, which were tied at 7% (Figure 8). Although they follow similar functional patterns, slightly fewer reads were assigned to metabolic functions in Oualidia (57%) than in Marchica (62%). Carbohydrates accounted for 11%, followed by amino acids and derivates and protein metabolism, with 9% of total reads in Oualidia (Figure 9). Genes encoding for essential biogeochemical processes were also present but in low proportions (1%). Functional gene properties were further assigned to microbes. For example, in Marchica, 25% of Rickettsia conorii 7 Ogata (belonging to the Proteobacteria phylum), 35% of Nostoc azollae 0708 Ran (Cyanobacteria phylum), 75% of Methylacidiphilum infernorum V4 Dunfield (Verrucomicrobia phylum), 30% of Candidatus Amoebophilus asiaticus 5a2 Schmitz-Esser (Bacteroidetes phylum), and 70% of Rothia dentocariosa 17931 Eisenberg (Actinobacteria phylum) proteins have a metabolic function (Figure 10). For comparison, in Oualidia, 15% of Candidatus Amoebophilus asiaticus 5a2 Schmitz-Esser (Proteobacteria phylum), 65% of Anaplasma marginale Maries Brayton (Proteobacteria phylum), 60% of Nostoc azollae 0708 Ran (Cyanobacteria phylum), 25% of Methylacidiphilum infernorum V4 Dunfield (Verrucomicrobia phylum), and 30% of Rothia dentocariosa 17931 Eisenberg (Actinobacteria phylum) genes have a metabolic function (Figure 11).
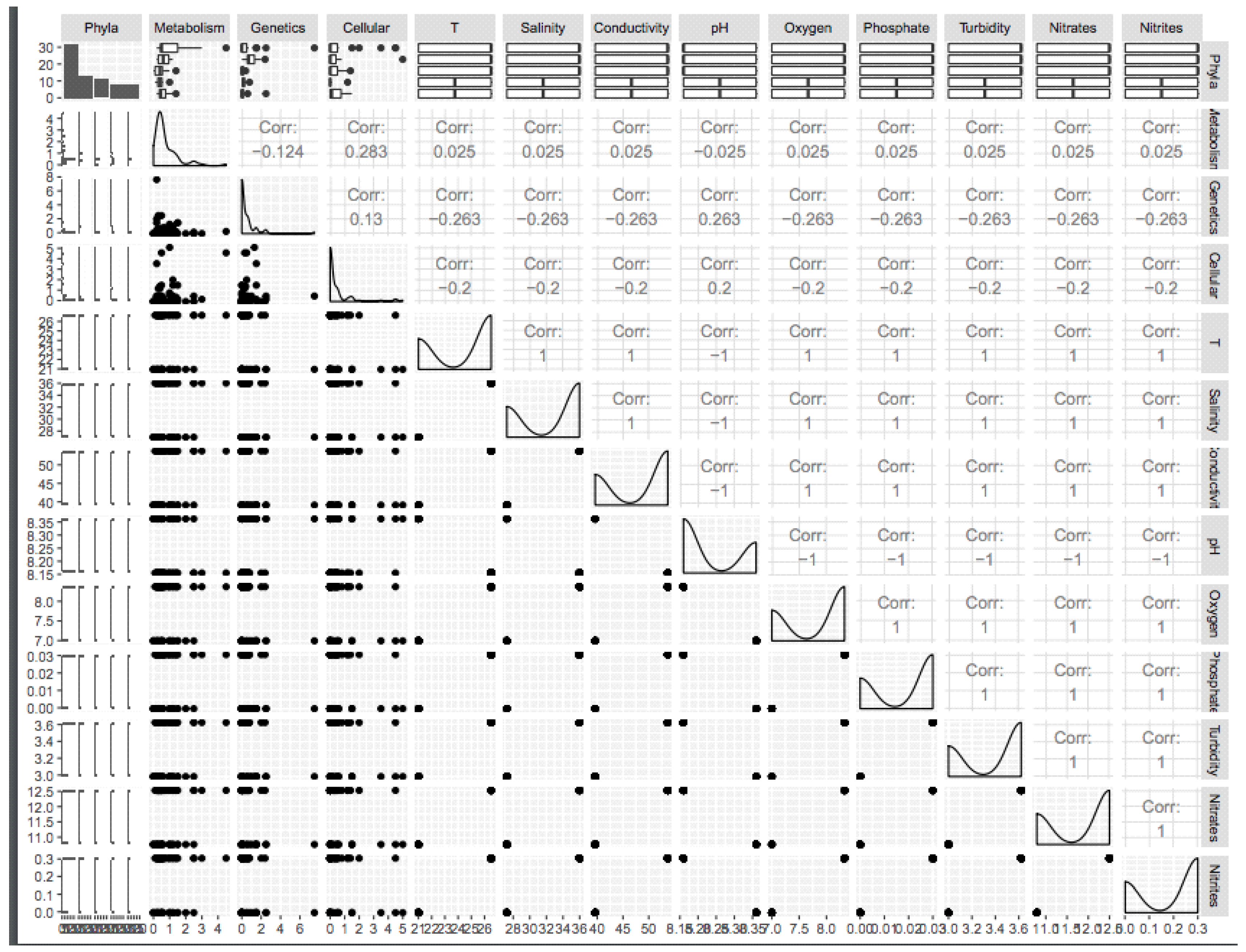
The correlation between detected gene functions and physical–chemical parameters was calculated using the R package ggally [59]. The analysis was based on nine water properties (pH, temperature, conductivity, turbidity, oxygen, salinity, nitrate, nitrite, and phosphate) and the type of gene function (metabolic, genetic, and cellular process). Seven out of the nine measured physicochemical parameters, namely, temperature, salinity, conductivity, oxygen, phosphate, nitrates, and nitrites, were positively correlated with metabolic functions (correlation = 0.025) and negatively correlated with genetic and cellular processes (correlation = −0.263 and −0.2, respectively); this correlation is almost the same among all previously cited parameters and functions. pH was positively correlated with genetic functions and cellular processes (correlation = 0.263 and 0.2, respectively) and negatively correlated with metabolic functions (correlation = −0.025) (Figure 12).
4. Discussion
In this project, we intended to delineate the microbial diversity and abundance within the surface waters of two Moroccan coastal lagoons, Marchica and Oualidia, depict their functional capabilities, and establish their variation patterns across environmental, temporal, and spatial gradients. Through the use of shotgun and amplicon sequencing metagenomics technologies, we obtained a fair amount of read data from marine water-filtered extracted DNA, which allowed us to identify the taxonomic structure and functional profiles of the microbial communities in these lagoons.
The degradation of water quality in lagoons by both natural and anthropogenic activities has a direct impact on the biodiversity and ecological balance, which has triggered the need for suitable conservation and management strategies. The physicochemical parameter measurements of temperature, salinity, electrical conductivity, pH, dissolved oxygen, phosphate, turbidity, nitrates, and nitrites were set out to assess the ecological health status of the Marchica and Oualidia lagoons. The higher rate of salinity (35 ppt) observed in Marchica in comparison to the Mediterranean Sea and Oualidia makes it a hypersaline lagoon. During the dry season, salinity increases because of the temperature increase. Other factors also impact salinity, such as shallow depth, evaporation, and volume of freshwater influx. However, in Oualidia, the absence of freshwater inflow to the lagoon suggests that the lower recorded salinity (27 ppt) is influenced by underground water resurgence [12]. pH is measured to assess the health status of aquatic ecosystems; it is known to influence the development and spread of microbes and has a direct impact on the environmental conditions that contribute to microorganism survival and expansion [60]. The pH of both lagoons was mildly alkaline, which is similar to results found by [61], where the pH of the Marchica lagoon oscillated between 8.1 and 8.7, corresponding to the wastewater treatment plant during the summer season [62]. In Oualidia, 8.03 was the pH value recorded for dry months, which implied an explicit marine influence [63]. The concentrations of dissolved oxygen in Marchica were remarkably low (8.3 mg/L) compared to those reported at Mohandis station (13.76 mg/L) in June 2013 by [64], which is possibly related to the fair oxygenation observed at the surface after the installation of an artificial inlet in 2010 that helped with recycling the Marchica lagoon waters. In addition, algal death is another factor that influences the decrease in DO in the Nador Lagoon [64]. In Oualidia Lagoon, samples were taken from the upstream region, which is known to be more oxygenated in spring and less oxygenated in summer. This is due to the growing rate of marine organisms’ respiration and aerobic biodegradation of dissolved organic matter by heterotrophic bacteria [65]. In reality, three essential factors influence the good oxygenation of Oualidia lagoon waters: freshwater inflows, depth, and surface water agitation [65]. The recorded turbidity in both lagoons (3 NTU) fell under the normal value set by the U.S. Geological Survey (USGS) (1–50 NTU). This means that the water is clear, which increases the amount of sunlight to penetrate the water layers, thereby increasing the photosynthetic rate [27]. The low phosphate concentrations recorded at both Marchica (0.03 mg/L) and Oualidia (0 mg/L) might be explained by the high renewal rate of water [65,66]. Nitrate concentrations in 2014 in Marchica and Oualidia lagoons (12.5 mg/L and 10.79 mg/L, respectively) showed slightly higher values compared to the maximum contaminant level (MCL) set in the United States for the concentration of nitrates in water (10 mg L−1) and to the World Health Organization (WHO) guidelines (11.3 mg L−1). This could be explained in Marchica by terrigenous inputs from Selouane and Caballo Oueds (streams). Furthermore, Oued Caballo carries urban wastewater from the Zeghanghane neighborhood, while Oued Selouane transports urban and toxic waste toward the lagoon [66]. Farming in the Bou Areg adjacent plain is the main surface and groundwater contamination source of nitrates, nitrites, and phosphate in Marchica. On the other hand, the wastewater treatment plant unwittingly increases these nutrients in the lagoon system [66]. In Oualidia, the quantity of fertilizers used in this agricultural area is generally the main source of nitrates [65] and heavy metals [63]. In 2015, we noticed a significant decrease in the nitrate level (from 12.5 mg/L to 4.8 mg/L in Marchica and from 10.79 mg/L to 2.68 mg/L in Oualidia), which could be due to the remediation actions by local authorities in Marchica by recycling the wastewater and materials, collecting waste, and improving the renewal of the lagoon waters. Indeed, the human impact was still noticed in Oualidia but was much less compared to 2014. This could be explained by the newly installed pit, which aimed to promote water renewal, improve water quality, and reduce the rate of suspended matter [65]. Almost certainly, our results suggest that the opening of the new inlet and the installation of waste treatment plants had a positive impact on the physicochemical properties, which indicates an improvement in the lagoon’s water quality.
Looking at the viral signals, we believe our results nicely illustrate the fact that we started to have a good “baseline database” of oceanic viral genomes (and genome fragments), although TOV_43 (Tara oceans) is highly biased toward the open ocean, whereas OSD is biased toward coastal sites. We also think that the fact that we detected the same viral genome/contingent in several distant OSD samples confirms that ocean viruses have a wide distribution (Figure 6 and Figure 7). This is consistent with what was observed in the TOV_43 study (https://www.ivirus.us/data accessed on 15 December 2020), with the added information that the OSD samples are microbial metagenomes, so should include only “actively infecting” viruses, and were synchronized (whereas Tara sampling was spread across multiple years), so we have the actual “proof” that the same virus was active at the same time in multiple places. Finally, we think we may correlate this viral signal to putative hosts: we will not have strong statistical power, but for the “known” viruses, we should be able to check if the corresponding host was indeed abundant in the same sample, and, for the “unknowns”, check which were the major microbial groups in the corresponding samples that would at least provide putative hosts.
By examining the community composition, samples from both studied locations appeared to be made up mostly of Proteobacteria members (α and γ), Cyanobacteria (Synechococcus), Bacteroidetes (Flavobacteriales), Verrucomicrobia (Verrucomicrobiae), and Actinobacteria (Actinobacteria). Alphaproteobacteria seemed to be the highest identified class across the different sampling locations, covering an average of 20% to 45% of the microbial communities in Oualidia and Marchica, respectively. The class consisted of Rhodobacteraceae, which are classified as denitrifiers capable of rapid growth and reducing nitrates in both oxic and anoxic conditions [40]. Their increasing abundance in lagoon coastal waters might be linked with the decrease in nitrogen content at both Marchica and Oualidia stations in 2015 (4 mg/L, 2 mg/L) compared to 2014 (14 mg/L, 2 mg/L). A sizeable proportion of reads belonged to Candidatus Pelagibacter Rappé (11% in Marchica; 7% in Oualidia) in 2014. Interestingly, in 2015, none were detected. This genus is known as an eminent member of SAR11 clusters, which are classified as the largest group of bacteria in the surface water of marine ecosystems [67]. This is in line with findings from Ghai et al. (2012) on Spanish Mar Menor Lagoon, where half of all 16S reads were assigned to Candidatus Pelagibacter (43%). In addition, SAR11 reads were encountered in all samples collected from the surface waters of different marine habitats either in the Pacific or Atlantic oceans, specifically coastal environments, as reported by Tara Oceans and the Global Ocean Sampling Expedition [3]. Different strains were within the Rhodobacterales group, particularly Rickettsia conorii 7 Ogata, which formed the most abundant strain identified in both lagoons. It is generally present on the Mediterranean side of Europe and is known to cause boutonneuse fever [68]. Gammaproteobacteria, in contrast, made up lower proportions compared to Alphaproteobacteria. This is typical of Gammaproteobacteria because they are found in large counts in benthic and lower pelagic marine environments [69]. This class was composed of Alteromonadales, which have been found to be related to sites affected by urbanization and eutrophication [70]. In addition, members of this group are resistant to metals [70]. Spatial and temporal variation in the distribution of Alteromonadales compared to Rhodobacterales might be assigned to competition for limiting nutrients [71]. The strain Xylella fastidiosa Temecula1 Sluys, classified among the top dangerous pathogenic bacteria infecting plants worldwide [72], was the dominant strain found in the Gammaproteobacteria group.
We found further evidence of the occurrence of almost exclusively the genus Synechococcus Nägeli as the most abundant Cyanobacteria, notably in Marchica, which was similarly observed in Mar Menor Lagoon, where Synechococcus formed a sizeable percentage and declined with increasing salinity in the Mediterranean Sea [7]. Almost no Prochlorococcus Penny populations were found in Oualidia waters, which is similar to the results in the parent Atlantic water body from which Oualidia waters are derived (OSD 93). This is the usual behavior of Prochlorococcus, which is commonly not found in coastal marine environments [73]. In addition, the Synechococcus growth rate increases with increasing temperature and nitrates, while increasing temperature and light levels might retard Prochlorococcus growth [71]. In fact, salinity and temperature are essential environmental factors for the composition and function of Synechococcus in the marine microbiome [74]. According to previous studies, Cyanobacteria grow in nutrient-enriched ecosystems, and their prevalence increases with eutrophication [75]. However, when the macronutrient concentration is low in the summer season, Synechococcus shows fast growth [75]. The highest number of Synechococcus read recruitment in Marchica in 2014 compared to 2015 could be explained by the lagoon’s intensive environmental rehabilitation effort, which includes cleaning wastes and depollution. Based on strain identification in the metagenome, we were able to identify the cyanobacterium Nostoc azollae 0708 Ran, which is famous for its nitrogen-fixation properties and is commonly used in farming [76].
Most of the remaining sequences were mainly assigned to moderately smaller phyla, including Bacteroidetes, Verrucomicrobia, and Actinobacteria. We noticed the unique existence of Bacteroides fragilis 9343 Coyne (3%) in Venice Lagoon, and Candidatus Amoebophilus asiaticus 5a2 Schmitz-Esser (9%) comprised the underrepresented Cytophagales group in the Atlantic Ria Formosa Lagoon. The former strain could potentially be used as an organic pollution bioindicator [77].
Archaeal assemblages made up a tiny percentage of the metagenome, forming less than 1% of the total sequences in both Marchica and Oualidia lagoons. These findings were in agreement with previous reports where barely detectable levels of archaeal sequences were found in surface marine environments [7,78,79]. In contrast, Archaea have a greater preference for deep waters, where they exist in significantly higher numbers [80,81]. Euryarchaeota were the dominant clade, which is normal as they are known to inhabit surface waters [82].
Phytoplankton sequence data showed dissimilarities between the two lagoons, where Marchica was dominated by dinoflagellates and Oualidia by Ochrophytes and Chlorophytes. The red dinoflagellate Gyrodinium Hulbert belongs to the family Dinophyceae, which lack armor protists responsible for red tides [83]. The Ochrophyta genus Pseudo-Nitzschia Peragallo, which belongs to diatoms, is responsible for amnesic shellfish poisoning (ASP); blooms were observed globally in coastal waters and were linked to elevated nutrient concentrations in marine environments [76]. While the Chlorophyta genus Tetraselmis Stein is a green alga within the order Chlorodendrales, the species members’ habitat preference depends on whether the water is deep or shallow because of their photosynthetic nature. Thus, they inhabit different water environments if sufficient light and nutrients are accessible [74]. In addition, many species belonging to the genus Tetraselmis have recently been investigated for use as biofuels because of their increased lipidic content [84]. Both Gyrodinium and Nitzschia were abundantly identified in the Mediterranean Spanish coastal lagoon Mar Menor using microscopic examination and enumeration. These diatom species are linked to high nutrient concentrations, especially phosphorus, and are hence associated with contaminated eutrophic waters [7]. Overall, the taxonomy of these groups is still to be elucidated [7]. In Oualidia, most hits were assigned to the Ostreococcus Courties genus, a unicellular green alga that has a crucial role in the oceanic carbon cycle, and a model to study the adaptation of green algae in the marine environment, Pelagodinium HJ Spero and Thalassiosira Cleve. The latter is widely distributed throughout the world’s oceans [85].
The functional diversity of microbial communities was explored in our study using a shotgun metagenomics approach. Across the different sampling sites, we observed similar patterns in the identified communities with regard to their functional features. The topmost functional genes were closely similar in Marchica and Oualidia, essentially comprising genes implicated in protein, carbohydrate, and amino acid metabolism. Previous reports supported these findings, as the surface microbial communities were found to be located in the Mediterranean Sea, Atlantic Ocean, and Pacific Ocean, where they share a close dissemination of genes encoded functions [3,78]. This would possibly suggest that the survival and adaptation of these microbial communities depends on the corset of genes they share in the surface water environments [58]. Remarkably, Methylacidiphilum infernorum V4 Dunfield and Rothia dentocariosa 17931 Eisenberg exhibited the highest proportions of genes with metabolic functions in Marchica. However, these amounts were related to Anaplasma marginale Maries Brayton and Nostoc azollae 0708 Ran in Oualidia. Furthermore, the correlation between microbiome gene expression and water properties showed that the latter, namely, temperature, salinity, conductivity, oxygen, phosphate, nitrates, and nitrites, influenced the expression of genes with metabolic functions, as the correlation was positive. This was indeed reported by [86], who studied five shallow coastal lagoons in Brazil and concluded that temperature and the concentration of nutrients are responsible for bacterial metabolism regulation. However, the pH in our study showed a negative impact on metabolic processes. This result is in accordance with a study by [87], which found that pH has no influence on bacterial abundance.
This metagenomics study enriched the Moroccan national catalog of marine microorganisms that can be used for bioindication, biodiversity protection, biotechnology valorization, biomonitoring, and lagoon health assessment. For instance, the presence of Synechococcus Nägeli and Bacteroides fragilis Coyne belonging to the Cyanobacteria and Bacteroides groups, respectively, could be associated with the eutrophic conditions of the lagoon Marchica. Indeed, the rate of nitrogen in the Marchica Lagoon is alarmingly high (12 mg/L). Hence, we may further suggest using these marine bacteria as bioindicators of deteriorated water quality in coastal lagoons. Notably, Cyanobacteria species are good candidates for environmental sustainability and an excellent agricultural bioresource [88]. Take, for example, Nostoc Azollae Ran, a biofertilizer used for either Taro or the cultivation of rice. It has also shown great bioremediation potential in reducing pollutants and improving waste management [77]. Cyanobacteria have been remarkably implemented in blue biotechnology, a newly emerging solution to the high demand for biomedicine, pharmaceutical and agricultural products, natural cosmetics, and sustainable energy sources by modern societies through exploiting marine biodiversity. They have shown their ability to generate molecules with antibacterial, antifungal, and antiviral traits. Today, they are being investigated for their anti-multiplicative effect on cancer cells. An average of 400 biomolecules secreted by Cyanobacteria strains have been listed [89]. Other identified marine bacteria in our study, although with low frequencies, expressed an impressive prospect for biotechnology application, for example, Alcanivorax Yakimov and Acinetobacter Brisou species. The former plays an important role in oil spill bioremediation [90] and when together they are of high interest for beauty technology since they have been shown to produce biosurfactants [91]. Proper management of lagoon health is another important aspect to take into consideration by implementing biomonitoring, which will help us eventually better understand exposures to environmental chemicals, shape public health, and prevent the development of specific diseases caused by particular microorganisms, such as Xylella fastidiosa Sluys, a pathogen infecting plants that in Brazil infected approximately 200 million citrus trees and caused damage estimated at USD 100 million in the grape industry in California [72]. We were also able to detect Mycobacterium leprae Hansen in the waters of both lagoons, which could possibly lead to leprosy infection. However, further investigations are needed to support this claim.
In connection with spatial variation, the microbial community was dominated by similar phyla identified in 2014 and 2015. However, clear differences were manifested at the genus, species, and strain divisions among samples. Hence, the bacterioplankton were diverse but modestly varied across samples, as shown by the alpha diversity. The observed richness in Oualidia was 1144 in 2014 and 704 in 2015, while in Marchica, we found 462 in 2014 and 316 in 2015. This also elucidated the importance of performing more sampling as the number of species captured fluctuates and the taxonomic richness increases with a high number of reads. Each lagoon chosen for this work displayed differences in physicochemical parameters such as temperature, salinity, DO, or nitrate concentrations in addition to reported anthropogenic pressures due to agricultural, industrial, and human impacts, which probably influenced microbial community distinctions at each site. The relationship between spatial variation and community structure needs more study for better understanding.
In conclusion, the results gained from this study provide the first comparison of microbial communities found in Moroccan coastal lagoon waters. They will serve as a valuable platform for further advanced studies to better understand the microbiome in lagoon ecosystems. More locations, various depths, and other lagoons should be sampled for more community descriptions within these waters. This will simultaneously strengthen existing findings of community patterns and confirm hypotheses that have been made on the relationship between microbial community structures, geography, and physical–chemical parameters. Seasonality studies can also be conducted to observe any changes in patterns by adding a broader time scale. The prokaryotic communities among a vertical profile can be analyzed and compared to those sampled across a horizontal gradient. It would also be interesting to further study microbial populations in surface lagoon waters and more deeply. A focus on minor and underrepresented taxa and a more profound and extensive analysis of the sequences obtained from these metagenomes can be carried out later to identify the taxonomic identity and functions of unknown reads and define more bioindicators/biomarkers of fragile marine ecosystems such as lagoons.
Author Contributions
Conceptualization, H.G., L.A.-Z.; methodology, H.G., L.A.-Z., B.C.; software, B.C., A.I.A.; validation, H.G., S.B., R.C.; formal analysis, B.C.; investigation: H.G., B.C., S.E., E.H.A., A.B.; resources, S.E., M.T., N.B., data curation, B.C.; C.E.H.; S.R.; writing—original draft preparation, B.C.; writing—review and editing, H.G., L.A.-Z., S.A., A.T., S.B.; R.C., F.E.O., A.E.H.; N.B., C.N.; visualization, B.C.; supervision, H.G., L.A.-Z., E.H.Z., N.H., F.B.; project administration H.G.; Funding acquisition, H.G., L.A.-Z., C.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the EU MicroB3 project, funded by the European Union’s Seventh Framework Programme (FP7; Joint Call OCEAN.2011-2: Marine microbial diversity-new insights into marine ecosystems functioning and its biotechnological potential) under grant agreement no 287589. Bouchra Chaouni is a recipient of a one year Fulbright fellowship at the Biological Marine Laboratory, Woods Hole, MA, USA for the academic year 2017/18. Hassan Ghazal is a US NIH grant recipient through the H3abionet/H3africa consortium U24HG006941. The APC was funded by the University Mohamed IV of Health Sciences.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The bacterial metagenome and 16S rRNA raw read sequence datasets were deposited in EMBL-EBI https://www.ebi.ac.uk/metagenomics/studies/MGYS00001977 accessed 1 December 2019 and OSD GitHub https://github.com/MicroB3-IS/osd-analysis/wiki accessed 1 December 2019.
Acknowledgments
We thank Richard Fox for his help with the bioinformatics analysis carried out on the MBL server and Andrew Voorhis and Anna Shipunova for their assistance with VAMPS. We are also grateful to Sullivan Matthew, Bolduc Benjamin, and Roux Simon from Ohio State University, Department of Microbiology, USA, for their help in the analysis of the viral phages, Kris Gunsalus for access to bioinformatics resources, and the Ocean Sampling Day consortium for supporting the overall work.
Conflicts of Interest
The authors declare that the research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Sánchez-Baracaldo, P.; Bianchini, G.; Wilson, J.D.; Knoll, A.H. Cyanobacteria and Biogeochemical Cycles through Earth History. Trends Microbiol. 2022, 30, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Amaral-Zettler, L.; Artigas, L.F.; Baross, J.; LokaBharathi, P.A.; Boetius, A.; Chandramohan, D.; Herndl, G.; Kogure, K.; Neal, P.; Pedros-Alio, C.; et al. A Global Census of Marine Microbes; Wiley-Blackwell: Hoboken, NJ, USA, 2010. [Google Scholar]
- Venter, J.C.; Remington, K.; Heidelberg, J.F.; Halpern, A.L.; Rusch, D.; Eisen, J.A.; Wu, D.; Paulsen, I.; Nelson, K.E.; Nelson, W.; et al. Environmental Genome Shotgun Sequencing of the Sargasso Sea. Science 2004, 304, 66–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunagawa, S.; Coelho, L.P.; Chaffron, S.; Kultima, J.R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D.R.; Alberti, A.; et al. Structure and Function of the Global Ocean Microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, H.; Lovell, C.R. Microbial Surface Colonization and Biofilm Development in Marine Environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Ren, H.; Shen, L.; Lou, L.; Tian, G.; Zheng, P.; Hu, B. PH Levels Drive Bacterial Community Structure in Sediments of the Qiantang River as Determined by 454 Pyrosequencing. Front. Microbiol. 2015, 6, 285. [Google Scholar] [CrossRef]
- Ghai, R.; Hernandez, C.M.; Picazo, A.; Mizuno, C.M.; Ininbergs, K.; Díez, B.; Valas, R.; DuPont, C.L.; McMahon, K.D.; Camacho, A.; et al. Metagenomes of Mediterranean Coastal Lagoons. Sci. Rep. 2012, 2, 490. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Knight, R. Global Patterns in Bacterial Diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 11436–11440. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Yan, Z.; Wang, P.; Zheng, X.; Fan, J. Comparative Metagenomics Reveals the Microbial Diversity and Metabolic Potentials in the Sediments and Surrounding Seawaters of Qinhuangdao Mariculture Area. PLoS ONE 2020, 15, e0234128. [Google Scholar] [CrossRef]
- Ruiz, F.; Abad, M.; Olías, M.; Galán, E.; González, I.; Aguilá, E.; Hamoumi, N.; Pulido, I.; Cantano, M. The Present Environmental Scenario of the Nador Lagoon (Morocco). Environ. Res. 2006, 102, 215–229. [Google Scholar] [CrossRef]
- El Madani, F.; Chiaar, A.; Chafi, A. Phytoplankton Composition and Abundance Assessment in the Nador Lagoon (Mediterranean Coast of Morocco). Acta Bot. Croat. 2011, 70, 269–288. [Google Scholar] [CrossRef]
- Natij, L.; Damsiri, Z.; Khalil, K.; Loudiki, M.; Ettahiri, O.; Elkalay, K. Phytoplankton Abundance and Diversity in the Coastal Waters of Oualidia Lagoon, South Moroccan Atlantic in Relation to Environmental Variables. Int. J. Adv. Res. 2014, 2, 1022–1032. [Google Scholar]
- Kopf, A.; Bicak, M.; Kottmann, R.; Schnetzer, J.; Kostadinov, I.; Lehmann, K.; Fernandez-Guerra, A.; Jeanthon, C.; Rahav, E.; Ullrich, M.; et al. The Ocean Sampling Day Consortium. GigaScience 2015, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every Base Matters: Assessing Small Subunit RRNA Primers for Marine Microbiomes with Mock Communities, Time Series and Global Field Samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Huse, S.M.; Mark Welch, D.B.; Voorhis, A.; Shipunova, A.; Morrison, H.G.; Eren, A.M.; Sogin, M.L. VAMPS: A Website for Visualization and Analysis of Microbial Population Structures. BMC Bioinform. 2014, 15, 41. [Google Scholar] [CrossRef] [Green Version]
- Huse, S.M.; Dethlefsen, L.; Huber, J.A.; Welch, D.M.; Relman, D.A.; Sogin, M.L. Exploring Microbial Diversity and Taxonomy Using SSU RRNA Hypervariable Tag Sequencing. PLoS Genet. 2008, 4, e1000255. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. MetaSPAdes: A New Versatile Metagenomic Assembler. Genome Res. 2017, 27, 824–834. [Google Scholar] [CrossRef] [Green Version]
- Mikheenko, A.; Saveliev, V.; Gurevich, A. MetaQUAST: Evaluation of Metagenome Assemblies. Bioinformatics 2016, 32, 1088–1090. [Google Scholar] [CrossRef] [Green Version]
- Hyatt, D.; Chen, G.-L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic Gene Recognition and Translation Initiation Site Identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [Green Version]
- Truong, D.T.; Franzosa, E.A.; Tickle, T.L.; Scholz, M.; Weingart, G.; Pasolli, E.; Tett, A.; Huttenhower, C.; Segata, N. MetaPhlAn2 for Enhanced Metagenomic Taxonomic Profiling. Nat. Methods 2015, 12, 902–903. [Google Scholar] [CrossRef] [PubMed]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The Metagenomics RAST Server—A Public Resource for the Automatic Phylogenetic and Functional Analysis of Metagenomes. BMC Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Brum, J.R.; Ignacio-Espinoza, J.C.; Roux, S.; Doulcier, G.; Acinas, S.G.; Alberti, A.; Chaffron, S.; Cruaud, C.; de Vargas, C.; Gasol, J.M.; et al. Patterns and Ecological Drivers of Ocean Viral Communities. Science 2015, 348, 1261498. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Aknaf, A.; Akodad, M.; Layachi, M.; El Madani, F.; Jaddar, A.; Mesfioui, A.; Baghour, M. Study of the Spatial and Temporal Variation of Physical-Chemical Parameters Characterizing the Quality of Surface Waters of the Lagoon Marchica–North-East Morocco. J. Mater. Environ. Sci. 2017, 8, 3216–3225. [Google Scholar]
- Padan, E.; Bibi, E.; Ito, M.; Krulwich, T.A. Alkaline PH Homeostasis in Bacteria: New Insights. Biochim. Biophys. Acta 2005, 1717, 67–88. [Google Scholar] [CrossRef] [Green Version]
- Kortz, A.R.; Magurran, A.E. Increases in Local Richness (α-Diversity) Following Invasion Are Offset by Biotic Homogenization in a Biodiversity Hotspot. Biol. Lett. 2019, 15, 20190133. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E. Measuring Biological Diversity; John Wiley & Sons: Hoboken, NJ, USA, 2003. [Google Scholar]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Kim, B.-R.; Shin, J.; Guevarra, R.; Lee, J.H.; Kim, D.W.; Seol, K.-H.; Lee, J.-H.; Kim, H.B.; Isaacson, R. Deciphering Diversity Indices for a Better Understanding of Microbial Communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, P.; Yarza, P.; Rapp, J.Z.; Glöckner, F.O. Expanding the World of Marine Bacterial and Archaeal Clades. Front. Microbiol. 2016, 6, 1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.S. The Phylogeny of Proteobacteria: Relationships to Other Eubacterial Phyla and Eukaryotes. FEMS Microbiol. Rev. 2000, 24, 367–402. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.; Lovell, C.R. Bacterial Primary Colonization and Early Succession on Surfaces in Marine Waters as Determined by Amplified RRNA Gene Restriction Analysis and Sequence Analysis of 16S RRNA Genes. Appl. Environ. Microbiol. 2000, 66, 467–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Yang, J.; Liu, L.; Zhang, W.; Amalfitano, S. Bacterioplankton Community Shifts Associated with Epipelagic and Mesopelagic Waters in the Southern Ocean. Sci. Rep. 2015, 5, 12897. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.A.; Steele, J.A.; Caporaso, J.G.; Steinbrück, L.; Reeder, J.; Temperton, B.; Huse, S.; McHardy, A.C.; Knight, R.; Joint, I.; et al. Defining Seasonal Marine Microbial Community Dynamics. ISME J. 2012, 6, 298–308. [Google Scholar] [CrossRef] [Green Version]
- Hartsock, A.; Shapleigh, J.P. Physiological Roles for Two Periplasmic Nitrate Reductases in Rhodobacter Sphaeroides 2.4.3 (ATCC 17025). J. Bacteriol. 2011, 193, 6483–6489. [Google Scholar] [CrossRef] [Green Version]
- Vincent, P.; Pignet, P.; Talmont, F.; Bozzi, L.; Fournet, B.; Guezennec, J.; Jeanthon, C.; Prieur, D. Production and Characterization of an Exopolysaccharide Excreted by a Deep-Sea Hydrothermal Vent Bacterium Isolated from the Polychaete Annelid Alvinella Pompejana. Appl. Environ. Microbiol. 1994, 60, 4134–4141. [Google Scholar] [CrossRef] [Green Version]
- Morris, R.M.; Rappé, M.S.; Connon, S.A.; Vergin, K.L.; Siebold, W.A.; Carlson, C.A.; Giovannoni, S.J. SAR11 Clade Dominates Ocean Surface Bacterioplankton Communities. Nature 2002, 420, 806–810. [Google Scholar] [CrossRef]
- Solano-Gallego, L.; Kidd, L.; Trotta, M.; Marco, M.D.; Caldin, M.; Furlanello, T.; Breitschwerdt, E.B. Febrile Illness Associated with Rickettsia Conorii Infection in Dogs from Sicily. Emerg. Infect. Dis. 2006, 12, 1985–1988. [Google Scholar] [CrossRef]
- Introduction to the Cyanobacteria. Available online: https://ucmp.berkeley.edu/bacteria/cyanointro.html (accessed on 27 February 2022).
- Bullerjahn, G.S.; Post, A.F. Physiology and Molecular Biology of Aquatic Cyanobacteria. Front. Microbiol. 2014, 5, 359. [Google Scholar] [CrossRef]
- Cooke, G.D.; Welch, E.B.; Peterson, S.; Nichols, S.A. Restoration and Management of Lakes and Reservoirs, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2005; ISBN 978-0-429-18923-4. [Google Scholar]
- Fernández-Gómez, B.; Richter, M.; Schüler, M.; Pinhassi, J.; Acinas, S.G.; González, J.M.; Pedrós-Alió, C. Ecology of Marine Bacteroidetes: A Comparative Genomics Approach. ISME J. 2013, 7, 1026–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GLOBAL: Marine Census Publishes Latest Findings. Available online: https://www.universityworldnews.com/post.php?story=20100903170738623 (accessed on 27 February 2022).
- Gómez-Pereira, P.R.; Fuchs, B.M.; Alonso, C.; Oliver, M.J.; van Beusekom, J.E.E.; Amann, R. Distinct Flavobacterial Communities in Contrasting Water Masses of the North Atlantic Ocean. ISME J. 2010, 4, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Mann, A.J.; Hahnke, R.L.; Huang, S.; Werner, J.; Xing, P.; Barbeyron, T.; Huettel, B.; Stüber, K.; Reinhardt, R.; Harder, J.; et al. The Genome of the Alga-Associated Marine Flavobacterium Formosa Agariphila KMM 3901T Reveals a Broad Potential for Degradation of Algal Polysaccharides. Appl. Environ. Microbiol. 2013, 79, 6813–6822. [Google Scholar] [CrossRef] [Green Version]
- Cardman, Z.; Arnosti, C.; Durbin, A.; Ziervogel, K.; Cox, C.; Steen, A.D.; Teske, A. Verrucomicrobia Are Candidates for Polysaccharide-Degrading Bacterioplankton in an Arctic Fjord of Svalbard. Appl. Environ. Microbiol. 2014, 80, 3749–3756. [Google Scholar] [CrossRef] [Green Version]
- Bünger, W.; Jiang, X.; Müller, J.; Hurek, T.; Reinhold-Hurek, B. Novel Cultivated Endophytic Verrucomicrobia Reveal Deep-Rooting Traits of Bacteria to Associate with Plants. Sci. Rep. 2020, 10, 8692. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.; Samudrala, R.; Anderson, I.; Hedlund, B.P.; Petroni, G.; Michailova, N.; Pinel, N.; Overbeek, R.; Rosati, G.; Staley, J.T. Genes for the Cytoskeletal Protein Tubulin in the Bacterial Genus Prosthecobacter. Proc. Natl. Acad. Sci. USA 2002, 99, 17049–17054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunfield, P.F.; Yuryev, A.; Senin, P.; Smirnova, A.V.; Stott, M.B.; Hou, S.; Ly, B.; Saw, J.H.; Zhou, Z.; Ren, Y.; et al. Methane Oxidation by an Extremely Acidophilic Bacterium of the Phylum Verrucomicrobia. Nature 2007, 450, 879–882. [Google Scholar] [CrossRef]
- Hou, S.; Makarova, K.S.; Saw, J.H.W.; Senin, P.; Ly, B.V.; Zhou, Z.; Ren, Y.; Wang, J.; Galperin, M.Y.; Omelchenko, M.V.; et al. Complete Genome Sequence of the Extremely Acidophilic Methanotroph Isolate V4, Methylacidiphilum Infernorum, a Representative of the Bacterial Phylum Verrucomicrobia. Biol. Direct 2008, 3, 26. [Google Scholar] [CrossRef] [Green Version]
- Ventura, M.; Canchaya, C.; Tauch, A.; Chandra, G.; Fitzgerald, G.F.; Chater, K.F.; van Sinderen, D. Genomics of Actinobacteria: Tracing the Evolutionary History of an Ancient Phylum. Microbiol. Mol. Biol. Rev. 2007, 71, 495–548. [Google Scholar] [CrossRef] [Green Version]
- Shivlata, L.; Satyanarayana, T. Thermophilic and Alkaliphilic Actinobacteria: Biology and Potential Applications. Front. Microbiol. 2015, 6, 1014. [Google Scholar] [CrossRef]
- Pinto, I.; Calisto, R.; Serra, C.R.; Lage, O.M.; Antunes, S.C. Bacterioplankton Community as a Biological Element for Reservoirs Water Quality Assessment. Water 2021, 13, 2836. [Google Scholar] [CrossRef]
- Pinheiro, R.O.; de Souza Salles, J.; Sarno, E.N.; Sampaio, E.P. Mycobacterium Leprae-Host-Cell Interactions and Genetic Determinants in Leprosy: An Overview. Future Microbiol. 2011, 6, 217–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewson, I.; Paerl, R.W.; Tripp, H.J.; Zehr, J.P.; Karl, D.M. Metagenomic Potential of Microbial Assemblages in the Surface Waters of the Central Pacific Ocean Tracks Variability in Oceanic Habitat. Limnol. Oceanogr. 2009, 54, 1981–1994. [Google Scholar] [CrossRef]
- Schloerke, B.; Cook, D.; Larmarange, J.; Briatte, F.; Marbach, M.; Thoen, E.; Elberg, A.; Crowley, J. Extension to “ggplot2” [R Package GGally Version 2.0.0]. 2020. Available online: https://ggobi.github.io/ggally/index.html (accessed on 27 March 2022).
- Thompson, L.R.; Sanders, J.G.; McDonald, D.; Amir, A.; Ladau, J.; Locey, K.J.; Prill, R.J.; Tripathi, A.; Gibbons, S.M.; Ackermann, G.; et al. A Communal Catalogue Reveals Earth’s Multiscale Microbial Diversity. Nature 2017, 551, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Matoir, M.; Belabed, A.; Najih, M.; Kada, O.; Rezzoume, N. Surrounding influence on the Ecological state of the lagoon of Marchica. J. Mater. Environ. Sci. 2015, 1260–1265. [Google Scholar]
- Riouchi, O.; Madani, F.E.; Abadie, E.; Skalli, A.; Baghour, M. The Spatio-Temporal Evolution of the Genus Nitzschia Longissima at the Level of the Lagoon in Nador, Morocco. E3S Web Conf. 2021, 234, 00081. [Google Scholar] [CrossRef]
- Hassou, N.; Boujaber, N.; Oumaskour, K.; Lakhdar, F.; Benba, J.; Benhaddou, Z.; Assoobhei, O.; Etahiri, S. Nutrients Variation in the Coastal Waters of Oualidia Lagoon, Morocco. Relation to the Contribution of Watershed and Hydro-Chemical Characteristics. Int. J. Adv. Res. 2016, 4, 1697–1706. [Google Scholar] [CrossRef] [Green Version]
- Chagas, G.G.; Suzuki, M.S. Seasonal Hydrochemical Variation in a Tropical Coastal Lagoon (Açu Lagoon, Brazil). Braz. J. Biol. 2005, 65, 597–607. [Google Scholar] [CrossRef] [Green Version]
- Damsiri, Z.; Natij, L.; Khalil, K.; Loudiki, M.; Rabouille, C.; Ettahiri, O.; Bougadir, B.; Elkalay, K. Spatio-Temporal Nutrients Variability in the Oualidia Lagoon (Atlantic Moroccan Coast). Int. J. Curr. Adv. Res. 2014, 2, 609–618. [Google Scholar]
- Aknaf, A.; Akodad, M.; Moumen, A.; Chekroun, K.B.; Elhamouti, C.; Bailal, A.; Baghour, M. Impact of the New Pass on the Eutrophication of the Lagoon Marchica: Study of the Two Sites Bou Areg and Mohandis. J. Mater. Environ. Sci. 2015, 6, 2939–2943. [Google Scholar]
- Giovannoni, S.J. SAR11 Bacteria: The Most Abundant Plankton in the Oceans. Annu. Rev. Mar. Sci. 2017, 9, 231–255. [Google Scholar] [CrossRef] [PubMed]
- MacConnachie, K.; Tishkowski, K. Boutonneuse Fever. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Zinger, L.; Amaral-Zettler, L.A.; Fuhrman, J.A.; Horner-Devine, M.C.; Huse, S.M.; Welch, D.B.M.; Martiny, J.B.H.; Sogin, M.; Boetius, A.; Ramette, A. Global Patterns of Bacterial Beta-Diversity in Seafloor and Seawater Ecosystems. PLoS ONE 2011, 6, e24570. [Google Scholar] [CrossRef] [PubMed]
- Juhmani, A.-S.; Vezzi, A.; Wahsha, M.; Buosi, A.; Pascale, F.D.; Schiavon, R.; Sfriso, A. Diversity and Dynamics of Seaweed Associated Microbial Communities Inhabiting the Lagoon of Venice. Microorganisms 2020, 8, 1657. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.-H.; Kang, I.; Yang, S.-J.; Cho, J.-C. Characterization of Spatial Distribution of the Bacterial Community in the South Sea of Korea. PLoS ONE 2017, 12, e0174159. [Google Scholar] [CrossRef]
- Schneider, K.; van der Werf, W.; Cendoya, M.; Mourits, M.; Navas-Cortés, J.A.; Vicent, A.; Oude Lansink, A. Impact of Xylella Fastidiosa Subspecies Pauca in European Olives. Proc. Natl. Acad. Sci. USA 2020, 117, 9250–9259. [Google Scholar] [CrossRef] [Green Version]
- Partensky, F.; Blanchot, J.; Vaulot, D. DifferentiaI Distribution and Ecology of Prochlorococcus and Synechococcus in Oceanic Waters: A Review. Bull. Inst. Océanographique Monaco 1999, 19, 457–475. [Google Scholar]
- Kim, Y.; Jeon, J.; Kwak, M.S.; Kim, G.H.; Koh, I.; Rho, M. Photosynthetic Functions of Synechococcus in the Ocean Microbiomes of Diverse Salinity and Seasons. PLoS ONE 2018, 13, e0190266. [Google Scholar] [CrossRef] [Green Version]
- Lürling, M.; Mello, M.M.E.; van Oosterhout, F.; de Senerpont Domis, L.; Marinho, M.M. Response of Natural Cyanobacteria and Algae Assemblages to a Nutrient Pulse and Elevated Temperature. Front. Microbiol. 2018, 9, 1851. [Google Scholar] [CrossRef] [Green Version]
- Brouwer, P.; Bräutigam, A.; Buijs, V.A.; Tazelaar, A.O.E.; van der Werf, A.; Schlüter, U.; Reichart, G.-J.; Bolger, A.; Usadel, B.; Weber, A.P.M.; et al. Metabolic Adaptation, a Specialized Leaf Organ Structure and Vascular Responses to Diurnal N2 Fixation by Nostoc Azollae Sustain the Astonishing Productivity of Azolla Ferns without Nitrogen Fertilizer. Front. Plant Sci. 2017, 8, 442. [Google Scholar] [CrossRef] [Green Version]
- Niestępski, S.; Harnisz, M.; Korzeniewska, E.; Osińska, A. Markers Specific to Bacteroides Fragilis Group Bacteria as Indicators of Anthropogenic Pollution of Surface Waters. Int. J. Environ. Res. Public Health 2020, 17, 7137. [Google Scholar] [CrossRef]
- Rusch, D.B.; Halpern, A.L.; Sutton, G.; Heidelberg, K.B.; Williamson, S.; Yooseph, S.; Wu, D.; Eisen, J.A.; Hoffman, J.M.; Remington, K.; et al. The Sorcerer II Global Ocean Sampling Expedition: Northwest Atlantic through Eastern Tropical Pacific. PLoS Biol. 2007, 5, e77. [Google Scholar] [CrossRef] [PubMed]
- Biers, E.J.; Sun, S.; Howard, E.C. Prokaryotic Genomes and Diversity in Surface Ocean Waters: Interrogating the Global Ocean Sampling Metagenome. Appl. Environ. Microbiol. 2009, 75, 2221–2229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signori, C.N.; Thomas, F.; Enrich-Prast, A.; Pollery, R.C.G.; Sievert, S.M. Microbial Diversity and Community Structure across Environmental Gradients in Bransfield Strait, Western Antarctic Peninsula. Front. Microbiol. 2014, 5, 647. [Google Scholar] [CrossRef] [PubMed]
- Luria, C.M.; Ducklow, H.W.; Amaral-Zettler, L.A. Marine Bacterial, Archaeal and Eukaryotic Diversity and Community Structure on the Continental Shelf of the Western Antarctic Peninsula. Aquat. Microb. Ecol. 2014, 73, 107–121. [Google Scholar] [CrossRef]
- Brown, M.V.; Philip, G.K.; Bunge, J.A.; Smith, M.C.; Bissett, A.; Lauro, F.M.; Fuhrman, J.A.; Donachie, S.P. Microbial Community Structure in the North Pacific Ocean. ISME J. 2009, 3, 1374–1386. [Google Scholar] [CrossRef] [Green Version]
- Gárate -Lizárraga, I.; Sevilla -Torres, G.; Álvarez -Añorve, M.; Aguirre -Bahena, F.; Violante -González, J.; Rojas -Herrera, A. First Record of a Red Tide Caused by Gyrodinium Instriatum (Dinophyceae: Gymnodiniales) in Bahía de Acapulco, Guerrero. CICIMAR Oceánides 2013, 28, 43–47. [Google Scholar] [CrossRef]
- Teo, C.L.; Jamaluddin, H.; Zain, N.A.M.; Idris, A. Biodiesel Production via Lipase Catalysed Transesterification of Microalgae Lipids from Tetraselmis Sp. Renew. Energy 2014, 68, 1–5. [Google Scholar] [CrossRef]
- Chappell, P.; Whitney, L.; Haddock, T.; Menden-Deuer, S.; Roy, E.; Wells, M.; Jenkins, B. Thalassiosira Spp. Community Composition Shifts in Response to Chemical and Physical Forcing in the Northeast Pacific Ocean. Front. Microbiol. 2013, 4, 273. [Google Scholar] [CrossRef] [Green Version]
- Scofield, V.; Jacques, S.M.S.; Guimarães, J.R.D.; Farjalla, V.F. Potential Changes in Bacterial Metabolism Associated with Increased Water Temperature and Nutrient Inputs in Tropical Humic Lagoons. Front. Microbiol. 2015, 6, 310. [Google Scholar] [CrossRef] [Green Version]
- Krause, E.; Wichels, A.; Giménez, L.; Lunau, M.; Schilhabel, M.B.; Gerdts, G. Small Changes in PH Have Direct Effects on Marine Bacterial Community Composition: A Microcosm Approach. PLoS ONE 2012, 7, e47035. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.S.; Kumar, A.; Rai, A.N.; Singh, D.P. Cyanobacteria: A Precious Bio-Resource in Agriculture, Ecosystem, and Environmental Sustainability. Front. Microbiol. 2016, 7, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajpai, V.K.; Shukla, S.; Kang, S.-M.; Hwang, S.K.; Song, X.; Huh, Y.S.; Han, Y.-K. Developments of Cyanobacteria for Nano-Marine Drugs: Relevance of Nanoformulations in Cancer Therapies. Mar. Drugs 2018, 16, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zadjelovic, V.; Chhun, A.; Quareshy, M.; Silvano, E.; Hernandez-Fernaud, J.R.; Aguilo-Ferretjans, M.M.; Bosch, R.; Dorador, C.; Gibson, M.I.; Christie-Oleza, J.A. Beyond Oil Degradation: Enzymatic Potential of Alcanivorax to Degrade Natural and Synthetic Polyesters. Environ. Microbiol. 2020, 22, 1356–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoniou, E.; Fodelianakis, S.; Korkakaki, E.; Kalogerakis, N. Biosurfactant Production from Marine Hydrocarbon-Degrading Consortia and Pure Bacterial Strains Using Crude Oil as Carbon Source. Front. Microbiol. 2015, 6, 274. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Marchica lagoon, North East of Morocco (N 35.11562, W 2.52803), (Image GeoEye from Google Earth, 2022).
Figure 1.
Marchica lagoon, North East of Morocco (N 35.11562, W 2.52803), (Image GeoEye from Google Earth, 2022).

Figure 2.
Oualidia lagoon, North West of Morocco (N 32.74675, W 9.036667), (Image GeoEye from Google Earth, 2022).
Figure 2.
Oualidia lagoon, North West of Morocco (N 32.74675, W 9.036667), (Image GeoEye from Google Earth, 2022).

Figure 6.
Proportion of reads mapping to viral contigs in OSD metagenomes. Legend: OSD24: Marchica lagoon.
Figure 6.
Proportion of reads mapping to viral contigs in OSD metagenomes. Legend: OSD24: Marchica lagoon.

Figure 7.
Heatmap of viral sequences abundance (when covered at >75%) across OSD samples. Only metagenomes with at least one viral sequence covered >75% were included.
Figure 7.
Heatmap of viral sequences abundance (when covered at >75%) across OSD samples. Only metagenomes with at least one viral sequence covered >75% were included.

Figure 8.
Functional category hits distribution annotated using MG-RAST Subsystems classification in Marchica lagoon (OSD24).
Figure 8.
Functional category hits distribution annotated using MG-RAST Subsystems classification in Marchica lagoon (OSD24).

Figure 9.
Functional category hits distribution annotated using MG-RAST Subsystems classification in Oualidia lagoon (OSD91).
Figure 9.
Functional category hits distribution annotated using MG-RAST Subsystems classification in Oualidia lagoon (OSD91).

Figure 10.
Top functional genes abundance in Marchica using Prokka [24].
Figure 10.
Top functional genes abundance in Marchica using Prokka [24].

Figure 11.
Top functional genes abundance in Oualidia using Prokka [24].
Figure 11.
Top functional genes abundance in Oualidia using Prokka [24].

Figure 12.
Correlation between functions and physicochemical parameters.

Table 1.
Physical and chemical data measured in waters of Marchica and Oualidia lagoons at the time of sampling.
Table 1.
Physical and chemical data measured in waters of Marchica and Oualidia lagoons at the time of sampling.
| Units | Marchica | Oualidia | ||||
|---|---|---|---|---|---|---|
| 2014 | 2015 | 2014 | 2015 | |||
| Parameters | Temperature | Celsius | 26.5 | 27.2 | 21 | 20 |
| Salinity | ppt | 35.98 | 35.96 | 27.24 | 29.41 | |
| Electrical conductivity | mS | 53.7 | 54.2 | 39.6 | 45.1 | |
| pH | <7.6 to 8.4> | 8.16 | 8.74 | 8.36 | 8.16 | |
| Dissolved Oxygen | mg/L | 8.33 | 8.50 | 7 | 7.5 | |
| Phosphate | mg/L | 0.03 | 0.04 | 0 | 0 | |
| Turbidity | NTU | 3.62 | 3.60 | 3 | 3.2 | |
| Nitrates | mg/L | 12.5 | 4.8 | 10.79 | 2.68 | |
| Nitrites | mg/L | <0.3 | 0.11 | <0.001 | <0.05 | |
Table 6.
16S metagenome sequences assigned to a Strain.
| Oualidia Lagoon | Marchica Lagoon | ||||||
|---|---|---|---|---|---|---|---|
| Taxa | Genus | Species | Strain | #Hits (bp) | % | #Hits (bp) | % |
| Alphaproteobacteria | Anaplasma | marginale | Maries | 3,521,109 | 8.28 | 2,149,110 | 2.21 |
| Alphaproteobacteria | Rickettsia | conorii | Malish 7 | 3,079,827 | 7.24 | 1,016,387 | 10.49 |
| Alphaproteobacteria | Anaplasma | phagocytophilum | HZ | 1,683,742 | 3.96 | 3,623,917 | 3.74 |
| Gammaproteobacteria | Xylella | fastidiosa | Temecula1 | 1,501,655 | 3.53 | 3,857,299 | 3.98 |
| Gammaproteobacteria | Xenorhabdus | nematophila | 1_9_0_6_1 | 1,117,290 | 2.62 | 4,068,450 | 4.19 |
| Alphaproteobacteria | Orientia | tsutsugamushi | Boryong | 829,248 | 1.95 | 3,639,647 | 3.75 |
| Gammaproteobacteria | Escherichia | colia | UMN026 | 490,326 | 1.15 | 1,533,000 | 1.58 |
| Betaproteobacteria | Burkholderia | rhizoxinica | 4_5_4 | 430,554 | 1.01 | 1,112,721 | 1.15 |
| Alphaproteobacteria | Brucella | canis | 2_3_3_6_5 | 283,501 | 0.66 | 592,700 | 0.61 |
| Alphaproteobacteria | Erythrobacter | litoralis | HTCC2594 | 277,397 | 0.65 | 725,363 | 0.75 |
| Betaproteobacteria | Ralstonia | solanacearum | PSI07 | 241,328 | 0.56 | 614,227 | 0.63 |
| Alphaproteobacteria | Rhizobium | sp | N_G_R_2_3_4 | 217,740 | 0.51 | 305,658 | 0.31 |
| Gammaproteobacteria | Shigella | dysenteriae | Sd197 | 124,696 | 0.29 | 346,084 | 0.35 |
| Gammaproteobacteria | Marinobacter | aquaeolei | VT8 | 664,804 | 1.56 | 2,030,270 | 2.09 |
| Cyanobacteria | Prochlorococcus | marinus | 9_3_1_3 | 1,387,027 | 0.3 | 3,261,370 | 3.36 |
| Cyanobacteria | Nostoc | azollae | 0_7_0_8 | 1,340,033 | 3.15 | 5,328,515 | 5.5 |
| Cyanobacteria | Synechococcus | sp | C_C_9_6_0_5 | 406,828 | 0.95 | 3,636,034 | 3.75 |
| Cyanobacteria | Synechococcus | sp | JA_2_1_3 | 287,486 | 0.67 | 1,310,559 | 1.35 |
| Bacteroidetes | Candidatus -Amoebophilus | asiaticus | 5_a_2 | 2,748,208 | 6.46 | 10,026,340 | 10.34 |
| Bacteroidetes | Bacteroides | fragilis | 9_3_4_3 | 956,572 | 2.25 | 2,783,907 | 2.87 |
| Verrucomicrobia | Methylacidiphilum | infernorum | V4 | 655,181 | 1.54 | 1,576,020 | 1.62 |
| Verrucomicrobia | Akkermansia | muciniphila | 8_3_5 | 517,529 | 1.21 | 571,583 | 0.58 |
| Actinobacteria | Rothia | dentocariosa | 1_7_9_3_1 | 2,513,106 | 5.91 | 3,917,261 | 4.04 |
| Actinobacteria | Mycobacterium | leprae | TN | 1,072,610 | 2.52 | 1,013,872 | 1.04 |
| Actinobacteria | Tropheryma | whipplei | 2_7 | 1,010,398 | 2.37 | 1,929,685 | 1.99 |
| Actinobacteria | Rhodococcus | jostii | RHA1 | 199,761 | 0.47 | 277,060 | 0.28 |
| Actinobacteria | Catenulispora | acidiphila | 4_4_9_2_8 | 194,558 | 0.45 | 614,181 | 0.63 |
Table 7.
18S SSU rRNA amplicons from selected high abundant sequences assigned to a phylum for both sampling campaigns 2014 and 2015 in both lagoons.
Table 7.
18S SSU rRNA amplicons from selected high abundant sequences assigned to a phylum for both sampling campaigns 2014 and 2015 in both lagoons.
| Marchica Lagoon | Oualidia Lagoon | |||||||
|---|---|---|---|---|---|---|---|---|
| 2014 | 2015 | 2014 | 2015 | |||||
| Taxa | #Hits | % | #Hits | % | #Hits | % | #Hits | % |
| Dinophyta | 4233 | 37.7 | 22,505 | 62.09 | 7617 | 22.7 | 1723 | 5.4 |
| Ochrophyta | 2725 | 24.2 | 6128 | 16.9 | 10,848 | 32.4 | 13,302 | 42.2 |
| Chlorophyta | 777 | 6.9 | 939 | 2.5 | 1827 | 5.4 | 6806 | 21.6 |
Table 8.
18S metagenomic rRNA from selected high abundant sequences assigned to a genus for both sampling campaigns 2014 and 2015 in both lagoons.
Table 8.
18S metagenomic rRNA from selected high abundant sequences assigned to a genus for both sampling campaigns 2014 and 2015 in both lagoons.
| Marchica Lagoon | Oualidia Lagoon | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2014 | 2015 | 2014 | 2015 | ||||||||
| Taxa | Genus | #Hits % | #Hits % | Taxa | Genus | #Hits | %#Hits % | ||||
| Dinophyta | Gyrodinium | 2844 | 25.3 | 12,160 | 33.5 | Dinophyta | Pelagodinium | 4051 | 12.1 | 20 | 0.06 |
| Ochrophyta | Pseudo-Nitzschia | 1776 | 15.8 | 752 | 2 | Ochrophyta | Thalassiosira | 2862 | 8.5 | 7979 | 25.3 |
| Chlorophyta | Tetraselmis | 503 | 4.4 | 61 | 0.1 | Chlorophyta | Ostreococcus | 8 | 0.02 | 5240 | 16.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chaouni, B.; Idrissi Azami, A.; Essayeh, S.; Arrafiqui, E.H.; Bailal, A.; Raoui, S.; Amzazi, S.; Twaddle, A.; El Hamouti, C.; Boukhatem, N.; et al. Moroccan Lagoon Microbiomes. Water 2022, 14, 1715. https://doi.org/10.3390/w14111715
AMA Style
Chaouni B, Idrissi Azami A, Essayeh S, Arrafiqui EH, Bailal A, Raoui S, Amzazi S, Twaddle A, El Hamouti C, Boukhatem N, et al. Moroccan Lagoon Microbiomes. Water. 2022; 14(11):1715. https://doi.org/10.3390/w14111715
Chicago/Turabian StyleChaouni, Bouchra, Abdellah Idrissi Azami, Soumaya Essayeh, El Houcine Arrafiqui, Abdelhakim Bailal, Sanae Raoui, Saaïd Amzazi, Alan Twaddle, Chahrazade El Hamouti, Noureddine Boukhatem, and et al. 2022. "Moroccan Lagoon Microbiomes" Water 14, no. 11: 1715. https://doi.org/10.3390/w14111715
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.