
Isolation of Bacteria Capable of Degrading Various AHLs for Biofouling Control in Membrane Bioreactors

1
Department of Water Resources and Environmental Engineering, Tamkang University, 151 Yingzhuan Road, Tamsui District, New Taipei City 25137, Taiwan
2
Water Treatment Science and Technology Research Center, Tamkang University, 151 Yingzhuan Road, Tamsui District, New Taipei City 25137, Taiwan
*
Author to whom correspondence should be addressed.
Water 2022, 14(11), 1712; https://doi.org/10.3390/w14111712
Submission received: 19 March 2022
/
Revised: 19 May 2022
/
Accepted: 25 May 2022
/
Published: 26 May 2022
(This article belongs to the Special Issue Contaminants of Emerging Concern in the Urban Water Cycle: Fate, Occurrence, Detection, Monitoring, and Control)
Abstract
:Membrane bioreactors (MBRs) are widely used to treat wastewater, mainly due to the production of high-quality effluent. However, biofilm forming on the surface of membranes can cause many problems, which remains one of the major limitations of this technique. Bacterial quorum quenching (QQ) has been proven to be a successful strategy to control biofouling in MBRs. However, for many QQ bacterial isolates, the detailed degradation rates of acyl homoserine lactones (AHLs) have rarely been reported. Therefore, this study aimed to isolate potential QQ bacteria and investigate their degradation rates against eight different AHLs. Results showed that four isolates (A9, A12, B11, and D3) exhibited consistent C8-HSL–(N-octanoyl-L-homoserine lactone) removal capabilities. These four isolates removed at least 70% of all AHLs tested within 180 min. They might have different QQ enzymes, based on our observation that the locations of enzyme activities differed. The bacteria most closely related to A9, A12, and B11 were Brucella anthropic, Bacillus cereus, and Bacillus toyonensis, respectively. Bacillus species have shown QQ activity in many studies, but AHL-reducing Brucella species have not been previously reported. Overall, this study extends our current knowledge of QQ bacteria that could be used to mitigate biofilm formation on MBR membranes.
1. Introduction
Membrane bioreactors (MBRs) are widely used in municipal and industrial wastewater treatment facilities due to several advantages, such as the production of high-quality effluent, small footprint, etc. In an MBR, a biological activated sludge process is combined with a membrane filtration unit. As a result, the membrane is prone to biofouling, which is caused by the formation of a biofilm on the surface of the membrane. Many techniques have been developed to mitigate biofouling in MBRs [1,2,3,4,5,6,7,8]. Among these techniques, bacterial quorum quenching (QQ) has been demonstrated to be an effective control strategy for suppressing the accumulation of transmembrane pressure during membrane filtration.
Most QQ bacteria used in MBRs degrade the signaling molecules of quorum sensing systems (QSs), which coordinate various synchronized bacterial group behaviors, including biofilm formation. Acyl homoserine lactones (AHLs) belong to one category of signaling molecules and are utilized by Gram-negative bacteria. Various and diverse QQ bacteria have been isolated [8,9,10,11,12,13,14,15]. Among these QQ bacteria, Rhodococcus sp. BH4 has been used in MBRs, and its ability to mitigate biofouling has been extensively investigated [8,16,17,18,19,20]. The degradation of C4-HSL, C6-HSL, C6-oxo-HSL, C8-HLS, and C12-oxo-HSL by Enterococcus sp. HEMM-1 was assessed using endpoint measurement at 2 h [21]. The QQ activity of HEMM-1 was better concerning the AHLs with shorter acyl chains. QS-inhibitory activity was 57% for C4-HSL, 19% for C6-HSL, 9% for C6-oxo-HSL, 10% for C8-HLS, and 4% for C12-oxo-HSL. Gu et al. [22] reported a QQ bacterium, Acinetobacter bereziniae strain, based on its removal rates of C6-HSL and C8-HSL in a cell suspension format and in microbial polyvinylidene difluoride (PVDF) bags. The authors found that the degradation rate of the Acinetobacter bereziniae strain in the microbial bag against C6-HSL and C8-HSL was 88% and 69%, respectively. The QQ ability of Bacillus sp. T5 and Delftia lacustris T6 was evaluated according to the inhibition of C8-HSL activity to the exogenic AHL reporter strain Agrobacterium tumefaciens A136 [23]. For the QQ bacteria, their AHL-degrading enzymes were identified and classified into three types: lactonase [14], acylase [13], and AHL oxidase/reductase [24].
Although many studies have aimed to isolate QQ bacteria for use in biofouling control in MBRs, detailed degradation rates of a range of AHLs have not been clearly reported. Since different AHL molecules might be used by different microbial communities in activated sludge, it may be more appropriate to use certain strains of QQ bacteria in specific systems. Therefore, the objectives of this study are to isolate potential QQ bacteria and to investigate the degradation rates of these bacteria against eight different AHLs. The results of this study could extend our current knowledge of QQ bacteria that are used to mitigate biofouling in MBRs.
2. Materials and Methods
2.1. Isolation of AHL-Degrading Bacteria
Samples collected from activated sludge tanks from a municipal wastewater treatment plant and campus drainage were used as sources for isolating potential AHL-degrading bacteria. The samples were serially diluted in sterile water and then 100 μL of the dilution was plated on Luria broth (LB) agar. After incubation at 30 °C for 24 h, colonies that had formed on the LB plates were picked from the agar and cultured in LB broth at 30 °C for 24–48 h. To screen potential AHL-degrading bacteria, 100 μL of the bacterial suspension was transferred into 100 μL of fresh LB broth with 10 μM of C8-HSL (N-octanoyl-L-homoserine lactone). After incubation overnight at 30 °C, 20 μL of supernatant was harvested after centrifugation at 8000× g for 10 min and transferred to 96-well plates containing 100 μL of LB broth, Agrobacterium tumefaciens A136 (Ti−) (pCF218) (pCF372) (OD600 ≥ 0.5), and X-Gal (20 mg/L). The plate was placed in an oven maintained at 30 °C and subjected to gentle shaking overnight. After incubation, supernatants containing residual C8-HSL would turn blue because of indigo produced from the hydrolysis of X-Gal, which was catalyzed by the enzyme produced by the reporter strain A136. In contrast, supernatants not containing C8-HSL remained the yellow color of LB broth. In addition, a QQ bacterium, Rhodococcus sp. BH4, was used as a positive control. Autoclave-killed BH4 and water were used as negative controls. When colonies showed the ability to degrade C8-HSL, the procedure was repeated two to three times to confirm this ability. Before the last confirmation test, four potential isolates (named A9, A12, B11, and D3) were further purified by plating on LB agar. The consistent appearance of the colonies (shape, color, size, etc.) was examined.
2.2. Assessing Degradation Efficiency of Isolated Bacteria against Various AHL Molecules
The degradation efficiency of the four isolated bacteria against different AHL molecules was evaluated in batch tests. Cell suspensions of the isolates (500 μL) were inoculated in 20 mL of fresh LB broth in 50 mL centrifuge tubes and cultured at 30 °C overnight in an orbital incubator. Relative cell density was estimated using an OD600 value measured by a spectrophotometer. The OD600 values of all the cell suspensions were then adjusted to the lowest one, to ensure that the initial cell concentrations were approximately similar (Table S1). Ten milliliters of cell suspension was centrifuged at 8000× g for 10 min. The supernatant was discarded and the cell pellet was then resuspended in 10 mL of fresh LB broth containing 100 nM of the indicated AHL molecules and 50 mM of PIPES (a pH buffer) in 50 mL centrifuge tubes. Additionally, water (designated H2O) and A12 bacteria killed by microwaving (designated DC) were used as negative controls to assess the fraction removed by nonbiological processes, such as AHL adsorption on the surfaces of cells or tubes. The batch tests were conducted in an incubator at 30 °C for 180 min with agitation at 120 rpm. Samples (200 μL) collected at 0, 30, 60, 120, and 180 min were centrifuged at 8000× g for 5 min, and the supernatants obtained were stored at −20 °C before analysis for residual AHL concentrations.
The AHLs used in the batch tests were C6-HSL, C8-HSL, C10-HSL, C12-HSL, 3-oxo-C6-HSL, 3-oxo-C8-HSL, 3-oxo-C10-HSL, and 3-oxo-C12-HSL (Sigma–Aldrich); please refer to Table S2 for the formal names, chemical formulae, and chemical abstract service (CAS) numbers of these AHLs. Concentrations of AHLs were determined using the reporter bacterial strain A. tumefaciens A136 and the Beta-Glo Assay System (Promega) [16]. In brief, A136 cultured overnight (OD600 > 0.5) was diluted with fresh LB broth to an adjusted cell concentration such that it had an OD600 value of approximately 0.1. In each well of the microwell plate, 90 μL of A136 and 10 μL of either the standard or a sample was mixed. The microwell plate was then covered with a sealing film and incubated at 30 °C for 1.5 h. After removing the sealing film, 30 μL of Beta-Glo reagent was added into each well. The plate was then covered with aluminum foil and placed in a dark at room temperature for 40 min. Luminescence produced in wells was measured in a microplate reader (Synergy HTX, BioTek, Inc., Taipei, Taiwan). The strength of luminescence signals was correlated with known concentrations of AHL to construct standard curves. For each sample, three analytical replicates were performed.
2.3. Tests for AHL-Degradation Enzymes
To evaluate whether the AHLs were actually degraded by the enzymes or if they remained intact but the biological pathway in the bioassay was inhibited or interfered with by other unknown small molecules secreted by the isolates, heat-inactivated methods were used to provide evidence to distinguish between these two possible mechanisms. The heat-inactivated methods were autoclaving (20 min at 121 °C and 15 psi) and microwaving (700 W for 5 min). In addition, extracellular and intracellular enzymes responsible for the degradation of AHLs were assessed. The spent media obtained in this test were generated by centrifugation (8000× g for 10 min) or by filtration using sterile PES syringe filters with a 0.2 μm pore size (Acrodisc PN 4612, Pall Corporation). After centrifugation, the supernatant was collected. The cell pellet at the bottom of the tube was washed to remove residual liquid. Ten microliters of sterile PBS solution (8 g/L of NaCl, 0.2 g/L of KCl, 1.44 g/L of Na2HPO4, and 0.245 g/L KH2PO4) was added into the tubes and the cell pellet was resuspended by vortexing for 30 s. The suspension was washed three times with PBS solution. Tubes containing a cell pellet were stored overnight at −20 °C to break the bacterial cell envelope. Finally, after equilibration with room temperature, the tubes were centrifuged at 8000× g for 10 min to harvest the cell lysate for the test.
2.4. Membrane Bioreactors
In this study, two lab-scale MBRs were operated in parallel to evaluate the performance of the potential QQ bacterial strain A12 in terms of the accumulation rate of transmembrane pressure. The A12 bacteria were entrapped in polyvinyl alcohol (PVA, 10%) alginate (1%) beads at an estimated dose of about 45.6 mg (dry weight) per liter of reactor volume. One reactor contained QQ beads and the other was run as a control. The reactor volume is 2 L. Hydraulic and sludge retention times are 8 h and 60 days, respectively. ZeeWeed 500D hollow fiber membrane (PVDF, pore size < 0.04 μm) was used and manually fabricated to create a small filtration module. When the TMP reached 30 kPa, it was considered the end of the test, and new membrane modules were installed for the next run. In total, three runs were conducted. The surface area of the hollow-fiber membrane in this module is 98 cm2. The flux was set at 25.5 L/m2/h (LMH) and the aeration rate under the membrane was 1 L/min. Synthetic wastewater containing 120 mg/L of glucose, 90 mg/L of peptone, 12 mg/L of yeast extract, 96 mg/L of (NH4)2SO4, 17 mg/L of KH2PO4, 300 mg/L of NaHCO3, 2.4 mg/L of CaCl2·2H2O, 24 mg/L of MgSO4·7H2O, 2.16 mg/L of MnSO4·5H2O, and 0.12 mg/L of FeCl3·6H2O was fed into the reactors using peristaltic pumps controlled by water-level sensors. Transmembrane pressure was monitored by pressure transducers and data loggers. Concentrations of COD, NH4+-N, NO2−-N, and NO3−-N were also monitored to evaluate the performance of the MBRs. Standard methods described by the Environmental Analysis Laboratory of Taiwan Environmental Protection Administration were used to analyze COD (NIEA 517.53B, closed reflux, titrimetric method), NH4+-N (NIEA W448.51B, phenate method), NO2−-N (NIEA W418.53C, colorimetric method), and NO3−-N (NIEA W419.51A, ultraviolet spectrophotometric method). Concentrations of extracellular polymeric substances (EPS) and soluble microbial products (SMP) in reactors were also processed [25] and measured [26,27].
2.5. Analysis of 16S rDNA of Isolates
Bacterial colonies of A9, A12, B11, and D3 that formed on the LB plates after overnight cultivation at 30 °C were sent to the nucleotide-sequencing service provider Genomics (Taipei, Taiwan) for sanger sequencing. A universal primer set 27F and 1525R [28] was selected to generate a near-full-length 16S rRNA gene sequence. Identification of the nucleobases was conducted via automated DNA sequencing, but assignment of the bases was manually conducted using the software Chromas v2.6.6. After ambiguous bases had been trimmed, the forward and reverse reads were merged to generate partial 16S rRNA gene sequences with lengths between 1327 nt and 1397 nt (Table S3). These sequences were used as queries to search against the NCBI nucleotide collection (nt/nr) database using the nucleotide BLAST with default settings to identify the most closely related bacterial species.
3. Results
3.1. Isolation of Potential AHL-Degrading Bacteria
Activated sludge samples collected from a local wastewater treatment plant were used as one source to screen for potential AHL-degrading bacteria. As shown in Figure 1a, 35 out of 101 bacterial colonies (the results for five colonies are not shown in the figure) exhibited the ability to degrade C8-HSL. After the first confirmation of their degradation ability (Figure 1b), 12 out of these 35 bacterial colonies retained their ability to break down C8-HSL. Ultimately, three colonies consistently removed C8-HSL from the LB broth (Figure 1c) and were collected for subsequent examination. These three colonies were designated as A9, A12, and B11. In addition to activated sludge, water samples collected from campus drainage were also used as another source for screening for potential AHL-degrading bacteria (results not shown) using the procedure described above. One out of 86 bacterial colonies consistently degraded C8-HSL in two screening runs. This culture was then designated as D3. In all of the tests, a known AHL degrader, BH4, was used as a positive control. Autoclave-killed BH4 and water were used as negative controls. Obtaining the expected results from positive and negative controls ensured that the bioreactions were performed properly.
Before final confirmation of C8-HSL-degradation ability, the bacteria were further processed to ensure their purity. Isolates A9, A12, B11, and D3 were cultured in LB broth again overnight. The cell suspension was then serially diluted and then plated on LB agar. Consistent colony morphology, such as shape, color, size, etc., was observed. The colonies of each isolate on plates were serially diluted to form cell suspensions of different concentrations and examined again for their ability to degrade C8-HSL. The result is illustrated in Figure 2. All isolates (except one replicate of 106-fold diluted D3) removed 5 μM of C8-HSL to below the detectable level in the X-Gal bioassay. Meanwhile, the expected results were observed in both the positive control (BH4) and the negative controls (dead BH4 and water).
In total, after screening 187 colonies and running three sets of confirmation tests, four C8-HSL-degrading bacteria (A9, A12, B11, and D3) were obtained. Their degradation efficiency toward various types of AHL molecules was then evaluated (see the following section).
3.2. Degradation Efficiency of Isolated Bacteria against Various AHL Molecules
The AHL-degrading bacterial isolates A9, A12, B11, and D3 were cultured in the presence of eight different AHL-type signaling molecules to evaluate their potential to degrade various AHLs. AHLs with different acyl-side chain lengths and with or without the keto oxygen on the C3 carbon were used. These AHLs were C6-HSL, C8-HSL, C10-HSL, C12-HSL, 3-oxo-C6-HSL, 3-oxo-C8-HSL, 3-oxo-C10-HSL, and 3-oxo-C12-HSL.
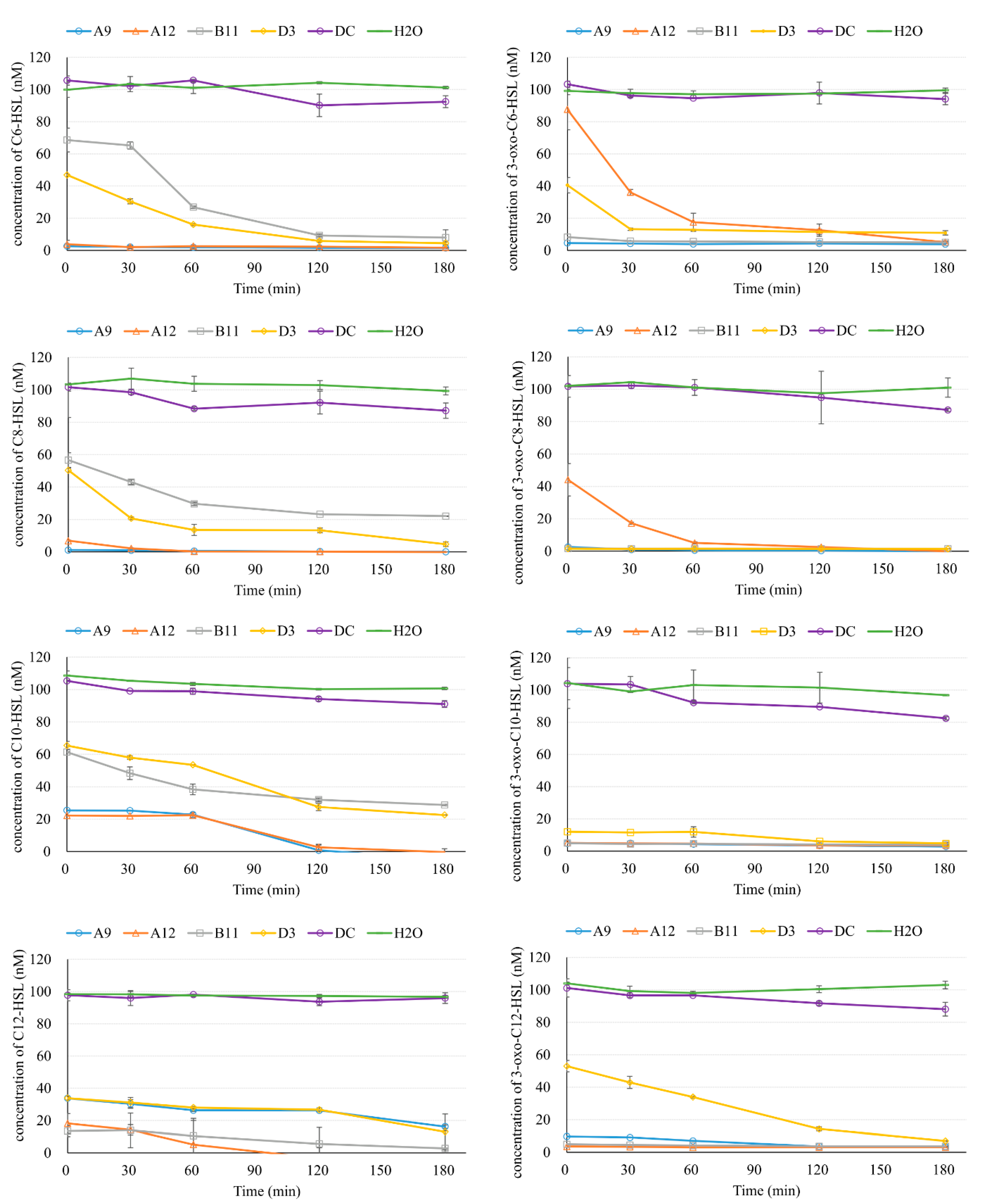
The degradation curves are illustrated in Figure 3. C6-HSL was rapidly consumed by A9 and A12; it had even been consumed by the first sampling point, when the AHL had just been mixed with the cell suspension (less than 1 min). C6-HSL was then maintained at under 4 nM throughout the rest of the test. Quick removal of C6-HSL was also observed for B11 and D3, but to a lesser extent. At the first sampling point (1 min), the concentration of C6-HSL was 68.6 nM and 46.9 nM for B11 and D3, respectively. More than 73% and 90% of the C6-HSL was degraded by 60 min and 120 min, respectively, for both B11 and D3. The removal curve of C8-HSL showed a similar trend to that of C6-HSL. A9 and A12 reduced C8-HSL to less than 7% at the first sampling point and almost completely removed C8-HSL by the end of the experiment. D3 was more effective than B11 in terms of degrading C8-HSL throughout the test. More than 20% of the C8-HSL was left (22.1 nM) for B11 at 180 min. C10-HSL was removed by all the isolates but with a slower rate than AHLs containing shorter side chains. For A9 and A12, C10-HSL was reduced to around 25% initially, and this amount remained constant for the first 60 min. After 120 min, the C10-HSL was reduced to less than 3 nM. For D3 and B11, >30% of the C10-HSL was degraded initially, but slower reduction rates were observed at 60 min than for C6-HSL and C8-HSL. For C12-HSL, A12 still showed the ability to quickly remove this compound (>80%) initially, and to reduce this molecule to extremely low levels after 120 min. In contrast, a slower removal rate of C12-HSL than for the other AHLs was observed for A9. In addition, B11 demonstrated a better degradation rate for C12-HSL than for AHLs containing shorter side chains.
The degradation of AHLs with a keto oxygen on the C3 carbon was also observed, showing that the four isolates are capable of degrading various types of AHLs (Figure 3). 3-oxo-C6-HSL was degraded very rapidly to a low level (<8.3 nM) by A9 and B11 from the beginning of the experiment. Fast degradation in the first 30 min was found for A12 and D3, but at a relatively slower rate. At the end of the test, 3-oxo-C6-HSL levels were at least below 11 nM. Better removal efficiency was observed in the test containing 3-oxo-C8-HSL. Concentrations of 3-oxo-C8-HSL were reduced to less than 3 nM in the first sample for all isolates except A12, which nonetheless still removed ~95% of this AHL by 60 min. In the test with signaling molecule 3-oxo-C10-HSL, all four isolates demonstrated an excellent ability to remove this compound rapidly. Only D3 exhibited a slightly slower degradation rate. A similar pattern was observed in the test of 3-oxo-C12-HSL: A9, A12, and B11 reduced 3-oxo-C12-HSL quickly, while D3 showed a significantly lower removal efficiency.
Since the lactone ring structure of AHLs is pH sensitive and their function as QS signaling molecules can be deactivated at higher pH values [29], pH values were monitored at the beginning and the end of the tests (Figure S1). Results showed that the variation in pH was less than ± 0.2. In addition, along with all the tests, water and sterile-A12 negative controls were also conducted (Figure 3). The concentrations of the AHLs in both controls were substantially higher than those in the tubes containing bacterial isolates. This evidence indicates that these compounds were likely removed through biological mechanisms. A noticeably higher AHL level in the water control than that in the killed-A12 control implies that nonbiological removal mechanisms exist, but the amount of AHL removed was much smaller and the effect of these mechanisms was negligible. The possible explanations are the adsorption of AHLs on the debris of dead cells and the existence of trace amounts of active enzymes even after the microwave treatment. Overall, this experiment demonstrated that all four bacterial isolates are capable of degrading a variety of AHL molecules, showing their potential to be used as QQ bacteria in MBRs to control biofilm formation and mitigate biofouling. Among the four isolates, A9 and A12 showed a better degradation efficiency than B11 and D3 in terms of degradation rate against the different AHLs tested.
3.3. Preliminary Tests of AHL-Degrading Enzymes
As shown in Figure 4a, wells receiving both autoclave- and microwave-inactivated spent supernatant (extracellular fraction) or cell lysate (intracellular fraction) turned blue, indicating that the C8-HSL was not degraded after the heat treatment. In the wells inoculated with active intact cell suspensions, the C8-HSL was reduced to lower levels, as indicated by the color yellow (original media) or light blue. The outcomes of the positive control (in the presence of C8-HSL) and the negative control (in the absence of C8-HSL) were both as expected. This result supports an enzyme-involved mechanism over inhibition of or interference with bioassays by unknown small molecules.
To further investigate whether the AHL-degrading enzymes in the isolates are extracellular or intracellular, supernatant collected by centrifugation or filtration and cell lysate obtained from freeze–thaw treatment were examined in the X-Gal bioassay (Figure 4b). The wells that received samples from overnight cultures of A9, A12, B11, or D3 with 5 μM of C8-HSL in the medium showed the color yellow or relatively light blue. This result indicated the reduction of C8-HSL during the growth of all four isolates. When using supernatant harvested using centrifugation, A9, B11, and D3 showed better C8-HSL-removal efficiency than A12. In contrast, when the supernatant was acquired by filtration, A12 exhibited the best C8-HSL reduction of the isolates. This result implies that at least some of the degrading enzymes are extracellular and that these enzymes might be intercepted by adsorption onto the filter membrane during the filtration process. For A12, better C8-HSL removal by the supernatant produced by filtration than that produced by centrifugation was observed. The actual reason for this is unknown, but there might be inhibitors that coexist with the AHL-degrading enzymes, and these inhibitors could be removed by the filtration membrane. However, this hypothesis requires future examination. For the cell lysate generated by freeze–thaw treatment, all isolates exhibited removal of C8-HSL, but to different degrees. A9 and A12 showed the best reduction, followed by D3 and B11. The negative control remained the color of the culture medium, as expected. In summary, the above results suggest that some AHL-degrading enzymes could be secreted from the cells (extracellular form) in all four isolates, while some enzymes were found inside the cells. From the current results, it is unclear whether these enzymes were produced to be transported to outside the cells (extracellular), or in an intracellular form (membrane-bound or freely suspended in the cytoplasm), or both.
3.4. Using the Isolated Potential Quorum Quenching Bacterium A12 for Biofouling Control in a Membrane Bioreactor
Bacterium A12 was selected and encapsulated within PVA-sodium alginate hydrogel beads to evaluate its biofouling-control capacity in a lab-scale membrane bioreactor (Figure 5). During three cycles of tests, the MBR with A12 showed lower increase of transmembrane pressure compared to the control MBR. Using 30 kPa as the end of each test, the fouling rates in the MBR with A12 were reduced by 64.7%, 111.6%, and 96.4% in the three cycles, respectively. Meanwhile, the removal efficiency of COD and NH4+-N and the concentrations of NO2−-N and NO3−-N were monitored in both reactors. Despite that the number of samples for MBR performance monitoring is rather low, the results (Table 1) did not show great differences between the two reactors, suggesting that the A12 bacterium does not seem to have significant observable negative effects on the system. Comparing the EPS and SMP obtained from the two MBRs (Table 1), protein and polysaccharide fraction of both SMP and EPS levels were similar. Only SMP protein was significantly higher in MBR with A12 (p = 0.02). Based on the correlation analysis of MBR fouling and other biological characteristics including EPS and SMP [30], SMP strongly correlated to fouling rate compared to EPS. The authors of this study suggested that soluble biopolymers presented in bulk liquid can directly deposit onto the surface of the membrane; thus, they are more critical to membrane fouling than EPS. It is unclear why the MBR with lower fouling rates had a higher level of SMP protein in the current study. A recent study reported inhibition of the protease secretion by the QQ effect [31], which might explain the higher SMP protein concentrations found in the A12 QQ reactor. However, a systematic analysis that includes SRT, MLSS levels, and dominant microbial species in MBRs is required to provide a more comprehensive discussion of the observed results.
3.5. Nucleotide Sequences of the 16S rDNA of Isolated Bacteria
To determine the taxonomic classification of A9, A12, B11, and D3, the genomic DNA of the four bacterial isolates was extracted, followed by 16S rDNA amplification and sequencing (Table 1). The 16S rDNA of A9, A12, and B11, which originated from activated sludge in a wastewater treatment plant, was successfully sequenced, but the quality of 16S rDNA sequences for D3 was very low. The ambiguous sequencing results for D3 were probably obtained because this isolate was not yet a pure culture, despite several separation processes having been performed. Low-quality sequences located at both ends were manually trimmed, and the resulting 16S rDNA sequences for A9, A12, and B11 ranged from 1327 nt to 1397 nt. After these sequences were BLASTed against a nucleotide collection (nt/nr) database, the most closely related bacterial species to A9, A12, and B11 were Brucella anthropic, Bacillus cereus, and Bacillus toyonensis, respectively. The sequence identities were high (>99.77%). Among these species, Brucella anthropic is Gram-negative, while Brucella anthropic and Bacillus cereus are Gram-positive bacteria.
4. Discussion
In this study, 187 bacterial colonies (101 from activated sludge and 86 from campus drainage) were screened for potential QQ activity that could be used to control biofouling in MBRs. Some of the colonies showed inconsistent results, i.e., an isolate might degrade C8-HSL in one run but not in the next run during the confirmation tests (Figure 1). One reason is that the colonies that formed on the LB plates may not have been a pure isolate, and another reason is that some bacteria have both AHL-degrading and AHL-producing pathways [15,32]. In view of this phenomenon, several confirmation tests should be performed during the isolation process before the QQ bacteria can be considered validated. Ultimately, four potential QQ bacterial isolates were obtained.
These four isolates were then used to degrade AHL molecules with a variety of acyl side-chain lengths and in the presence or absence of the keto oxygen on the C3 carbon (Figure 3). Although they all showed positive and similar results, in that they removed 5 μM of C8-HSL after overnight incubation (Figure 1 and Figure 2), the degradation rates against different AHLs varied substantially (Figure 3). For example, A9 and A12 removed the selected AHLs more efficiently than B11 and D3, except in the cases of C12-HSL when treated with A9 and 3-oxo-C6-HSL and 3-oxo-C8-HSL when treated with A12. As opposed to the X-Gal bioassays, which measured only the endpoint concentrations of the AHLs, the degradation curves provide detailed information, especially for selecting the bacterial strains for future applications. However, this information is rarely reported in the literature. The degradation rates of the eight types of AHL by the most widely used QQ bacterium, Rhodococcus sp. BH4, have been examined [16]. Those results showed that 3-oxo-C12-HSL was reduced most efficiently, while the degradation rate of 3-oxo-C6-HSL was the lowest. From an initial 0.4 μM of 3-oxo-C6-HSL, only about 1% and 10% of this AHL was degraded at 3 min and 10 min, respectively. The AHL-degrading activity of another QQ bacterium, Pseudomonas sp. 1A1, was evaluated against eight kinds of AHL by Cheong et al. [15]. That study found that the degradation rate increased with the length of the acyl side chain (C6, C8, C10, and C12). The AHL with the longest acyl chain (C12-HSL) was removed most efficiently. For the four AHLs that contained an oxo-functional group at the C3 position (3-oxo-(C6, C8, C10, or C12)-HSL), the highest degradation rate was also observed for the AHL with the longest acyl chain, 3-oxo- C12-HSL. However, the reduction rates for the rest of the AHLs were less than 10% when measured at 10 min. In the present study, all eight of the tested AHLs were at least 70% removed within 180 min by all four isolates. The best initial degradation rates against all eight different AHLs (at least 80%) were produced by A9, which was effective even for the AHL with the keto oxygen on the C3 carbon.
The high initial degradation rates that were observed (Figure 3) raised the question of whether the AHLs were actually degraded enzymatically or remained intact but were inactivated by inhibitors secreted by the isolates that interfered with the biological pathway in the bioassay. The possibility of the presence of inhibitors has been reduced since C8-HSL treated with heat-inactivated spent supernatant (extracellular fraction) or cell lysate (intracellular fraction) could still induce β-galactosidase, as evidenced by the production of blue-indigo precipitation. Temperature-sensitive enzymes might be responsible for the observed reduction in AHLs in the degradation experiments. The high initial removal percentages of the AHLs could be explained by the inoculation with physiologically active cultures and/or abundant AHL-degrading enzymes being carried from the cultivation prior to the tests.
The location of the AHL-inactivating enzymes was also assessed in this study (Figure 4). A9 and D3 showed both extracellular and intracellular enzymatic activity. For A12, better intracellular QQ activity was observed. In contrast, higher external-enzyme activity was observed for B11. Two types of AHL-lactonases, encoded by the qsdA and jydB genes, have been identified in the cells of Rhodococcus sp. BH4 [16,33]. It has been reported that the lactonase of Rhodococcus sp. BH4 is intracellular, while the acylase of Pseudomonas sp. 1A1 is extracellular [15]. Most bacteria isolated from saltern, pond, and marine habitats exhibit much higher levels of intracellular than extracellular enzyme activity [23]. In the same study, the authors also demonstrated that most of the C8-HSL removal was the result of QQ enzymes belonging to the lactonase family, except for the QQ enzyme of Delftia lacustris T6, which was isolated from a saltern. QQ bacteria have been isolated from river sediments containing penicillin and from activated sludge treating penicillin-containing wastewater [34]. In these penicillin-containing environments, five out of the six isolates can degrade C10-HSL with their AHL-acylase activities, while one isolate might possess AHL-lactonase activity. These results imply that our four isolates may contain different QQ enzymes, because of our observation of dissimilar locations of their enzyme activities (Figure 4). Confirming the exact types of the QQ enzymes in our isolates will require further investigation.
A12 was applied to an MBR and the results showed that this reactor had a slower transmembrane pressure increase rate than the control MBR (Figure 5). This preliminary result suggests the potential of QQ effects for controlling biofouling in MBRs, but the long-term effects of the QQ bacteria require further examination. Comparing the 16S rDNA sequences in the NCBI nucleotide collection (nt/nr) database, the most closely related bacteria to A9, A12, and B11 were Brucella anthropic, Bacillus cereus, and Bacillus toyonensis, respectively (Table 2). Genus Brucella is in the class Alphaproteobacteria and the phylum Proteobacteria. To the best of our knowledge, QQ activity in Brucella has not been previously reported. QQ activities of Bacillus species have been identified in many studies. For example, Bacillus sp. strain 240B1 has an AHL-lactonase gene (aiiA) that can hydrolyze the lactone ring [14]. Bacillus thuringiensis subspecies aizawai, galleriae, kurstaki, kyushuensis, ostriniae, and subtoxicus were capable of degrading 3-oxo-C6-HSL [35]. Bacillus sp. T5, which also shows the highest identity with Bacillus cereus (99%), had the greatest AHL-removal capability of 19 isolated QQ bacteria [23]. In this study, among the four isolates exhibiting strong QQ activity, two (A12 and B11) are Gram-positive Bacillus species. It seems that a strong QQ capacity is widespread in Bacillus, although the actual role of QQ enzymes such as lactonase in this genus awaits future exploration.
5. Conclusions
To investigate the bacterial QQ technique for controlling biofouling in MBRs, this study aimed to isolate potential QQ bacteria and to investigate the degradation rates of these bacteria against eight different types of AHL. Of 187 bacterial colonies screened, the four isolates (A9, A12, B11, and D3) that showed consistent C8-HSL-removal capabilities were studied in detail. These four isolates can remove at least 70% of all AHLs within 180 min. Moreover, the best initial AHL degradation rates were observed for A9, which can remove more than 80% of AHLs, even the AHLs with the keto oxygen on the C3 carbon. A9, A12, B11, and D3 might have different QQ enzymes, as suggested by our observation of dissimilar locations of the enzyme activities in these isolates. The most closely related bacteria to A9, A12, and B11 were Brucella anthropic, Bacillus cereus, and Bacillus toyonensis, respectively. QQ activities in Bacillus species have been identified in many studies, while AHL reduction by Brucella species has not been previously reported. Preliminary tests showed a reduction in transmembrane pressure increase rate in an MBR treated with Bacillus sp. A12. In conclusion, results of this study extend our current knowledge of QQ bacteria that could be used to mitigate biofilm formation on filtering membranes in MBRs.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/w14111712/s1. Figure S1: pH monitored at the beginning and the end of the AHL-degradation tests. H2O: a negative control in which the cell suspension was replaced with water. DC: A dead-cell control, in which A12 killed by microwaving served as another negative control. Table S1: Concentration of cell suspensions of A9, A12, B11, and D3 in the AHL-degradation tests. Values shown are OD600. Table S2: Acyl homoserine lactones used in this study. Table S3: Nucleotide sequences of the 16S rRNA genes of bacterial isolates A9, A12, and B11.
Author Contributions
Conceptualization, I.-C.C.; methodology, I.-C.C., C.-Y.C. and C.-Y.H.; validation, I.-C.C. and C.-Y.C.; investigation, I.-C.C., C.-Y.C. and C.-Y.H.; data curation, C.-Y.C. and C.-Y.H.; writing—original draft preparation, I.-C.C.; writing—review and editing, I.-C.C.; visualization, I.-C.C. and C.-Y.C.; supervision, I.-C.C.; funding acquisition, I.-C.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Taiwan Ministry of Science and Technology (MOST 105-2218-E-032-006 and MOST 106-2221-E-032-011).
Data Availability Statement
Not applicable.
Acknowledgments
The kind provision of the bacteria Rhodococcus sp. BH4 and Agrobacterium tumefaciens A136 by Professor Kwang-Ho Choo’s lab (Kyungpook National University) is gratefully acknowledged.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Muthukumaran, S.; Kentish, S.; Lalchandani, S.; Ashokkumar, M.; Mawson, R.; Stevens, G.W.; Grieser, F. The optimisation of ultrasonic cleaning procedures for dairy fouled ultrafiltration membranes. Ultrason. Sonochem. 2005, 12, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Le-Clech, P.; Fane, A.; Leslie, G.; Childress, A. MBR focus: The operators’ perspective. Filtr. Sep. 2005, 42, 20–23. [Google Scholar] [CrossRef]
- Judd, S. The status of membrane bioreactor technology. Trends Biotechnol. 2008, 26, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Pasmore, M.; Todd, P.; Smith, S.; Baker, D.; Silverstein, J.; Coons, D.; Bowman, C.N. Effects of ultrafiltration membrane surface properties on Pseudomonas aeruginosa biofilm initiation for the purpose of reducing biofouling. J. Membr. Sci. 2001, 194, 15–32. [Google Scholar] [CrossRef]
- Mansouri, J.; Harrisson, S.; Chen, V. Strategies for controlling biofouling in membrane filtration systems: Challenges and opportunities. J. Mater. Chem. 2010, 20, 4567–4586. [Google Scholar] [CrossRef]
- Jiang, W.; Xia, S.; Liang, J.; Zhang, Z.; Hermanowicz, S.W. Effect of quorum quenching on the reactor performance, biofouling and biomass characteristics in membrane bioreactors. Water Res. 2013, 47, 187–196. [Google Scholar] [CrossRef]
- Kim, J.-H.; Choi, D.-C.; Yeon, K.-M.; Kim, S.-R.; Lee, C.-H. Enzyme-Immobilized Nanofiltration Membrane To Mitigate Biofouling Based on Quorum Quenching. Environ. Sci. Technol. 2011, 45, 1601–1607. [Google Scholar] [CrossRef]
- Oh, H.-S.; Yeon, K.-M.; Yang, C.-S.; Kim, S.-R.; Lee, C.-H.; Park, S.Y.; Han, J.Y.; Lee, J.-K. Control of Membrane Biofouling in MBR for Wastewater Treatment by Quorum Quenching Bacteria Encapsulated in Microporous Membrane. Environ. Sci. Technol. 2012, 46, 4877–4884. [Google Scholar] [CrossRef]
- Torres, M.; Rubio-Portillo, E.; Antón, J.; Ramos-Esplá, A.A.; Quesada, E.; Llamas, I. Selection of the N-Acylhomoserine Lactone-Degrading Bacterium Alteromonas stellipolaris PQQ-42 and of Its Potential for Biocontrol in Aquaculture. Front. Microbiol. 2016, 7, 646. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-W.; Gan, H.M.; Yin, W.-F.; Chan, K.-G. Genome Sequence of Roseomonas sp. Strain B5, a Quorum-Quenching N-Acylhomoserine Lactone-Degrading Bacterium Isolated from Malaysian Tropical Soil. J. Bacteriol. 2012, 194, 6681–6682. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.Z.; Morohoshi, T.; Someya, N.; Ikeda, T. AidC, a novel N-acylhomoserine lactonase from the potato root-associated cytophaga-flavobacteria-bacteroides (CFB) group bacterium Chryseobacterium sp. strain StRB126. Appl Environ. Microbiol. 2012, 78, 7985–7992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Tang, K.; Shi, X.; Zhang, X.H. Flaviramulus ichthyoenteri sp. nov., an N-acylhomoserine lactone-degrading bacterium isolated from the intestine of a flounder (Paralichthys olivaceus), and emended descriptions of the genus Flaviramulus and Flaviramulus basaltis. Int. J. Syst. Evol. Microbiol. 2013, 63, 4477–4483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leadbetter, J.R.; Greenberg, E.P. Metabolism of Acyl-Homoserine Lactone Quorum-Sensing Signals by Variovorax paradoxus. J. Bacteriol. 2000, 182, 6921–6926. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.H.; Xu, J.L.; Li, X.Z.; Zhang, L.H. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. USA 2000, 97, 3526–3531. [Google Scholar] [CrossRef] [PubMed]
- Cheong, W.-S.; Lee, C.-H.; Moon, Y.-H.; Oh, H.-S.; Kim, S.-R.; Lee, S.H.; Lee, C.-H.; Lee, J.-K. Isolation and Identification of Indigenous Quorum Quenching Bacteria, Pseudomonas sp. 1A1, for Biofouling Control in MBR. Ind. Eng. Chem. Res. 2013, 52, 10554–10560. [Google Scholar] [CrossRef]
- Oh, H.S.; Kim, S.R.; Cheong, W.S.; Lee, C.H.; Lee, J.K. Biofouling inhibition in MBR by Rhodococcus sp. BH4 isolated from real MBR plant. Appl. Microbiol. Biotechnol. 2013, 97, 10223–10231. [Google Scholar] [CrossRef]
- Kim, S.-R.; Oh, H.-S.; Jo, S.-J.; Yeon, K.-M.; Lee, C.-H.; Lim, D.-J.; Lee, C.-H.; Lee, J.-K. Biofouling Control with Bead-Entrapped Quorum Quenching Bacteria in Membrane Bioreactors: Physical and Biological Effects. Environ. Sci. Technol. 2013, 47, 836–842. [Google Scholar] [CrossRef]
- Lee, S.; Park, S.K.; Kwon, H.; Lee, S.H.; Lee, K.; Nahm, C.H.; Jo, S.J.; Oh, H.S.; Park, P.K.; Choo, K.H.; et al. Crossing the Border between Laboratory and Field: Bacterial Quorum Quenching for Anti-Biofouling Strategy in an MBR. Environ. Sci. Technol. 2016, 50, 1788–1795. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, S.; Lee, K.; Nahm, C.H.; Kwon, H.; Oh, H.-S.; Won, Y.-J.; Choo, K.-H.; Lee, C.-H.; Park, P.-K. More Efficient Media Design for Enhanced Biofouling Control in a Membrane Bioreactor: Quorum Quenching Bacteria Entrapping Hollow Cylinder. Environ. Sci. Technol. 2016, 50, 8596–8604. [Google Scholar] [CrossRef]
- Kim, S.-R.; Lee, K.-B.; Kim, J.-E.; Won, Y.-J.; Yeon, K.-M.; Lee, C.-H.; Lim, D.-J. Macroencapsulation of quorum quenching bacteria by polymeric membrane layer and its application to MBR for biofouling control. J. Membr. Sci. 2015, 473, 109–117. [Google Scholar] [CrossRef]
- Ham, S.-Y.; Kim, H.-S.; Cha, E.; Park, J.-H.; Park, H.-D. Mitigation of membrane biofouling by a quorum quenching bacterium for membrane bioreactors. Bioresour. Technol. 2018, 258, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Huang, J.; Zeng, G.; Shi, Y.; Hu, Y.; Tang, B.; Zhou, J.; Xu, W.; Shi, L. Quorum quenching activity of indigenous quorum quenching bacteria and its potential application in mitigation of membrane biofouling. J. Chem. Technol. Biotechnol. 2018, 93, 1394–1400. [Google Scholar] [CrossRef]
- Yavuztürk Gül, B.; Koyuncu, I. Assessment of new environmental quorum quenching bacteria as a solution for membrane biofouling. Process Biochem. 2017, 61, 137–146. [Google Scholar] [CrossRef]
- Uroz, S.; Chhabra, S.R.; Cámara, M.; Williams, P.; Oger, P.; Dessaux, Y. N-Acylhomoserine lactone quorum-sensing molecules are modified and degraded by Rhodococcus erythropolis W2 by both amidolytic and novel oxidoreductase activities. Microbiology 2005, 151, 3313–3322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Cao, B.; Wang, D.; Ma, T.; Xia, H.; Yu, D. Influence of wastewater sludge treatment using combined peroxyacetic acid oxidation and inorganic coagulants re-flocculation on characteristics of extracellular polymeric substances (EPS). Water Res. 2016, 88, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Peterson, G.L. A simplification of the protein assay method of Lowry et al. which is more generally applicable. Anal. Biochem. 1977, 83, 346–356. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Yates, E.A.; Philipp, B.; Buckley, C.; Atkinson, S.; Chhabra, S.R.; Sockett, R.E.; Goldner, M.; Dessaux, Y.; Camara, M.; Smith, H.; et al. N-acylhomoserine lactones undergo lactonolysis in a pH-, temperature-, and acyl chain length-dependent manner during growth of Yersinia pseudotuberculosis and Pseudomonas aeruginosa. Infect. Immun. 2002, 70, 5635–5646. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.S.A.; De Simone, L.; Bruno, G.; Park, H.; Lee, K.; Fabbricino, M.; Angelidaki, I.; Choo, K.-H. Quorum quenching, biological characteristics, and microbial community dynamics as key factors for combating fouling of membrane bioreactors. npj Clean Water 2021, 4, 19. [Google Scholar] [CrossRef]
- Salehiziri, M.; Amalfitano, S.; Gallipoli, A.; Gianico, A.; Rad, H.A.; Braguglia, C.M.; Fazi, S. Investigating the influences of quorum quenching and nutrient conditions on activated sludge flocs at a short-time scale. Chemosphere 2020, 248, 125917. [Google Scholar] [CrossRef]
- Chong, T.-M.; Koh, C.-L.; Sam, C.-K.; Choo, Y.-M.; Yin, W.-F.; Chan, K.-G. Characterization of quorum sensing and quorum quenching soil bacteria isolated from Malaysian tropical montane forest. Sensors 2012, 12, 4846–4859. [Google Scholar] [CrossRef] [Green Version]
- Ryu, D.H.; Lee, S.W.; Mikolaityte, V.; Kim, Y.W.; Jeong, H.Y.; Lee, S.J.; Lee, C.H.; Lee, J.K. Identification of a Second Type of AHL-lactonase from Rhodococcus sp. BH4, belonging to the α/β Hydrolase Superfamily. J. Microbiol. Biotechnol. 2020, 30, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Kusada, H.; Zhang, Y.; Tamaki, H.; Kimura, N.; Kamagata, Y. Novel N-Acyl Homoserine Lactone-Degrading Bacteria Isolated From Penicillin-Contaminated Environments and Their Quorum-Quenching Activities. Front. Microbiol. 2019, 10, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.J.; Park, S.Y.; Lee, J.J.; Yum, D.Y.; Koo, B.T.; Lee, J.K. Genes encoding the N-acyl homoserine lactone-degrading enzyme are widespread in many subspecies of Bacillus thuringiensis. Appl. Environ. Microbiol. 2002, 68, 3919–3924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Screening and confirmation of potential AHL-degrading bacteria originating from activated sludge from a local municipal wastewater treatment plant. C8-HLS was used in all screening and confirmation experiments. (a) Screening of potential AHL-degrading bacteria from colonies on LB agar plates; (b) first confirmation of degradation ability; (c) second confirmation of degradation ability; (d) Rhodococcus sp. BH4 was used as a positive control, and autoclave-killed BH4 and water were used as negative controls.
Figure 1.
Screening and confirmation of potential AHL-degrading bacteria originating from activated sludge from a local municipal wastewater treatment plant. C8-HLS was used in all screening and confirmation experiments. (a) Screening of potential AHL-degrading bacteria from colonies on LB agar plates; (b) first confirmation of degradation ability; (c) second confirmation of degradation ability; (d) Rhodococcus sp. BH4 was used as a positive control, and autoclave-killed BH4 and water were used as negative controls.

Figure 2.
Final confirmation of degradation ability of four bacterial isolates (A9, A12, B11, and D3). To ensure their purity, each isolated bacterium was cultured in LB broth and then plated on LB agar before this test. Well-separated colonies formed on the plates, and 104-, 105-, and 106-fold diluted cell suspensions cultured in LB broth, were selected for the test, except for B11, for which only 105- and 106-fold dilutions were used. Each isolate was triplicated and the assay was duplicated. Rhodococcus sp. BH4 was used as a positive control. Autoclave-killed BH4 and water were used as negative controls.
Figure 2.
Final confirmation of degradation ability of four bacterial isolates (A9, A12, B11, and D3). To ensure their purity, each isolated bacterium was cultured in LB broth and then plated on LB agar before this test. Well-separated colonies formed on the plates, and 104-, 105-, and 106-fold diluted cell suspensions cultured in LB broth, were selected for the test, except for B11, for which only 105- and 106-fold dilutions were used. Each isolate was triplicated and the assay was duplicated. Rhodococcus sp. BH4 was used as a positive control. Autoclave-killed BH4 and water were used as negative controls.

Figure 3.
The degradation curves of eight different AHLs using the A9, A12, B11, and D3 bacterial isolates. H2O: A negative control that replaced the cell suspension with water. DC: A dead-cell control, in which A12 killed by microwaving served as another negative control. The error bars show the standard deviation of three analytical replicates. Designed initial AHL concentrations were 100 nM.
Figure 3.
The degradation curves of eight different AHLs using the A9, A12, B11, and D3 bacterial isolates. H2O: A negative control that replaced the cell suspension with water. DC: A dead-cell control, in which A12 killed by microwaving served as another negative control. The error bars show the standard deviation of three analytical replicates. Designed initial AHL concentrations were 100 nM.

Figure 4.
Characterization of AHL-degrading enzymes. Each assay was triplicated and the initial concentration of C8-HSL was 5 μM. (a) Evaluation of whether the AHLs were degraded by heat-sensitive enzymes or if there were other inhibitors that might interfere with the biological pathway in the bioassay. (b) Investigation of whether the AHL-degrading enzymes are extracellular or intracellular.
Figure 4.
Characterization of AHL-degrading enzymes. Each assay was triplicated and the initial concentration of C8-HSL was 5 μM. (a) Evaluation of whether the AHLs were degraded by heat-sensitive enzymes or if there were other inhibitors that might interfere with the biological pathway in the bioassay. (b) Investigation of whether the AHL-degrading enzymes are extracellular or intracellular.

Figure 5.
Transmembrane pressure (TMP) monitored in an A12 MBR and a control MBR. When the TMP reached 30 kPa, it was considered the end of the test and new membrane modules were installed for the next cycle. There were three cycles of tests.
Figure 5.
Transmembrane pressure (TMP) monitored in an A12 MBR and a control MBR. When the TMP reached 30 kPa, it was considered the end of the test and new membrane modules were installed for the next cycle. There were three cycles of tests.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Performance and characteristics of an MBR with A12 QQ bacteria and a control MBR.
| Parameter | A12-Inoculated MBR | Control MBR | Note |
|---|---|---|---|
| COD removal rate (%) | 96.83 | 95.56 | n = 1 |
| NH4+-N removal rate (%) | 99.69 ± 0.09 | 99.59 ± 0.13 | n = 3 |
| Effluent NO3−-N (mg/L) | 33.25, 24.96 | 34.98, 24.96 | n = 2 |
| Effluent NO2−-N (mg/L) | 0.11 ± 0.05 | 0.36 ± 0.03 | n = 3 |
| MLSS (mg/L) | 3661.1 ± 846.0 | 3846.7 ± 602.5 | n = 3 |
| EPS protein (mg/g MLSS) | 2.73 ± 0.46 | 3.0 ± 0.47 | n = 4 |
| EPS polysaccharide (mg/g MLSS) | 14.84 ± 4.36 | 11.93 ± 1.13 | n = 4 |
| SMP protein (mg/g MLSS) | 2.62 ± 0.68 | 2.35 ± 0.78 | n = 4 |
| SMP polysaccharide (mg/g MLSS) | 15.82 ± 2.13 | 15.73 ± 4.01 | n = 4 |
MLSS: mixed liquor suspended solids; EPS: extracellular polymeric substances; SMP: soluble microbial products. p-value of EPS protein, EPS polysaccharide, SMP protein, and SMP polysaccharide between A12 MBR and control MBR are 0.23, 0.14, 0.02, and 0.48, respectively.
Table 2.
16S rDNA sequences of bacterial isolates and BLAST results.
| Isolate | Source | Most Closely Related Bacterium 1 | Sequence Identity (%) | G(+)/G(−) |
|---|---|---|---|---|
| A9 | Activated sludge | Brucella anthropic | 1363/1365 (99.85) | G(−) |
| A12 | Activated sludge | Bacillus cereus | 1397/1398 (99.93) | G(+) |
| B11 | Activated sludge | Bacillus toyonensis | 1327/1330 (99.77) | G(+) |
| D3 | Campus drainage | – | – | – |
1 Only the name of the first microorganism is shown if more than one sequence shared the same percentage identity.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chien, I.-C.; Chu, C.-Y.; Huang, C.-Y. Isolation of Bacteria Capable of Degrading Various AHLs for Biofouling Control in Membrane Bioreactors. Water 2022, 14, 1712. https://doi.org/10.3390/w14111712
AMA Style
Chien I-C, Chu C-Y, Huang C-Y. Isolation of Bacteria Capable of Degrading Various AHLs for Biofouling Control in Membrane Bioreactors. Water. 2022; 14(11):1712. https://doi.org/10.3390/w14111712
Chicago/Turabian StyleChien, I-Chieh, Chiao-Yun Chu, and Chien-Yu Huang. 2022. "Isolation of Bacteria Capable of Degrading Various AHLs for Biofouling Control in Membrane Bioreactors" Water 14, no. 11: 1712. https://doi.org/10.3390/w14111712
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.